CHAPTER 16
Protected cultivation
Summary
This chapter deals with the benefits of protecting plants from low temperatures as well as from rain, hail, sleet and snow, and discusses the various forms of protection that are available. It examines the ways in which the provision of protection will modify the plant’s environment, how plants will respond to these changes, and how any harmful effects can be mitigated. It also looks at the respective benefits of the various systems that can be added to a basic greenhouse, such as heating, ventilation, carbon dioxide enrichment, lighting and irrigation. It does not look at the cultivation of mushrooms, which are grown entirely in the dark. Finally, it considers the crops that are grown under protection on commercial nurseries and whether the techniques that are used can be employed by gardeners.
Why protect plants?
Most gardeners recognise that, even if they plant trees and hedges as windbreaks, the plants growing in their gardens are still at the mercy of the weather. In the United Kingdom, the high temperatures and bright sunshine that should be the norm in summer are inevitably followed by dull winter days that are very likely accompanied by freezing conditions. Garden plants are also likely to be exposed to strong winds and heavy rain at any time of year and are liable to be nibbled by birds, rabbits and other animals as well being attacked by insects, bacteria and viruses. All of these factors can damage plants and restrict their growth. Although it is true that native plants have evolved to tolerate and to survive most of these conditions, the plants grown in gardens have often been introduced from areas with warmer climates. For these plants in particular, exposure to low temperatures is often very damaging, especially if the temperature falls below 0°C, the freezing point of water. Such plants may even die if the freezing conditions continue for more than a few days.
Plants have evolved various ways of limiting the impact of freezing conditions. The most common of these is shown by deciduous trees and shrubs, which lose their leaves in autumn and enter a period of dormancy (see Chapter 9). In this state, the growing points at the tips of their shoots are protected within buds enclosed by bud-scales. However, even this approach does not always ensure survival especially if an early period of warm weather occurs after winter dormancy has been broken by exposure to low temperature, and consequently bud growth occurs prematurely. If the young developing shoots then experience a period of frost they may be damaged or even killed. For all of these reasons, if the plants themselves or their leaves, flowers or fruits are valuable to the grower in financial, ornamental or even sentimental terms, it is worth considering how to protect them (protected cultivation).
The most usual way of doing this is either to cover the whole plant with an appropriate material or to move the plants to a location where they can be more readily protected. The degree of protection depends upon the materials that are available or can be bought, and the degree of control over the environment that the gardener wishes to provide. If covering of the whole plant is either too expensive or too difficult to do, it may be sufficient to cover just the parts considered most valuable, such as the flowers.
What are the benefits?
The most obvious benefit is that protection lessens the physical damage caused by wind or by rain, hail and snow, and even animals, and so helps to maintain the quality and appearance of the above-ground parts of plants. Even relatively simple forms of protection enable the gardener to protect plants from frost, but the gardener has more control over air temperature with more sophisticated forms of protection, such as a greenhouse. A greenhouse can be used to improve the speed and uniformity of propagation from seeds or cuttings, and it allows propagation to begin earlier in the growing season thus conferring a head start on those plants. The gardener’s ability to control temperature is greatly improved if a heater is installed, and this confers many advantages, including the ability to advance the time of flowering and to improve the quality and the quantity of the harvestable products. Finally, tender plants can be moved into a heated greenhouse to keep them alive over winter or simply to extend their growing season.
Temperature
Control of temperature is beneficial because most of the biochemical processes that occur in plants and that enable them to grow in size and to develop in complexity are highly dependent on temperature (see Chapter 8). Almost all of them proceed more quickly as the temperature rises and so the plant then grows more quickly, produces leaves faster, comes into flower earlier, and its fruits mature and ripen earlier. On the other hand, if the temperature falls and approaches 0°C, most of these same biochemical processes will slow and eventually stop altogether, especially when the temperature falls below 0°C.
