THERE CANNOT BE a more important event than the greening of the world, for it prepared the way for everything that happened on land thereafter in the evolutionary theatre. A love of green is not just a sentimental attachment to rural holidays remembered from youthful days, green days. It runs deeper than that. In desert countries the rich sheikh celebrates his fortune with a garden sequestered away from the sun. We admire grandeur in wild scenery, mountains, canyons, deserts and glaciers. In such territory eventually this majesty begins to pall; a vague sense of dissatisfaction creeps in. Something is missing. But in greenness there is repose. It has been proved that the green wavelengths are least irritating to the retina. Red is angry, blue is cold, but green is restful. In The Golden Bough Sir James Frazer catalogued endless human ritual variations on the theme of welcoming the return of spring, of rain, of foliage and growth. There is the medieval Green Man, an atavistic figure to be found carved in stone or wood (rarely, coloured) in European churches and cathedrals. On his wild face tendrils grow from nose and brow. As they branch out they bifurcate, and give off simple leaves. They might be a representation of an early plant, like Cooksonia or Rhynia. All terrestrial animal life grew up alongside green plants, and we are still bound to them. Green still represents fertility and fecundity, and even amid the sophistication of city life, there is still some urge that drives people to parks and gardens. In China I joined a queue of hundreds of quiet Chinese in order to admire the plants in the great garden of Hangzhou. This was in the time before self-expression was encouraged, and the comrades lined up patiently in their blue serge suits. In a recess of the unconscious mind there may still be an awareness of the greening of the landscape in Silurian and Devonian times, that first Eden. Andrew Marvell’s wonderful lines are worth recalling:
The mind, that Ocean where each kind
Does straight its own resemblance find;
Yet it creates, transcending these,
Far other worlds, and other seas;
Annihilating all that’s made
To a green thought in a green shade.
During the Cambrian one-third of the world was devoid of life. The barren area was the land surface away from the sea. There may well have been bright stains of bacteria around springs, and covering such rocks as were washed regularly by showers. But the landscape would have been devoid of any softening tones of green. It would have seemed, to our eyes, naked and harsh. Nothing would have been there to consolidate loose soils, to absorb the worst of the weather, so that every rainstorm would have prompted a small flood, and stones and pebbles cascaded down slopes and tumbled freely into the choked beds of rivers. This was a world of erosion, one that left the elements free to wreak their worst. We might catch a glimpse of such a world in the remoter parts of the Arabian Peninsula today, where no trees break the hard horizon. Sandstorms whistle across the plains, or slowly build dunes. The rock is sculpted by wind and sand, which finds out every weakness. When there is a storm the flood carries great quantities of rock away; the waters gather finally into basins, where they evaporate into glistening salt pans. There is no sound save the wind, nor any sense that it was not always thus from the beginning of time. In mountainous regions in the Cambrian we must imagine a high and unclothed Himalaya, where frost prised slabs of rock away, and rain carried off its stony booty almost as fast as tectonics could uplift the ranges. Great fans of scattered rocks would have carpeted the plains beyond. There might have been a certain stark beauty to the landscape, but our eyes would soon have sought the comfort of an oasis, or a green valley.
The greening of the land was an extraordinary transformation to the beauty of the world, not merely an opportunity for a great expansion in the compass of life. I like to imagine the breakthrough happening as a dramatic change of scene, so that plains and mountain slopes alike were verdured in a geological instant, and all the bleakness of the ancient landscape was suddenly painted over with a thin carpet of photosynthetic green. In fact it was probably not like this, neither so sudden nor so complete. The colonization of the land by plants entailed surmounting several important and tricky physiological obstacles. If one plant species made such a breakthrough away from water it must have had a hundred precursors which had solved these difficulties before it. It is unquestionable that algal ancestors lay below the first land plants. It was once thought that the first truly terrestrial plants were Devonian in age, but then examples were found from Silurian rocks. Now, students of this early history are looking for the kind of spores which might have been dispersed not through water but through the atmosphere, a sure indicator of terrestrial habits, at least at some stage in development. These spores are more easily preserved than the delicate fronds of whole plants. They can be extracted from sedimentary rocks by dissolving them in hydrofluoric acid. Spores* with the right kind of design and ornament have now been found in rocks of Ordovician age. This confirms, sadly, that my dramatic vision was incorrect, and that the greening happened over tens of millions of years.
There was not merely the business of moving from an environment supported by water to one of thin air. Tissues are delicate, and desiccation is the worst kind of destruction that can happen to them. So it was necessary for plants to develop a thin, waxy coat on the outside of their green fronds, which could cut out, or at least much reduce, water loss. Surely the first plants would have hovered tentatively at the water’s edge, ever vulnerable to just too much roasting by the sun, crisping when the conditions deteriorated, plumping up with rain or dew. Breathing through such a waxy covering then presented a different set of problems—after all, what retains moisture also excludes air. The compromise is a special arrangement of cells in the surface of the plant to make stomata, minute holes guarded by sausage-like cells which can admit air but which expand to close the apertures when conditions get too drying—a natural air-conditioning system at a microscopic level. Once armed with waxy coat and stomata, a flat shoot could creep over mud, and put up shoots into the air, extending short roots to absorb nutrients even as the green surface photosynthesized and grew, expanding into a new world of air and light.
If you kneel down by a small stream bank where the atmosphere is moist and dank, listening to the tumbling water, brush aside overhanging ferns and on the bare bank above the water line you will very likely see such a prostrate green leaf. Dark green, and perhaps slightly crimped, this plant will make little effort to lift itself off the muddy bank. It forms a verdant crust. It seems to revel in the dim light, soaked, and skulking where nothing else grows. This is a liverwort. It may be as close as we can get in the living flora to those ancient plants that first made the break from water, though there is little enough in a liverwort that can be preserved as a fossil. It is a plant so primitive that it does not even have stomata. Now, liverworts are confined to the edges of water and to dim caves, once more occupying only the niches in which they may have started. Photosynthetic pads like these have a two-dimensional spread over mud and soil. Exposed thus to the sun’s heat and radiation, like a prostrate sun-bather on the beach, they must resist the effects of harmful ultraviolet rays. It will be recalled that these rays are partly filtered by the ozone layer, which is formed high in the atmosphere by transformation of oxygen molecules. Presumably, there must have been enough oxygen in the atmosphere of the Ordovician to start such a protective process, the long, patient result of 3,000 million years of algae and “blue-green” bacteria releasing oxygen into the atmosphere. This reminds us of a link with the earliest days of the Earth: in a sense, leaves were conceived in the first, tacky microbial mats.
The task of almost any other plant you care to name is to grow upwards, towards the light, aspiring to the sky. This at once creates a whole environment: protected, sheltered places beneath stems and foliage. But prostrate mats require some stiffening before they can rise above the mud, and they need points of growth where shoots can be concentrated so that they do not grow haphazardly. As shoots get longer they need a system of transporting water from the roots to the shoots, to keep the whole structure turgid; contrariwise, the products of photosynthesis need to be distributed to the rest of the plant from the leaves. The plant, in short, is little more than a photosynthetic factory with problems of distribution and supply like any manufacturer. I wonder if it is coincidence that complex and large factories are referred to as “plants.” Lest this seems a drearily reductionist view of the wonders of plants, consider what happens when any part of the system fails: that wilting pot plant deprived of water; the etiolated and spindly specimen deprived of light; the floppy failure forced to grow too fast. Plants are what late-twentieth-century systems analysts would call integrated feedback systems, and if the feedback fails they let you know by wilting, and eventually by dying. The technical problems that need to be overcome to make a viable, upright plant are problems of plumbing and engineering, chemistry and aerodynamics.
In the first place, there is support. Some plant organs achieved a certain measure of rigidity by their cells clubbing together, leaning on one another for mutual support. When I was a child there were still a few farms on which the wheat was harvested using equipment less sophisticated than the combine harvester. The cut sheaves were stacked into standing bunches which dotted the fields. They were known as “stooks.” It is odd to reflect that this is a truly obsolete word, for other than in the background of a painting by Constable or Brueghel I cannot imagine where a stook will be seen in the future. But the principle is not so different from that behind the construction of the stem of a plant. In cross section most stems are composed of stacked, long tubes, which associate together for strength. The tubes conduct fluids—sap, the “blood” of plants. The cell walls acquired a thickening, often making a characteristic spiral pattern. The tough, organic building material lignin further reinforced the structure. Plumbing and engineering combined to allow upward growth. This full complement of features is typical of vascular plants that clothe the world today.
