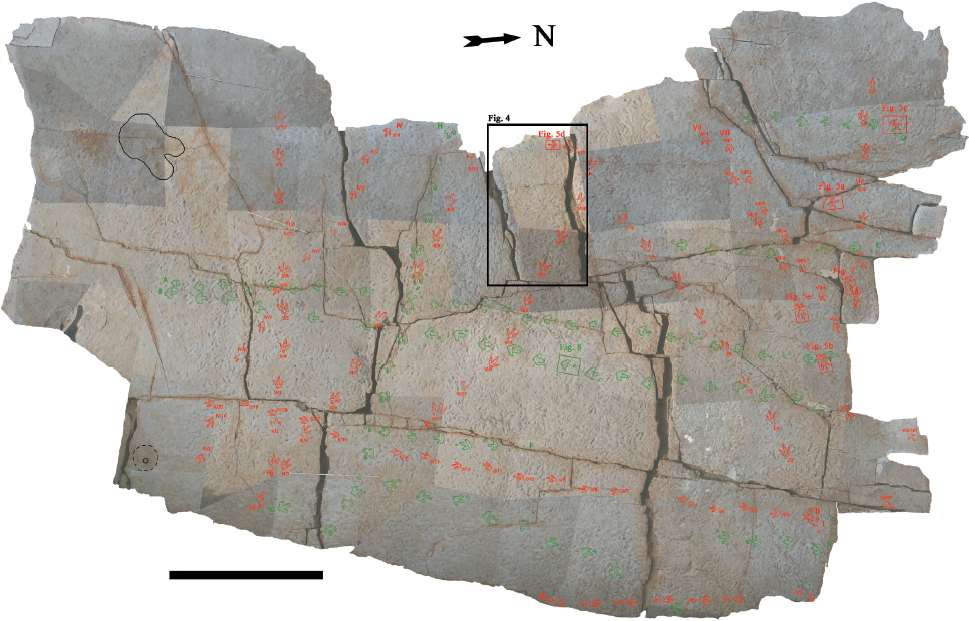
17.1. The Chicken Yard level at night, photograph from a higher layer on the east side; artificial low-angle light from the northern margin. The surface is moderately to heavily dinoturbated. Courtesy Tobias Landmann/Schaumburger Zeitung, 2011.
17.1. The Chicken Yard level at night, photograph from a higher layer on the east side; artificial low-angle light from the northern margin. The surface is moderately to heavily dinoturbated. Courtesy Tobias Landmann/Schaumburger Zeitung, 2011.
Too Many Tracks: Preliminary Description and Interpretation of the Diverse and Heavily Dinoturbated Early Cretaceous “Chicken Yard” Ichnoassemblage (Obernkirchen Tracksite, Northern Germany) |
THE MODERATELY TO HEAVILY DINOTURBATED BERRIAsian Chicken Yard level from the Obernkirchen tracksite (Lower Saxony, northern Germany) is preliminarily described and analyzed. Its ichnoassemblage is characterized by an extraordinary high track density composed of several different morphotypes and size classes of theropod and ornithopod true tracks with an overall similar preservation quality. The occurrence of didactyl tracks of a new, so far unnamed ichnotaxon that can be attributed to deinonychosaurian dinosaurs is particularly remarkable. Despite the high track density and associated frequent overprinting of tracks, several trackways were identified. Their orientation analysis tends toward a primarily bimodal orientation pattern despite the overall chaotic appearance. Also, the history and development of the term “dinoturbation” and its application to Mesozoic dinosaur tracksites are reviewed, and the different factors and scenarios that may have led to high dinoturbation in general and at the Chicken Yard level in particular are discussed together with some recommendations for the analysis of heavily dinoturbated paleosurfaces.
INTRODUCTION
Early Cretaceous (Berriasian) dinosaur tracksites have been known from Lower Saxony, Northwestern Germany, since the 19th century (Struckmann, 1880a, 1880b). Hornung et al. (2012) and Hornung, Böhme, and Reich (2012) have restudied and comprehensively listed 13 localities with terrestrial deposits of Berriasian age in the County of Lower Saxony. Within a terrestrial succession of mainly limnic claystones and black shales, dinosaur tracks have so far only been discovered in sandstones, which are excellent building stones and have therefore been quarried at different locations for the last 1000 years (Graupner, 1977; Lepper, 1997; Lepper and Richter, 2010). Today, only two main areas are still exploited: the Wesling quarry at Münchehagen (Wings, Lallensack, and Mallison, 2016) and – on a larger scale – the Obernkirchener Sandsteinbrüche quarries (Obernkirchen sandstone quarries).
The majority of the historical track discoveries are isolated natural track casts on mostly small quarried slabs (Ballerstedt, 1920). However, the modern quarrying activities are extending horizontally and have uncovered large track-bearing paleosurfaces, where dinosaur tracks and trackways are preserved as impressions or negative epirelief (Wings, Broschinski, and Knötschke, 2005).
Generally, a low to moderate dinoturbation consisting of easily identifiable trackways, with isolated tracks (that cannot be assigned to any given trackway) being the exception, is typical for the Berriasian ichnoassemblages of northwest Germany. Ornithopod and sauropod trackways are often aligned parallel and with regular intertrackway spacing (Fischer, 1998; Wings, Broschinski, and Knötschke, 2005) potentially indicating gregariousness, and preferred walking directions can be determined with only a few exceptions where tracks overprint each other. The majority of the trackways are characterized by short stride lengths and are attributed to herbivores, whereas the theropod trackways generally have longer stride lengths and are scarcer. Often the tracks have a considerable depth and most levels also exhibit ripple-marks and occasionally large fossil tree trunks.
Currently, in the Obernkirchen sandstone quarries (named hereafter Obernkirchen tracksite), two levels, an upper level with mainly iguanodontian and a lower level with a variety of tridactyl (chiefly theropod) tracks and even didactyl tracks, crop out and have been excavated. The latter was discovered by the authors in 2007 and briefly first mentioned by Richter, Böhme, and van der Lubbe (2009). It is an extraordinary track-bearing level that is moderately to heavily dinoturbated and was named the “Chicken Yard”level (Fig. 17.1).
The term “dinoturbation” was coined by Dodson et al. (1980) and was derived from the term “bioturbation,” which is used for sediment disruption caused by diverse invertebrate burrowing organisms. Bioturbation was introduced by the German geologist Rudolf Richter (1936) for “all kinds of displacement within sediments and soils produced by the activity of organisms and plants” (English translation from Flügel, 2004:185). Bioturbation leads to alteration of bedding and lamination structures and contributes to the homogenization of the substrate (Reineck, 1963). The term is meant and used for horizontal (sediment surface) as well as vertical (cross-sections) dimensions of the substrate, the latter being widely applied to thin section analyses. The primary definition and use of the term “bioturbation” arose from marine geosciences (Flügel, 2004). Interestingly, in most of the sediment cases, the focus lies on the vertical component of the textural or fabric changes.
Droser and Bottjer (1986) additionally developed a semiquantitative “bioturbation index,” using the burrow density of substrates for classifying bioturbation fabrics, which they called “ichnofabric.” They showed that a differentiation of bioturbation types is possible by defining categories between two extremes: 0% bioturbation (no evidence of biogenic mixing of sediments) and 100% (complete biogenic mixing of sediments; Flügel, 2004). Between these two end members, a continuous spectrum can be subdivided, for instance, in 10% increments.
Seen in the context of the primary term, dinoturbation can be seen as a special case of bioturbation. When introduced first by Dodson et al. (1980), it was regarded from two viewpoints: dinoturbation must have had an impact (1) on the sediment and (2) on the remains of small biota (small animals and plants) because, as the comparison to extant African biota showed, cropping and “trampling” of large vertebrates – mainly herbivores – in certain savannah areas keeps the diversity of microvertebrate remains and their tracks and traces low (Dodson et al., 1980). Accordingly, dinoturbation may also obscure or delete previously left tracks, notably those of smaller vertebrates, as well as body fossils of invertebrates (clams, snails), plants, and even vertebrates (Lockley et al., 1986; Lockley, 1993) at Mesozoic tracksites.
At the time the term was coined, the focus was on the strong horizontal impact of animal activities on the substrate surface. Thus, considering the locomotor activities of dinosaurs on a substrate, dinoturbation was applied mainly to two-dimensional bedding planes. Moderate to heavy dinoturbation is known from many sites, but the majority of ichnoassemblages are characterized by a rather low dinoturbation index.
Lockley and Conrad (1989) proposed a useful approach toward characterizing dinoturbated tracksites by estimating the percentage of tracks per surface unit, which they called the “dinoturbation index,” subdivided into three major categories: lightly (0%–33%), moderately (34%–66%), or heavily (67%–100%) trampled, which is comparable to the approach of Droser and Bottjer (1986). Accordingly, the most heavily dinoturbated surfaces would correspond to complete superficial trampling or alteration in a way that most remaining structures (tracks) would be very incomplete and poorly defined, whereas only a few could easily and clearly be identified as dinosaur tracks (Fig. 17.1). Typically, such surfaces are characterized by a very high number of isolated and incomplete, overprinted tracks when compared to the total number of recognizable trackways, which is then typically low because it is so difficult to unambiguously identify single trackways.
As mentioned, a low to moderate dinoturbation is typical for German Berriasian tracksites. The Obernkirchen lower Chicken Yard level strongly differs from all of them (including the Obernkirchen upper level) because of its extraordinarily high track density, being the first site from the “German Wealden” that is heavily dinoturbated. On the excavated area (approximately 400 m2) more than 900 rather complete tracks can be counted. Comparable tracksites such as the Australian Lark Quarry site (Thulborn and Wade, 1984; Romilio, Tucker, and Salisbury, 2013) among others are significant witnesses of extraordinarily high levels of dinosaur activity within the fossil record, and possibly of a longer recording timespan.
At the time of writing, negotiations about the protection of the Chicken Yard level were still ongoing, whereas the upper track-bearing level was already protected and open to the public. A first brief description of the Chicken Yard level and the discussion about the origin of its high dinoturbation index including the importance of directional analysis is the focus of this work.
GEOLOGICAL SETTING
The Obernkirchen tracksite is located within the Obernkirchen Sandstone, a thin subunit of the Bückeberg Formation (informally described as the German Wealden) exposed in the northern and western vicinities of Hannover (Hornung et al., 2012). The absolute age of the Obernkirchen Sandstone is estimated between 142 and 138 Ma, spanning ~3.4 Ma (Hornung and Böhme, 2012).
The Lower Saxony Basin, where the Bückeberg Formation was deposited, was a southern sub-basin of the North German Basin. The prograding deltaic sediments from one of two major river systems indicate the deposition of a barrier (p. 394) and delta system (p. 397) to the West of Hannover (Pelzer, 1998; Hornung and Böhme, 2012), which is believed to have matured the sand grains of the Obernkirchen sandstones. It is likely that the formation of the barrier facies was related to landward transport of sand during storm events, and maybe even longshore currents (Pelzer, 1998). This is also confirmed by the occasional occurrence of large tree trunks in the Obernkirchen Sandstone at the Münchehagen tracksite (back-barrier storm deposits sensu Hornung et al., 2012) and in the Obernkirchen Sandstone from Obernkirchen/Bückeberg (landward deltaic succession sensu Hornung et al., 2012; Richter and Knötschke, pers. obs., August, 2007; Richter et al., 2012). In the Early Cretaceous (during the Berriasian and earliest Valanginian), the Bückeberg Formation was deposited at a paleolatitude of about 32°–33° N (Hornung and Böhme, 2012) under warm and humid subtropical to paratropical conditions with seasonal droughts (Pelzer and Wilde, 1987; Hornung and Böhme, 2012).
