Evolution and Notions of Human Race
Alan R. Templeton
OUTLINE
1. The biological meaning of race
2. Do biological races exist in chimpanzees?
3. Do biological races exist in humans?
4. Do adaptive traits define human races?
5. Do human races exist: The answer
Races exist in humans in a cultural sense, but it is essential to use biological concepts of race that are applied to other species to see whether human races exist in a manner that avoids cultural biases and anthropocentric thinking. Modern concepts of race can be implemented objectively with molecular genetic data, and genetic data sets are used to see whether biological races exist in humans and in our closest evolutionary relative, the chimpanzee.
GLOSSARY
Admixture. Reproduction between members of two populations that previously had little to no reproductive contact.
Alleles. Alternative forms of homologous genes within a species that constitute the most basic type of genetic diversity.
Evolutionary Lineage. A population that maintains genetic continuity and identity over many generations because of little to no reproductive interchange with other populations.
Evolutionary Tree. A depiction of the ancestral relationships that interconnect a group of biological entities through a diagram in which ancestral nodes can split into two or more descendant types but that does not allow fusion of previously split types.
Gene Flow. Movement of individuals or gametes from the local population of birth to a different local population followed by successful reproduction.
Genetic Differentiation. Differences among populations based on particular alleles they possess, the frequencies of shared alleles, or both.
Haplotype. A specific nucleotide sequence existing among the homologous copies of a defined DNA region, whether a gene or not.
Isolation By Distance. A model of gene flow in which most genetic interchange is between neighboring populations but in which genes can spread to distant populations over many generations because there are no absolute barriers to movement between any pair of neighboring populations.
Local Population. A collection of interbreeding individuals of the same species that live in sufficient proximity that most mates are drawn from this collection of individuals.
Race. A subpopulation within a species, also called a subspecies, that has sharp geographic boundaries separating it from the remainder of the species, with the boundaries characterized by a high degree of genetic differentiation defined either through a quantitative threshold or qualitatively as a separate evolutionary lineage.
1. THE BIOLOGICAL MEANING OF RACE
Do human races exist? Many people would answer yes because they have a strong sense of their own racial identity and feel they can classify other people into racial categories. However, the ability to classify oneself and others into races does not mean that races actually exist as a culture-free, biological category. For example, Lao and coworkers (2010) assessed the geographic ancestry of self-declared “whites” and “blacks” in the United States by the use of a panel of genetic markers. It is well known that the frequencies of alleles (different forms of a gene) vary over geographic space in humans. The differences in allele frequencies are generally so modest that any one gene yields only a little information about the geographic origins of one’s ancestors. However, with modern DNA technology, it is possible to infer the geographic ancestry of individuals by scoring large numbers of genes. Self-identified “whites” from the United States are primarily of European ancestry, whereas US “blacks” are primarily of African ancestry, with little to no overlap in the amount of African ancestry between US “whites” and “blacks.” In contrast, Santos and coworkers (2009) did a similar genetic assessment of Brazilians who self-identified themselves as “whites,” “browns,” and “blacks” and found extensive overlap in the amount of African ancestry among all these “races.” Indeed, many Brazilian “whites” are surprised to learn that they are considered to be “blacks” when they visit the United States, and similarly, some US “blacks” are considered to be “whites” by Brazilians. Obviously, the culturally defined racial categories of “white” and “black” do not have the same genetic meanings in the United States and Brazil. It is clear that an objective, culture-free definition of race is required before the question about the existence of biological races can be answered.
One way of ensuring a culture-free definition of race is to use a definition that is applied to species other than humans. The word race is not commonly used in the nonhuman literature; instead, the word subspecies is used to indicate the major types or subdivisions within a species. There is no consensus on what constitutes a species (see chapter VI.1), much less a subspecies. Because the US Endangered Species Act mandates the protection of endangered subspecies of vertebrates as well as endangered species, conservation biologists have developed operational definitions of race or subspecies applicable to all vertebrates. We will apply these culture-free definitions to humans to avoid an anthropocentric definition of race.