Shoots may have come before leaves. Some of the earliest plants in the Silurian were small and creeping, sending up upright shoots that bifurcated, once or twice, like candelabras. None was identical with any plant still alive, although there was a passing resemblance to some of the simplest mosses. Leaves were often nothing more than small scales that clothed the shoots. It is known that the earliest fossils of land plants are associated with sediments that accumulated in rivers and estuaries. It was around some ancient lake or river that shoots first poked their heads above water. But why did they leave the sheltering water? What prompts life to nudge its way across such thresholds?
The question can be asked as appropriately here as at any other point in this narrative. Some writers, especially geneticists, invoke the imperative drive for reproductive success, the urgent bidding of DNA to propagate itself until it subdues the very stars. But then the time was also undoubtedly ripe. If the planet had an evolutionary history which was both part of, and engendered by, the organisms that shaped it, then this was the moment when new things almost had to happen. The atmosphere was ready, the climate was ready. But chance alone could equally be cited as the prime mover. Ponds and lakes are notoriously unreliable habitats, drying out at some times, overflowing at others. The crucial engineering changes possibly happened as a way of coping with unpredictability. How much better to weather a dry period than to die out completely, and what better incentive to favour plants which could reproduce by means of spores, dispersed through the air? The earliest stages of the story are indeed recorded only by the spores that spread beyond the habitats of the first land plants and were left behind in shallow seas and deltas. The microscopic spores themselves had to be specially constructed to survive out of water: they have a tough coat, the exine. This enveloping cover is also impalpably thin, but still serves to protect delicate reproductive contents. Its toughness is confirmed by the survival of these tiny objects for hundreds of millions of years. Maybe one day a fortunate hammer blow will reveal the Archaeopteryx or Australopithecus of plants.
Early plants are preserved as black films on shales; often they look little more than dark scribbles on the rock surface. Their fine detail has to be teased out by means of preparations which clear their ancient tissues. Diane Edwards, professor at the University of Wales, waxes ecstatic over a dark smear conjured from the early Devonian rocks that crop out along the mouth of the river Towy in South Wales, because she can guess what details they may yet reveal. She has contributed more than anyone else to our knowledge of the vital steps taken by humble plants in ancient times.
Plant fossil hunters particularly like to find sporangia, the vessels that held the spores. These little sacs can be picked apart and the spores they contain extracted. In this way we can attempt to link the common fossils known only from spores with actual species of extinct, primitive plants. Sadly, many fossil shoots are sterile. But the cases that are reliably connected show that there were progressive adaptations to wind dispersal through early plant history, as proved by smaller spores. Occasionally, petrifactions are discovered which allow for a complete reconstruction of plants, down to the last cell. In the most famous of these, the Devonian Rhynie Chert, a hard, greyish rock discovered in the early years of the twentieth century in Scotland near the village that gives it its name, each cell has been filled immediately after death by silica, so that when the chert is sliced thinly, the minute structure of the plants can be examined. In some of these plants the vascular tissue is well developed, and it is not so difficult to imagine them being related to ferns. Ferns are presumed to have a history almost as long as that of liverworts. Living ferns, too, belong to the realm of green shade, although some species are tough and can endure drought. I was astonished to find them growing on arid slopes in the middle of Australia, in the company of Spinifex and Eucalyptus. On the underside of the fronds the little spore sacs appear as brown dots. If they are dusted on to moist peat the spores will germinate; but the little plant that arises is not the spore producer. Instead, specialized portions of this reproductive generation produce male sperm; eggs are produced in another. Only when the one has been fertilized by the other can the mature fern grow. This fertilization happens as sperm swim through the medium of water, be it ever such a thin film. Hence the fern still has a link to its algal forebears, and an obligation to the water from which it crept, and, appropriately, ferns are still at their most extravagant in places like the forests of New Zealand, where there is abundant and continuous rainfall. There, tree ferns erupt skywards like fireworks; epiphytic ferns hang from branches, as delicate as lace; creeping ferns cover every patch of untenanted soil. There is nothing obsolete about ferns, even though their fossil record stretches back to the days of the first tenancy of the land.
There are some places which are holy sites for palaeontologists. The town of Ludlow in the Welsh Borderland is one of these shrines: it was in the vicinity of Ludlow that many of the important stories concerning the conquest of dry land by plants and animals were constructed from clues found in the Late Silurian rocks. The town retains a character that neither one-way systems nor superstores can obliterate. Timber-framed houses still lean out over the streets, surprising in their top-heaviness, and showing off their oak timbers with an ostentation that signifies centuries of comfortable and peaceful affluence. The Feathers is an old and fine hotel decked out in this black-and-white livery. Geologists stay there, and have done so since the early days. You can mention Ludlow to Czech or Estonian geologists and they will grin with recognition, pat you on the shoulder and say: “Ah … the Feathers! Very good.” There are rooms with eccentric angles, and watercolours that are difficult to hang straight. There is good ale, without which no scientist can draw any conclusions of any worth or lasting value. People still assume that you are honest there, and do not demand twelve kinds of identification before they will cash a cheque.
There is a tract of land running from Church Stretton to Ludlow in the east, to Bishop’s Castle in the west, along the old border with Wales. It is not grand country, if by grandeur you mean spectacle. But I find that the Long Mynd, an ancient and wild inlier of Precambrian rocks that stands high above the surrounding low hills, is all that is needed in the way of spectacle. I am always trying to find excuses to return to Bishop’s Castle, where Miss Sheckleton used to provide the perfect bed and breakfast in a Tudor house with flag floors and improbable staircases. Her breakfast was neither too much nor too little, the egg yolks were soft and the whites were cooked, the toast warm and cut into triangles on which to spread thick, home-made marmalade, and pots of tea arrived unbidden. At such a breakfast time nobody could be more content than a geological researcher looking forward to the day’s discoveries. In Shropshire, wandering up streams, hammer in hand, you could be forgiven for thinking that little had changed since the Devonian period, for there are still ferns and liverworts galore: male ferns stacked against the banks, polypodies sprouting from rock surfaces like so many feathers, and all along the stream sides there is still a crust of richly green liverworts.
At the end of the Silurian period, and in the following Devonian, this part of the Welsh Borderland lay to the south of a growing Caledonian mountain range which stretched away to where the Highlands of Scotland now spread the northern part of Britain with wild, boggy moorland and glens. The peaks that remain there now are but ground-down relics of what once extended as far as northern Norway. Towering like the Alps, snow-clad peaks fed streams which carved away the mountains even as they grew, carrying material away to be deposited on plains beyond and, ultimately, to the sea. Gradually, the sea retreated southwards as the apron of debris, mostly sand and silt, accumulated from the slow demise of the Caledonian Mountains. In many areas the iron that weathered away under the ancient tropical Sun colours the rocks a rich, deep red, or even purple. These rocks have long been known as the Old Red Sandstone. In South Wales it was these rocks that yielded to Diane Edwards some of her favourite fossil plants. Where cows and sheep now graze in the peaceful fields of Shropshire there was then a maze of estuaries, streams and lakes, over which the sea stepped from time to time, although with less and less frequency. The deposits of the riverbank, or the floors of estuaries, have yielded many of the remains that record the very transition from sea to land, by way of intermediate habitats. What is now a domestic landscape was witness to the most profound events in the history of life. It somehow seems too tame, this comfortable countryside: one almost demands a spectacular geological feature to match the profundity of the events recorded in the rocks. For here there are cadavers of early fish, some still so incompletely known as to be mysteries. There were fish the size of herrings without hinged jaws, just a simple opening for a mouth, and many, like Cephalaspis, were more or less protected by hard, bony plates on the outside, and were almost certainly feeding on algae and other organic materials grubbed from the mud. During Silurian times fish must have made a transition from marine habitats to being capable of coping with both fresh and brackish water. Once they had overcome the physiological problems of making this transition, of swapping salt water for fresh, one can visualize these sluggish animals grubbing their way upstream, following the nutritious mud, tapping willingly into a resource that hitherto had been exploited by nothing more advanced than bacteria. For a brief heyday, it was a good time to be a simple fish.
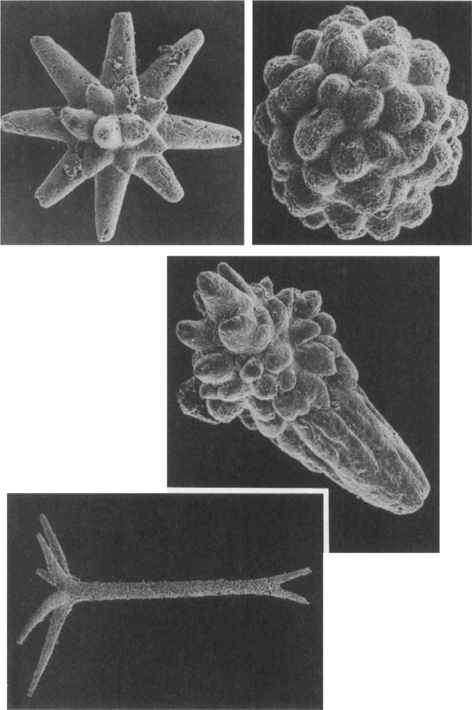
20. Skeletal fossils of animals from the early Cambrian of South Australia (photos courtesy Stefan Bengtson); enlarged between 50 and 150 times. Small shelly fossils show a great variety of shapes, but can be problems to interpret in terms of living relatives.
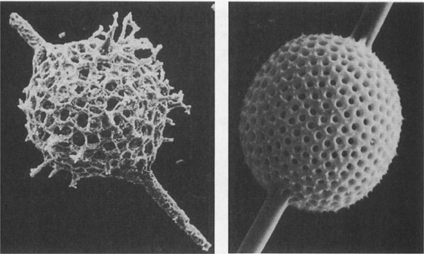
21. Radiolarians—a protistan group with a long history and siliceous skeletons. The example on the left, a millimetre across, is from Ordovician rocks in Spitsbergen; on the right is a living radiolarian from Barbados.
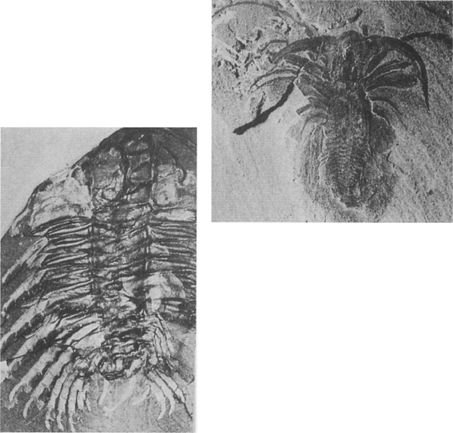
22. Burgess Shale (Cambrian) arthropods preserving amazing details of their limbs: Olenoides (trilobite), left, and Marrella, above