MATERIALS AND METHODS
The documentation of the excavated Chicken Yard level was achieved after cleaning the surface and taking off the sedimentary infillings of the majority of tracks. In 2008, high-resolution digital photographs were taken from approximately 3–4 m above the ground in cooperation with the Leibniz Universität Hannover and by using a Canon EOS 400 camera. The photogrammetry software Eos Systems PhotoModeler Pro 5.23 was used to produce (with assistance of manually referenced tie-points) a geometric transformation of 95 photographs into one reference system in order to generate an orthophoto (mosaic of photographs without distortion) as a basis for the final track map. To ensure recognition of the tracks on the orthophoto, the tracks were outlined with chalk before the photographs were taken.
Finally, a classical track outline site map was drawn on the basis of the orthophoto using Adobe Photoshop Elements 7.0, outlining the most complete tracks – approximately 900 and much the same count of partially preserved tracks (~700, only partially documented in the map). Then, 715 of the most complete tracks were selected for the total track directional analyses and are represented in fine-scaled (1°) and linear rose diagrams, constructed with Microsoft Excel and modified with Adobe Photoshop Elements 7.0. Within the 715 analyzed tracks, tridactyl tracks of theropods (424, isolated and within-trackway tracks), medium-sized and large iguanodontid ornithopods (196, isolated tracks and tracks of longest trackways), indeterminate medium and large tridactyl tracks (20), deinonychosaurian tracks (55, only tracks from trackways), and small ?ornithopod tracks (20, only tracks from trackways) were considered.
Four simplified rose diagrams show the directions of the most pronounced trackways of tridactyl theropods, medium-sized and large iguanodontid ornithopods, deinonychosaurid dinosaurs, and small ?ornithopods.
Due to the high track density, many of the partially preserved, heavily overprinted tracks and those represented only by weak toe tip impressions were not considered on the main site map and in the count, but they are given as a separate exemplified area containing all textural characters of the surface in detail.
The best-preserved “elite” tracks and trackways of tridactyl theropods and iguanodontid dinosaurs, with all three toes recognizable and in the best case the metatarsal “heel” region, were selected for measurements. Overprinting, however, leads to lacking parameters even for some of the best-preserved tracks. The following track and trackway parameters were measured in the field using standard rulers, calipers (metric units) and goniometers (e.g., following Romilio and Salisbury, 2011; Hübner, 2016): maximum track length (from central posterior border of the metatarsal imprint and the outmost distal tip of digit III), maximum track width (from outmost left and right track margins), digits I–II–III lengths (from central posterior border of the metatarsal imprint and the outmost distal tip of the digit), depth (from track floor of the anterior-most phalange of digit III to an iron bar laid out across the track), digit divarication angles (between the central posterior border of the metatarsal imprint and the outmost distal tip of each digit), pace, stride (both from corresponding points, usually from and to outmost distal tip of digit III or from and to central posterior border of the metatarsal imprint). Tracks of didactyl theropods and small ?ornithopods were not measured in detail, as these tracks are in the focus of Lockley et al. (2016) and Hübner (2016).
Outline drawings of trackways (all of tridactyl theropods and the longest ones from the medium-sized and large iguanodontid dinosaurs) and some individual elite tracks are superimposed onto the photogrammetric map. The trackways are numbered consecutively (roman numbers: tridactyl theropods; letters: iguanodontid ornithopods) including missing tracks in gaps.
Indeed, it proved problematic to number the tracks at all – the sheer mass made it very difficult to put a logical order into the number rows. The decision was made to use the natural weathering of the sandstone into large slabs (German “Blöcke,” B) for numbering each of them (B1, B2, etc.) and then label the tracks within each block in a clockwise direction, starting at “North/0°/360°,” therefore easily being able to redetect them, resulting in numbers such as “B3/27”–block 3, track 27. These numbers will be published in a forthcoming, more detailed publication about the Chicken Yard level and are used here only for three isolated examples not belonging to trackways (“B2e/24,” B1a/14,” and “B8/2”), located on three different sandstone slabs.
The Eos Systems PhotoModeler Scanner computed three-dimensinoal contour line graphics for selected tracks, each calculated out of five photographs taken from different positions with a Canon EOS 550D camera. The graphics were revised with Corel Draw X5 and Adobe Photoshop Elements 7.0. For the work with PhotoModeler, calibrating the cameras and lenses used is obligatory.
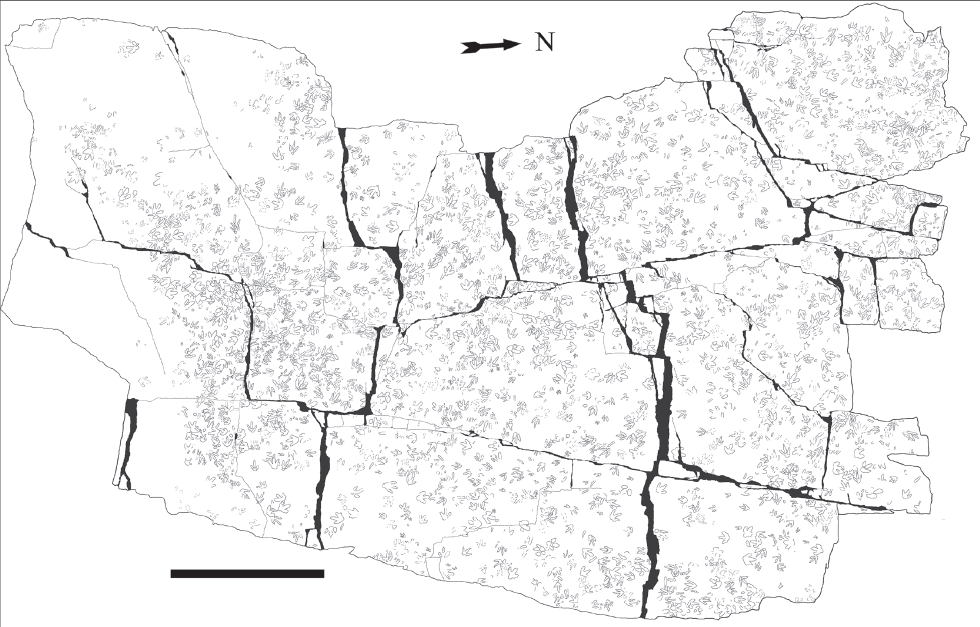
17.2. Sitemap with outline drawings of selected complete and well-preserved dinosaur tracks at the Chicken Yard level, Obernkirchen tracksite, based on the photogrammetric orthophoto. Scale bar: 5 m. Courtesy Schaumburger Landschaft, 2011.
The size classes for tracks (based on pes length [PL]) follow those cited in Marty, Strasser, and Meyer (2009): minute: PL < 10 cm; small: 10 < PL < 20 cm; medium-sized: 20 < PL < 30 cm; large: 30 < PL < 50 cm. Huge tracks (PL > 50 cm) were not found on the Chicken Yard level.
The dinoturbation index scheme developed by Lockley and Conrad (1989) was applied for the Chicken Yard level.
INTRODUCING THE CHICKEN YARD LEVEL – PRELIMINARY DESCRIPTION AND INTERPRETATION
At present, the Chicken Yard level is approximately 400 m2 in size, and it is located on top of a very thick, homolithic and homogenous, fine-grained quartzose sandstone bank of a thickness of approximately 1.6 m. Most of its surface is covered with tracks (Figs. 17.1, 17.2).
Viewed from above, the edge of the overhanging sandstone cliff, which is approximately 6.5 m high, the hundreds of tridactyl tracks look like bird tracks, almost like Gallus imprints in a chicken yard. For this reason, one of the museum volunteers coined the nickname Chicken Yard (German: Hühnerhof) and from then on this name has been used for this level.
In the southwestern corner of the Chicken Yard level and along its southern edge, the surface is quite smooth and lacks any tracks, but some neomiodontid clams are preserved here. Neither ripple-marks nor desiccation cracks are preserved on the Chicken Yard level. Some few plant remains (stems and chaff) are scattered on the surface of the Chicken Yard.
TRACK PRESERVATION
Description
The majority of tracks from the Chicken Yard level show a similar quality of preservation. The preservation quality of the tracks is generally high, as most of the tracks preserve claw impressions and many of them even delicate and clear digital pad impressions. At low-angle light conditions, a marked fine-scale alteration of the surface can be observed.
Most of the tracks from the whole area originally had sedimentary infillings. The natural track casts had approximately the same grain size (slightly finer) than the track-bearing sandstone itself. After removal of the track infills, some track floors additionally showed tiny adhesion spikes sensu Jackson, Whyte, and Romano (2009); they will be described in detail elsewhere (Richter et al., in prep.). No mudcracks can be observed on the Chicken Yard level. Some small areas were slightly damaged by the machines of the quarry workers rolling on the surface, originating in artificial “textures.”
No evidence for the former presence of microbial mats, such as calcareous “biofilm remains,” microbial wrinkle marks (e.g., Dai et al., 2015), or microbial lamination (e.g., Marty, 2008), has so far been found on the Chicken Yard level and underlying and overlying sandstones.
Interpretation
As Hornung et al. (2012) pointed out, it is important to note that the vertebrate (dinosaur) track-bearing deposits of the Obernkirchen Sandstone are all located within a limnic depositional system, in contrast to former publications (e.g., Diedrich, 2004) that have suggested marine influences. Within a cyclic, limnic, depositional environment, the preservation of tracks is common (Souza Carvalho, Borghi, and Leonardi, 2013).
The complete and well-preserved tracks of the Chicken Yard preserve substantial anatomical details. Even if the Chicken Yard tracks do not preserve skin impressions, the fine anatomical details such as digital pad and claw impressions are diagnostic for true tracks, which have not suffered from significant erosion and/or diagenetic modification. Because of the numerous track interferences where complete tracks systematically overprint incomplete tracks and the abundance of distinctive sedimentary infillings even within incomplete tracks, the possibility that incomplete tracks are undertracks can be ruled out.
The sedimentary infillings are comparable to extant tracks and their infillings from Lake Manyara, especially in the vicinity of the lakeshore and the shallow water zone (Cohen et al., 1991). This corresponds well with the observed adhesion spikes and the absence of mudcracks. More or less synchronous sedimentary track infillings originate right after the foot impression when the substrate is still moist to a certain degree (up to saturated; Jackson, Whyte, and Romano, 2009). They are sometimes called ‘Plomben’ in historical German track literature around 1900. Literally plugs, they are sometimes incorrectly associated with the ‘infillings’ of layers above the track horizons, originating after the track-making time, building up the next, completely new layer.
Table 17.1. Some representative examples, with focus on track data from continuous trackways; depth from digit III (theropods: distal phalange)
Trackways (number of datasets) |
Depth ranges distal phalange III (cm) |
Average depth (cm) |
I (8) |
1.8–4.3 |
2.7 |
II (17) |
0.8–2.2 |
1.4 |
VII (7) |
1.3–2.4 |
2.0 |
B (6) |
2.3–3.5 |
2.9 |
Notes: Roman numbers indicate theropod trackways. B is the ornithopod trackmaker. For the position of trackways, see Figure 17.11.
Although there is no evidence for their presence, it cannot be excluded that microbial mats may have played a role during track formation and preservation on the Chicken Yard level, as well as those of other German Berriasian localities with less homogeneous sandstones than at the Chicken Yard level.
TRACK DEPTH
Description
All tracks from the Chicken Yard level are rather shallow (on average ranging in depth between 1.2 to 2.5 cm) (Table 17.1). There is a slight size-independent trend to deeper tracks in the outermost northwestern quadrant of the level (2.5 to 3.8 cm).