Biologically, races are geographically circumscribed populations within a species that have sharp boundaries that separate them from the remainder of the species. In traditional taxonomic studies, the boundaries were defined by morphological differences, but increasingly these boundaries are defined in terms of genetic differences that can be scored in an objective fashion in all species. Most local populations within a species show some degree of genetic differentiation from other local populations by having either some unique alleles or different frequencies of alleles. If every genetically distinguishable population were elevated to the status of race, then most species would have hundreds to tens of thousands of races. This would make the concept of race nothing more than a synonym for a local population. There is a consensus that race or subspecies should refer to a degree or type of genetic differentiation that is well above the level of genetic differences that exist among local populations. Both quantitative and qualitative criteria are used to define these racial genetic boundaries.
Quantitatively, one commonly used threshold is that two populations with sharp boundaries are considered to be different races if 25 percent or more of the genetic variability that they collectively share is found as between-population differences. One of the oldest measures used to quantify these differences is a statistic known as fst. Consider drawing two homologous genes at random from all the genetic variation collectively shared by both subpopulations. The frequency with which these two randomly drawn genes from the total population are different alleles is designated by Ht, the expected heterozygosity of the total population. Now consider drawing two genes at random from just a single subpopulation. Let Hs be the average frequency with which these randomly drawn genes from the same subpopulation are different alleles. Then, fst = (Ht−Hs)/Ht. In many modern genetic studies, the degree of DNA sequence differences between the randomly drawn genes is measured, often with the use of a model of mutation, instead of just determining whether the two genes are the same or different alleles. When this done, the analysis is called an analysis of molecular variance (AMOVA). Regardless of the specific measure, the degree of genetic differentiation can be quantified in an objective manner in any species. Hence, human “races” can indeed be studied with exactly the same criteria applied to nonhuman species. The main disadvantage of this definition is the arbitrariness of the threshold value of 25 percent.
A second definition of race defines the genetic differences qualitatively. Sharp boundaries exist in this case because the species is subdivided into two or more evolutionary lineages. An evolutionary lineage is created within a species when an ancestral population is split into two or more subpopulations, often by some sort of geographic barrier, such that there is no or extremely limited genetic interchange after the split. This means that the subpopulations tend to evolve mostly independently of one another, causing the lineages to accumulate genetic differences with increasing time since the split. Immediately after the split, the subpopulations would share most ancestral polymorphisms (gene loci with more than one allele) and would therefore be difficult to diagnose as separate lineages. With increasing time since the split, genetic divergence accumulates, and diagnosing the separate lineages becomes easier. Unlike the fst definition of race, no arbitrary threshold of differentiation is set a priori. A split into separate lineages also means that the genetic differences among the races would define an evolutionary tree analogous to an evolutionary tree of species. Statistical methods exist for testing the null hypothesis that the genetic variation within a species has a treelike structure, and other statistics test the null hypothesis that the entire sample defines a single evolutionary lineage. Therefore, just as with the fst definition, the lineage definition of race can be implemented for all species in an objective fashion using uniform criteria, thereby avoiding a human-specific or cultural definition of race.
2. DO BIOLOGICAL RACES EXIST IN CHIMPANZEES?
Before addressing the existence of human races, we first apply these definitions of race to our closest evolutionary relative, the chimpanzee. In this manner, the definitions can be applied in a context that avoids the emotion and cultural biases that inevitably creep into discussions of human race.
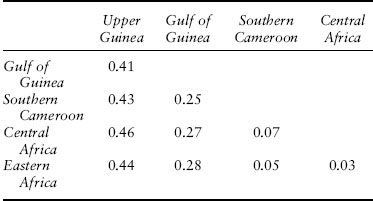
Based on morphological differences, the common chimpanzee (Pan troglodytes) has been subdivided into five races or subspecies: P. t. verus in the Upper Guinea region of western Africa, P. t. ellioti in the Gulf of Guinea region (southern Nigeria and western Cameroon), P. t. troglodytes in central Africa, P. t. schweinfurthii in the western part of equatorial Africa (mostly southern Cameroon), and P. t. marungensis in central and eastern equatorial Africa. Gonder and coworkers (2011) genetically surveyed chimpanzees throughout their range. They discovered sharp genetic differences separating the Upper Guinea and Gulf of Guinea populations from all other populations, but with less sharp genetic boundaries between the equatorial African populations. Table 1 shows the pairwise AMOVA results for these populations. The Upper Guinea and Gulf of Guinea populations are above the 25 percent threshold for contrasts with each other and with all other chimpanzee populations. However, the three regions sampled in equatorial Africa are all well below the 25 percent threshold. Hence, there are three races or subspecies of common chimpanzees using the threshold criterion: P .t. verus in the Upper Guinea region, P. t. ellioti in the Gulf of Guinea region, and the chimpanzee populations from equatorial Africa.