Respiration is probably the main biochemical process that responds to temperature. During this process, some of the sugars created by photosynthesis are broken down again to release energy (see Chapter 2), which is then used to convert other sugars into the proteins and cell walls required for plant growth. At a temperature of 5°C or so, respiration proceeds very slowly, but as the temperature rises the rate increases very rapidly. Indeed, as a general rule, the rate of most biochemical processes doubles for every 10°C rise in temperature, a response that is called exponential (Fig. 16.1). The increase in temperature eventually reaches an optimum at which the rate of the biochemical process is at its maximum. Thereafter, the rate falls again as the temperature continues to rise and eventually ceases if the temperature becomes too high.
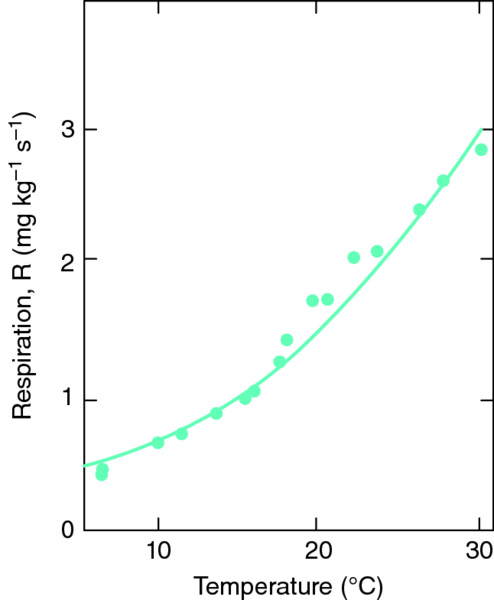
Figure 16.1 The effect of temperature on a biochemical process such as respiration.
Development
The relationship between the rate of plant development and temperature is similar to that described above but is often less curvilinear. Indeed, over the range that is most relevant to protected cultivation (10–25°C), it is simpler to assume that the rate of production of flowers and fruits increases linearly with increasing temperature (Fig. 16.2). Furthermore, the optimum temperature for a developmental process is often much lower than the optimum for respiration. In order to use a heating system in the most efficient way, it is obviously important also to know whether a plant process responds more to the temperature by day than by night, or vice versa. Unfortunately, it is usually quite difficult to answer this question as the lengths of the day and the night continually change in a natural environment.
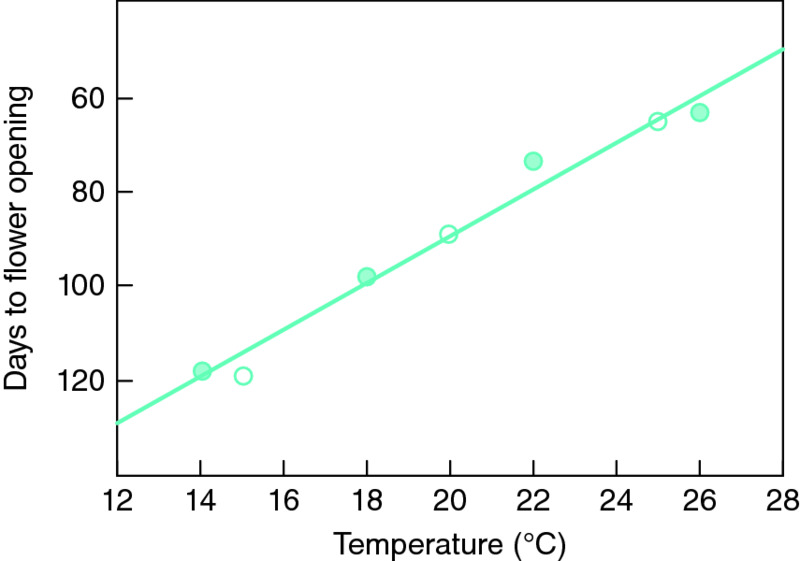
Figure 16.2 The effect of temperature on a developmental process such as flowering of chrysanthemum.
