The capacity to attribute mental states to others (“mindreading” or “theory-of-mind”) was once thought by most empirical researchers and philosophers to be a distinctively human talent. The tide has changed, and in the past eight or nine years there has been a growing consensus in comparative psychology and philosophy that humans are not alone in being mindreaders. It is now generally believed that a number of highly intelligent social animals, from apes to scrub jays, are capable of attributing simple perceptual states, such as seeing and hearing, as well as goals and intentional actions (e.g., Buttelmann et al. 2007; Emery and Clayton 2008; Hare et al. 2000; Hare, Call, and Tomasello 2001; Santos, Flombaum, and Webb 2007; Uller 2004; Wood and Hauser 2008).1
Yet, despite this largely accepted opinion, there is considerable debate in the field over whether animals are capable of attributing beliefs. On the one hand, there are some philosophical arguments (Bermúdez 2003, 2009; Davidson 2001) that purport to show the a priori impossibility of belief attribution in animals and a few empirical studies with chimpanzees that appear to confirm this conclusion (Call and Tomasello 1999; Kaminski, Call, and Tomasello 2008; Krachun et al. 2010). On the other hand, there are at least two empirical studies with chimpanzees that have produced what appear to be opposite findings (Krachun et al. 2008; O’Connell and Dunbar 2003).
I do not believe that the philosophical arguments or the empirical studies on either side of this debate are particularly persuasive. Since I have argued elsewhere (Lurz 2007, 2008, 2011) that the main philosophical arguments against belief attribution in animals are invalid, my focus here will be on the empirical side of the debate. What is needed to move the debate and the empirical field forward, I propose, is a fundamentally new experimental approach to testing belief attribution in chimpanzees, one that is capable of distinguishing genuine belief-attributing subjects from their perceptual-state and behavior-reading counterparts. I present just such an experimental protocol in the last section of this chapter.
Belief Attribution Studies with Chimpanzees
Two types of experimental protocols have been used to test for belief attribution in chimpanzees and other animals. The first type, commonly called the cooperative knowledge-ignorance (or bait-and-switch) paradigm, is designed to test an animal’s ability to use a human experimenter’s communicative gestures to choose between two containers, one of which covers food underneath. Call and Tomasello (1999) ran one of the first of such experiments, but found no evidence that their apes could discriminate between the containers on the basis of whether the experimenter was knowledgeable or ignorant of the food’s true location.
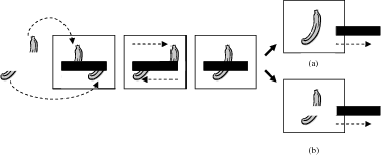
It was thought by some that the chimpanzees failed the cooperative knowledge-ignorance test because it involved cooperative communication regarding the location of hidden food, a type of communicative act that is not part of the natural behavioral repertoire of chimpanzees (see Hare and Tomasello 2004). This, in turn, prompted Kaminski, Call, and Tomasello (2008) to run a competitive version of the knowledge-ignorance experiment. In their experiment two chimpanzees (a subject and a competitor) participated in a competitive game over a highly desirable piece of food (e.g., a grape) hidden beneath one of three cups on a sliding table in a middle area (see figure 17.1).
At the start of the game, the chimps watched while an experimenter baited one of three cups on the table. After the baiting, a screen was positioned (or lowered) in front of the competitor, blocking its view of the middle area. Once the screen was in place, the experimenter performed in front of the subject chimp one of two types of manipulations on the cups and bait. In the unknown-lift trials the experimenter removed the bait from underneath the cup and then replaced it underneath the same cup. In the unknown-shift trials the experimenter removed the bait from underneath the cup and placed it under a different cup. After this, a screen was positioned in front of the subject chimp, blocking its view of the middle area. The screen before the competitor was then removed, and the table with the cups was slid to the competitor’s side. The competitor was allowed to choose a cup by pointing at it. If the baited cup was chosen, the food underneath it was removed and given to the animal; the empty cup was then returned to the table. If an unbaited cup was chosen, the animal was shown that the cup was empty and received no reward; the empty cup was then returned to the table. The subject chimp, of course, could not see which cup the competitor had chosen; although from its prior training in the experiment it knew that the competitor had selected a cup at this point and had received the hidden bait if it had chosen correctly. After the competitor had made its choice and the cups were returned to their original locations, the screen before the subject chimp was removed. The table was then slid to the subject chimp’s side. The subject chimp was allowed to make one of two choices at this point: It was allowed to choose a cup on the table that (depending upon the competitor’s choice) may or may not have the highly desirable piece of food underneath or it was allowed to choose a cup, which was placed inside its booth, that had a less preferred but guaranteed food reward inside (e.g., monkey chow).

FIGURE 17.1. Experimental setup from Kaminski et al. (2008). Reprinted from Cognition, vol. 109, J. Kaminski, J. Call, and M. Tomasello, “Chimpanzees Know What Others Know, But Not What They Believe,” pp. 224–234, 2008, with permission from Elsevier.