Table 1. Genetic differentiation among populations of chimpanzees as measured by Rst
Source: Modified from Gonder et al. 2011.
Note: Rst is related to fst but incorporates a mutational model for microsatellites.
If chimpanzees are subdivided into separate evolutionary lineages, the genetic differences among lineages should define a treelike structure characterized by splits and isolation. There are genetic differences between different geographic areas (table 1), but such genetic differentiation can also arise when gene flow (genetic interchange associated with individuals who disperse from their birth population) occurs but is restricted by geography. For example, gene flow can be restricted when most dispersal is limited to nearby local populations. Because genes are passed on from generation to generation, a new allele can still spread throughout a species’ range over multiple generations by using local populations as “stepping-stones” to reach more distant local populations. Such stepping-stone models yield a pattern of isolation by distance in which the degree of genetic differentiation between two populations increases with increasing geographic distance between them.
Genetic differentiation structured by isolation by distance can be distinguished from genetic differentiation due to lineage splits by testing for constraints on genetic distances. Consider three hypothetical populations (A, B, and C) such that A is closer to B than to C, and B and C are the closest geographic pair. Under isolation by distance, the genetic distance (measured, say, by the fst value between a pair of populations) should increase with increasing geographic distance; that is, the fst between A and B should be less than the fst between A and C. In contrast, suppose populations A, B, and C represent separate evolutionary lineages (races) such that A split from the common ancestral population of B and C in the past, followed by a more recent split between populations B and C. This results in an evolutionary tree of populations such that genetic distance between populations increases with the time since their split from a common ancestral population. In this hypothetical case, the genetic distances between populations A and B and between populations A and C should be the same, since they both involve a split from the same ancestral population. Hence, the expected pattern of genetic distances differs for trees versus isolation by distance, and formal statistical tests exist to determine whether the pattern of genetic differentiation is consistent with the special constraints imposed by an evolutionary tree.
Another method for testing for a treelike structure is based on finer geographic sampling. As more sites are sampled under an isolation-by-distance model, the geographically intermediate populations should also have intermediate genetic distances. In contrast, when the populations are grouped into a smaller number of evolutionary lineages, genetic distances among populations within a lineage should be relatively small, although they may show an isolation-by-distance pattern within the geographic range occupied by a particular lineage. However, the genetic distances are expected to show a large, sudden increase when the geographic boundary between two lineages is crossed.
When the chimpanzee genetic data are used to estimate an evolutionary tree of populations, the resulting tree has the Upper Guinea population splitting off first, followed by the Gulf of Guinea population, and then splits among the equatorial Africa populations. This tree predicts that the Upper Guinea population should be equally distant from all the other populations, and table 1 shows that this prediction is supported when the error in estimating the distances is taken into account. This tree also predicts that the Gulf of Guinea population should be equally distant from all the equatorial African populations but that this distance should be smaller (less time since the split) than the distances involving the Upper Guinea population. Table 1 shows that this prediction is also supported. However, the genetic distances among the three equatorial African populations show the isolation-by-distance pattern on an east-west axis. These three populations are therefore collapsed into a single lineage. Hence, chimpanzees do show a treelike structure of genetic differentiation with three lineages: Upper Guinea, Gulf of Guinea, and the combined equatorial African populations. Hence, races do exist in chimpanzees under the lineage definition, and they correspond exactly to the same three races defined by the quantitative threshold definition of race.