No overprinting of deeper tracks by shallower tracks occurs. The majority of the dinosaur tracks have small or no displacement rims, and only in those areas with intensive overprinting, can slight displacement rims be observed.
The track floors are rather even (anteroposteriorly as well as transversal) and show only very weakly (maximum 0.5 to 0.6 cm, mostly not at all) a “classical” deeper anterior part as interpreted as result of the third phase of walking ([1] heel-down, [2] forward rotation, and [3] toe-off, Thulborn and Wade, 1989), and as exemplified by the template experiments of Manning (2004).
Interpretation
Having such flat track floors can be interpreted in different ways: (1) a slippery surface might necessitate a deliberate flat placement of the dinosaur feet (at a high angle, approximately 90°) to prevent instability (Manning, 2004), or (2) a (semi) consolidated, firm underground will not react with a high degree of plasticity (Falkingham et al., 2011; Falkingham, 2014).
As all preserved tracks of the Chicken Yard level are comparably shallow with only a few exceptions, this can be interpreted as a proof of similar to identical substrate conditions all over the level (Lockley, 1993) except in the southern part, where no tracks are preserved at all.
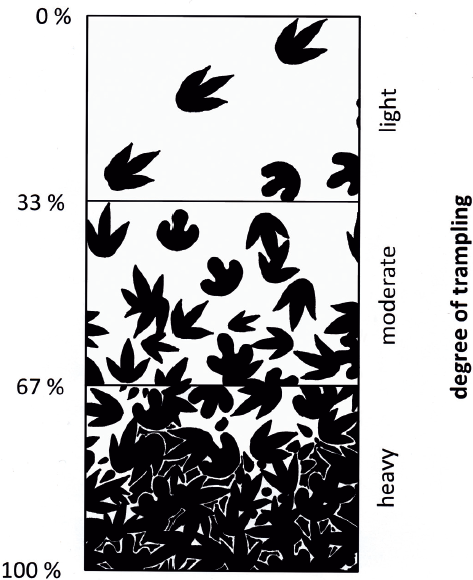
17.3. Dinoturbation index scheme; the track density and associated degree of overprinting are estimated. Drawing after Lockley and Conrad (1989), with schematic tridactyl tracks as observed on the Chicken Yard level.
Even if substrate conditions were possibly more or less the same with a semiconsolidated firm underground, it should also be kept in mind that the foot impact of heavy animals leads to an additional densification of the substrate (Manning, 2004), which is of importance for areas heavily walked upon. From one walking event to the next, the underground must have become increasingly more dense and firm. Therefore, the dinosaurs must altogether have been “paving” their way through the river delta sediments. The last dinosaurs to step on that layer probably found the best and comparably most solid substrate conditions to walk on, leaving shallow tracks and involving less locomotion energy compared to wet, loose sand (Bates et al., 2013).
The tracks are neither deep enough, nor are they found in channels, to discuss avulsion (“dinovulsion”) phenomena such as those known from a Jurassic locality from Utah (Jones and Gustason, 2006). Moreover, computational weathering experiments of Henderson (2006) suggest that it is most unlikely to consider very flat tracks from homolithic rocks to be undertracks. Additionally, the flatter tracks never overprint the deeper ones on the Chicken Yard level, indicating no complete drying out (at least in the preserved “track recording window”).
DINOTURBATION INDEX
Description
The dinoturbation index of the Chicken Yard level is moderate (southern third) to heavy (two northern thirds and middle part of southern area) according to the scheme (Fig. 17.3) developed by Lockley and Conrad (1989). In the southernmost parts, there are no tracks, which represents the “lightly” dinoturbated situation.
A high dinoturbation index and associated degree of overprinting characterizes most areas of the Chicken Yard level, and a small, representative area was chosen to depict the overall density of complete and incomplete (overprinted) tracks as well as impressions that cannot readily be identified as tracks (Fig. 17.4). Roughly estimated, one-third are well-preserved and complete tracks and the remainder are overprinted, incomplete tracks.
Interpretation
An exact count of the incomplete (partially overprinted) tracks was not made, but this seems to be an approach worthwhile for the future work in order to estimate a degree of overprinting. Unless this will be done, the overprinted tracks can be estimated as abundant and with high numbers. Altogether, taking the variable dinoturbation indexes into account, there seems to have been a strong lateral change of either substrate/paleoenvironment or animal behavior.
GROSS TRACK MORPHOTYPES
Due to the overall similar track preservation on the Chicken Yard level, several different morphotypes of theropod and ornithopod tracks can clearly be discriminated. The majority of the tracks are medium-sized to large and primarily tridactyl. There are no tracks from obligatory quadrupedals such as ankylosaurs (in spite of the rediscovery of the Metatetrapodus-type tracks by Hornung, König, and Reich (2012) in the Göttingen University collections) or of sauropods, as observed at the Münchehagen tracksite (Fischer, 1998:plate 2). The different morphotypes are briefly described hereafter.
Tridactyl Theropod Tracks
Description Many of these tridactyl tracks show the typical toe pads and some also show claw marks. Different morphotypes can be differentiated, not all of them forming trackways though. Future multivariate analysis (such as in Castanera et al., 2015) will help to show whether there is just preservational variation or taxonomic reason for the differences, and they will be described in detail elsewhere (Richter et al., in prep.). Hereafter, we give descriptions for some representative morphotypes.
Morphotype 1: Mostly represented by single tracks, characterized by a slender appearance of the digit impressions, marked but not broad digital pads, and a digit IV/III length ratio of 0.80. The length of the track figured here is 36.5 cm. (See Fig. 17.5A.)
Morphotype 2. Not only represented by single tracks, but also by one rather long trackway (no. VII) with 7 tracks. They show extremely marked, broad, and very round digital pad impressions. Thus, the trackmaker must have had rather fleshy feet. The digit IV/III length ratio of VII/6 is 0.83, of VII/5 is 0.84. Mean length is approximately 35.5 cm. (See Figs. 17.5B and 17.5E.)
Morphotype 3. Generally not found in trackways, this morphotype can be attributed to a theropod in a heavily dinoturbated zone (northwestern part of the Chicken Yard) and represents the type 3 here. There are only a few other isolated tracks of this morphotype. This is the largest and most robust morphotype. The mean digit IV/III length ratio is 0.80. Mean length is approximately 44.5 cm. (See Fig. 17.5C.)
Morphotype 4. A more slender appearing theropod morphotype with narrow digit impressions, and a digit IV/III length ratio of 0.75, indicating a comparatively longer digit III. Mean length is 30 cm. (See Fig. 17.5D.)
Interpretation For the medium-sized and some of the large theropod tracks, slightly younger theropod dinosaurs from the Isle of Wight (Wessex Formation, Valanginian and Hauterivian), such as Neovenator or Baryonyx and even the more gracile Eotyrannus might be considered as possible trackmakers insofar as both localities are geographically and temporally relatively close to each other.
The enigmatic Bueckeburgichnus from the German Berriasian (ichnotaxonomy is discussed in Hornung, Böhme, and Reich, 2012) shows hallux impressions, which seem unlikely to be preserved at all within the depositional conditions of the Chicken Yard level with its comparably shallow tracks. However, the gross overall morphology of all tracks of the basic theropod morphotype resembles Megalosauripus rather than Bueckeburgichnus, as the latter is characterized by more stout digits II and III and a less robust digit IV. A more detailed ichnotaxonomic discussion for typical Early Cretaceous medium- to large-sized theropods of Germany will be the focus of a forthcoming work about the Obernkirchen theropods (Richter et al., in prep.).
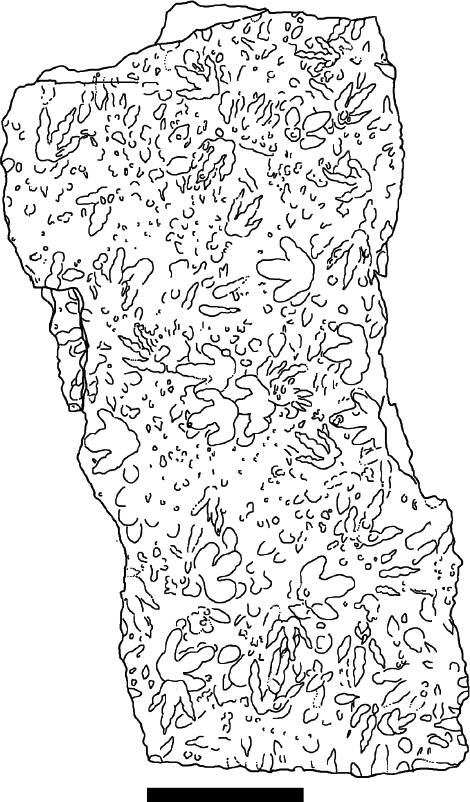
17.4. Detail of a heavily dinoturbated area from the northwestern margin of the Chicken Yard level (to identify on the trackway map, see marked zone in Fig. 17.11). The dinoturbation index can be estimated at >80%. Scale bar: 1 m.
Didactyl Theropod Tracks
Description On the Chicken Yard level, the most remarkable discovery was didactyl tracks (Fig. 17.6) left by deinonychosaurian dinosaurs (van der Lubbe, Richter, and Böhme, 2009). This striking and completely new track morphotype is characterized by the lack of a proper digit II impression, with only a small but distinctive rounded groove around the area of the toe articulation of digit II (see van der Lubbe, Richter, and Böhme, 2009; Lockley, et al., 2016). The mean pes lengths of the didactyl tracks from the seven trackways range between 13 and 24.5 cm, with a mean length of 20 cm.
Interpretation The good preservation quality and quantity (systematic occurrence in seven trackways each with more than three tracks) of these didactyl tracks exclude the possibility that they are all preservational artifacts of tracks left by a trackmaker that usually leaves tridactyl tracks or are due to a paleopathology on both feet of a habitually tridactyl trackmaker. The first scenario was observed in walking experiments with emus by Milàn (2006): emus tended to lower the load on digit II on firm substrates, thus producing didactyl tracks. This can also be observed on recent tidal flats with extant sea gulls when the typical stiff, hard state of sand appears during the approaching phase of the lowering tide (on a very homogenous quartz grain beach), thus allowing producing pseudodidactyl tracks with only very weak traces of the second toes by healthy, normal gull feet (Richter, pers. obs., June, 2010; Falkingham, et al., 2010:fig. 5) (Figure 17.7).

17.5. Four track morphotypes of tridactyl theropods from the Chicken Yard level. (A, B) Tracks are shown as (a) contour line models, (b) photogrammetric photographs, and (c) grazing light photographs. (C, D) Tracks are shown as (a) contour line models, (b) basic photogrammetric photographs, and (c) additional daylight photographs. (E) Track is shown as (a) a contour line model and (b) a photogrammetric photograph. (A) Morphotype 1, medium-sized “standard type” (nicknamed “Mr. Jimmy” during the Obernkirchen Symposium 2011; dextral [dex.]; B2e/24). (B) Morphotype 2: digital pad type (trackway VII/6, sinistral]). (C) Morphotype 3, large and robust theropod track (dex.; B1a/14). (D) Morphotype 4, gracile theropod (dex.; slight overprinting of digit III by another track; B8/2). (E) Morphotype 2: second specimen from the digital pad type (trackway VII/5, dex.). All tracks pictured are localized on the trackway map, Figure 17.11; all black scale bars 10 cm; all line distances 0.1 cm. Grazing light photographs are courtesy of Oliver Gerke, 2010.