The question was answered very clearly in relation to the flowering of chrysanthemum (Chrysanthemum morifolium) and it was done using cabinets equipped both with heating and cooling. The walls and the roof of these cabinets were transparent, so that the plants grew in daylight. The day and the night periods were kept the same length, namey 12 hours, either by placing blackout covers over the cabinets or by using artificial lighting as required, depending upon the time of year. In a series of experiments, the plants were given either a high day temperature (20°C) followed by a low night temperature (10°C) or a low day temperature (10°C) followed by a high night temperature (20°C). As the day and the night periods were the same length in both treatments, their average temperature was 15°C and this was given throughout the day and the night in the third treatment.
It was expected that the plants would be more sensitive to the night temperature but most of the plants flowered at the same time in all three treatments (Fig. 16.3). A few varieties showed a slight delay in flowering in the treatment with the low night temperature but the delay was small. Overall, the result indicated that flowering was responding to the actual temperature the plant was exposed to at any moment, regardless of whether the plant was in the light or the dark. Similar responses have now been shown to occur in tomato (Solanum lycopersicum) and in many other plants. For all of these, it seems reasonable to conclude that their rate of development over a period of days or weeks is controlled by the average temperature to which they have been exposed over the whole of that time, rather than to either the day or the night temperature alone. Consequently, air temperature in a greenhouse does not have to be set at a specific day or night temperature to control flowering or fruit development but can be set to achieve a given average temperature.

Figure 16.3 The separate effects of day and night temperature on flowering and stem growth of chrysanthemum. The treatments are from left to right; 10°C day/20°C night; 15°C day/15°C night; 20°C day/10°C night. The night and the day each lasted 12 hours. Reproduced by permission of Cockshull, Hand and Langton. Further details in Cockshull, Hand, and Langton (1982) Acta Horticulturae 125, 101–10.
Stem growth
The above conclusion evidently does not apply to all developmental processes, as the plants in the chrysanthemum experiment differed greatly in height (Fig. 16.3). Indeed, stem elongation was strongly influenced by whether a particular temperature occurred in the day or in the night. In particular, the high temperature of 20°C was much more effective at promoting stem length if it was given during the day rather than the night. Indeed, the combination of high day and low night temperature, which does occur naturally in summer, gave very tall, leggy plants, and it is now thought that this kind of response occurs in most plants. The shortest plants were produced by the treatment with the day temperature (10°C) lower than the night temperature (20°C). Short stems are particularly desirable in flowering pot plants, and although this sequence of temperatures is rarely found in nature, it can be produced in a heated greenhouse.
Further research with other plants has shown that compact plants are almost always produced when the night temperature is warmer than that by day. Furthermore, it appears that the low day temperature does not have to be given all day, as compact plants are also obtained if the low temperature is given for just a short period in the early morning. This treatment has been described as a DROP treatment, as the temperature is lowered for an hour or so at the time of the change from night to day and is then raised again. The DROP treatment is now regularly used by many producers of pot plants as it lessens the need to control plant height by applying chemical growth retardants (see ‘Plant growth regulators’, below).
Cold tolerance
The subject of cold tolerance is a very complex one as species vary in how well they can tolerate a particular low temperature and whether they can tolerate it over a long period of time. One component of cold tolerance is evidently related to a plant’s ability to stop ice crystals forming in its cells. However, this is not the only cause of plant death as the temperature does not have to fall below 0°C to cause the death of the tomato (Solanum lycopersicum). Indeed, many tomato varieties will not tolerate a temperature of even 5°C and so must be grown under protection in many countries.
Principles of protection
Energy transfer
Before discussing the different types of protection available it is necessary first to consider energy transfer. Plants that are growing in the open are exposed to energy radiating from the sun by day. This radiant energy covers an enormous range of wavelengths but the range that gardeners are mainly concerned with extends from the short wavelengths of ultraviolet light (UV) to the very long wavelengths of infrared radiation (IR). As described in Chapter 8, ‘visible radiation’ or ‘light’ is radiation that can be detected by pigments in the eyes of humans and other animals to produce the sense of vision. Visible radiation includes short wavelengths that are sensed as blue light and extends through yellow and red light to the longer wavelengths of far-red light. Almost one-half of all the radiant energy emitted by the sun is at wavelengths that are visible, whereas the remainder is emitted at very long wavelengths and constitutes the thermal or infrared radiation.