Kaminski, Call, and Tomasello (2008) hypothesized that if the subject chimp was capable of attributing beliefs, then it should understand that the competitor in the unknown-shift trials was likely to choose the empty cup on the table, given its false belief regarding which cup was baited, and thus the subject chimp in such trials would be expected to choose the high-quality baited cup on the table. But in the unknown-lift tests the subject chimp should understand that its competitor was likely to choose the baited cup on the table, given its correct belief regarding which cup was baited, and thus the subject chimp in these trials would be expected to choose the low-quality but guaranteed baited cup inside its booth. Kaminski and colleagues found no such preferential choosing on the part of the subject chimp. In fact, the subject chimps were just as likely to choose the high-quality baited cup on the table in the unknown-shift tests as they were in the unknown-lift tests. The authors took these results, plus the findings from Call and Tomasello (1999), as positive support for the hypothesis that chimpanzees are unable to attribute beliefs.
Negative results are always tricky to interpret. And although Kaminski, Call, and Tomasello’s interpretation of the findings is plausible, it is not the only plausible interpretation. It is quite possible that the knowledge-ignorant paradigm (either competitive or cooperative) is simply poorly suited to elicit belief attribution in chimpanzees. Arguably if animals are capable of mindreading, then this is for the express purpose of producing reliable expectations of others’ behaviors. So it stands to reason that mindreading animals should be expected to excel at realistic (perhaps competitive) mindreading tasks that explicitly test their ability to anticipate others’ behavior. And, as a matter of fact, that is what we do seem to find with perceptual-state attribution tests (see Hare 2001; Hare, Call, and Tomasello 2001; Santos, Flombaum, and Webb 2007). However, the knowledge-ignorance paradigm is not designed to test an animal’s ability to anticipate another’s behavior; it is principally a discrimination task, designed explicitly to test an animal’s ability to choose between different containers.2 Thus it is quite plausible that in Kaminski, Call, and Tomasello’s study the subject chimp’s capacity to attribute beliefs was not piqued by the discriminatory nature of the task problem, forcing the animal to employ a nonmindreading strategy to make its selection of cups—perhaps some “high-risk strategy in favor of the high-quality reward in preference to the safe, low-quality reward,” as Kaminski, Call, and Tomasello suggest.
Most recently, Krachun et al. (2010) received negative results from a knowledge-ignorance (bait-and-switch) protocol that was purposively designed to be neither a communicative-cooperative nor competitive paradigm. In the training phase of the experiment, a chimp observed while an experimenter (the baiter) placed a grape or a piece of banana inside a small yellow box with a lid. The baiter closed the lid on the box and placed it underneath one of two colored cups according to the following rule: if the food in the box was a grape, the baiter placed the box under the blue cup; if the food in the box was a piece of banana, the baiter placed the box under the white cup. After the baiter placed the box under the correct cup according to the protocol, the chimp was then allowed to select one of the cups on the table. Five of the six chimps tested on this phase of the experiment eventually learned to select the white cup if they had observed the baiter place a banana piece in the yellow box and the blue cup if they had observed the baiter place a grape in the yellow box.
Once the chimps reached criterion on this phase of the experiment, they were then given a false-belief test and a true-belief test. In the former the chimp observed while the baiter placed a grape or a banana piece inside the yellow box (as in the training phase), closed the lid of the box, and then left the testing room. While the baiter was absent, the chimp then observed another experimenter (the switcher) replace the contents of the box with the alternative food item (e.g., if the box contained a grape, the switcher removed the grape and replaced it with a banana piece). After the switcher switched the contents of the box and closed its lid, the baiter returned to the room. A screen was then raised in front of the chimp, blocking its view while the baiter placed the yellow box under the appropriate cup according to what item the baiter (not the switcher) had placed inside it. The screen was then removed and the chimp was allowed to choose one of the cups. The design of the true-belief test was the same except that the baiter remained in the room and watched while the switcher switched the contents of the box.
The researchers reasoned that if the chimps understood that the baiter in the false-belief trials had a mistaken belief regarding the contents of the yellow box, then they should choose the cup that corresponded to the yellow box’s original contents and not to its actual contents. Likewise, if the chimps understood that the baiter in the true-belief trials had a correct belief regarding the contents of the yellow box (having witnessed the switch by the switcher), then they would be expected to choose the cup that corresponded to the box’s actual contents, not its original contents.
What Krachun and colleagues discovered, however, was that, across the different test trials, their chimps chose the cup according to the yellow box’s actual contents. One rather plausible explanation of these results, the researchers suggest, is that the chimps “might have unthinkingly carried the strategy they learned during the training trials over to the true and false belief test trials (e.g., to choose based on the current contents of the yellow container)” (2010:162).3 That is to say, the chimps may have failed to comprehend the task presented to them in the training trials as one requiring the prediction of the baiter’s future behavior in light of what the baiter currently believes about the contents of the box, but rather one requiring them to discriminate between colored cups according to a conditional rule that had nothing directly to do with the mental states of the baiter. Hence it is not implausible to suppose that the failure of the chimpanzees on the Krachun et al. (2010) test is due not to their inability to attribute beliefs but to their inability to understand the tasks at hand as requiring or involving mindreading as opposed to requiring or involving discrimination according to a nonmindreading conditional rule.