In the case of the Chicken Yard level, this scenario can be excluded, as several long trackways cross the entire area. Also, these didactyl tracks always and unambiguously exhibit a distinct, circular impression below the area of toe II (van der Lubbe, Richter, and Böhme, 2009; Lockley, et al., 2016). The didactyl tracks have been assigned to the Troodontidae by van der Lubbe, Richter, and Böhme (2009) and not to the more frequent Dromaeosauridae (see also Lockley, et al., 2016).
Although the absolute lengths of the tracks assigned to Troodontidae are much shorter than those from other theropods, their trackmakers must have been quite large by standards of their family (2–3 m body length). The tracks provide evidence for the existence of these sickle-clawed dinosaurs in the Berriasian of northwest Germany, but skeletal remains are still unknown. For didactyl dromaeosaurs, in the Early Cretaceous of Central Europe, there is only one skeletal tax-on known from velociraptorine teeth from the Isle of Wight (Sweetman, 2004).
It is worth mentioning that one right track and one left track of a troodontid are located almost directly beside each other, evoking the picture of a “standing” animal. Their heel (= metatarsal pad) impressions are longer than those from all other didactyl tracks. They were shortly mentioned in van der Lubbe, Richter, and Böhme (2009), and tridactyl tracks with similar, elongated “metatarsal impressions” are generally interpreted as “crouching traces” (Gierlinski, 1994; Milàn, Loope, and Bromley, 2008; Milner, et al., 2009).
17.6. Grazing light photograph of a troodontid, didactyl, pes track (dextral) from the Chicken Yard level. Joint print at toe II marked by white arrow. Localized on the trackway map, Figure 17.11. Scale bar: 10 cm. Courtesy Oliver Gerke, 2010.
Medium-Sized to Large Ornithopod Tracks
Description These are medium-sized to large tridactyl pes tracks (Fig. 17.8) with petaloid-shaped and rounded toes (hardly recognizable ungual zone) and a small- to mid-sized “metatarsal heel” impression. The pes lengths tend to be slightly longer than the widths. Trackway B (B13–B18), for example, shows a mean length of 40.5 cm and a mean width of 36.2 cm. Mean pace length is 100.7 cm and mean stride length about 203.3 cm. The pes tracks are commonly associated with oval- to kidney-shaped manus tracks, located slightly in front of the pes tracks. More tracks from the Chicken Yard level still have to be measured and additionally investigated with multivariate analysis (compare Hornung, et al., 2016).
Interpretation To date, these tracks match most closely to the ichnogenus Iguanodontipus that is commonly assigned to iguanodontid dinosaurs (Díaz-Martínez et al., 2015; Hornung, et al., 2016; former Amblydactylus, see discussion in Lockley, et al., 2014). In the Early Cretaceous, Iguanodontipus tracks are commonly found in Europe (England, Portugal, Spain, Switzerland). However, there is also an ongoing debate to which dinosaurs (Iguanodon, Mantellisaurus, Delapparentia) these tracks are best assigned to (Santos, Callapez, and Rodrigues, 2012).
Maybe, these dinosaurs reacted upon a wet underground. This would be corroborated by the quadrupedal stance of the iguanodontians, proven by the existence of manus imprints, using their forefeet for stabilization (as described by Wright, 1999).
The presence of shallow manus tracks indicates that there was only a limited influence of erosion on the Chicken Yard level, as the usual preservational bias for ankylopollexian manus impressions would greatly hamper their existence when minimal erosion was present or as undertracks on lower levels (Castanera et al., 2013).
17.7. A trackway of the extant seagull Larus argentatus. A healthy-looking, tridactyl trackmaker was walking on the tidal flat at the time of approaching low tide, producing a pseudodidactyl trackway. Island of Langeoog, German North Sea Coast, 2010. Scale: 1-Euro coin = 2.3 cm.
17.8. A pes and manus track of the Iguanodontipus-type trackway B (localized on the trackway map, Fig. 17.11; sinistral). (A) Contour line model. (B) Basic photogrammetric photograph. (C) Field photograph during dusk. Scale bar: 10 cm; distance between neighboring contour lines is 0.1 cm.
Small ?Ornithopod Tracks
Description There are a few small ?ornithopod tracks on the Chicken Yard level, but they are difficult to spot within the wealth of the larger tracks. There are short trackways with only a few consecutive tracks. The maximum length of the tracks is 16.2 cm and the maximum width is 13.5 cm. Mean length and width according to measurements of Hübner (2016) is 13.1 cm and 10.9 cm, respectively (see chap. 16 for a detailed description of the 28 tracks).
Interpretation Hübner (2016) shows that it is very difficult to assign these tracks even tentatively to either heterodontosaurids, basal ornithopods, dryosaurid ornithopods, and/or basal marginocephalians. Known skeletal genera from the slightly younger Isle of Wight are Iguanodon, Valdosaurus, and/or Hypsilophodon (Martill and Naish, 2001), and the enigmatic Stenopelix from the same horizons of the Obernkirchen area is currently interpreted as basal Marginocephalian or even a very basal Ceratopsian (Butler et al., 2011).
TRACK SIZE CLASSES
Description
Track size classes from the Chicken Yard level include a few small (10 to 20 cm), many medium-sized (20 to 30 cm), and some larger (30 to 50 cm) tracks. Regarding the 715 most complete tracks, they consist of 67.0% theropods (including 7.7% troodontids), 27.4% medium-sized and large iguanodontid ornithopods, 2.8% indeterminate taxa (of medium-sized and large tridactyl dinosaurs), and 2.8% small ?ornithopods.
Interpretation
On the Chicken Yard level, the track sizes are rather widely scaled. In general, track surfaces are controlled by the substrate properties (Falkingham et al., 2011), and they are rather small-scaled. However, size-class exceptions in spite of substrate-dependences are known for example from a carbonate platform tracksite, the Late Jurassic Chevenez-Combe Ronde tracksite from northwest Switzerland, with many small- and medium-sized sauropod tracks that are associated with minute and small tridactyl tracks (Marty, 2008), and this seems to be far more common in the vertebrate ichnological record so that it could be considered an exception (Marty, pers. comm., February, 2015). For widely scaled track sizes, a firm underground underlying the moist upper layer may be a possible explanation.
TRACK AND TRACKWAY ORIENTATION
At first glance, the tracks on the Chicken Yard level look as if they were randomly placed and multimodal, resulting in a chaotic pattern. However, a total directional analysis of 715 clearly distinguishable and plus/minus complete individual tracks delivers a different picture (Fig. 17.9). The majority of tracks point to the north, and a second maximum to the south with a slightly southwestward trend, resulting in an overall bimodal pattern. Less pronounced but still recognizable is another, bimodal east–west trend and some more separate, individual walking directions.
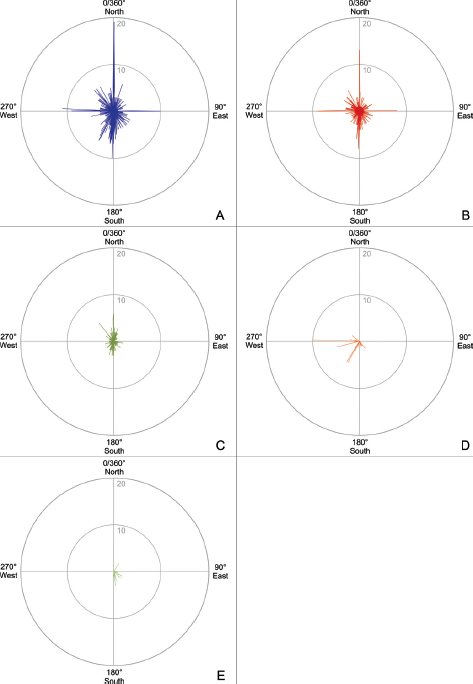
17.9. (A–E) Rose diagrams with track orientation (including tracks of trackways) on the Chicken Yard level; fine-scaled view in 1°-steps, linear scale. (A) Rose diagram for 715 clearly distinguishable and complete tracks (theropods, medium-sized and large iguanodontid ornithopods, few indeterminate medium-sized and large tridactyl dinosaurs, troodontids, and small ?ornithopods sensu Hübner, 2016). The marked north–south–orientation is dominant, a second east-west trend and much more scattered rest of orientations are visible. (B) Rose diagram for 424 tridactyl theropod tracks. Visible are a primary, bimodal north–south, a secondary east–west, and a tertiary, weak southeast–northwest orientation. (C) Rose diagram for 196 medium-sized and large iguanodontid ornithopod tracks. A well-pronounced, bimodal north–south pattern is visible with an additional northwest trend. (D) Rose diagram for 55 troodontid tracks from the eight trackways with marked west and southwest trend. (E) Rose diagram for 20 small ?ornithopod tracks and their mainly southeastward movements.
The picture becomes more differentiated when splitting up the tracks into tridactyl theropod (424), ornithopod (only medium-sized and large iguanodontid ornithopod, 196), deinonychosaurian dinosaur (55), and small ?ornithopod (20) sensu Hübner (2016) tracks. The iguanodontids were walking mainly north–south in both directions, delivering a strong bimodal pattern. This correlates with the “Upper Level” of the Obernkirchen tracksite walking directions.
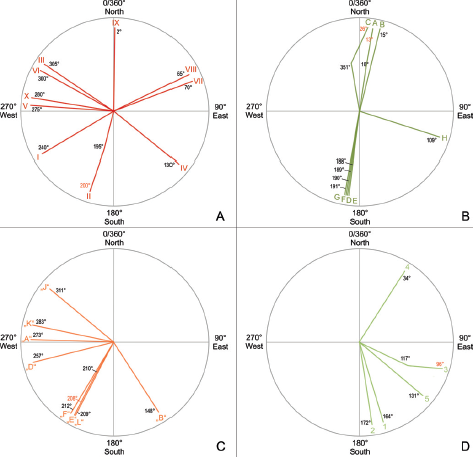
17.10. (A–D) Simplified rose diagrams with trackway directions on the Chicken Yard level. (A) Direction of tridactyl theropod trackways I–X. (B) Directions of medium to large ornithopod trackways A–H. (C) Directions of eight troodontid trackways (following the preliminary naming of van der Lubbe, Richter, and Böhme [2009], A, B, D–F, J–L). (D) Directions of five small ?ornithopod trackways 1–5 sensu Hübner (2016). Changes in direction were depicted stylized. The points of change in direction are the “zero-points” for reading the new direction (red numbers).
The tridactyl theropod tracks show three trends. There seems to be a strong bimodal north–south orientation but also a marked west–east tendency and a third trend toward a west-northwest–east-southeast direction. The tracks of the deinonychosaurian dinosaurs show a marked trend to the west and southwest, those from the small ?ornithopod tracks reflect mainly southeastward movements.
Trackways
Despite the high track density, some trackways of all observed dinosaur groups can clearly be identified on the Chicken Yard level. The 10 tridactyl theropod trackways from the Chicken Yard are represented by straight trackways with only slight turns and iguanodontid ornithopods by 8 almost straight trackways (Fig. 17.10 and 17.11; Table 17.2). Only the longest trackways (minimum five tracks) of the latter category are analyzed here.
Theropod trackways V and X represent the strict “Westerners.” The trackmaker of trackway II went southward but made a slight turn from 195° to 200°, and only the trackway IX points more or less exactly northward.
The longest of them all, trackway A consists of 28 tracks and is located very close and parallel to trackway B with 21 tracks. Both belong to iguanodontian trackmakers which headed toward the north.