Most of the visible radiation can be absorbed by pigments in plant leaves, especially chlorophyll, and this is the first step in photosynthesis (see Chapter 2). Indeed, all of the radiant energy emitted by the sun can be absorbed by at least some of the molecules in plant tissues, and especially by water. Much of the energy that is absorbed serves to warm plant tissues and it is their increase in temperature that drives the evaporation of water. The water vapour escapes through the minute holes in leaves called stomata, and the process of water loss is called transpiration (see Chapters 2 and 8). All objects that are exposed to solar radiation will absorb some radiation and be warmed by it, but this energy can quickly be lost again to the surrounding air by means of conduction and convection. Energy can also be lost by radiation to the sky, and such losses occur most rapidly when the sky is cloudless.
One of the purposes of protection is to reduce all of these losses of energy so that the protected objects are kept warmer for longer. One way to do this is to employ a protective covering that blocks the loss of radiant energy. This blocking mechanism is frequently called the ‘greenhouse effect’ and is shown particularly well by glass as it is transparent to visible radiation but not to the long wavelengths of infrared radiation. This mechanism is also exhibited by some gases in the atmosphere, especially carbon dioxide (CO2) and methane (CH4), and such gases are popularly known as ‘greenhouse gases’ (see Chapter 15).
The importance of light
Although plants grow slowly if the temperature is low, most will not grow at all if they are deprived of light. It is important, therefore, that any form of protection must allow some light to reach the leaves of the plants that are being protected. Light is required to power photosynthesis, the process by which CO2 in the atmosphere is fixed into sugars (see Chapter 2), the basic building blocks from which all the other compounds required for plant growth are synthesised.
Because light is of such overriding importance for green plants, if maximum productivity is required as well as protection it is essential that any covering material should transmit most of the light that falls upon it. That is not to say that translucent materials, i.e. those that scatter light, are not useful, especially in summer. Such materials cause the direct beam of sunlight to be diffused and cause more light to fall on the lower leaves of a plant’s leaf canopy. Sometimes, gardeners want a plant to develop in the dark, as with the ‘forcing’ of rhubarb. In such cases, the part of the plant that is going to be eaten is etiolated rather than green, as would be the case if it had been exposed to light in the usual way (see Chapter 8). When forced in this way, the compounds required for new shoot growth must come from reserves stored in the root or in a storage organ such as a seed, bulb, or tuber.
Cladding materials
Any material that covers a structure placed over plants is usually referred to as its cladding. An ideal cladding material, therefore, is one that provides some thermal insulation while allowing light to reach the plants within the structure. Most claddings provide some thermal insulation by limiting air movement beneath them. Air movement causes warm air to be moved to the inner surface of the cladding from where energy will be lost to the outside, mainly by conduction through the cladding. Warm objects also emit long-wave, IR radiation, and the most efficient claddings stop this from being radiated to the sky.
Glass
Glass is still a favourite cladding material, partly because of its transparency and partly because it is relatively sturdy and provides physical protection against rain, hail, snow and wind. The glass intended for greenhouses is normally supplied as 3 mm thick panes, and it should be checked to ensure its iron content is not too high. Glass with a high content of iron transmits less light than horticultural glass of good quality, and this can readily be detected by placing the two next to one another and then viewing the sky through them.
As described above, plants grown under glass will be warmer than in the open partly because the air around them is still, which limits the loss of heat through the cladding. However, the main advantage of glass is that it absorbs the long wavelength, IR radiation emitted by the warm plants and soil, and so keeps most of that energy within the protective structure. Glass also blocks the transmission of UV radiation coming from the sun, but this can be a disadvantage if the spread of a particular fungal disease is inhibited by UV.