23. A trilobite (Phacops from the Devonian) showing its sophisticated eye—note the individual lenses exquisitely preserved for hundreds of millions of years.

24. Reefs ancient and modern: the structure of the Silurian reef in Shropshire is very like that of the modern reef (left) even though the organisms of which it is composed are only distantly related.

25. Conodonts—an assemblage of tooth-like conodonts, each a millimetre or so long. For many years their precise nature remained an enigma. Now they are thought to be allied to vertebrates.
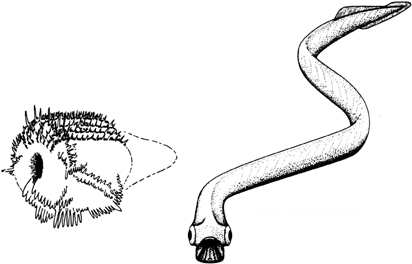
26. Professor Lindström’s reconstruction (far left) and the real conodont animal as recently reconstructed by Mark Purnell (left).

27. Graptolites “writing in the rocks.” The saw-like edges were the tubes inhabited by small living animals. Ordovician Didymograptus on black shale (natural size)

28. Charles Lapworth, namer of the Ordovician System

29. Tree ferns still form an exuberant canopy in New Zealand.

30. The Feathers in Ludlow, a well-known rendezvous for Silurian palaeontologists

31. A thin section through Devonian Rhynie Chert reveals the minute details of each plant cell—a snapshot of the early colonization of land. This is a section through Rhynia, magnified forty times.

32. Willi Hennig, the originator of cladistics

33. An early and distant relative of the spiders, a trigonotarbid, painstakingly reconstructed from sections by Jason Dunlop of Manchester University

34. Part of the skeleton of “Boris” from the late Devonian of Greenland, the many-fingered early land vertebrate. Its scientific name is Acanthostega gunnari.

35. Jawless fish, Spizbergaspis, from the Devonian of Spitsbergen. The head of this animal clearly shows the paired eyes behind the forward nose.

36. A spiny trilobite, Ceratarges, from the marine Devonian rocks

37. The wonderfully regular bark of the Carboniferous tree Lepidodendron

38. FAR LEFT: One of the ornamental columns in the Natural History Museum, London, based on exactly the same species of Lepidodendron—a case of craft imitating nature