The troodontid trackways are subject to another publication (see Lockley et al., 2016, and van der Lubbe, et al., in prep.) but will be also mentioned briefly (Fig. 17.10C). They consist of seven trackways with more than three tracks, or even eight trackways if the shortest one with just three consecutive tracks is also included. The longest one runs 273° (almost exactly west), and there is a second trend toward the southwest (208°–210°, 212°, and 209°). Opposite walking directions (e.g., east or northeast) are not represented. A comparable picture is delivered by the small ?ornithopods (Fig. 17.10D, mostly having moved in southeastern directions (117/96°, 131°, 164°, and 172°), with only one exception to the north-northeast (34°; see also Hübner, 2016). Note that directional changes of some dinosaurs (theropods: II; large ornithopods: A, C; small ?ornithopods: 3; and troodontids: E) have been worked into the same diagrams, respectively, for simplification and recognition that they belong each to one trackway.
Interpretation of Track and Trackway Directions
The marked north–south trend of especially larger tracks and trackways is unambiguous and may indicate a preferred walking direction or pathway. Besides that, each of the dinosaur groups worked out here shows astonishing details that could not be expected a priori by the superficially “chaotic” arrangement of tracks at the beginning of research. In marked contrast to the single tracks, the long theropod trackways do not represent a north–south trend but rather an east–west trend. Due to the completeness of these long trackways, this may imply that these trackways – as a result of a possible “final trackmaking phase” – represent a later change from one preferred walking direction to another over a certain time span. Also, among the larger tridactyl theropod trackmakers, the trackway directions allude that trackmakers VI and III and additionally VIII and VII could possibly have been walking parallel to each other as well as V and X, which, however, are not very close to each other.
A much more strikingly north–south trend is represented by all single large ornithopod tracks and trackways (Figs. 17.9C and 17.10B; Table 17.2) with their north-northeast and south-southwest directions. The mostly parallel arrangement of the iguanodontid ornithopod trackways is also obvious and documents once more the social behavior of the iguanodontian dinosaurs and their roaming in groups. Three trackways (A, B, C) are oriented to the north and four (D–G) to the south, expressing a clear bimodal north–south orientation pattern. Another (H) crosses the main north–south walking direction in an east–southeastern direction. Trackways A and C both make slight turns to the right.
Table 17.2. Directions of trackways for the tridactyl theropods (roman numbers: trackways) and iguanodontid ornithopods (letters A–H); Chicken Yard level
Trackway Number |
Orientation (0°/360° = North) |
I |
240° |
II |
195–200° |
III |
305° |
IV |
130° |
V |
275° |
VI |
300° |
VII |
70° |
XIII |
65° |
IX |
2° |
X |
280° |
A |
10–13° |
B |
15° |
C |
351–26° |
D |
189° |
E |
188° |
F |
190° |
G |
191° |
H |
109° |
Four of five small ?ornithopod trackways show a strong preference to southeastern directions, only one at 34° (north-northeast), and none with a north–south direction.
On the Chicken Yard level, many of the didactyl tracks are organized in trackways, too, and they also do not mirror the north–south trend of some of the other groups but depict a distinct western and southwestern trend. Three (E, F, L) of these are even parallel to each other. Parallel didactyl trackways (ichnotaxon Dromaeopodus) were already reported from the Early Cretaceous of Shandong Province (China) and interpreted as evidence of gregarious behavior (Li et al., 2015). The parallel didactyl trackways from the Chicken Yard level could thus be further evidence that these animals frequently occurred in groups (refer also to van der Lubbe, et al., in prep.).
The different but mostly straight walking directions and the lack of centrally arranged, radial trackways (see “Discussion”) reject the presence of a shrinking water hole scenario as was proposed for instance for the Isle of Wight dinoturbation layers (Martill and Naish, 2001). The presence of straight trackways, some of which are parallel, seem to indicate a “passing-by” without much interaction between individual trackmakers. For the theropod tracks, this also excludes rambling paths such as have been suggested for the Hettangian tridactyl tracks from Dinosaur State Park (Galton and Farlow, 2003). For the iguanodontid ornithopods, the presence of manus tracks, the short pace/stride values and the gentle turns of the trackways indicate a slow walking gait.
17.11. Outline drawings of the 10 tridactyl theropod (I–X) and 8 ornithopod (A–H) trackways of the Chicken Yard level superimposed onto the photogrammetric map. Also, the examples for the gross track morphotypes are marked here (see Gross track types) and the exact position of “slab 8 (B8)” with the realistic textural density from Figure 17.4 is shown. Scale bar: 5 m.
Two groups of iguanodontid trackways on the upper track level of Obernkirchen (the Upper Level approximately 4 m above the Chicken Yard level) show a clear bimodal north–south orientation (Böhme et al., 2009). This corresponds very well with the directions gathered from the tracks of the Chicken Yard level and may be a first hint toward a possible longer-term trend of displacement along a stable landscape marker, such as the shoreline of a large lake.
There may have been two different dinosaur walking events altogether: a rather random primary one and then a second, with a strong primary north–south bimodal pattern and a weaker secondary east–west ‘bimodal’ trend, together forming almost a quadrimodal configuration. The more marked north–south orientation of the iguanodontian trackways could also indicate an additional species-related, different behavior, such as a preferred route.
The Early Cretaceous dinosaur tracks from Glen Rose/Paluxy River (United States) serve as good comparison. Besides the large sauropod tracks, the tridactyl tracks show a mirror-image, bimodal orientation pattern, with most of them heading generally northward or southward, in approximately equal numbers. This bimodal pattern of dinosaur trackways is commonly observed; it suggests that the trackmakers were moving back and forth along the local coastline (Lockley, 1993; Farlow et al., 2011). This could fit well into a small-scale, regional wandering due to the aforementioned nutritional aspects. Dinosaurs wandering alongside a shoreline or a lakeside (or here, a large lake’s delta) could also have done so in order to avoid possible geographical barriers in the hinterland (e.g., dense forests). This interpretation could also fit for the second tracksite of the Obernkirchen sandstone quarries (Upper Level), where iguanodontid dinosaurs walked in groups besides each other in a comparable bimodal pattern (Böhme et al., 2009).
Taking the bimodal trackway pattern into account, it can also be postulated that returning individuals left two trackways. Interpretations such as this have been made and seem logical (Farlow et al., 2011), but bidirectional walking zones could only be proven by unambiguous special cases of individual pathology, such as “a hurt, biped dinosaur” (Lockley, 1993:99) or the tyrannosaurid of McCrea et al. (2014).
17.12. Vertical cross-section of a theropod track from the northern continuation of the Chicken Yard level showing the homolithic texture of the fine-grained sandstone. Only small coal ribbons are recognizable on a macroscopic scale within the sandstone. Scale bar: 10 cm.
Additional Remark
In spite of the excellent and numerous horizontal data, the homolithic nature of the Chicken Yard locality level sandstones (see Fig. 17.12) does not enable the inclusion of data sets concerning the vertical aspects.
DISCUSSION
Fossil localities with a high dinoturbation index and thus high numbers of tracks received iconic nicknames such as “dinosaur disco” (Moutier tracksite, northwest Switzerland: Lockley and Meyer, 2000), “the Raceway” (Fleming Fjord Formation, Jameson Land, East Greenland: Milàn, Clemmensen, and Bonde, 2004), the “Blue Hole Ballroom” (Paluxy River, United States: Farlow et al., 2011), or the controversial “Dinosaur stampede” (Lark Quarry tracksite, Australia: Thulborn and Wade, 1979, 1984; Romilio and Salisbury, 2011). The Chicken Yard level is one of the ichnoassemblages with the highest dinoturbation index worldwide (Fig. 17.13).
Important Parameters for Moderate to High Dinoturbation
The most important factors that may have led to the exceptionally high degree of dinoturbation observed on the Chicken Yard level, sensu Lockley and Conrad (1989), are paleoenvironment, duration of exposure of the sediment surface, moisture content of the substrate at time of track formation, population density, and animal activity and behavior. For the following discussion (following Marty, pers. comm., November, 2014), we propose a slightly simplified parameter list:
1. Paleoenvironment – physical barriers, such as shorelines or water holes, renewed wetting on tidal flats, and so on.
2. Track recording – time of exposure, substrate properties sensu lato, climatic conditions, and so on.
3. Trackmakers – behavioral parameters of animals: frequency, level of activity, overprinting, carrion, feeding grounds, and so on.
17.13. Detail from the Chicken Yard level at night, photograph taken from a higher level on the east side, artificial grazing light from the northern margin (lower right in the photograph), showing the general aspect of a moderately to heavily dinoturbated ichnoassemblage. Courtesy Tobias Landmann/Schaumburger Zeitung, 2011.
Paleoenvironment Behrensmeyer et al. (1992:48) stated, “The margins of large, fluctuating lakes are generally poor in organic remains, although wood and small vertebrate remains may occur; but they provide an optimal context for ichnofossils, including invertebrate and vertebrate tracks, trails, fish nests, and root traces.” As the Obernkirchen Chicken Yard level belonged to a large former lacustrine area, it fits well into this scenario. Driftwood, root traces, and the dinosaur tracks are the trademarks of the Berriasian deposits of Obernkirchen, although some small freshwater-to-brackish clams occur as well (Neomiodontidae; see Richter et al., 2012). For an actualistic comparison, an African lake system chosen by Cohen et al. (1991, 1993) proves ideal. Like the many known fossil track–bearing lake-margin strata, Lake Manyara in Tanzania represents an aggrading, closed depositional system with numerous and diverse trackmakers (birds, mammals). The shores of Lake Manyara show trampled (= “heavily bioturbated”) areas in different distances to the shoreline, so that the aspect of trampling and thus large-scale bioturbation delivered useful observations transferrable into dinoturbation criteria (for more details see Cohen et al., 1991, 1993).
The dinosaurs leaving their tracks on the Chicken Yard level must have crossed a large river delta system at the margin of a large lake some 140 million years ago and thus were probably only temporarily present in a paleoenvironment composed of emerged sand bars and barriers (Hornung et al., 2012; Hornung and Böhme, 2012) when they have passed alongside the southern and southeastern shore of the Lower Saxony Basin. In the hinterland of Obernkirchen, forests were present and probably sustained some of the herbivorous dinosaurs (Hornung et al., 2012). It remains questionable whether there were narrow zones, leading to migration corridors in a true sense, or if a passing-by in different directions just followed the actual paleogeographical situation. According to the paleoenvironmental interpretation of Hornung et al. (2012), a network of fluvial channels developed toward the lagoon and sandy mouthbars divided the many arms, some of which were in the process of being abandoned and reduced to oxbow lakes (see pp. 397, 398). It is likely that dinosaurs passing by had to avoid certain areas of these sandbars or deeper oxbow lakes.
Track Recording An important parameter that influences dinoturbation is time (Lockley and Conrad, 1989). In order to produce a heavily dinoturbated level, a certain time span was required for track formation before the environmental conditions changed, track formation became no longer possible, and the next depositional event covered and preserved the track-bearing surface (e.g., growth of microbial mats, tempestitic influences: see Hornung et al., 2012; and discussions herein).
However, the duration of exposure is not measurable and can greatly vary in different extant depositional systems between some days and up to several weeks, months, and even years (Cohen et al., 1991; Marty, 2008). Moreover, it is possible that due to repeated drying and wetting cycles, tracks may form at several different occasions (track recording windows) and lead to the formation of a heavily dinoturbated level that consists of several different track generations left by unrelated noncontemporaneous trackmakers.
Time periods on the order of days to weeks are often considered as track-formation windows in carbonate depositional environments (e.g., Kvale et al., 2001; Marty, 2008). Cohen et al. (1991) provided interesting “track survivorship” curves showing that in an extant lake system from Africa, for small vertebrates, a maximum of 20 days of general track persistence was valid but only in a very restricted zone. On the other hand, microbial mats that are ubiquitous on modern and ancient carbonate and siliciclastic tidal flats may lithify by the precipitation of calcium carbonate and consequently enhance the preservation potential of tracks and other traces (Marty, Strasser, and Meyer, 2009). Such consolidated mats may be exposed for longer time periods during which they may withstand trampling by large and heavy trackmakers and even resist heavy rainfall (Marty, Strasser, and Meyer, 2009). Scott et al. (2007) noted that microbial mats do temporarily protect tracks due to fluid chemistry influences allowing calcification.