Film plastics
Film plastics have the advantage of lower cost compared to glass but their lifespan may be as short as 3–5 years. Film plastics also provide a more flexible cladding than glass and are well suited to covering curved structures. However, they must still be secured to the frame to stop them being blown away. As with glass, a film-plastic cladding stops air movement under the material and so limits heat loss through the cladding.
Modern film plastics are usually laminates of three or more layers, and it is feasible to add specific chemicals to one or more of the layers to give them special properties. One very successful approach has been to incorporate additives that cause any condensation on the inner surface of the film to form a film of liquid rather than droplets. These ‘anti-fog’ or ‘anti-drip’ films create a more pleasant environment for people working underneath the film, and it is better for the plants too as the droplets could cause damage and encourage disease. Most film plastics designed as cladding materials are translucent rather than transparent and are now treated to resist degradation by the UV in sunlight.
Other additives that have been used include some that selectively absorb certain regions of the solar spectrum. As mentioned earlier, some films block the transmission of UV to the plants below. However, there is evidence that UV wavelengths can induce an increase in the concentration of health-giving compounds in some plants and so it may be beneficial to use a film that permits UV transmission when growing such plants. There are also some very specialist applications where the transmission of only red or blue light is desirable in order to elicit a particular plant response. For example, plants tend to be more compact if grown under a film plastic that allows blue light to be transmitted while reducing the transmission of all other colours.
Additives that block the outward transmission of the long-wave, infrared (IR) radiation emitted by warm objects are also widely used. Cladding materials that exhibit this ‘greenhouse effect’ enable the plants to remain warmer for longer, and so the plants grow and develop more quickly. Although most film plastics are made solely of polyethylene, some commercially available films are made from ethylene-vinyl acetate (EVA) together with polyethylene, which gives them greater mechanical strength.
Bubble plastic sheets
Bubble plastic is manufactured by trapping bubbles of air between the layers of plastic. Most types allow reasonable light transmission and have the effect of scattering sunlight, which many people consider to be an advantage. One type that is especially made for cladding tunnels and small greenhouses is manufactured from the very strong sheet plastic ‘Keder’. This bubble plastic has good thermal insulation properties and a much longer useful life than conventional film plastics.
Polycarbonate sheets
Polycarbonate can be manufactured as twin-walled sheets, which have good light transmission combined with much better insulation properties than single sheets of glass. Polycarbonate sheets are also lighter and stronger than panes of glass and are now treated to minimise the discoloration that used to occur as the sheets aged. Although they have many desirable properties, they are a relatively expensive cladding material.
Types of protection
Although it is relatively easy to describe the benefits associated with providing plants with an environment that is warmer and more protected than in the open, it is more difficult to suggest how this might be achieved without too great a cost.
Mulches
Traditionally, mulching was the action of putting a protective covering of organic matter around plants to prevent the growth of weeds, and to conserve water by restricting evaporation from the soil. Originally the mulch would probably have been leaves or straw, but now the simplest way to mulch is to lay a sheet of film plastic on the soil. Holes or slits can then be made in the sheet through which the roots of seedlings and young plants can be pushed into the soil. Plastic sheets perform the same functions as organic mulches but they are better at warming the soil and thus enhancing root activity. With strawberries (Fragaria × ananassa), there is the additional benefit that a plastic mulch also stops the fruit from making contact with the soil.
Black plastic sheets have the advantage of being good at absorbing solar radiation, and the accompanying increase in soil temperature under the mulch enables the roots to function better than if they were in bare soil. However, if laid too early in the year, especially if the nights are cloudless and clear, a black sheet will radiate energy to the clear sky, which may cause the covering and the plant to become frosted. A clear polythene sheet may, therefore, be a safer option in the very earliest part of the growing season.
Horticultural fleeces and meshes
A horticultural fleece is a lightweight, unwoven fabric that allows some sunlight to pass through it together with rain and irrigation sprays. Horticultural fleeces create a still microenvironment around the protected plants that helps to keep the air around them warmer. On nights when there is a cloudless sky, the plants and their surrounding soil will radiate less energy to the sky if they are beneath a horticultural fleece, and so they will be warmer than unprotected plants. Consequently, wrapping tender plants in horticultural fleece is a very effective way of protecting them from frosts.