3. DO BIOLOGICAL RACES EXIST IN HUMANS?
Do human races exist according to the same criteria applied to chimpanzees? In 2002, Rosenberg and others performed a genetic survey of 52 human populations. They used a computer program to sort individuals or portions of their genomes into five groups and discovered that the genetic ancestry of most individuals was inferred to come mostly from just one group. Moreover, the groups corresponded to five major geographic populations: (1) sub-Saharan Africans; (2) Europeans, Near and Middle Easterners, and Central Asians; (3) East Asians; (4) Pacific populations; and (5) Amerindians. This paper was the most widely cited article from the journal Science in 2002, and many of these citations claimed that this paper supported the idea that races were biologically real in humans. However, Rosenberg and coauthors were more cautious in their interpretation. When they increased the number of groups beyond five, they also obtained an excellent classification into smaller, more regional groups. Hence, they showed that with enough genetic markers, it is possible to discriminate most local populations from one another. Recall that genetic differentiation is necessary but not sufficient to define races, so even if there is a consensus that five groups is the right number, genetic discrimination alone does not necessarily mean that these five groups are races.
Assuming for now that the five groups are the meaningful populations, do these groups satisfy the quantitative threshold definition of race? Table 2 shows the AMOVA results for these five groups, along with a comparable analysis of the three races of chimpanzees that satisfy both the threshold and lineage definitions of race. Table 2 shows how the genetic variation is hierarchically partitioned into differences among individuals within the same local population, differences among local populations within the same “race,” and among “races.” Table 2 confirms the reality of race in chimpanzees using the threshold definition, as 30.1 percent of the genetic variation is found in the among-race component, a result expected from the pairwise analysis shown in table 1. In contrast with chimpanzee races, the five major “races” of humans account for only 4.3 percent of the genetic variation—well below the 25 percent threshold. The genetic variation in our species is overwhelmingly variation among individuals (93.2 percent). According to the threshold definition, there are no races in humans.
Table 2. AMOVA of genetic variation in chimpanzees and in humans

Sources: Chimpanzees—data from Gonder et al. 2011; humans—data from Rosenberg et al. 2002.
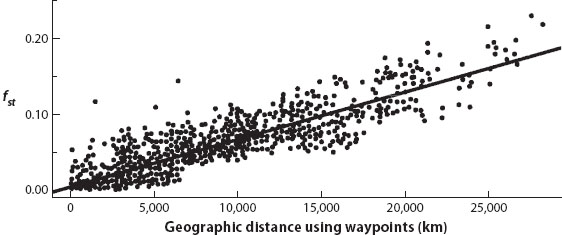
As for the lineage definition, a treelike structure of genetic differentiation has been strongly rejected for every human data set subjected to testing for the constraints expected from an evolutionary tree of populations. Increased geographic sampling further undermines the idea of separate lineages. When Rosenberg and coworkers published their results in 2002, their geographic sampling was coarse. It is now known that the computer program used in these studies generates well-differentiated populations as an artifact of coarse sampling from species characterized by isolation by distance. Figure 1 shows a plot of the pairwise fst values of humans as a function of geographic distance. The results fit well with the predictions of an isolation-by-distance model. Consequently, it is not surprising that when Behar and coworkers (2010) sampled Old World populations more finely and used the same computer program used in the 2002 study, most individuals showed significant genetic inputs from two or more populations, indicating that most human individuals have mixed ancestries. The “races” so apparent to many who cited Rosenberg and coworkers simply disappeared with better sampling. These results and figure 1 falsify the hypothesis that humans are subdivided into evolutionary lineages.
Figure 1. Isolation by distance in human populations. (Modified from Ramachandran et al. 2005. Copyright 2005 National Academy of Sciences, USA.)