The conclusion I wish to draw here is that a more feasible belief-attribution test for chimpanzees, one that is more likely to trigger their belief-attributing capacities, is one in which the test animal is required to anticipate a target’s behavior. This is the method used in the experimental protocol described in the last section.
Theoretical Background: The ARM Theory
The experimental protocol I will outline is based upon an appearance-reality mindreading (ARM) theory, various versions of which have been defended over the years by Humphreys (1980), Gallup (1982), and Carruthers (1996). According to theorists of ARM, mental-state attribution in animals evolved for the express purpose of anticipating a target’s (e.g., a conspecific’s, predator’s, or prey’s) behavior in those situations in which the animal’s behavior-reading counterparts (i.e., conspecifics that lack mental-state concepts) could not. In many cases the way things perceptually appear to a target is a better predictor of its behavior than the way things objectively are. Behavior-reading animals—animals that are unable to attribute mental states to targets to predict their behaviors—can appeal only to the latter sorts of objective, observable, mind-independent facts (e.g., facts about a target’s past behaviors or its current line of eye gaze to an object in the environment) to predict targets’ future behaviors. Mindreading animals, however, are able to appeal to the subjective way things perceptually appear to a target to predict its behavior. It is hypothesized that mental-state attribution in animals evolved as a result of their coming to introspect their own ability to distinguish appearance from reality, using this introspective distinction for the purpose of anticipating others’ behaviors.
Of course, targets can sometimes see through deceptive appearances and act on what they believe to be really occurring in the environment. And so, on the ARM theory, mental-state attribution in animals should reflect two levels of development. The first level involves perceptual-state attribution, wherein the animal is able to anticipate a target’s behavior in terms of how it thinks the world perceptually appears to the target. The second level of development is belief attribution, wherein the animal is able to anticipate a target’s behavior in terms of how it thinks the target believes things are, the way things perceptually appear to the target notwithstanding.
To test ARM empirically, we need an experimental method capable of distinguishing (a) those animals capable of attributing perceptual states (e.g., seeing or visually appearing) from (b) those capable of attributing perceptual states and beliefs. All the belief-attribution studies just surveyed, unfortunately, rest upon a particular view of the difference between belief attribution and perceptual-state attribution that prevents them from experimentally distinguishing (a) and (b). The view itself, quite correctly, is based upon an undeniable fact about beliefs and perceptions—namely, that past veridical perceptions (e.g., having seen food placed in location x) can sometimes lead a subject to later act on a false belief at a time when the veridical perception no longer exists (e.g., after the deceptive switching of the bait to location y, the competitor/cooperator now incorrectly believes that the bait is still in location x). The studies, therefore, assume that if chimpanzees are capable of attributing beliefs, and not just current perceptual states, then they should be able to predict a target’s behavior on the basis of his/her currently held false belief in the absence of the veridical perception that caused it. The chief problem with this approach is that a target’s currently held false belief will always be confounded with his/ her having had a past veridical perception (as well as his/her having had a direct-line-of-gaze to the perceived object), and, as a result, the animal’s successful prediction of the target’s action can just as well be explained in terms of it attributing a past veridical perception (or a past direct-line-ofgaze to an object) to the target as it can in terms of it attributing a currently held false belief.

FIGURE 17.2. Müller-Lyer diagram.
Luckily there are two other ways of understanding the difference between belief attribution and perceptual-state attribution that escape this problem. Both these approaches reflect a distinctive feature of belief in relation to perception—namely, revisability and abstractness. A distinctive feature of beliefs (at least as a kind of mental state) is that they are revisable in light of countervailing evidence in a way that perceptual states (as a kind) largely are not.4 As a result of measuring the lines in a Müller-Lyer diagram (see figure 17.2), for example, one’s belief about the lines’ unequal length changes despite their continuing to look unequal, and this change in one’s belief naturally leads to a change in one’s behavior.
Thus an animal capable of attributing beliefs, and not just perceptual states, is expected to be sensitive to the behavioral difference between 1. a target that currently perceives an object as F but has countervailing evidence indicating that the object is really not F, and 2. a target that currently perceives an object as F but has no such countervailing evidence. In Lurz (2009, 2011) I have described a number of innovative experimental protocols for testing chimpanzees’ and other animals’ sensitivity to this epistemic difference; so I shall not discuss those here.
The other distinctive feature of belief (and thought in general) is its ability to allow subjects to represent facts or properties in the world that cannot be represented by means of perception alone. Such facts and properties are routinely called “abstract.” Of course, abstractness comes in degree, and so it is unlikely that there will be any sharp distinction between perception and cognition on this feature. Be this as it may, the point here is merely that the more abstract a fact or property a mental state represents, the more the state belongs to cognition. And this is surely correct. Although I do not have a metric of abstractness to offer, it is quite clearly the case that representing an object as having a second-order (or determinable) property (i.e., a property that is defined in terms of other properties) is more abstract than representing an object’s first-order (determinate) properties (i.e., properties that are not defined in terms of other properties). Consider, for example, the second-order (determinable) property of having shape in general. Intuitively, representing an object as having shape in general is more abstract than representing it as square or circular, for example. And, for similar reasons, representing a pair of objects as being same/different in shape in general is more abstract than representing them as having the same/different determinate shape (e.g., as both being square).