The term ‘mesh’ is usually reserved for plastic materials that are woven. Their properties are similar to those of fleeces but they are more permeable to water and are often stronger. They can also be used to protect plants from rabbits and birds and from some insect pests if their pore size is small enough.
Cloches and cold frames
Cloches consist of a protective covering that is either self-supporting or that employs very simple supports. The simplest arrangement is probably to lean pairs of glass sheets against one another to form an inverted V-shape, although it is important that the ends of cloches are closed either partially or completely as otherwise their effectiveness is reduced. One alternative to glass is to use a film-plastic sheet draped over supports in the shape of an arch. The sheet should be wide enough to cover at least one row of young plants.
Cold frames are constructed with robust walls of wood or brick, and the protective covering is stretched over a frame that rests on these walls. This arrangement has the advantage that the frame and its cladding can easily be removed, either partially or completely, whenever it is felt that the air within the cold frame should be mixed with the outside air. This is usually done to allow the plants to become acclimatised to a colder environment, that is to ‘harden them off’, before planting them in the open.
Ideally, the protective coverings used in cloches and cold frames should be transparent or translucent and they should restrict air movement underneath them so as to reduce heat loss. The covering should also provide protection against rain, hail and snow. If glass is used, the enclosed air will also become warmer because of the ‘greenhouse effect’ described earlier, but the disadvantage of using glass for cloches and cold frames is that it can be broken relatively easily. Twin-walled polycarbonate sheets are tougher than glass but are more expensive.
Tunnels
Tunnels provide a reasonably low-cost form of protection. Small tunnels are similar to cloches, but the larger, walk-in tunnels are made by erecting a series of galvanised steel tubes shaped into arched frames (sometimes called ‘hoops’) and then stretching a film-plastic cover over the length of them. Tunnels can be of any width up to the maximum size of film-plastic sheet that is available. The most commonly used film plastic is polythene, and its insulation properties can be improved if the manufacturer incorporates into it an additive that blocks the outward transmission of IR, thermal radiation. The disadvantage of film plastics is that their life is limited and so tunnels may have to be recovered every 3–5 years.
A relatively recent development is the use of tunnels to protect soft fruit, including raspberries (Rubus idaeus), strawberries (Fragaria x ananassa) and cherries (Prunus avium). The primary purpose of these tunnels is to protect the fruit so the polythene covering need only be present while the fruits are developing. The production of strawberries is dealt with later, but with cherries the trees are grafted onto a dwarfing rootstock and the tunnels can be up to 5.5 m high and 9 m wide (c. 19 × 30 ft). Many tunnels can be joined together at their gutters to create multi-bay arrays with no side walls except around the periphery of the array. The film-plastic cover is drawn over the trees just before they begin to flower and is removed again once the cherries have been picked (Fig. 16.4). The merits of this system are that the fruits are protected from rain-induced cracking as well as from wind, birds and even insects, so the yield of high-quality fruit is greatly increased. The fruit can also be picked earlier.

Figure 16.4 Cherry trees flowering in a tunnel clad with a film-plastic. Reproduced by permission of Pink Sky, courtesy of Lower Hope Farm, Herefordshire.
Greenhouses
The most advanced of the conventional forms of protection is the greenhouse. These range from small structures in domestic gardens (Fig. 16.5) to the very large structures in which commercial crops are grown (Fig. 16.5). The smaller structures can be clad with any of the materials used to cover tunnels but the larger structures are mainly clad with glass and are then usually called glasshouses. The structure and the glazing bars of small greenhouses can be made of cedar wood but it is more common for all greenhouses now to be constructed largely from aluminium extrusions. Modern commercial glasshouses are mainly based upon the standard ‘Venlo’ structure with a span of 3.2 m (c. 10 ft 6 in). The individual units are joined together at their gutters and have no side walls, except around the periphery of the whole structure. As it is more economical to have large structures for the commercial production of crops, each glasshouse can cover an area of 5 hectares (12 acres) or more and their gutters can be 4.5–5 m above ground level.