Another way of testing for distinct lineages is through a technique known as multilocus nested-clade phylogeographic analysis (ML-NCPA). Many regions of the human genome experience little to no recombination. The distinct genetic states that exist in such regions (called haplotypes) reflect the accumulation of various mutations during evolutionary history. This evolutionary history, called a haplotype tree, is the history of the genetic variation in that genomic region and is not necessarily the history of the populations that bear this variation. Indeed, if a species has sufficient gene flow, there can be no evolutionary tree of populations because there are no population splits; however, there will still be haplotype trees for each nonrecombining region of the genome. Haplotype distributions can be influenced by population-level history, but the population-level information embedded in a haplotype tree must be extracted carefully. It is never justified to equate a haplotype tree directly to an evolutionary history of populations. ML-NCPA provides a statistically rigorous method for making inferences about population history from haplotype trees. No other technique of phylogeographic inference has been so extensively validated as ML-NCPA by both positive controls (data sets for which outside information exists that indicates a known historical event or process) and computer simulation. These validations show that ML-NCPA is not prone to making false-positive inferences about past splits and is very powerful in detecting separate lineages, even when the split is relatively recent and results in haplotype trees affected by retention and sorting of ancestral haplotypes. Moreover, ML-NCPA can detect lineages even when there is some, but very limited, genetic interchange. ML-NCPA does not require an a priori model of the evolutionary history of a species; rather, the history is inferred directly from the haplotype trees using explicit criteria applicable to all species. Finally, each inference made with ML-NCPA is subject to a statistical test for significance.
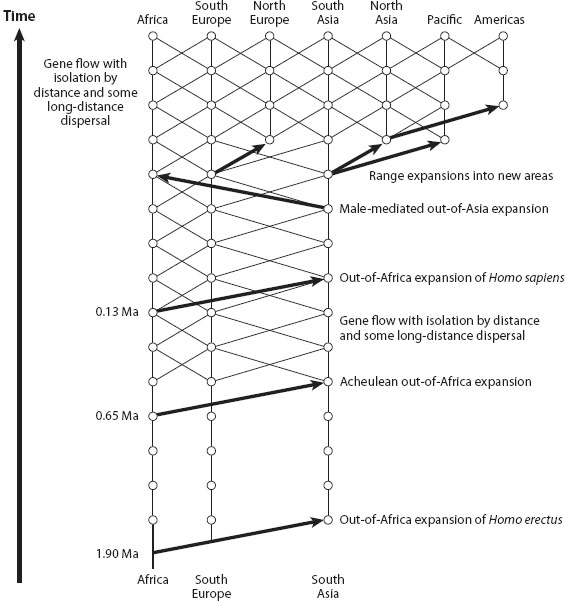
Figure 2 shows the inferences from ML-NCPA about human evolution. The oldest inferred event is an out-of-Africa range expansion into Eurasia genetically dated to about 1.9 million years ago—the same time that the fossil evidence indicates that Homo erectus spread out of Africa into Eurasia during a major wet period in the Sahara. The paleoclimatic data indicate that the Sahara region experienced repeated minor wet periods such that the Sahara is unlikely to have been a dispersal barrier on a timescale of tens of thousands of years. Consistent with these paleoclimatic data, ML-NCPA infers limited genetic interchange with isolation by distance between sub-Saharan Africa and Eurasia starting no later than 700,000 years ago in the Pleistocene. The null hypothesis of complete genetic isolation during the Pleistocene is decisively rejected. Consequently, even during the Pleistocene, Old World human populations were not subdivided into isolated and independently evolving lineages.
The next major event shown in figure 2 is a second population expansion out of Africa into Eurasia around 700,000 years ago, corresponding to the spread of the Acheulean tool culture out of Africa into Eurasia during the second major Saharan wet period of the Pleistocene. The null hypothesis of no admixture between the expanding population and the Eurasian populations is rejected. Hence, the Acheulean expansion was marked by further genetic interchange between African and Eurasian populations, further weakening the hypothesis of isolated Pleistocene lineages of humans. Gene flow then continued until a third major expansion of humans out of Africa into Eurasia occurred around 130,000 years ago, the time of the last major Saharan wet period. The fossil record indicates that modern humans began expanding out of sub-Saharan Africa at 130,000 years ago and reached China no later than 110,000 years ago. The null hypothesis of no admixture is overwhelmingly rejected for this expansion event. This strong rejection of strict replacement without admixing was the most controversial ML-NCPA inference, because the dominant model of human evolution at the time was the out-of-Africa replacement model (see chapter II.18). The inference of admixture has since been supported by studies on fossil DNA of archaic Eurasian populations.