Consequently, ichnoassemblages are commonly affected by a certain degree of time-averaging. Even though, the degree of time-averaging is very low when compared to most other sedimentary depositional units (e.g., a single bed), it is still high enough to make it generally impossible to determine – based on tracks – whether on a given surface two animals were really present at the same time or not (Marty, 2008). To make that determination, other characteristics such as (very long) parallel trackways with a very regular intertrackway spacing, synchronous turns, or very similar track preservation need to be present.
The longer a drying phase (track recording window) is, the more animals can pass through a given zone, and the less that can be said about the faunal diversity, as time-averaging increases (Falkingham et al., 2011). On the other hand, it is probable that additional densifying processes such as those deriving from many animals walking upon a substrate during a certain time span simultaneously stabilizes the substrate as a whole, strengthening it against contemporary erosion, which is known from African flamingo populations and their innate substrate influence (Scott, Renaut, and Owen, 2012).
Studies about extant African land vertebrate faunas from lakes show that within transects from lake shores to landward areas, the depth of the tracks increases with increasing water content, and some “mass track occurrences” such as those from flamingos compare well to fossil dinoturbation areas concerning the density of the tracks and the interactions of tracks with the substrate (Cohen et al., 1991, 1993; Scott et al., 2007) and each other (overprinting).
The different size classes of the tracks on the Chicken Yard level indicate that quite different sizes and types of small, medium-sized, and large dinosaurs were able to walk on the surface, implying a moist thin layer that overlaid a firmer subsurface layer, resulting in a certain stability for different weight groups. The vertical cross-section shows the homolithic sandstone entity (see Fig. 17.12). The low displacement rims of the track additionally hint to this, as seemingly there was not much substance to be squeezed at the margins. Concerning the sizes, a similar situation with huge and small tracks is known from Lark Quarry, Australia (Thulborn and Wade, 1984).
Concerning the time of exposure (parameter 2) and the activity of the dinosaurs (parameter 3), it remains questionable what caused the high number of incomplete and/or overprinted tracks at the Chicken Yard. Random overprinting during a short but high-activity time period can be discussed. The shallow tracks with their flat or lacking displacement rims could point to a “short” drying-out event, the surface buried before drying out completely. The marked similarity and equal preservational quality of all tracks with no shallower tracks overprinting deeper tracks might result from that short event, which necessarily implies a high biological activity at the very end of that phase.
Also, another scenario can be constructed. Several walking cycles result in several track-recording events modified afterward by alternating drying out and rewetting, along with the associated erosion. In that case, only the last “track generation” would not undergo erosion and could become ichnotaxonomically the most useful group of tracks in “elite preservation quality,” including rather complete trackways. If the latter had been the case on the Chicken Yard level, it would explain a longer time span to develop bimodal traveling paths, but the preservational quality of all recognizable tracks should be quite different, assuming that there are recognizable but eroded tracks preserved from theoretical cycle(s) before the last one.
Thus, a third scenario seems more probable for the Chicken Yard. All recognizable tracks show a similar quality of preservation, which speaks against multiple drying-out cycles plus erosion. Also, there are indicators such as the lack of desiccation cracks, sedimentary infillings in the tracks (possibly originating from shallow water), and in certain tracks adhesion spikes, which give additional evidence for a high moisture or even permanent water cover of the uppermost substrate surface and no final drying out at all.
Additionally, supported by the single theropod track north–south–directional trend contrasting with the east-west one for the long theropod trackways, plus many walking directions for the four gross dinosaur track types, this most probably points at a changing environment during the track recording time (e.g., instable and migrating river delta) and thus a “longer” time span. This could have resulted in slight changes of abundantly used travel paths as for the theropods, which changed from north-south to east-west. Altogether this evidence would suggest a “longer-term” exposure (e.g., several weeks as in Farlow et al., 2006, or even more), likewise allowing for many different dinosaurs to pass by in all possible directions without the need to hypothesize an intensified biological activity during a short time. This interpretation is preferred here.
Trackmakers The behavior of animals plays a most crucial role, and even a few animals can leave many tracks if there is a driver such as a water hole or carrion. Martill and Naish (2001) reconstructed a water hole scenario from the terrestrial Valanginian Wessex formation from the Isle of Wight, representing a formerly subtropical environment with seasonal droughts: “As the [fresh] water body became smaller, more dinosaurs would have gathered around the shrinking water hole, churning up the mud, as occurs in modern-day seasonal wetlands such as the South American Pantanal. Eventually, the margin of the water body would have become an intensely churned quagmire” (319). This scenario represents an abundant environmental situation that is not restricted to South America. The shrinking, drying-up water reservoirs are suboptimal preservational zones for light-weight animals (the “Goldilocks effect;” Falkingham, et al., 2011) and also preferred ambush hunting zones for predators and thus, mostly avoided by small animals, which consequently do not leave tracks. But all other, larger animals gather exactly the way Martill and Naish (2001) describe it and leave their tracks. Thus, for terrestrial deposits, this could be a favorable reason to explain dinoturbated areas.
Carrion scenarios can easily be observed today. Dead seals (Phoca vitulina) or harbor porpoises (Phocoena phocoena) regularly attract large groups of seagulls at the North Sea Coast. The latter produce high track densities around the carcasses (in analogy to dinoturbation, this could be named “aviturbation”). Their foot skeletons are anatomically almost identical, and the different genera and species cannot easily be distinguished (Richter, pers. obs., August, 2010) (Fig. 17.14).
Another scenario indicates that many dinosaurs could have been looking for special nourishment conditions, for instance, upon regularly emerged supratidal flats – not carrion but living fish trapped in tidal ponds (Petti et al., 2011) similarly to extant shore birds moving restlessly along the shoreline and along tide pools (Lockley and Meyer, 2000). Thus, they represent trackways of animals feeding in special environments (“foraging behavior”).
For theropod-dominated heavily dinoturbated levels, carrion-feeding could be proposed, but an ichnoassemblage providing evidence for this kind of behavior has not been found so far. However, theropods can be abundant in general without dinoturbation, such as marked majorities of theropods have lately been confirmed again for the important Shandong Tianjialou Formation (?Aptian-Albian) from China (Li et al., 2015).
Judging from seagulls gathering around carrion leaving dense track zones as pictured in Figure 17.14, the majority of the tracks should be arranged radially to the (former) center of the food and just around this area. This scenario can clearly be ruled out for the Chicken Yard ichnoassemblage, because the heavily dinoturbated area has a considerable size and extension. Also, within the chaos of trampling caused by carrion-feeding, trackways should not be expected. The rather distinct trackways from the Chicken Yard clearly speak against such a scenario.
CONCLUDING REMARKS AND OUTLOOK
The Chicken Yard level at the Obernkirchen tracksite is a good example of a moderately to heavily dinoturbated paleosurface and as such delivers the typical advantages and disadvantages of mass track occurrences. The density of tracks is so high that the degree of overprinting exacerbates the general analysis, but on the other hand it exhibits a high ichnodiversity despite the rather small size of this site.
17.14. Dense track zone of extant sea gulls at a carrion feeding place, a dead subadult seal (Phoca vitulina). The gull Chroicocephalus ridibundus was observed feeding together with the Larus-species L. marinus, L. canus, and L. argentatus, and other than minor size differences, their tracks are virtually nondistinguishable. Island of Langeoog, German North Sea Coast, 2010. Below sternal region: 2-Euro coin as scale = 2.5 cm.
Although there is some mixed preservation with many incomplete and/or overprinted tracks (sensu Castanera et al., 2013), the overall track preservation quality is very good and the Chicken Yard level tracks exhibit enough anatomical details to differentiate among several, gross track morphotypes: tridactyl and deinonychosaurian theropods (the latter formerly unknown from the German Berriasian), iguanodontian ornithopods, and small basal ornithopods or basal marginocephalians (compare Fig. 17.15). Even within the tridactyl theropods, different morphotypes could be recognized, though future investigations (such as multivariate analysis) have to show whether there is a taxonomic reason for the observed differences or to what degree these morphotypes may be affected by a preservational influence. As the skeletal record of Early Cretaceous dinosaurs is poor in Germany, the tracks are an important element for reconstructing the local dinosaur fauna.
Although the precise reason for the unique preservation of this densely dinoturbated level with its strong predominance toward theropod tracks remains unknown, at least the “track chaos” definitively has a hidden trend toward changing bimodal orientation patterns of the tracks and trackways. This and the linear trackways without any pronounced turns support a passing-by scenario.
The paleoenvironment of the Chicken Yard level is interpreted as a former delta environment at the fringe of the Lower Saxony Basin. We can imagine that oxbow lakes with calm and shallow water and eventually changing sandbars could have been the ideal place for dinosaurs leaving their tracks (following Hornung et al., 2016, see pp. 397, 398) and also for preserving them (Fig. 17.15). But also some indicators such as the lacking desiccations tracks, the changing trackway directions, and others point toward this interpretation.
Different size classes and the shallow and similar preservation of the tracks fit best to the explanation of a thin moist top layer stabilized by a firm underground, perhaps further consolidated by dinoturbation.
Despite the presence of some obvious trackways, one should bear in mind that a big problem of heavily dinoturbated surfaces will always be the difficulty to unambiguously recognize trackways. Especially in the case of “accelerating, decelerating or turning trackways,” the trackway identification is hampered by the high track density, and only more or less regular trackways can “easily” be detected. Nonetheless, the case of the German Chicken Yard proves that it is indeed rewarding to search carefully for these few trackways and to analyze their orientations. In comparison to the “total directions analysis” with a strong north–south and weaker east–west trend, the trackways prove different behaviors of different dinosaur groups; for instance, the large iguanodontian ornithopod trackways mostly represent the marked north–south trend, whereas the orientation of the larger theropod tracks are much more variable (at least within the trackway data) and even show a change from north–south to the lately produced, last-generation east–west trackways.
17.15. The different dinosaurs of the Chicken Yard level. Non-naturalistic grouping to show the different gross track morphotypes. Artwork by Frederik Spindler, Freiberg, Germany, 2014.
Thus, for heavily dinoturbated surfaces, we recommend a detailed directional analysis for single tracks as well as for trackways. In forthcoming works, the degree of overprinting deserves a closer look and could ideally be transferred into a measurable “degree” and thus a parameter for the assessment of dinoturbated surfaces. This might allow further conclusions toward the behavior of the trackmakers. For instance, around a water hole or carrion or other feeding place, tracks may be arranged very chaotically and result in a high degree of overprinting (not many trackways visible), whereas in an area that is a regular pathway or migration corridor (even if that area is changing from time to time), the last animals that went over the surface will still leave straight trackways that can easily be recognized. The latter case would possibly be the better scenario to explain the Chicken Yard level ichnoassemblage than any “water hole” or “feeding ground” scenario would.
The Chicken Yard sandstone is homolithic and delivered no additional vertical data. Though, if the substrate of a dinoturbation layer is heterolithic and allows for vertical investigations (subsurfaces), this should be done level by level. Details of a cross-sectional analysis must be taken into account, as undertracks may contribute toward an even better understanding of the origin of the tracksite than if it were only based on the true tracks. This way, it will become clearly visible whether the dinoturbation is a true signal from only one paleosurface or whether it is an accumulated (composite signal) from tracks left on different layers with undertracks and/or overtracks, which would lead to different interpretations (see also Marty, 2008).