Figure 16.5 (a) A small greenhouse suitable for a garden. Photograph by K.E. Cockshull. (b) A large, modern glasshouse suitable for growing tomatoes commercially. Photograph by K.E. Cockshull.
There are also large speciality glasshouses such as ‘The Glasshouse’ at the RHS Garden at Wisley, Surrey. This is made largely from curved panes of toughened glass and its roof is 12.5 m (c. 41 ft) above the ground at its highest point (Fig. 16.6). Even larger are the domed biomes of the Eden Project in Cornwall, the largest of which is 35 m high (c. 115 ft) (Fig. 16.7). These biomes are constructed largely from hexagonal, inflated plastic cells made from ETFE (ethylene tetrafluoroethylene), a UV-transparent film plastic.

Figure 16.6 ‘The Glasshouse’ at the Royal Horticultural Society’s Garden, Wisley, Surrey. Photograph by K.E. Cockshull.

Figure 16.7 Biomes at the Eden Project in Cornwall. Photograph by K.E. Cockshull.
In general, where there is a choice it is best to site greenhouses where they will encounter the minimum of shading from external objects such as walls, trees and other buildings. It is also desirable that they are sheltered from wind, as wind-speed is a major determinant of heat loss. With large commercial glasshouses it is an advantage to orient the ridges of the component Venlo structures so that they run east to west, as this should maximise light transmission into the structure in winter. However, it is also desirable to run crop rows from north to south so that one side of the row benefits from the sun in the morning and the other in the afternoon; on average, therefore, both sides should receive the same amount of light.
The conventional greenhouse normally spreads horizontally over the ground with the plants firmly based in the ground or resting on it, but over the years it has often been suggested that glasshouses should be built vertically. The usual arguments for this are that the footprint of the structure can then be smaller and it could perhaps be sited closer to where edible crops are consumed. Unfortunately the cost of building vertically rather than horizontally is much greater and the engineering problems are much more complex. These problems are made worse if the crops are all to be given the same amount of light each day by growing them in troughs on endless-belt systems that pass up and down the greenhouse. Although such structures have been built, and a relatively low-level installation for producing animal feed does currently exist in the United Kingdom, none designed to produce crops for human consumption has yet proved to be commercially viable.
Systems to be added
Heating systems
The first system to consider adding to a greenhouse is some means of heating the air so that it is possible to keep the air temperature above 0°C at all times. It is also desirable and more economical to control the heating system via a thermostat. If the main objective of installing heating is simply to provide frost protection in a gardener’s small greenhouse, the thermostat should probably be set at about 5°C. This will allow plenty of time for the system to react and for heat to reach all parts of the greenhouse. Of course, if the cost of heating is of less concern, the thermostat can be set much higher than 5°C and a wide range of tender plants can be grown. Various heat sources are available to gardeners, including fan-assisted electric heaters (provided that they are approved for greenhouse use), and also propane burners, or even paraffin heaters, although the heat output of the latter cannot readily be controlled by a thermostat.
Most commercial growers have systems that rely on heating water in a boiler, and then circulating the hot water around the glasshouse through pipes laid mostly on the floor. The pumps that circulate the water are usually controlled via one or more aspirated thermostats within the glasshouse. Natural gas is the main fuel used but oil and propane are still used by some growers, and there is always interest in using wood chips and other forms of biomass as well as less conventional fuels.
Ventilation
For complete control of air temperature it is also necessary to have a controllable system of ventilation. It has already been stated that the air temperature will rise as radiant energy from the sun is trapped within a greenhouse, and there are circumstances when the air temperature can become too high, especially in summer. Ventilation allows this hot air to be exchanged for cooler air from outside the greenhouse, and this is usually accomplished by opening ventilators sited in the greenhouse roof. Roof-mounted ventilators generally pivot around the ridge, and in small greenhouses they are opened either manually or by pistons activated by a gas or a liquid that expands when heated. Electric motors are employed to open the ventilators in large greenhouses.