39. The living Selaginella— a humble survivor from the days of the Carboniferous giant trees

40. Carboniferous cockroach Aphthoroblattina johnsoni. This specimen comes from Coseley, Staffordshire.

41. The “Bolsover dragonfly” Tupus diluculum—a giant of its kind—flying in a Carboniferous swamp
It is interesting to reflect on how historical accidents played their part in the story of life. Were it not for the convulsions of the Earth which threw up Caledonia and the Appalachian chain* there would not have been the myriad lakes and streams in which nurseries for new species could be established. Species are produced by isolation, and in the Devonian mountain chains these unique historical circumstances allowed for little sequestered pockets which in turn permitted animals to go their own way. To take an appropriate linguistic analogy, there are more extreme dialects in Norway among a few million people than there are in the whole of the vast United States. These dialects developed along the thin, but impassable Caledonian chain that follows the present Atlantic coast of Norway. Glacial erosion has sculpted this coast into a series of spectacular and deep fiords. But contact between settlements on the shores of adjacent fiords is limited, other than by sea. Before modern communications had become established, local Norwegian populations developed their own ways of speaking, almost their own languages, in isolation from their neighbours.
It is not difficult to imagine that, tucked into equally isolated crannies of the ancient Caledonian range, there were places where adventurous, innovative species could thrive away from too much direct competition. Land life was born and bred at the edge of dying mountain chains. These vanished Alps and Andes themselves were thrown up by what Charles Lapworth called the “great Earth engine” of tectonics. The great breakthrough to land was stimulated by the restless crust, upon which life was carried as a flea may be carried by a bison. It was the disappearance through subduction of oceans which had been wide in the Ordovician that brought continent into collision with continent and threw up mountains that stretched through the Appalachians of the southern United States, through Vermont to New Brunswick and Newfoundland, and ultimately through highland Scotland to Norway. Genesis was as much about geography as genetics.
EVEN AS THE PLANTS WERE CREEPING over the land, animals were leaving the water, too. Their early history is, if anything, even more obscure than that of the plants. It will come as no surprise to learn that the arthropods were among the vanguard. Some have even claimed that terrestrial tracks showing the whiskery or pleated series of marks typical of arthropods indicate that they may have accompanied—even predated—the landward march of plants. Such tracks are certainly known from the Silurian rocks of Western Australia. But it is probably special pleading to claim such precocity for the arthropods, because it will be recalled that evidence of land plants in the form of their cryptospores has been found in rocks as old as Ordovician. But it is doubtless true that there are fossil footprints of arthropods in localities where the arthropods themselves are reluctant to put in an appearance. The animals that made these footprints may have been too delicate, easily washed away, their skeletons dispersed and broken. But their tracks were evidently rather easily preserved on the sandy flats that surrounded temporary lakes, or on the levees that flanked the rivers. Thus may the most casual of actions in the life of an animal be destined for immortality. The transient may be linked to the eternal:
To see a World in a grain of sand,
And a Heaven in a wild flower,
Hold Infinity in the palm of your hand,
And eternity in an hour
as the poem by William Blake described it.
Arthropods enjoyed a brief residence at the apex of the pyramid of life during the period of transition from water to land. Water scorpions—eurypterids—haunted the estuaries, lakes and rivers of the Silurian and Devonian. They were the giants of their jointed-legged kind. The longest species of the genus Slimonia was the size of a man. At its front were pincers; the flattened body was carried on spindly legs; there were eyes on the top surface of the head; at the back end there was a leaf-like “paddle.” Other species had a spike at their posterior end which probably functioned as a sting. The appellation of these animals as scorpions is no coincidence—they were indeed relatives of those most poisonous predators in hot climates. In Mexico scorpions are still one of the most frequent causes of death, far outstripping snakes; the notion of a two-metre subaqueous scorpion is one that chills the blood. The largest specimen of Slimonia in the Natural History Museum in London is in a six-foot-long slab of sandstone, a mighty animal by any standards. It once fell on the foot of my assistant, turning his big toe blue for weeks, and proving that it could still do damage nearly 400 million years after its demise. These animals must have terrified jawless fishes, lurking in the shallows until some unfortunate creature crawled into the compass of their arms. The lungs lay on the underside of the body, oxygen-absorbing tissue tucked away, and folded and crimped like a napkin in a posh restaurant. Slimonia needed to husband its oxygen. I imagine it lying motionless for hours or even days at a time, much as some species of crocodile do today, until a misguided creature approached close enough to be grasped. Then … a sudden lunge, fast as the snap of a mousetrap.
There are limits to arthropodan horror, but they are imposed by the laws of physics rather than the constraints of imagination. The arthropod skeleton is a wonderful construction—but it works best at small size. The arthropods have an exoskeleton, a series of tubes that house the flesh and muscles inside. That is why we have to pick at lobster legs with delicate forks to get out the last of the meat. But as legs built on this plan get bigger—and this applies especially to support in air and on land—so the efficiency of such tubes as support dwindles; they need to have thicker and thicker walls, and then the operation of any kind of joint becomes progressively difficult. So does breathing. As in plants, oxygen has to be absorbed into the animal through an impervious skin. “Lungs” may be no more than small tubes—trachea—connecting with the surface of the body, through which air may pass and be absorbed by way of soft membranes into the body cavity. At very small sizes, its subsequent diffusion is sufficient. But as arthropods get larger the surface area of the animal increases in proportion to the square of its dimensions, but its volume requiring to be serviced by breathed oxygen increases as the cube. The arithmetic is obvious. A doubling in size requires an eightfold increase in oxygen absorption; there has to be an upper size limit beyond which breathing becomes almost impossible. So it seems likely that our eurypterid was torpid most of the time, lying in wait in some muddy ditch or stream, biding its time, harbouring its strength for a swift kill. (It will be recalled that the living parasitic arthropods known as bed bugs are capable of living without food for long periods of time until a meal presents itself, when they spring, or rather crawl into action with surprising alacrity.) Venom also helps to reduce the struggling-time of prey, a technique that many snakes still employ to advantage. The sea scorpions were soon outclassed as predators by jawed fish, but I like to think that the frisson of terror we still feel when we disturb a scorpion is akin to that felt by a Devonian fish disturbed from sucking up Devonian mud by the claws that lurked beneath it.
The Rhynie Chert preserved more than just early plants. The extraordinary seal from decay provided by siliceous water could entrap the most inconspicuous of animals. Plants and animals alike were petrified—turned to stone—by a hot spring, one that dribbled siliceous fluids around stems before they could decay. The silica replicated each cell with absolute fidelity. The plants probably grew around warm pools, fed by nutrients that accompanied the waters welling up from deep within the Earth. Beneath them there would have been a humid microclimate, and plant debris would soon have rotted to form the first humus-rich soil. This was a habitat, a haven from the hardships of a young world; a green arbour of opportunities to earn a living. In the Rhynie Chert there are spider-like creatures called trigonotarbids that were certainly equipped with massive jaws, and must surely have been predators. They did not spin webs, but probably simply crawled over their food and chomped it. But they prove that already by the Devonian there must have been a food chain. Then there are mites. These are both the most numerous and least conspicuous of arthropods, being at most a millimetre or two long. Today they teem in the soil, they clean up small particles, they feed on human skin and impinge on our awareness only because the droppings of house mites cause asthma. They are a food source for thousands of other creatures of the loamy dark. They share a distant common ancestor with the spiders. There is another tiny animal, a springtail, which has been claimed as the most primitive of insects. It has no wings, but like all insects it has only three pairs of legs. Living springtails can easily be observed as tiny, dark “bugs” around the edges of ponds; disturb the dank vegetation there and you will see little insects “hopping.” The “spring” in the tail allows for this powerful escape reaction. The Rhynie Chert specimen comes complete with its “spring.” They have almost no other fossil record, but their ubiquity today probably indicates that they have been pursuing their inconspicuous life habits since very early times. In another Devonian age deposit, from Gilboa, in the eastern USA, there are other small animals, including a strange creature called a pseudoscorpion. These miniature predators are still common in almost all damp places today, where they cull other tiny animals that live in soil. Their arms seem hardly to have changed since the Devonian.
What is extraordinary about these early communities of small arthropods is not how different they are from their equivalents in the living world, but how remarkably similar. It can be concluded that once animals and plants left the water they set up a mutual dependency which has endured ever since. This is the decomposing community of animals that live in the soil, breaking down organic matter, recycling it, returning it to the soil once again. In its own way it is an unspectacular little ecosystem, conservative, even dull. But it is vital to the health of the planet. If the springtails were to undergo a mysterious demise, together with the mites that live in soil and the minute fungi upon which they feed, quite soon there would be an ecological crisis of a magnitude we can scarcely imagine. Nutrients would become locked away, the soil would become progressively impoverished, larger plants would die, and soon animals would follow suit. Novels that seek to portray post-holocaust worlds always seem to assume that the soil will magically survive, and that a bean cast into seared soil will quietly proceed to a successful crop. But the soil is not a passive medium; it is alive. I doubt whether there would be many readers for a post-holocaust novel that was concerned with the hero’s desperate search for a mite. But alas for the world if the mites and their diminutive allies failed to prosper!
There are other survivors. After all, liverworts, mosses, clubmosses and ferns are still found in many places around the world, surviving messengers from the days of the great greening. The meek may, after all, inherit the Earth. But they remain meek—through hundreds of millions of years. It is as if lack of ambition somehow secured longevity. Live and let liverwort! These organisms are reminiscent of the Good Soldier Schweik, the soldier who survived by dint of always being somewhere away from the front line. Stolidly avoid conflict, sit on the sidelines, look for the uncompetitive option. Become a liverwort. If persistence has to be purchased at the price of lack of adventure, so be it.