Following Premack (1983), cognitive scientists have come to call relations of same/difference in second-order (determinable) properties of color or shape second-order relations and relations of same/difference in determinate colors or shape first-order relations, taking mental representations of the former (abstract) relations as a mark of genuine cognition. It would be ideal, then, in developing a test for belief attribution in chimpanzees, to run an experiment that tested the apes’ capacity to attribute representations of second-order relations to targets. Just such a protocol can be developed, I believe. Chimpanzees and other great apes, after all, are known for their distinctive talent in solving and responding to relational match-to-sample (RMTS) tasks that arguably require representing abstract, second-order relations or properties (see Premack 1983; Smith et al. 1975; Thompson and Oden 2000; Thompson, Oden, and Boysen 1997; Vonk 2003; Vonk and MacDonald 2002, 2004).5 The idea behind the experimental protocol I am proposing here is to exploit this unique cognitive talent in chimpanzees and see whether they can attribute such abstract representations (i.e., beliefs) to others.
Testing the Attribution of Abstract Representations by Chimpanzees
The following belief-attribution test employs a violation-of-expectancy (VOE) paradigm with video-animated stimuli using looking time as a measure—a methodology that is commonly used in developmental psychology and becoming more so in animal studies (see e.g., Csibra 2008; Gergely et al. 1995; Heider and Simmel 1944; Kuhlmeier, Wynn, and Bloom 2003; Premack and Premack 1997; Premack and Woodruff 1978; Surian, Caldi, and Sperber 2007; Uller 2004). However, unlike any VOE experiment to date, the following experiment requires the test animal to be able to make an appearance-reality distinction involving occluded objects. Therefore, the belief-attribution experiment will include a series of pretests designed to probe the chimpanzees’ ability to discriminate appearances from reality involving partially occluded objects.
APPEARANCE-REALITY (A/R) DISCRIMINATION TEST USING COMPUTER ANIMATED STIMULI
Abundant empirical evidence shows that chimpanzees and monkeys are susceptible to some of the same perceptual illusions as humans (for review see Fujita 2006). One well-known example of this is the case of seeing occluded objects as unified, typically referred to as amodal completion.6 Using a two-choice delayed matched-to-sample task, Sato, Kanazawa, and Fujita (1997), for example, showed that an adult chimpanzee reliably matched a unified straight bar when presented with an occluded figure (see figure 17.3a) whose top and bottom portions were aligned and moving in a congruent manner, but matched a broken bar when presented with an occluded figure (see figure 17.3b) whose top and bottom portions were misaligned and moving in an incongruent manner.
This and other amodal completion experiments have been taken as evidence that chimpanzees see occluded figures as having certain features (e.g., being straight or unified) even though the figures may not in fact have such features. Chimpanzees appear to be subject to a type of amodal completion illusion. What researchers have not investigated, however, is whether chimpanzees (or any other animal) can distinguish the apparent from the real shape of an occluded object in those cases where they are given clear evidence that appearances are deceptive.7 The appearance-reality (A/R) task I will describe asks whether chimpanzees, when given evidence that an occluded figure is actually bent or disjointed, despite it looking straight or whole, take the figure to be as it really is (i.e., as bent or disjointed) or as it appears to be (i.e., as straight or whole). If chimpanzees are capable of distinguishing appearances from reality in such cases, then they should take the occluded figure to be bent or disjointed, appearances to the contrary. If they are incapable of making the distinction, then they should take the occluded figure to be as it finally appears to be (i.e., as a straight or whole object behind an occluder).

FIGURE 17.3. Amodal completion stimuli.
Figure 17.4. Simple A/R test using computer animation.
Although different methods can be used to answer these questions (e.g., match-to-sample or delay match-to-sample tasks, such as used in Sato, Kanazawa, and Fujita 1997), I present here a habituation/dishabituation procedure using computer-animated stimuli. Figure 17.4 shows a series of still frames from the videos that could be used in the experiment. In the habituation phase of the experiment, the animal is shown a video of two pieces of a disjointed object (e.g., two pieces of banana or two pieces of a rod) being placed on top of an occluder. (Note that the width of the rectangular occluder is the same size as the missing section between the two banana pieces. Thus, when the banana pieces align themselves on top of the occluder in the third frame, there will still be a missing portion between them that is the width of the occluder.) The object’s continued disjointedness can be further demonstrated by moving the pieces on top of the occluder in opposite directions. The pieces are then brought into alignment, giving the illusion of a whole occluded object, as illustrated by the third frame in figure 17.4. This ends the habituation phase of the experiment. The testing phase occurs immediately afterward. In this phase the occluding bar is removed to reveal either the occluded object (17.4a) as whole or (17.4b) as disjointed.8
If the animal is able to make an appearance-reality distinction in the case of occluded objects, then it should be able to comprehend that, from the countervailing evidence given in the habituation phase, there really is not a whole object (banana) behind the occluder, despite it coming to look that way once the pieces are aligned. Such an animal, then, should be surprised (look longer) to see a whole object (17.4a) emerge from behind the occluder once the occluder is removed but unsurprised (look less) to see a disjointed object (17.4b) emerge. On the other hand, if the animal is unable to make an appearance-reality distinction and simply thinks “if it looks like a whole object (banana) behind the occluder, then it is a whole object (banana) behind the occluder,” then it should be surprised (look longer) to see a disjointed object (17.4b) emerge from behind the occluder but unsurprised (look less) to see a whole object (17.4a) emerge.