Hopefully, the ongoing quarrying activities will open up the connection of the current Chicken Yard level to the east in the nearby future. The Chicken Yard level should definitely be protected, which we hope to support by this research.
ACKNOWLEDGMENTS
Our sincerest and primary gratitude goes to Klaus Köster, the owner of the Obernkirchen sandstone quarries, who supported our work so generously in spite of many obstacles. His employees are thanked as well. A big thank-you to all of our good-witted volunteers who have been strong and full of enthusiasm and have never lost faith in the project. Among those, Uwe Stratmann deserves an extra thank-you for his massive support since the beginning of the project in winter 2007/2008 and the discovery of the Upper Level. We acknowledge the permanent background counseling activities of our cherished colleague Dr. Jahn Hornung and some fruitful discussions and exchange with our colleague Dr. Tom Hübner. The Lower Saxony Ministry of Science and Culture generously supported the Obernkirchen digging campaigns 2009, 2010, 2011, and donated even more money for the color illustrations and the overall quality of this “life of the past” book. Dr. Manfred “Sir” Wiggenhagen taught us the primary photogrammetric methodology – his sedulous help deserves a “grand merci!” The rural county of Schaumburg, namely, Fritz Klebe and his collaborators, continuously helped through difficult political times – thank you for that! Also, the erection of the public area Upper Level Obernkirchen/“DinOS” as project from the European Union is their merit. A huge thanks goes to the foundation Schaumburger Landschaft, namely, Sigmund Graf Adelmann, who made the Dinosaur Track Symposium Obernkirchen 2011 possible and who financially supported the making of the main site map by Torsten van der Lubbe, the discoverer of the troodontid tracks, whom we also thank for his support from 2008 until 2011. The foundation Klosterkammer Hannover, namely, Dr. Stephan Lüttich, is thanked for parts of the symposium as well as for supporting the making of scientifically correct, new landscape illustrations for the Lower Saxony Lower Cretaceous. These have been brilliantly done by Frederik Spindler, Freiberg, whom we thank for his enormous skills and his patience. The foundation Stiftung Niedersachsen generously funded the Obernkirchen symposium, is thanked, and in particular, Dr. Joachim Werren. Oliver Gerke is thanked for providing three photographs of the Chicken Yard level at night. We also thank Tobias Landmann, Schaumburger Zeitung, for allowing us to use two of his Chicken Yard level night photographs. Martin G. Lockley, an anonymous reviewer, Peter Falkingham, and especially Daniel Marty deserve our deepest gratitude because their comments substantially enhanced the quality of the manuscript – Daniel, merci beaucoup! Robert Fischer helped with one of the figures. Finally, A.R. wishes to express her gratitude toward her family, who proved to be very patient from the symposium through the finishing of this book.
REFERENCES
Ballerstedt, M. 1920. Dinosaurierfährten im Wealdensandstein des Harrl bei Bückeburg und eine zurzeit freiliegende Spur eines ‘vierfüs-sigen,’ plumpen Dinosauriers. Zeitschrift der Deutschen Geologischen Gesellschaft 72: 8–10.
Bates, K. T., R. Savage, T. C. Pataky, S. A. Morse, E. Webster, P. L. Falkingham, L. Ren, Z. Qian, D. Collins, M. R. Bennett, J. McClymont, and R. H. Crompton. 2013. Does footprint depth correlate with foot motion and pressure? Journal of the Royal Society Interface 10: 20130009, doi: 10.1098/rsif.2013.0009.
Behrensmeyer, A. K., J. D. Damuth, W. A. DiMichele, R. Potts, H.-D. Sues, and S. L. Wing. 1992. Terrestrial Ecosystems through Time: Evolutionary Paleoecology of Terrestrial Plants and Animals. The University of Chicago Press, Chicago, Illinois, 588 pp.
Böhme, A., U. Stratmann, M. Wiggenhagen, A. Richter, and T. van der Lubbe. 2009. New tracks on the rock: parallel trackways of a new type of Iguanodontipus-Caririchnium-like morphology from the Lower Cretaceous sandstones of Obernkirchen, northern Germany. Journal of Vertebrate Paleontology 29(3, Supplement): 66A.
Butler, R. J., L. Jin, J. Chen, and P. Godefroit. 2011. The postcranial osteology and phylogenetic position of the small ornithischian dinosaur Changchunsaurus parvus from the Quantou Formation (Cretaceous: Aptian-Cenomanian) of Jilin Province, northeastern China. Palaeontology 54: 667–683.
Castanera, D., J. Colmenar, V. Sauqué, and J. I. Canudo. 2015. Geometric morphometric analysis applied to theropod tracks from the Lower Cretaceous (Berriasian) of Spain. Palaeontology 58(1): 183–200.
Castanera, D., B. Vila, N. L. Razzolini, P. L. Falkingham, J. I. Canudo, P. L. Manning, and À. Galobart. 2013. Manus track preservation bias as key factor for assessing trackmaker identity and quadrupedalism in basal ornithopods. PLoS One 8(1): e54177.
Cohen, A. S., J. Halfpenny, M. G. Lockley, and E. Michel. 1993. Modern vertebrate tracks from Lake Manyara, Tanzania and their paleobiological implications. Paleobiology 19(4): 433–458.
Cohen, A. S., M. G. Lockley, J. Halfpenny, and A. E. Michel. 1991. Modern vertebrate track taphonomy at Lake Manyara, Tanzania. Palaios 6: 371–389.
Dai, H., L. Xing, D. Marty, J. Zhang, W. S. Persons IV, H. Hu, and F. Wang. 2015. Microbially-induced sedimentary wrinkle structures and possible impact of microbial mats for the enhanced preservation of dinosaur tracks from the Lower Cretaceous Jiaguan Formation near Qijang (Chongqing, China). Cretaceous Research 53: 98–109.
Díaz-Martínez, I., X. Pereda-Suberbiola, F. Pérez-Lorente, and J. I. Canudo. 2015. Ichnotaxonomic review of large ornithopod dinosaur tracks: temporal and geographic implications. PLoS One 10(2): e0115477.
Diedrich, C. 2004. New important iguanodontid and theropod trackways of the tracksite Obernkirchen in the Berriasian of NW Germany and the megatracksite concept of Central Europe. Ichnos 11(3–4): 215–228.
Dodson, P., A. K. Behrensmeyer, R. T. Bakker, and J. McIntosh. 1980. Taphonomy and paleoecology of the dinosaur beds of the Jurassic Morrison formation. Paleobiology 6(2): 208–232.
Droser, M. L., and D. J. Bottjer. 1986. A semiquantitative field classification of ichnofabric. Journal of Sedimentary Petrology 56: 558–559.
Falkingham, P. L. 2014. Interpreting ecology and behaviour from the vertebrate fossil track record. Journal of Zoology 292(4): 222–228.
Falkingham, P. L., K. T. Bates, L. Margetts, and P. L. Manning. 2011. The ‘Goldilocks’ effect: preservation bias in vertebrate track assemblages. Journal of the Royal Society Interface 8(61): 1142–1154.
Falkingham, P. L., L. D. Agenbroad, K. Thompson, and P. L. Manning. 2010. Bird tracks at the Hot Springs Mammoth Site, South Dakota, U.S.A. Ichnos 17: 34–39.
Farlow, J. O., W. Langston Jr., E. E. Deschner, R. Solis, W. Ward, B. L. Kirkland, S. Hovorka, T. L. Reece, and J. Whitcraft. 2006. Texas giants: dinosaurs of the Heritage Museum of the Texas Hill Country. The Heritage Museum of the of the Texas Hill Country, Canyon Lake, Texas, 105 pp.
Farlow, J. O., M. O’Brien, G. J. Kuban, B. F. Dattilo, K. T. Bates, P. L. Falkingham, L. Piñuela, A. Rose, C. Freels, C. Kumagai, C. Libben, J. Smith, and J. Whitcraft. 2011. Dinosaur tracksites of the Paluxy River Valley (Glen Rose Formation, Lower Cretaceous), Dinosaur Valley State Park, Somervell County, Texas; pp. 41–69 in P. Huerta, F. T. Fernández-Baldo, and J. I. Canudo Sanagustín (eds.), Proceedings of the V International Symposium about Dinosaur Palaeontology and Their Environment. Colectivo Arquelógico y Paleontológico Salense, Burgos, Spain.
Fischer, R. 1998. Die Saurierfährten im Naturdenkmal ‘Saurierfährten Münchehagen’; pp. 3–60 in R. Fischer (ed.), Das Naturdenkmal ‘Saurierfährten Münchehagen.’ Mitteilungen des geologischen Instituts der Universität Hannover 37. Universität Hannover, Hannover, Germany.
Flügel, E. 2004. Microfacies of Carbonate Rocks: Analysis, Interpretation and Application. Springer, Berlin, Germany, 976 pp.
Galton, P. M., and J. O. Farlow. 2003. Dinosaur State Park, Connecticut, USA: history, footprints, trackways, exhibits. Zubia 21: 129–13.
Gierlinski, G. 1994. Early Jurassic theropod tracks with metatarsal impressions. Przeglad Geologiczny 42: 280–284.
Graupner, A. 1977. Natursteine an hannoverschen Bauwerken. Hannoversche Geschichtsblätter, Neue Folge 31(1/2): 105–152.
Henderson, D. M. 2006. Simulated weathering of dinosaur tracks and the implications for their characterisation. Canadian Journal of Earth Sciences 43: 691–704.
Hornung, J. J., and A. Böhme. 2012. The ‘German Wealden’ and the Obernkirchen sandstone: an introduction; pp. 62–72 in A. Richter and M. Reich (eds.), Dinosaur Tracks 2011. An International Symposium, Obernkirchen, April 14–17, 2011, Abstract Volume. Universitätsdrucke Göttingen, Göttingen, Germany.
Hornung, J. J., A. Böhme, and M. Reich. 2012. The type material of the theropod ichnotaxon ‘Bueckeburgichnus’ maximus Kuhn, 1958 – reconsidered; p. 27 in A. Richter and M. Reich (eds), Dinosaur Tracks 2011. An International Symposium, Obernkirchen, April 14–17, 2011, Abstract Volume. Universitätsdrucke Göttingen, Göttingen, Germany.
Hornung, J. J., J. König, and M. Reich. 2012. The type material of the ankylosaurian ichnospecies Metatetrapous valdensis Nopcsa, 1923 (Early Cretaceous, northern Germany); p. 29 in A. Richter, and M. Reich (eds.), Dinosaur Tracks 2011. An International Symposium, Obernkirchen, April 14–17, 2011, Abstract Volume. Universitätsdrucke Göttingen, Göttingen, Germany.
Hornung, J. J., A. Böhme, N. Schlüter, and M. Reich. 2016. Diversity, ontogeny, or both? A morphometric approach to iguanodontian ornithopod (Dinosauria: Ornithischia) track assemblages from the Berriasian (Lower Cretaceous) of northwestern Germany; chap. 12 in P. L. Falkingham, D. Marty, and A. Richter (eds.), Dinosaur Tracks: The Next Steps. Indiana University Press, Bloomington, Indiana.
Hornung, J. J., A. Böhme, T. van der Lubbe, M. Reich, and A. Richter. 2012. Vertebrate tracksites in the Obernkirchen Sandstone (late Berriasian, Early Cretaceous) of northwest Germany: their stratigraphical, palaeogeographical, palaeoecological, and historical context. Paläontologische Zeitschrift 86: 231–267.