So the backboned animals followed shortly after the plants made their excursions on to land. By the Devonian, there were fish equipped both with backbones and with jaws alongside their jawless relatives, swimming through freshwater lakes, probing and exploring. These jawed fish included one species that was the ancestor of all land animals, be it lizard, bat, bird or dinosaur. Genetic similarities between all land-living animals equipped with backbones tell us that it was indeed likely that the step was made but once. This is not to say that other animals may not have lingered about the edge of the two worlds, trying their luck. In mangrove swamps in the tropics curious little fish called mud skippers can waddle out of water on their fins, but somehow they do not seem to be contenders to usurp the place of lions. It is difficult now to imagine the circumstances of that swampy exodus that changed the face of the Earth in so many respects. The hand that types this page was born in a limping, stumbling appendage that triumphed only by virtue of being first. I am reminded of the temptation, that any biographer has to resist, of making the tale seem too pat, the steps—in this case the most literal steps—too logically arranged. The first vertebrate limb was probably a poor thing; it was natural selection that refined it. We can imagine a whole range of flippers and fins that might be transformed into legs and hands by shifting this bone or that. All land vertebrates are termed tetrapods—literally “four feet”—and it is obvious that most amphibians, reptiles and mammals do indeed have four limbs. In creatures where they are lost, as in snakes, this loss is an evolutionary change from ancestors that originally also had four limbs. Where a pair of limbs is transformed, as wings are in birds, it is equally apparent that the wing is the equivalent of a limb, but changed for a different function—wings were not sprouted anew. Glance at a struggling, naked chick in a nest, and the bones—the arms—that go to make the wing are only too clear. A biologist would say that the wing and the limb are homologous structures—that is, they share a deep evolutionary identity. Limbs may be lost, but they cannot be reinvented, because the blueprint is fixed. Further, cats and humans alike have five toes. So do nearly all the tetrapods: where the number has changed—as it has, for example, in the horse, which has but a single toe—this is because of a proven reduction and loss of toes; the primitive number was still five. But why five? After all, it is well-known that there are genetic “sports” among humans which may result in whole villages with six digits. It does not seem to be a lethal mutation. It poses no major difficulties other than making trickier the early learning of the decimal system. The number five may seem to be the “right” number for digits in some indefinable way, but this may be no more than wayward chance, the fickle selection of one individual rather than another on a Devonian shore. I remember the circumstances which made me a palaeontologist and how they depended on one, crucial examination result; there could so easily have been a different slip of paper, another outcome.
It is now known that some of the early tetrapods had seven toes. The fossils of these ichthyostegids—an awkward name perhaps not altogether inappropriate for animals which probably waddled rather uncomfortably across the Devonian landscape—were recovered from sandstones and siltstones in east Greenland, rocks which were not so different from those in South Wales or the Welsh Borderland, being another part of the great Caledonian chain. The climate in Greenland was warm then. Ichthyostegids have been claimed to be little more than “walking fish,” and it is true that they retained from their aqueous ancestors both a fish-like tail and long jaws with rather undifferentiated teeth. But their limbs had become at least workable legs, even if they operated akimbo and spreadeagled. Their toes were the surprise. Early workers may have assumed that there had to be five toes because we tend to see what we expect to see, and, after all, everybody knows that tetrapods have five toes. Jenny Clack and her colleagues at the University of Cambridge reexamined some of the early collections of these animals, as well as making elegant new finds of their own. What they discovered was a surprising variation in digit numbers. In this lumbering world seven was as good as five. Per Ahlberg tells me that he is studying a species with eight toes. At some stage one of these fish-like land-dwellers with five toes pulled ahead of the rest, and this was the founding father (or mother) of five-ness in all subsequent terrestrial vertebrates. If one of its other waddling contemporaries had been the successful stock for all evolutionary lineages that followed we might now be struggling with seven-fingered gloves, or be able to play an octave with one hand without crossing our fingers. Or else villages with six digits might be the norm, and whispered remarks would be made about those freakish places in which the inhabitants were born with only five.
Lungs were necessary, too. Terrestrial breathing presents some of the same problems that stomata solved in plants—it is a question of taking the process of oxygen absorption inside the organism. External gills would soon dry out, even on a mud flat. Small insects and millipedes solved the problem by incorporating small tubes called tracheae into their bodies, but as we have seen this could not work at large sizes. Among the vertebrates, lungfish had already developed in early times the necessary air-breathing organ which gives them their name. Oxygen is absorbed through the moist, sequestered walls of lungs and tracheae alike. Some lungfish live in Africa, and when the rivers dry out, as they do in the dry season, these types can wrap themselves in cool mud and breathe air until the weather improves again. Was some such ruse the prompt that helped natural selection to improve the first lung? After all, we know that the Caledonian climate was hot, and the rocks are full of evidence of drying-out pools, leaving behind the tessellated testimony of mud cracks. In living animals the lung is crimped into folds to help oxygen absorption, and it is quite easy to see how, when this extraordinary organ had been developed, improvements in its design would have been at a premium. Once equipped with a scaly skin that would not dry out, with four waddling legs (regardless of toe number), and with lungs, the tetrapod vertebrate that was our ancestor—and ichthyostegids are a good approximation to it—could lumber away from the water. It was probably no herbivore, in which case it must have eaten other animals washed up on the shore, or caught the arthropods that had made the same journey away from water. It would, presumably, have returned to the water to breed, because a larval stage under water would have been retained from its piscine past. Not every change has to be made at the same time.
In the Mall, not far from St. James’s Park, there is an imposing square building at the end of a Regency terrace, which houses the Athenaeum Club, favoured by bishops and professors and senior civil servants. But there is another club that meets there from time to time in the dining room in the basement. This club is called the Tetrapods. It was founded by Julian Huxley and his friends in the 1930s. Biological matters of the day are addressed by a visiting speaker, and the chairman of the night bangs the table for order with the pubic bone of a whale. The fortunes of the dining club, like those of many other tetrapods, have gone up and down. At its low ebb, it met in a pub called the Goat. Some of the members have been coming for a very long time—in fact, nobody remembers who some of the oldest members are. But from such places ideas do diffuse through the scientific community, and I remember hearing about the important discoveries of the multiple digits on one of those very evenings. The chairman afterwards made some poor jokes (also a tradition) about some of the members returning to primitive tetrapodous modes of gait by the end of the evening.
THE LANDWARD TRANSFORMATIONS WERE probably already underway in the Silurian, but were completed in the Devonian period—that is, between about 410 and 360 million years ago. I should briefly describe the life and times of this crucial time period. The Devonian was named after the county of Devon, in southwest England. The countryside is singularly lovely, and much of it owes its character to the Devonian rocks. They underlie most of Exmoor, where even the constant inroads made by tourists cannot dull the charm of the Barle Valley, or overwhelm the friendliness of Dulverton. Tougher sandstones make for moorland, where sheep and heather flourish. Modern farming methods still seek to nibble away at the untamed stretches, but enough remains to help us to understand how wild it must once have been. Much of the charm resides in the contrast between the open moors and the sequestered valleys with their ancient cottages, many of them thatched. Westwards again, marine Devonian shales make up many of the sculpted cliffs of Cornwall. In south Devon, around Torquay, there is a completely different set of Devonian rocks, which contain marine fossils; little of the story of the colonization of land is preserved in this area. It presents instead a continuation of the riches of the Silurian seas, and there is the testament of corals, clams, brachiopods and trilobites to prove it. The Devonian rocks in parts of Ohio, New York State and Ontario are similar in character, but even more prolific. There is a famous trilobite called Phacops rana which teems in certain localities, and which has great eyes with spherical lenses and a knobbly head. There are dozens more such animals in great areas of Brazil, or Morocco, or Turkey. Looking at these creatures you would never guess that elsewhere during their lifetime a great greening was in the process of softening the face of the Earth.
The two different faces of the Devonian—marine versus the deposits of lakes and mountain basins—are a kind of temporal schizophrenia. The non-marine rocks were lumped together as Old Red Sandstone, and it was some time before it was proved to everybody’s satisfaction that these richly coloured red rocks recording life’s greatest adventure were the exact contemporaries of unremarkable pale limestones and dark shales. They looked so different, and had so few fossils in common. There was precious little to link the plants and fish of the Caledonian basins with the marine rocks which contained trilobites and brachiopods galore. This led to a grand controversy in the nineteenth century as to the identity and age of the Old Red Sandstone. Martin Rudwick has described how the Old Red problem was settled, using as his source prolific contemporary letters and documents to bring alive a debate every bit as fiercely contested at the time as the origin of species a little later or the discovery of the AIDS virus today. At its end, one of England’s most peaceful counties was attached to one of the most revolutionary periods in life’s long history.
One of the problems any thoughtful person has in understanding this momentous breakthrough is how to account for such a profound shift by a series of small, stepwise changes in body design. Yet the more evidence that accumulates, the more species are discovered as fossils which show one or another of the features associated with making the change to life on land. Clever anatomists can show that the bones in a reptilian arm can be compared closely with another set of bones in an ichthyostegid fossil. Molecular biologists discover more and more deep similarities in the genes which attest to the common heritage of tetrapods. No, we can take shared descent now as a fact, at once as commonplace as our own parentage and as miraculous as birth itself. The only satisfactory explanation as to why plants and animals pushed further and further into hostile territory is that their daring was rewarded by overwhelming success in reproduction in this virgin habitat. They had more babies, which prospered more, and in turn bred more freely. Such species were, in a word, fitter. This is natural selection at work. If the cradle of these changes was some isolated basin at the edge of a vanished Himalaya, some place where the first ill-engineered limbs could try out their new function, their subsequent reproductive success is attested to by the appearance of tetrapods practically everywhere in the Carboniferous period that followed the Devonian. There are other reasons for this, too, which we shall come to.