Follow-up tests can be run using a set of different types of objects, both familiar and unfamiliar to the animal, as well as different types of occluders and settings (e.g., vertical as opposed to horizontal occluders, three-dimensional backgrounds as opposed to two-dimensional or blank backgrounds). Those animals that pass this A/R test will be allowed to participate in the belief-attribution test.
BELIEF ATTRIBUTION TEST
Those animals that test positive on the A/R task will participate in the following belief-attribution test using a VOE paradigm with computer ani-mation. In the familiarization phase of the experiment, the test animal is habituated to the movements of a computer-generated image (CGI) of a conspecific (perhaps one with which the test animal is familiar). Figures 17.5 and 17.6 show a still frame from the habituation videos. The chimpanzee in the still frames is the CGI figure that the test animal is watching, not the test animal viewing the video. The video sequence opens with the CGI figure standing in front of two trees. The CGI figure at this point is facing away from the trees and toward the test subject watching the video. While the CGI figure’s head is turned away from the trees, two sample stimuli (e.g., a red and blue rectangle) from offscreen silently move onto the screen and place themselves behind the trees, as shown in step 1 in Figures 17.5 and 17.6. After the stimuli are in place, the CGI figure turns around to face the occluded rectangles, as shown in step 2 in figures 17.5 and 17.6. The CGI figure now has the same view of the occluded stimuli as the test subject. After the CGI figure has turned around and has looked at each of the occluded stimuli, it knuckle walks down one of the two paths. Thus there will be two types of habituation videos shown to the test animal. In those videos where the sample stimuli are the same in shape (as illustrated in figure 17.5), the CGI figure knuckle walks down the right path.

Figure 17.5. Habituation phase with same-shape stimuli. While the CGI figure is on screen and facing away from the trees (not shown here), 1. the same-shaped stimuli (red and blue rectangles) come from offscreen and place themselves behind the trees. 2. The CGI figure then turns around to face the occluded stimuli (as shown here). After looking toward each occluded shape, the CGI figure knuckle walks down the right path (white arrow).
In those videos where the sample stimuli are different in shape (as illustrated in figure 17.6 below), the CGI figure knuckle walks down the left path.9
The test animal is shown these two types of habituation videos—those in which the sample stimuli are the same in shape and the CGI figures takes the right path and those in which the sample stimuli are different in shape and the CGI figure takes the left path—a number of times to induce habituation (i.e., the expectation of the CGI figure’s actions in the presence of same- or different-shaped stimuli). Various shapes (e.g., circles and triangles) can be used as sample stimuli in the habituation videos.
Figure 17.6. Habituation phase with different-shaped stimuli. While the CGI figure is on screen and facing away from the trees (not shown here), 1. the different-shaped stimuli (red rectangle and yellow crescent) come from offscreen and place themselves behind the trees. 2. The CGI figure then turns around to face the occluded stimuli (as shown here). After looking toward each occluded shape, the CGI figure knuckle walks down the left path (white arrow).
FIGURE 17.7. “Look-same-but-are-different” test video. While the CGI figure is on screen and facing away from the trees (not shown here), 1. the different shaped stimuli come from offscreen and place themselves behind the trees. 2. The CGI figure then turns around to face the occluded stimuli (as shown here).
In the testing phase of the experiment, the test animal watches two types of similar videos in which a new set of occluding stimuli is used. In the “look-same-but-are-different” test video, two sample stimuli (e.g., an amputated orange oval and a whole blue oval) come on the scene. The pieces of the amputated orange oval align themselves to the sides of the left tree while the whole blue oval moves behind the right tree (see figure 17.7).
FIGURE 17.8. “Look-different-but-are-same” test video. While the CGI figure is onscreen and facing away from the trees (not shown here), 1. the same-shaped stimuli come from offscreen and place themselves behind the trees. 2. The CGI figure then turns around to face the occluded stimuli (as shown here).
To reinforce to the viewing chimpanzee (test animal) that the disjointed pieces that are attached to the left tree do not constitute an occluded whole oval, the pieces themselves can be moved in opposite directions before aligning, much in the way that the banana pieces were moved in the A/R task present earlier. The final placement of the stimuli gives the illusion that there are two similarly shaped objects behind the trees (see figure 17.7). After the placement of the different sets of shapes, the CGI figure turns around to look at the sample stimuli, just as in the habituation videos. At this point the test animal is shown one of two different endings to the video. In the “surprise” ending the CGI figure is shown knuckle walking down the left path.10 In the “expected” ending the CGI figure is shown knuckle walking down the right path. For each ending, the test animal’s looking time is measured.