Hübner, T. 2016. Elusive ornithischian tracks in the famous Berriasian (Lower Cretaceous) ‘Chicken Yard’ tracksite of northern Germany: quantitative differentiation between small tridactyl trackmakers; chap. 16 in P. L. Falkingham, D. Marty, and A. Richter (eds.), Dinosaur Tracks: The Next Steps. Indiana University Press, Bloomington, Indiana.
Jackson, S. J., M. A. Whyte, and M. Romano. 2009. Laboratory-controlled simulations of dinosaur footprints in sand: a key to understanding vertebrate track formation and preservation. Palaios 24: 222–238.
Jones, L. S., and E. R. Gustason. 2006. Dinosaurs as possible avulsion enablers in the Upper Jurassic Morrison Formation, East-Central Utah. Ichnos 13: 31–41.
Kvale, E. P., G. D. Johnson, D. L. Mickelson, K. Keller, L. C. Furer, and A. W. Archer. 2001. Middle Jurassic (Bajocian and Bathonian) dinosaur megatracksites, Bighorn Basin, Wyoming, U.S.A. Palaios 16: 233–254.
Lepper, J. 1997. Naturwerksteine in Niedersachsen. Zeitschrift für angewandte Geologie 41(1): 3–10.
Lepper, J., and A. Richter (eds.). 2010. Steine an der Leine: Naturwerksteine im Stadtbild von Hannover. E. Schweizerbart’sche Verlagsbuchhandlung, Stuttgart, Germany, 90 pp.
Li, R., M. G. Lockley, M. Matsukawa, and M. Liu. 2015. Important dinosaur-dominated footprint assemblages from the Lower Cretaceous Tianjialou Formation at the Houzuoshan Dinosaur Park, Junan County, Shandong Province, China. Cretaceous Research 52: 83–100.
Lockley, M. G. 1993. Auf den Spuren der Dinosaurier. Dinosaurierfährten – Eine Expedition in die Vergangenheit. Birkhäuser Verlag, Basel, Switzerland, 313 pp.
Lockley, M. G., and C. Meyer. 2000. Dinosaur Tracks and Other Fossil Footprints from Europe. Columbia University Press, New York, New York, 323 pp.
Lockley, M. G., and K. Conrad. 1989. The palaeoenvironmental context, preservation, and palaeoecological significance of dinosaur tracksites in the western USA; pp. 121–134 in D. D. Gillette and M. G. Lockley (eds.), Dinosaur Tracks and Traces. Cambridge University Press, Cambridge, UK.
Lockley, M. G., K. J. Houck, and N. K. Prince. 1986. North America’s largest trackway site: implications for Morrison Formation paleoecology. Geological Society of America Bulletin 97: 1163–1176.
Lockley, M. G., L. Xing, J. A. F. Lockwood, and S. Pond. 2014. A review of large Cretaceous ornithopod tracks, with special reference to their ichnotaxonomy. Biological Journal of the Linnean Society 113: 721–736.
Lockley, M. G., J. D. Harris, R. Li, L. Xing, and T. van der Lubbe. 2016. Two-toed tracks through time: on the trail of ‘raptors’ and their allies; chap. 11 in P. L. Falkingham, D. Marty, and A. Richter (eds.), Dinosaur Tracks: The Next Steps. Indiana University Press, Bloomington, Indiana.
Lubbe, T. van der, A. Richter, and A. Böhme. 2009. Velociraptor’s sisters: first report of troodontid tracks from the Lower Cretaceous of northern Germany. Journal of Vertebrate Paleontology 29(3, Supplement): 194A.
Manning, P. L. 2004. A new approach to the analysis and interpretation of tracks: examples from the Dinosauria; pp. 93–123 in D. McIlroy (ed.), The Application of Ichnology to Palaeoenvironmental and Stratigraphic Analysis. Special Publication 228. Geological Society, London, UK.
Martill, D. M., and D. Naish. (eds.) 2001. Dinosaurs of the Isle of Wight. Field Guide No. 10. The Palaeontological Association, London, UK, 433.
Marty, D. 2008. Sedimentology, taphonomy, and ichnology of Late Jurassic dinosaur tracks from the Jura carbonate platform (Chevenez-Combe Runde tracksite, NW Switzerland): insights into the tidal-flat palaeoenvironment and dinosaur diversity, locomotion, and palaeoecology. PhD dissertation, University of Fribourg, Fribourg, Germany. GeoFocus 21, 278 pp.
Marty, D., A. Strasser, and C. A. Meyer. 2009. Formation and taphonomy of human footprints in microbial mats of present-day tidal-flat environments: implications for the study of fossil footprints. Ichnos 16(1–2): 127–142.
McCrea, R., L. Buckley, J. O. Farlow, M. G. Lockley, P. J. Currie, N. A. Matthews, and S. G. Pemberton. 2014. A ‘Terror of Tyrannosaurs’: the first trackways of tyrannosaurids and evidence of gregariousness and pathology in Tyrannosauridae. PLoS One 9(7): e103613.
Milàn, J. 2006. Variations in the morphology of emu (Dromaius novaehollandiae) tracks reflecting differences in walking pattern and substrate consistency: ichnotaxonomic implications. Palaeontology 49(2): 405–420.
Milàn, J., D. B. Loope, and R. G. Bromley. 2008 Crouching theropod and Navahopus sauropodomorph tracks from the Early Jurassic Navajo Sandstone of USA. Acta Palaeontologica Polonica 53: 197–205.
Milàn, J., L. B. Clemmensen, and N. Bonde. 2004. Vertical sections through dinosaur tracks (Late Triassic lake deposits, East Greenland): undertracks and other subsurface deformation structures revealed. Lethaia 37: 285–296.
Milner, A. R. C., J. D. Harris, M. G. Lockley, J. I. Kirkland, and N. A. Matthews. 2009 Bird-like anatomy, posture, and behaviour revealed by an Early Jurassic theropod dinosaur resting trace. PLoS One 4: e4591.
Pelzer, G. 1998. Sedimentologie und Palynologie der Wealden-Fazies im Hannoverschen Bergland. Cour. Forschungsinstitut Senckenberg 207: 211 pp., 27 app.
Pelzer, G., and V. Wilde. 1987. Klimatische Tendenzen während der Ablagerungen der Wealden-Fazies in Nordwesteuropa. Geologisches Jahrbuch, Serie A 96: 239–263.
Petti, F.M., M. Bernardi, P. Ferretti, R. Tomasoni, and M. Avanzini. 2011. Dinosaur tracks in a marginal marine environment: the Coste dell’Anglone ichnosite (Early Jurassic, Trento Platform, NE Italy). Italian Journal of Geosciences 130(1): 27–41.
Reineck, H. 1963. Parameter von Schichtung und Bioturbation. Geologische Rundschau 56: 420–438.
Richter, A., A. Böhme, and T. Van der Lubbe. 2009. ‘Chicken Run’: a new unusual, heavily dinoturbated tracksite from the Lower Cretaceous sandstones of Obernkirchen, northern Germany. Journal of Vertebrate Paleontology 29(3, Supplement): 171A.
Richter, A., J. J. Hornung, A. Böhme, and U. Stratmann. 2012. Excursion Guide A1: Obernkirchen sandstone quarries: a natural workstone lagerstaette and a dinosaur tracksite; pp. 73–99 in A. Richter and M. Reich (eds), Dinosaur Tracks 2011. An International Symposium, Obernkirchen, April 14–17, 2011, Abstract Volume. Universitätsdrucke Göttingen, Göttingen, Germany.
Richter, R. 1936. Marken und Spuren aus dem Hunsrückschiefer. II Schichtung und Grundleben. Senckenbergiana 18: 215–244.
Romilio, A., and S. W. Salisbury. 2011. A reassessment of large theropod dinosaur tracks from the mid-Cretaceous (late Albian-Cenomanian) Winton Formation of Lark Quarry, centralwestern Queensland, Australia: a case for mistaken identity. Cretaceous Research 32: 135–142.
Romilio, A., R. T. Tucker, and S. W. Salisbury. 2013. Reevaluation of the Lark Quarry dinosaur tracksite (late Albian-Cenomanian Winton Formation, central-western Queensland, Australia): No longer a stampede? Journal of Vertebrate Paleontology 33(1): 102–120.
Santos, V. F., P. M. Callapez, and N. P. C. Rodrigues. 2012. Dinosaur footprints from the Lower Cretaceous of the Algarve Basin (Portugal): new data on the ornithopod palaeocology and palaeobiogeography of the Iberian Pensinsula. Cretaceous Research 40: 158–169.
Scott, J. J., R. W. Renaut, and R. B. Owen. 2012. Impacts of flamingos on saline lake margins and shallow lacustrine sediments in the Kenya Rift Valley. Sedimentary Geology 277/278: 32–51.
Scott, J. J., R. W. Renaut, R. B. Owen, and W. A. S. Sarjeant. 2007. Biogenic activity, trace formation, and trace taphonomy in the marginal sediments of saline, alkaline lake Bogoria, Kenya Rift Valley; pp. 311–332 in SEPM (ed.), Sediment-Organism Interactions: A Multifacetes Ichnology. Special Publication 88. Society for Sedimentary Geology, Tulsa, Oklahoma.
Souza Carvalho, I. de, L. Borghi, and G. Leonardi. 2013. Preservation of dinosaur tracks induced by microbial mats in the Sousa Basin (Lower Cretaceous), Brazil. Cretaceous Research 44: 112–121.
Struckmann, C. 1880a. Die Wealden-Bildungen der Umgegend von Hannover. Eine geognostische Darstellung. Hahn’sche Buchhandlung, Hannover, Germany, 122 pp.
Struckmann, C. 1880b. Vorläufige Nachricht über das Vorkommen grosser vogelähnlicher Thierfährten (Ornithoidichnites) im Hastingssandsteine von Bad Rehburg bei Hannover. Neues Jahrbuch für Mineralogie, Geologie und Palaeontologie Jahrgang 1881 (I. Band): 125–128.
Sweetman, St. C. 2004: The first record of velociraptorine dinosaurs (Saurischia, Theropoda) from the Wealden (Early Cretaceous, Barremian) of southern England. Cretaceous Research 25: 353–364.
Thulborn, R. A., and M. Wade. 1979. Dinosaur stampede in the Cretaceous of Queensland. Lethaia 12: 275–517.
Thulborn, R. A., and M. Wade. 1984. Dinosaur trackways in the Winton Formation (mid-Cretaceous) of Queensland, Memoirs of the Queensland Museum 21: 413–517.
Thulborn, R. A., and M. Wade. 1989. A Footprint as a History of a Movement; pp. 51–56 in D. D. Gillette and M. Lockley (eds.), Dinosaur Tracks and Traces. Cambridge University Press, Cambridge, UK.
Wings, O., A. Broschinski, and N. Knötschke. 2005. New theropod and ornithopod dinosaur trackways from the Berriasian of Münchehagen (Lower Saxony, Germany). Kaupia, Darmstädter Beiträge zur Naturgeschichte 14: 105.
Wings, O., J. N. Lallensack, and H. Mallison. 2016. The Early Cretaceous dinosaur trackways in Münchehagen (Lower Saxony, Germany): 3D photogrammetry as basis for geometric morphometric analysis of shape variation and evaluation of material loss during excavation; chap. 3 in P. L. Falkingham, D. Marty, and A. Richter (eds.), Dinosaur Tracks: The Next Steps. Indiana University Press, Bloomington, Indiana.
Wright, J. 1999. Ichnological evidence for the use of the forelimb in iguanodontid locomotion; pp. 209–219 in A. R. Milner and D. J. Batten (eds.), Life and Environments in Purbeck Times. Special Papers in Palaeontology 68. The Palaeontological Association, London, UK.