Somehow the colonization of land is a more graspable breakthrough than the evolution of the cell, vastly important though that was. It is easier to appreciate the problems that had to be faced, to imagine ourselves as that first doddering tetrapod, to feel the need for breath, or imagine ourselves sliding gratefully beneath dark, damp vegetation to escape the worst of the Sun’s attentions. But just as we can empathize with the animals at the moment of invasion of the land, so we almost inevitably throw our own human values into the scene. We incorporate the language of ambition. We talk, as I have just done several times in the previous paragraph, of achievement and attainment and success. It is easy to turn this story into a drama of the race to be first. I myself find it almost impossible to filter out the idea of the will to succeed from my narrative of life’s changes. Science itself is so often motivated by the will to be first, to attach one’s name to a discovery. The language is strikingly the same. A scientific achievement is described as a “breakthrough.” It allows for the development of a new field, the exploration of a new landscape of discovery. The race to be first is what motivates teams of hollow-eyed experimenters to sit up all night watching the bubble chamber in the hope of catching the fleeting evidence of a fundamental particle. The race to be first pushes hardy palaeontologists into the bleak Gobi Desert in search of new dinosaurs. Teams of medical researchers fight to beat the other team to be the first to publish the structure of a gene, or the properties of a chemical which can suppress the growth of cancerous cells. It is no wonder that I have conflated evolutionary achievement with the process of scientific advance. Of course, there are important differences from natural selection; after all, scientific research deals with things other than reproductive success (unless you stretch a point and regard the successful promulgation of an idea as a kind of fecundity). Temporarily at least, some scientists have even got there first by usurping the credit from somebody else, or by claiming the work of a clever student as their own, or by pulling strings in the Academy of Sciences.
In Nature, on the contrary, there is no sin. Nor can Nature cheat. While there are areas where the morality of scientific discovery is murky, and many a bright mind who has been wronged by a greedy professor, there remains one golden rule: you must not fake your results. Palaeontology is particularly vulnerable to false claims, or to deliberate attempts to mislead. The trust that is axiomatic to science means that if a fossil is reported as coming from a site, and being genuine, well, then you do not examine its bona fides. The Piltdown Man scandal is the best-known case where this trust was abused. The fabrication of a “missing link” between man and ape was a bold conceit, to say the least, and in retrospect it is astonishing that the hoax was not discovered sooner. But the history of palaeontology has been marred, and in some ways enlivened, by other cases of fabrication which are less well-known to the public. There are even several at the stage we have reached in our history.
The case of Professor V. J. Gupta emerged during the 1980s. Gupta’s name was a familiar one to those concerned with the geology of the Himalaya. He had written a textbook on the subject, and had perfectly respectable academic qualifications from the University of Wales, at Aberystwyth, where his former colleagues will vouch for his easy charm of manner and intelligence. When he returned to India and attained high standing at the University of Chandrigar, it may well have seemed he had been given just reward for a valuable career. He published many scientific papers. Of these, a number were concerned with establishing the ages of rocks in remote areas in the Himalaya. Clearly, this bore closely upon the understanding of the geology of the highest mountain chain in the world. The publications described fossils collected by Gupta himself, at some personal risk, because the fossils were snatched boldly from politically difficult areas, or from very inaccessible mountains. The majority of these publications were written with joint authors known for their expertise in one or another group of fossil animals. There were papers on trilobites among them. What happened was that V. J. would send a parcel to the appropriate professor including the fossil material, indicating its importance. The delighted professor would recognize many of the fossils without much difficulty; this was very satisfactory, because the age of the rocks in question was thus established. A fairly effortless paper would be the result, one more for the professor’s list, and another for V. J., and so everyone was happy. The only problem was that some of the fossils did not originate from where they were said to have been found. There were combinations of fossils unique to other, well-known areas—such as Bohemia—and it looked as if they were even preserved in the identical way, as shown, for example, by a distinctive colour or chemistry. It began to appear suspiciously as if the “Himalayan” fossils might have been collected (or even purchased) in other countries during the course of V.J.’s travels. After all, he had spent time in what was then Czechoslovakia, and nothing would have been easier than to pocket a few fossils from one of the well-known localities there. John Talent, an Australian academic, collected together a number of examples of such jiggery-pokery and published a scathing exposé in Nature. Some of the allegedly Himalayan fossils were pinned down to other, well-known localities far from Asia. It seemed that success had even made V. J. careless, because some of the same photographs of specimens appeared more than once, though allegedly from different localities! When you have got away with improbable feats of duplicity then it begins to feel as if you have total immunity; you can get away with anything. So it was with V. J. Some other facts fell into place. The trilobite co-author, Fred Shaw from Lehman College in the Bronx, had been sufficiently interested in the trilobites he had described with Gupta that he wished, not unreasonably, to do some fieldwork in the Himalaya in order to collect more and better ones. He travelled all the way to Chandrigar, on the promise of doing this, only to find that the professor was not available. He had mysteriously disappeared. Poor Fred was rather miffed to make this discovery, but even then he did not immediately assume that he had been the victim of a ruse, such is the belief in the probity of the scientific community. But after Talent’s case had been made, all the work carried out by Gupta was tainted by uncertainty, and if there was good there, it was condemned along with the bad. There was, of course, a scandal. V. J. was stripped of certain of his titles. His regular attendance on the international conference circuit stopped. Certainly, his work in future will be quoted only in the context of doubt. But India is a large country, and Chandrigar is a long way away from London and New York. I am told that V. J. Gupta still draws his salary and still has a position at the university.
As to motive, advancement was probably the most important. Academics rarely become rich, but they may at least become comfortable enough, and influential:
Fame is the spur that the clear spirit doth raise
(That last infirmity of noble mind)
To scorn delights, and live laborious days …
For some, the fame is tempting, but the laborious days are too exhausting, and the delights irresistible.
SUCH DIVERSIONS DO not help the efforts of hundreds of honest workers who are attempting to unravel the story of life. But the Gupta case brings up the question of evidence, and it is an interesting one. How do we know when we are nearing the truth about history? It is quite clear that we know a lot more about the landward move of life than we did fifty years ago. New discoveries in the field are part of it. So are new perceptions of old facts. But even facts are slightly nebulous. After all, it was a “fact” that five toes were universal until seven toes were discovered by looking again at an “old” fossil. Without impugning the honesty of any individuals, all palaeontologists know that some observers are more reliable than others. Some scientists, you might say, are better at recognizing facts. We are becoming used to the idea in particle physics that the observer is part of the equation. Perhaps this is a more general phenomenon than we admit. In my own history, I can reinterpret the events of my life in retrospect, as I have doubtless modified the autobiographical facts interwoven within this book, if only in the interests of improving the story. Perhaps our fakers lie at the extreme end of a spectrum of reliability in the face of the facts.
If facts are treacherous things, how can we know that we have arrived at the truth? I suspect that we will never know enough about the colonization of land. Year after year there have been surprises, and there is no sign of an end. The next Devonian plant is never quite what we would have predicted from the last one. Every new early land vertebrate springs some sort of surprise. There are good reasons for this. Serendipity is one: the mere chances of finding a fossil are low, and poorer still for early animals that were rare. The rivers in Wales or the glaciers in Greenland carve this way and that at the behest of the elements, not to accommodate scientific researchers. So each discovery is some kind of small miracle, a jackpot on the wheel of chance. Facts may be slippery things, but one thing we know is that we do not have enough of them. Then there is preservation: delicate animals like insects and spiders require a Rhynie Chert to preserve them. The most primitive insects were without doubt wingless, and we know one or two fossils that prove it. They were also very small, and size alone militates against survival as a fossil. So when wings appeared in the fossil record they did so as fully developed structures. A specialist can point to various primitive features, but a non-specialist can recognize them for what they are without prompting. There are no linking fossils yet known, and the chances of finding them now must be slight. So in this case we can say that we have a shortage of facts; we know what we are missing. This leaves the way open for speculation as to the origin of wings, which there has already been in abundance, as we shall see. In other cases, it might not be so clear what still remains to be discovered, such as what plants might have lived around strange pools that may yet serve to link mosses and liverworts or ferns with their ancestors. I am forced back on to a thoroughly practical and rather unsophisticated definition of the limits of ignorance. When continued and persistent collecting reveals only more of what we already know, the searcher eventually gives up through sheer exhaustion. Facts still unknown may be hidden from sight, but it becomes unproductive to wish for what can be sought but not found. The complete reconstruction of history will for ever elude our grasp. There will be little chance of ever discovering direct evidence of insubstantial and tiny worms like nematodes, even if the search were carried on to the end of time. When land was colonized, an invisible phalanx of small animals and plants moved in alongside Ichthyostega and Rhynia. It is salutary to remember that there was always, and will always be, this hidden history.
There are ways of coping with the impossibility of ever knowing history, or completely reconstructing a narrative of the past; a way round being an eternal supplicant for more facts. The solution involves turning speculation about evolutionary history away from arguments about actual descent to theories about relationships. This is relativity rather than narrative. This practice is embodied in the discipline of cladistics. I have spent much of my working life near the centre of cladistic theory. I have been berated for not being a pure enough practitioner of the method by some of the more demanding zealots. Equally, I have been condemned by other traditionalists as a “damned cladist.” Passions have been stirred about the manner in which we gain knowledge of the past that are astonishing in their intensity. Some scientists have not spoken to other colleagues for years, other than in grumbling undertones. Yet cladistics is in no way offensive or unpalatable.