The second test video is the “look-different-but-are-same” video, in which two sample stimuli of the same shape come onto the screen and move behind the trees (see figure 17.8).
After the placement of the stimuli, the CGI figure turns around and looks at them, as in the habituation videos. The test animal is then shown one of two endings. In the “surprise” ending the CGI figure is shown knuckle walking down the right path.11 In the “expected” ending the CGI figure is shown knuckle walking down the left path. Again, for each of these endings, the test animal’s looking time is measured.
It goes without saying that different types of “deceptive” shapes can be used in the test videos (see figure 17.9) and that the quality of the moving images in the videos themselves will be more detailed than the schematic representations given here.
The beauty behind the belief-attribution protocol is that it is capable of discriminating between belief-attributing chimps and their perceptual-state and behavior-reading counterparts. If those chimps that pass the A/R task are also capable of attributing beliefs involving second-order relations, then they should be able to understand that the CGI figure takes the right path when it believes the occluded objects are the same in shape (irrespective of their determinate shapes and colors) and the left path when it believes the occluded objects are different in shape (again, irrespective of their different determinate shapes or colors). Armed with this understanding of the CGI figure’s behavior in the habituation videos, the chimps should be able to understand that in the look-same-but-are-different video the CGI figure likely believes that the occluded objects are the same in shape, since they look to be the same in shape and the CGI figure did not see the objects go behind or align themselves to the trees. And so, on the basis of this second-order belief attribution, the chimps should anticipate the CGI figure taking the right path and look longer at the surprise ending than the expected one.
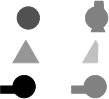
FIGURE 17.9. Various deceptive shapes.
Likewise, in the look-different-but-are-same video, the chimps should understand that the CGI figure likely believes that the occluded objects are different in shape—again, since they look different in shape and the CGI figure did not see them go behind the trees. The chimps should thus predict that the CGI figure will move down the left path and should look longer at the surprise ending than at the expected one.
The chimpanzees would not have such expectations of the CGI figure’s actions, however, were they capable of attributing only first-order representations (i.e., perceptual states) to the CGI figure. Although such chimpanzees would be capable of understanding that the CGI figure mistakenly sees the occluded objects as having the same determinate shape (both oval) in the first test video and different determinate shapes (circle and packman) in the second, they would not be able to predict the CGI figure’s actions on the basis of either of these first-order perceptual-state representations. Since the determinate shapes of the occluded objects on these test trials are novel, the CGI figure was never shown to the chimps as ever having such first-order perceptual representations in the habituation videos. Thus first-order perceptual-state-attributing chimps would have no way of knowing how the CGI figure might behave on the basis of such novel first-order perceptual states. The point of introducing novel shapes (real and apparent) into the test videos is precisely to test the chimpanzees’ ability to rise to the more abstract second-order level of mental-state attribution in their understanding of the CGI figure’s actions.
The last prediction we need to consider is that of the behavior-reading chimpanzee that passed the A/R task. These chimps cannot attribute first-order or second-order representations to targets (such as the CGI figure), but they can themselves represent first-order and second-order relations in the world. Hence these chimps, through their viewing of the habituation videos, could come to understand that the CGI figure takes the right path when the occluded objects actually have the second-order relation of being same in shape, taking the left path when they have the second-order relation of being different in shape. The question, then, is how these chimps would represent the second-order relation of the occluded objects in the test videos. Do they represent them as they actually are or as they appear to be? If they represent them as they are, then these behavior-reading chimps would make the exact opposite predictions of the CGI figure’s behavior in the test videos from their belief-attributing counterparts. They would expect the CGI figure in the look-same-but-are-different test video to knuckle walk down the left fork, since the sample stimuli are in fact incongruent shapes, and this is how the CGI figure responded to incongruent shapes in the habituation videos, and they would expect the CGI figure in the look-different-but-are-same test video to knuckle walk down the right fork, since the sample stimuli are in fact congruent shapes, and this is how the CGI figure responded to congruent shapes in the habituation videos.
On the other hand, if the behavior-reading chimps represent the sample stimuli as having the second-order relations that they appear to have, then these behavior-reading chimps would make the same predictions of the CGI figure’s actions as their belief-attributing counterparts. However, it needs to be remembered that the point of the prescreening A/R test was to screen out such chimps in the first place. If the chimps take the occluded objects as having the second-order relation that they appear to have, despite their being shown that the objects really do not instantiate these second-order relations, then these chimps are not able to make an appearance-reality distinction involving occluded objects, and so they would not be included as subjects in the belief-attribution test. Thus the only behavior-reading chimps that could plausibly squeeze through the A/R test are those that would be expected to represent the occluded objects in the test videos as having the second-order relations that they actually have, appearances to the contrary. But these chimps, as noted, would be expected to react differently to the different endings in the test videos from their belief-attributing counterparts.