Figure 2. Significant inferences about human evolution from multilocus, nested-clade phylogeographic analysis. Geographic location is indicated on the x-axis, and time on the y-axis, with the bottom of the figure corresponding to 2 million years ago. Vertical lines indicate genetic descent over time, and diagonal lines indicate gene flow across space and time. Thick arrows indicate statistically significant population range expansions, with the base of the arrow indicating the geographic origin of the expanding population. Lines of descent are not broken, because the population range expansion events were accompanied by statistically significant admixture when they involved expansion into a previously inhabited area. (Modified from Templeton 2005.)
Following the expansion with admixture of modern humans from Africa, there have been additional expansions, mostly into areas not formerly occupied by humans (figure 2). Wherever humans lived, gene flow was established, mostly limited by isolation by distance but with some long-distance dispersal as well. On a timescale of tens of thousands of years, there is not one statistically significant inference of splitting or isolation during the last 700,000 years. Because of gene flow and admixture, humans are a single evolutionary lineage. Hence, there are no races in humans under the lineage definition.
4. DO ADAPTIVE TRAITS DEFINE HUMAN RACES?
Races or subspecies, when they exist, always occupy a subset of the geographic range of their species. Sometimes, environmental factors vary over the geographic range of the species, and some of these environmental factors can induce natural selection that results in local adaptation. Hence, when races exist, they sometimes display local adaptations to the environment associated with their geographic subrange that are not adaptive in the remainder of the species’ geographic range. This reasoning leads to the idea that local adaptations can sometimes be biological markers of racial status.
Variation in environmental factors can still induce natural selection that results in local adaptations in species with sufficient gene flow and admixture to prevent race formation. However, in this case, the geographic distributions of the local adaptations reflect the geography of the environmental factors and not racial boundaries. Frequently, different adaptive traits display discordant geographic distributions, thereby indicating that these are simply adaptations of local populations and not markers of higher groupings such as race.
Because humans are not subdivided into races by any of the definitions applied to other species, the locally adaptive traits of humans are not “racial” traits. Skin color is historically the locally adaptive trait most commonly considered a “racial” trait in humans. Skin color is an adaptation to the amount of ultraviolet (UV) radiation in the environment: dark skins are adaptive in high-UV environments to protect cells from radiation damage, and light skins are adaptive in low-UV environments to make sufficient vitamin D, which requires UV radiation. Skin color varies continuously among humans and does not fall into a few discrete “racial” types. Moreover, the geographic distribution of skin color follows the environmental factor of UV incidence and does not reflect overall genetic divergence. For example, the native peoples with the darkest skins live in tropical Africa and Melanesia. The dark skins of Africans and Melanesians are adaptive to the high UV found in these areas. Because Africans and Melanesians live on opposite sides of the world, they are more highly genetically differentiated than many other human populations (figure 1). Europeans, who are geographically close to Africa, are more similar at the molecular genetic level to Africans than Melanesians are to Africans, despite the fact that Europeans have light skins that are adaptive to the low-UV environment of Europe. Hence, skin color is not an indicator of the degree of genetic differentiation, as a true racial trait would be.
Adaptive traits in humans do not define coherent populations. For example, the adaptive trait of dark skin is widespread in sub-Saharan Africa. Another adaptive trait found in Africa is resistance to sleeping sickness, and the responsible gene is found at frequencies up to 80 percent in parts of western Africa where the parasite that causes sleeping sickness is common. However, this adaptive trait is virtually absent in East African populations. Hence, the distribution of sleeping sickness resistance is only a subset of the geographic distribution of dark skin in Africa. Another adaptive trait is resistance to malaria, which is widespread in African populations. However, malaria is also common in some areas outside Africa, and malarial resistance is found in many European and Asian populations as well. Indeed, one of the alleles underlying malarial resistance, the sickle-cell allele, is most frequent in certain populations on the Arabian Peninsula despite often being regarded as a disease of “blacks.” Each adaptive trait in humans has its own geographic distribution that reflects the distribution of the underlying environmental factor for which it is adaptive. The discordance in the distributions of adaptive traits in humans makes them useless in defining races.