We are all, perhaps, used to seeing evolutionary trees in accounts of the evolution of life. These are similar in intention and design to our own family trees, where it is known whom Uncle Cuthbert fathered, and Aunt Mildred’s sons sired. These are literal, historical narratives of descent, of genealogy. They comprise the unexciting parts at the front of the minor books in the Bible where who begat whom is laboriously chronicled. With ancient history, prior to written records, genealogy is much more problematical. So it is with species. But what we do know is that if evolution happened, then evolutionary changes—whether they affect limbs, such as described in this chapter, or genes, such as periodically emerge throughout this book—will be handed down to future species which shared the same common ancestor. These derived features are the basis of cladistics. Instead of trying to reconstruct actual trees of descent, cladograms are drawn which map out the relationships between species as a series of regularly splitting branches based upon the characteristics of the animals themselves. Every branching point is where a new characteristic was acquired, which is handed down to any species lying above it on the diagram. The animals or plants that descended from a common ancestor comprise a “clade.” Names of groups of animals or plants are given to clades that share a set of characteristics. The groups can be very large—a good fraction of the animal kingdom—or very small, such as a genus comprising a handful of species, but the principle is the same. We have already met the tetrapods, those animals that share four limbs, a vast group if ever there was one. On the other hand, Homo is a genus within the tetrapods comprising intelligent apes sharing a capacity to make tools, walk upright and copulate at all times, not only at oestrus. To construct a cladogram one does not need all the species because the only concern is relative common ancestry. One can easily construct a cladogram showing that humankind is more closely related to pigs than to fish, without implying that men descended from pigs, or that pigs are more human than we might have thought. Such a cladogram would be based on our shared characteristics: with pigs we share (inter alia) our warm blood, four limbs, ear structure, giving birth live and suckling our young. These characteristics are sufficient to show that man and pig belong together in a clade—we call this group the mammals. With fish and pigs we share backbones and paired eyes, and these are just two of the characteristics that show that all three animals belong in a still more inclusive clade—of vertebrates.
If the principle is no more than just stated, then the practice is a good deal more complicated. The more species that are treated on the cladogram, the more potential diagrams of relationships there are, especially because not all characteristics point the same way. For example, we know that both bats and birds have wings; yet bats are not birds, they are mammals. We recognize this because bats share more characteristics with other mammals (suckling their young, fur, and so on) than they do with birds (flight). Our common sense appreciates that bats are flying mammals rather than furry birds. What this natural nous does intuitively is to apply Occam’s razor to the question of the ancestry of bats. We recognize more characteristics linking bats with other mammals than bats with birds; it seems a simpler and preferable arrangement to assume that Nature would have to make fewer alterations to turn an insect-eating mammal into a bat, than to turn a bird into a bat. This simple guiding principle is what William of Occam (fl. early fourteenth century) is said to have determined: given the choice, we prefer an explanation that is simpler. As cladograms get more complicated because of the inclusion of more species and more characters a computer will be required to determine which of many hundreds of possibilities is the simplest arrangement of species.
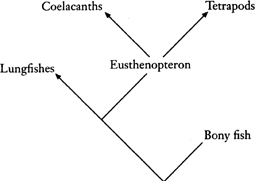
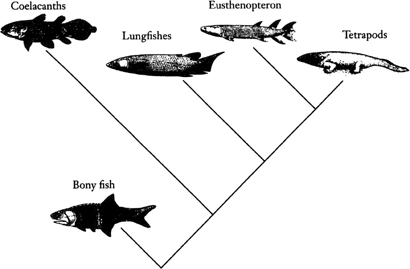
This might seem to be pretty esoteric stuff, and hardly likely to stimulate fury and controversy. Cladistics, however, examines cherished notions about descent from a new standpoint. Quite often new answers emerge, and those answers are not necessarily popular with people who have championed another, traditional view. The origin of terrestrial habits is an example. The tetrapods had been regarded as descending from a curious group of lobe-finned fishes, a view often repeated in popular books. The most well-known lobe-fins have fins with a fleshy central axis which look superficially leg-like, and it seemed a reasonable suggestion that these were the basis for the evolution of that most necessary of terrestrial organs. The living lobe-finned fish, the coelacanth Latimeria, is possibly the most famous of “living fossils,” and its nickname, “old four legs,” speaks clearly of its traditional place in the history of life. It is, indeed, a most lugubrious survivor from a distant age, sluggishly plying its trade in a few places in the Indian Ocean. But there were other kinds of fish living in the streams and lakes of the Devonian. Among them were the earliest lungfishes. There are several species of lungfish still living today, of which the most primitive is Neoceratodus, a slow-moving animal clothed in large scales which now lives only in Australia and copes with low oxygen conditions in water by breathing air, an adaptation that may have seen it through many hard times since the Permian period when its fossil relatives lived. Neoceratodus lacks limb-like fins altogether. The cladists got to work, analysing all the characteristics of lungfish, including subtle features of the bones of the head as well as obvious ones like lungs. They compared these features with what they shared with lobe-finned fish on the one hand, and tetrapods on the other. A heterodox answer was returned: it was the lungfishes, not the classical lobe-fins, that were more closely related to the tetrapods. Our coelacanth was shuffled sideways off the main line of descent: “old would-be four legs.” Naturally, those who had espoused the lobe-fin theory for most of their lives were disbelieving; there were arguments, there were tantrums. But the cladists were probably right. Now that the molecular sequences of molecules can be elucidated (providing yet another list of characteristics) there is additional evidence supporting their view. A return to the fossils showed that early fish like Eusthenopteron had fins which might, after all, be candidates for conversion into limbs. Later descendants of the Devonian lungfish, like Neoceratodus, had lost them over the passage of millions of years. In sum, it was a revolution about the understanding of those seminal days on Devonian shores.
The early protagonists of cladistic theory in the 1970s were united against what was largely a hostile “establishment.” Opposition threw them together, convinced (rightly, in my view) of the value of their perceptions. The founding father of cladistics, the German entomologist Willi Hennig, was elevated to the status of secular saint. His works, I might say, are not an easy read. The new discipline acquired a language of its own, which further alienated those who had not been converted to the methods. (I have avoided all such language here, but the inelegance of it—symplesiomorphy, synapomorphy, semaphoront, to give a few examples—may have had not a little to do with the alienation some have felt.) Cladists talked to other cladists, and those who disbelieved the canon grumbled or raged among themselves. It was a fine example of what the philosophers call a change in paradigm.
A change in a way of seeing the world follows a kind of trajectory. The early, heroic days unite a number of dedicated scientists who follow the developments of the new theory. The invention of a language for the new concepts serves both to reinforce their own identity and to exclude heretics. The inner group of leading practitioners then becomes a kind of high priesthood. Acolytes are admitted if they embrace the canon. Research students are always the most fervent of converts, because they have the energy of youth combined with a desire to please their new masters. They like to be part of the coterie. The Natural History Museum in London, and the American Museum of Natural History in New York, became the centres for theoretical developments of cladistics in the heroic days. Colleagues would enquire sotto voce of a friend, “Are you or aren’t you?” You became a cladist the way you might have become a Buddhist. Some of the most trenchant of the high priesthood were almost preternaturally designed for the role. Fish experts, in particular, dominated; Gareth Nelson, from the American Museum, radiated the fervour of a slightly demented prophet. His lectures had a kind of insouciant brilliance that made you feel really stupid for not understanding what on earth he was on about. Surely, you felt, anything so incomprehensible must be worth unscrambling. The opposite approach was embodied by the fish expert in the museum in London, Colin Patterson, whose presentations combined clarity and charisma in equal measure. Then there was Dick Jefferies, who has esoteric theories of his own about the origin of all vertebrates. Dick is the very embodiment of professordom, with an extensive knowledge of almost everything. He is given to quoting slabs of Chaucer, and can read embryology textbooks in the original German. For added spice there was David Hull, a philosopher of science from New York, who was following the cladistic debate as a kind of living experiment in the way ideas are worked through the scientific community, and who stalked around the department in London listening to everything that was being said.
Some of the antagonists were more colourful than the protagonists. Beverley Halstead was a professor at the University of Reading with a knack for publicity and a capacity for making mischief. He was also strangely likeable considering the trouble he caused. He retained a quirky boyishness and great energy into middle age; he seemed to appeal to women. He had worked extensively on Silurian and Devonian fossil fish, and was thus directly concerned with the implications of cladistics. He did not approve of them. I recall his outburst at a meeting at which Dick Jefferies outlined his theories about the origins of all vertebrates. “Stop this madness, Jefferies, before it’s too late!” he cried. His mischief-making impacted on the Natural History Museum in London after an exhibition opened in 1981, which included representations of the evolution of mankind illustrated by cladistic diagrams. MARXISTS TAKE OVER AT THE NATURAL HISTORY MUSEUM was the kind of headlines that Halstead conjured out of his critique of the exhibition. He had rather ingeniously combined his own two personal bugaboos (cladistics and Marxism) in a rant against those in charge of the exhibits at the museum, comparing the intellectual grip of these people with the kind of implacable orthodoxy which was associated with the then Soviet regime. It was an absurd charge, but it made good publicity. Another charge that hit the papers was that DARWIN WAS DEAD at the Natural History Museum. This was a different misreading of the use of cladograms—the point being that you do not have to assume that evolution has taken place to construct them. Equally, of course, the shared characteristics on which cladograms are based argue for a shared common ancestry; that is, that evolution has taken place. But the debate ran for several months. I sought out the cuttings file, which is quite a handful. In the end, the exhibition was modified somewhat, which no doubt Bev Halstead regarded as a triumph. He would have been nearer the mark if he had noted that cladograms make very dull exhibits, however interesting they may be to the scientists who made them. Bev had an endearing habit of turning up in the Palaeontology Department after one of his salvos looking as pleased as punch, like a naughty but confident schoolboy who had just been caught doing something rude in the school gym. It was a loss to the richness of our intellectual entertainment when he was tragically killed by a lorry a few years ago.
Traditional Evolutionary Tree