Thus, unlike any test used in the past or currently being used, this belief-attribution test has the power to distinguish genuine belief-attributing chimpanzees (as well as other great apes) from their behavior-reading and perceptual-state attributing counterparts. It has been the challenge of designing an experimental approach capable of making this distinction that has severely retarded progress in animal mindreading research (see Fitzpatrick 2009; Heyes 1998; Lurz 2009, 2011; Penn and Povinelli 2007; Povinelli and Vonk 2006). The experimental protocol proposed here has the potential to move the field forward. Finally, it is relevant to note that this experimental approach can be used to test belief-attribution in infants and severely aphasic subjects, which are areas of research in need of more sensitive diagnostic measures of belief attribution (see Song and Baillargeon 2008; Varley 2001).
Notes
I wish to thank Marietta Dindo, Bob Mitchell, and Julie Smith for their very helpful comments on an earlier draft of this essay and Whitney Pillsbury for creating the images for figures 17.5–17.8.
References
Bennett, J. 1976. Linguistic Behaviour. Indianapolis, IN: Hackett.
Bermúdez, J. L. 2003. Thinking Without Words. Oxford: Oxford University Press.
——. 2009. “Mindreading in the Animal Kingdom.” In R. Lurz, ed., The Philosophy of Animal Minds, pp. 145–164. Cambridge: Cambridge University Press.
Burdyn, L., and R. Thomas. 1984. “Conditional Discrimination with Conceptual Simultaneous and Successive Cues in the Squirrel Monkey (Saimiri sciureus).” Journal of Comparative Psychology 4:405–413.
Buttelmann, D., M. Carpenter, J. Call, and M. Tomasello. 2007. “Encultured Chimpanzees Imitate Rationally.” Developmental Science 10:F31–F38.
Call, J., and M. Tomasello. 1999. “A Nonverbal False Belief Task: The Performance of Children and Great Apes.” Child Development 70:381–395.
Carruthers, P. 1996. Language, Thought and Consciousness. Cambridge: Cambridge University Press.
Csibra, G. 2008. “Goal Attribution to Inanimate Agents by 6.5-Month-Old Infants.” Cognition 107:705–717.
Davidson, D. 2001. Subjective, Intersubjective, and Objective. Oxford: Oxford University Press.
Emery, N. J., and N. S. Clayton. 2008. “How to Build a Scrub-Jay That Reads Minds.” In S. Itakura and K. Fujita, eds., Origins of the Social Mind, pp. 65–98. Tokyo: Springer.
Fagot, J., E. Wasserman, and M. Young. 2001. “Discriminating the Relation Between Relations: The Role of Entropy in Abstract Conceptualization by Baboons (Papio papio) and Humans (Homo sapiens).” Journal of Experimental Psychology 27:316–328.
Fitzpatrick, S. 2009. “The Primate Mindreading Controversy: A Case Study in Simplicity and Methodology in Animal Psychology.” In R. Lurz, ed., The Philosophy of Animal Minds, pp. 258–277. Cambridge: Cambridge University Press.
Fujita, K. 2006. “Seeing What Is Not There: Illusion, Completion, and Spatiotemporal Boundary Formation in Comparative Perspective.” In E. A. Wasserman and T. R. Zentall, eds., Comparative Cognition, pp. 29–52. Oxford: Oxford University Press.
Gallup, G. 1982. “Self-Awareness and the Emergence of Mind in Primates.” American Journal of Primatology 2:237–248.
Gergely, G., Z. Nadasdy, G. Csibra, and S. Biro. 1995. “Taking the Intentional Stance at Twelve Months of Age.” Cognition 56:165–193.
Hare, B. 2001. “Can Competitive Paradigms Increase the Validity of Experiments on Primate Social Cognition?” Animal Cognition 4:269–280.
Hare, B., J. Call, B. Agnetta, and M. Tomasello. 2000. “Chimpanzees Know What Conspecifics Do and Do Not See.” Animal Behaviour 59:771–785.
Hare, B., J. Call, and M. Tomasello. 2001. “Do Chimpanzees Know What Conspecifics Know?” Animal Behaviour 61:139–151.
Hare, B., and M. Tomasello. 2004. “Chimpanzees Are More Skillful in Competitive Than in Cooperative Cognitive Tasks.” Animal Behaviour 68:571–581.
Harman, G. 1978. “Studying the Chimpanzee’s Theory of Mind.” Behavioral and Brain Sciences 4:576–577.
Heider, F., and M. Simmel. 1944. “An Experimental Study of Apparent Behavior.” American Journal of Psychology 57:243–259.
Heyes, C. 1998. “Theory of Mind in Nonhuman Primates.” Behavioral and Brain Sciences 21:101–148.
Humphrey, N. 1980. “Nature’s Psychologists.” In B. Josephson and V. Ramachandran, eds., Consciousness and the Physical World, pp. 57–75. Oxford: Pergamon.