5. DO HUMAN RACES EXIST: THE ANSWER
Using culture-free, objective definitions of race, the answer to the question whether races exist in humans is clear and unambiguous: no. Human evolutionary history has been dominated by gene flow and admixture that unifies humanity into a single evolutionary lineage. This finding does not mean that all human populations are genetically identical. Isolation by distance ensures that human populations are genetically differentiated from one another, and local adaptation ensures that some of these differences reflect adaptive evolution to the environmental heterogeneity that our globally distributed species experiences. However, most of our genetic variation exists as differences among individuals, with between-population differences being very small. There are no biological races in humans; indeed, despite our global distribution, we are one of the most genetically homogeneous species on this planet.
FURTHER READING
Gonder, M. K., S. Locatelli, L. Ghobrial, M. W. Mitchell, J. T. Kujawski, F. J. Lankester, C.-B. Stewart, and S. A. Tishkoff. 2011. Evidence from Cameroon reveals differences in the genetic structure and histories of chimpanzee populations. Proceedings of the National Academy of Sciences USA 108: 4766–4771.
Lao, O., P. M. Vallone, M. D. Coble, T. M. Diegoli, M. van Oven, K. J. van der Gaag, J. Pijpe, P. de Knijff, and M. Kayser. 2010. Evaluating self-declared ancestry of U.S. Americans with autosomal, Y-chromosomal and mitochondrial DNA. Human Mutation 31: E1875–E1893.
Long, J. C., and R. A. Kittles. 2009. Human genetic diversity and the nonexistence of biological races. Human Biology 81: 777–798. Formal statistics tests of the hypothesis that human genetic variation is structured in a treelike fashion. They show that the hypothesis of a tree of populations is strongly rejected for humans.
Ramachandran, S., O. Deshpande, C. C. Roseman, N. A. Rosenberg, M. W. Feldman, and L. L. Cavalli-Sforza. 2005. Proceedings of the National Academy of Sciences USA 102: 15942–15947. Support from the relationship of genetic and geographic distance in human populations for a serial founder effect originating in Africa
Relethford, J. H. 2009. Race and global patterns of phenotypic variation. American Journal of Physical Anthropology 139: 16–22. This paper shows that skin color variation and other morphological traits are clinal and are not well described by discrete racial categories.
Rosenberg, N. A., J. K. Pritchard, J. L. Weber, H. M. Cann, K. K. Kidd, L. A. Zhivotovsky, and M. W. Feldman. 2002. Genetic structure of human populations. Science 298: 2381–2385.
Santos, R. V., P. H. Fry, S. Monteiro, M. C. Maio, J. C. Rodrigues, L. Bastos-Rodrigues, and S. D. J. Pena. 2009. Color, race, and genomic ancestry in Brazil: Dialogues between anthropology and genetics. Current Anthropology 50: 787–819.
Templeton, A. R. 2003. Human races in the context of recent human evolution: A molecular genetic perspective. In A. H. Goodman, D. Heath, and M. S. Lindee, eds. Genetic Nature/Culture, 234–257. Berkeley: University of California Press. This paper covers many of the same issues as this chapter, but with older data sets. These older data sets yield the same conclusions found in this chapter.
Templeton, A. R. 2005. Haplotype trees and modern human origins. Yearbook of Physical Anthropology 48: 33–59. The results of multilocus nested clade phylogeographic analysis of humans, showing that gene flow and admixture have been such common features of recent human evolution that there is only one evolutionary lineage of humanity.
Wolpoff, M., and R. Caspari. 1997. Race and Human Evolution. New York: Simon & Schuster. The authors strongly argue against typological thinking in anthropology that explains human variation in terms of a few types or “races” rather than dealing with the entire range of variation found in living humans. Although the book ignores the nonhuman literature and tends to hyperbole, it remains an excellent introduction to the intertwined topics of racism and models of modern human origins.