1981 Cladogram
1996 Cladogram
The next phase in the trajectory of scientific change begins as the new ideas become part of the mainstream. They become incorporated into undergraduate courses. Cladograms illustrating the relationships between fish and early tetrapods become the norm; those who once fulminated against the ideas retire or go mad, or, as in the case of poor Bev, die early. It is not long before almost everyone is using the new methods. At about the same time, schisms appear among the original adherents. Points of difference become magnified into separate schools. A few of the faithful are excommunicated. Rows break out among people convinced of errors in methodological detail. This is particularly the case when computers come into play because there are few people clever enough to understand algorithms. The whole movement begins to resemble a political party or a religious movement.
In retrospect, it is clear that the cladistic movement is analogous to those that affected other fields of intellectual endeavour in the latter half of the twentieth century. The main thrust was a search for rigour in the way relationships between animals are described: in sum, if you want to classify some organism you should do so in relation to evolved characteristics it shares with other organisms, the features of the thing itself. The modernists in literary criticism attracted an aversion among rival scholars, similar to the brouhaha directed at cladists, in the importance they attached to the primacy of text itself. They tried to unbuckle literature from its historical baggage, the better to understand its workings. As with cladistics, they invented a language which, in the hands of advanced practitioners like Jacques Derrida, could seem at once opaque and infuriating. The beginning of the whole process, in so far as beginnings can be identified at all, was in the structuralism pioneered by the anthropologist Claude Lévi-Strauss. Descriptive anthropology was previously rich with case studies of tribal habits and systems, which, being almost infinite in diversity, required a reductionist view, a discerned set of interpretative principles, to render them more than a mere testimony to mankind’s inventiveness. The change from water to land, this unscrambling of history I have described, is not merely about the discovery of facts; it is also an anthropological study, every bit as human as the intention of a novel.
Discovery does not stop. Some of the more extreme cladists seemed at one time to think that their new methods might render almost obsolete the need to discover new extinct animals. This is dangerous nonsense. There can be no better illustration of the possibility and importance of new discovery than the application of X-rays to the Devonian Hunsrück Shale of Germany. This dark slate contains slightly smeared examples of various animals—trilobites, sea stars, crinoids—as fossils go, not especially appealing. It was known that “soft parts” of animals were preserved in the Hunsrück, apparently coated or replaced by iron pyrites at a very early stage, which painted over the parts that normally decay to invisibility. In the 1970s Wilhelm Stürmer reasoned that the pyrites should register on an X-ray of the slabs. He could scarcely have anticipated the parade of wonderful details his X-rays revealed. These animals were as if frozen on the point of death: starfish petrified with their minutest details conserved; sea lilies with their most delicate, feathery pinnules stretched out to gather seafood, more graceful than a peacock feather; and, just as showy, trilobites with limbs and antennae still scrabbling; and other, strange arthropods that had no record since the Burgess Shale of the Cambrian. These were sea-dwellers, but how much could a discovery of like kind add to our knowledge of the invasion of the land? Whose hammer may yet uncover further secrets?