Kaminski, J., J. Call, and M. Tomasello. 2008. “Chimpanzees Know What Others Know, But Not What They Believe.” Cognition 109:224–234.
Kanizsa, G. 1979. Organization in Vision: Essays on Gestalt Perception. New York: Praeger.
Krachun, C., J. Call, and M. Tomasello. 2009. “Can Chimpanzees Discriminate Appearances from Reality?” Cognition 112:435–450.
Krachun, C., M. Carpenter, J. Call, and M. Tomasello. 2008. “A Competitive Nonverbal False Belief Task for Children and Apes.” Developmental Science 12:521–535.
——. 2010. “A New Change-of-Contents False Belief Test: Children and Chimpanzees Compared.” International Journal of Comparative Psychology 23:145–165.
Kuhlmeier, V., K. Wynn, and P. Bloom. 2003. “Attribution of Dispositional States by Twelve-Month-Old.” Psychological Science 14:402–408.
Lurz, R. 2007. “In Defense of Wordless Thoughts About Thoughts.” Mind and Language 22:270–296.
——. 2009. “If Chimpanzees Are Mindreaders, Could Behavioral Science Tell? Toward a Solution to the Logical Problem.” Philosophical Psychology 22:305–328.
——. 2011. Mindreading Animals. Cambridge: MIT Press.
O’Connell, S., and R. Dunbar. 2003. “A Test for Comprehension of False Belief in Chimpanzees.” Evolution and Cognition 9:131–140.
Penn, D., and D. Povinelli. 2007. “On the Lack of Evidence That Non-human Animals Possess Anything Remotely Resembling a ‘Theory of Mind’.” Philosophical Transactions of the Royal Society B 362:731–744.
Penn, D., K. Holyoak, and D. Povinelli. 2008. “Darwin’s Mistake: Explaining the Discontinuity Between Human and Nonhuman Minds.” Behavioral and Brain Sciences 31:109–178.
Povinelli, D., and J. Vonk. 2006. “We Don’t Need a Microscope to Explore the Chimpanzee’s Mind.” In S. Hurley and M. Nudds, eds., Rational Animals, pp. 385–412. Oxford: Oxford University Press.
Premack, D. 1983. “The Codes of Man and Beast.” Behavioral and Brain Sciences 6:125–167.
Premack, D., and A. J. Premack. 1997. “Motor Competence as Integral to Attribution of Goal.” Cognition 63:235–242.
Premack, D., and G. Woodruff. 1978. “Does the Chimpanzee Have a Theory of Mind?” Behavioral and Brain Sciences 1:515–526.
Santos, L., J. I. Flombaum, and P. Webb. 2007. “The Evolution of Human Mindreading: How Nonhuman Primates Can Inform Social Cognitive Neuroscience.” In S. M. Platek, J. P. Keenan, and T. K. Shackelford, eds., Evolutionary Cognitive Neuroscience, pp. 433–456. Cambridge: MIT Press.
Sato, A., S. Kanazawa, and K. Fujita. 1997. “Perception of Object Unity in a Chimpanzee (Pan troglodytes).” Japanese Psychological Research 39:191–199.
Smith, H., J. King, E. Witt, and J. Rickel. 1975. “Sameness-Difference Matching from Sample by Chimpanzees.” Bulletin of the Psychonomic Society 6:469–471.
Song, H., and R. Baillargeon. 2008. “Infants’ Reasoning About Others’ False Perceptions.” Developmental Psychology 44:1789–1795.
Surian, L., S. Caldi, and D. Sperber. 2007. “Attribution of Beliefs by Thirteen-Month-Old Infants.” Psychological Science 18:580–586.
Thompson, R., and D. Oden. 2000. “Categorical Perception and Conceptual Judgments by Primates: The Paleological Monkey and the Analogical Ape.” Behavioural Processes 35:149–161.
Thompson, R., D. Oden, and S. Boysen. 1997. “Language-Naive Chimpanzees (Pan troglodytes) Judge Relations Between Relations in a Conceptual Matching-to-Sample Task.” Journal of Experimental Psychology 23:31–43.
Uller, C. 2004. “Disposition to Recognize Goals in Infant Chimpanzees.” Animal Cognition 7:154–161.
Varley, R. 2001. “Severe Impairment in Grammar Does Not Preclude Theory of Mind.” Neurocase 7:489–493.
Vonk, J. 2003. “Gorilla (Gorilla gorilla) and Orangutan (Pongo abelii) Understanding of First- and Second-Order Relations.” Animal Cognition 6:77–86.
Vonk, J., and S. MacDonald. 2002. “Natural Concept Formation by a Juvenile Gorilla (Gorilla gorilla gorilla) at Three Levels of Abstraction.” Journal of the Experimental Analysis of Behaviour 78:315–332.
——. 2004. “Levels of Abstraction in Orangutan (Pongo abelii) Categorization.” Journal of Comparative Psychology 118:3–13.
Wood, J., and M. Hauser. 2008. “Action Comprehension in Non-human Primates: Motor Simulation or Inferential Reasoning?” Trends in Cognitive Sciences 12: 461–465.