Ideally, the ventilators should occupy an area equivalent to about one-sixth of the floor area, but this is very difficult to achieve in a small greenhouse. Hence, such houses are prone to get very warm in summer even if side ventilators are also fitted. Air exchange is desirable not only to lower the greenhouse air temperature but also to allow the CO2 concentration of the greenhouse air to be replenished. This may be necessary if the photosynthetic activity of the plants in the greenhouse depletes the CO2 concentration below its normal atmospheric level (see ‘Carbon dioxide’, below). It may also be desirable to open the ventilators to allow atmospheric pollutants to escape or to lower the humidity of the air. A very high relative humidity of 95% or more at a temperature of 18°C can be detrimental to plants in a variety of ways, the most obvious being that it will encourage the development of fungal diseases. A less obvious one is that such conditions will restrict the loss of water from the leaves by transpiration. This in turn will produce localised calcium deficiencies within the plant, as calcium moves mainly in the transpiration stream, and a localised lack of calcium can cause tip-burn of lettuce leaves and blossom-end-rot of tomato fruit.
Energy saving
Saving energy and thereby reducing heating costs is always an issue for gardeners and commercial growers alike. For gardeners, once the obvious sources of heat loss in greenhouses have been dealt with, such as broken and poorly overlapping glass panes, and ill-fitting doors, the most practicable approach is to improve the structure’s thermal insulation. There are various ways of achieving this in winter, but probably the most satisfactory is to line the roof and the inner walls of the greenhouse with either film-plastic sheets or bubble plastic. The latter usually has the better insulating properties. It might even be worth considering replacing glass with twin-wall polycarbonate sheets, which have much superior insulation properties.
For commercial growers, the cost of heating a glasshouse can account for more than 25% of all production costs, depending on the crop and the length of time the glasshouse is heated. Consequently many commercial growers have now invested in alternative heat sources. Currently one of the most popular is ‘combined heat and power’ (CHP), which consists of an engine that burns a fuel such as natural gas for the purpose of generating electricity. The electricity is then sold to the National Grid, whereas the heat is used in the glasshouse. With some crops it may be worthwhile to use the electricity on site, for example to power supplementary lighting. In Holland there are communal systems where a large central unit generates electricity and the heat that is co-generated is distributed to numerous glasshouses. In other countries geothermal heat may be used to power individual or communal heating systems.
The use of alternative fuels such as biomass or even methane gas from an anaerobic digester of vegetable waste is also being considered but supplies of the alternative fuel need to be near-at-hand and inexpensive. A few growers are able to use the waste heat from an industrial operation. One such example is British Sugar in Norfolk, where the parent company generates heat for refining sugar and some of this heat is then used in its own glasshouse complex.
Another solution to the problem of conserving energy is reducing the volume of the glasshouse by installing an insulating screen from gutter to gutter across its width. This is awkward in small greenhouses as the gutters are too low and the screen will impede access, but it suits modern commercial glasshouses where the gutters are likely to be at least 4 m above the ground. A very popular ‘thermal screen’ is one manufactured by alternating thin strips of aluminised plastic and clear plastic to create a curtain-like material. The ratio of aluminised to clear strips can be prescribed, depending on whether the screen is required primarily for energy saving or for shading. When used for energy saving, the screen is drawn over the crop before nightfall and is then withdrawn again after dawn, although not necessarily immediately, unless the day is expected to be sunny.
Carbon dioxide (CO2)
As described in Chapter 2, photosynthesis is a light-dependent process by which carbon dioxide (CO2) is fixed into sugars, which can then be used to power plant growth. The response of leaf photosynthesis to CO2 concentration shows that the amount of CO2 that is fixed increases as the CO2 concentration rises, up to about 1000 parts per million (ppm CO2) (Fig. 16.8). Above this concentration there are some further benefits but they are small. These responses led to the idea of enriching the glasshouse atmosphere with CO2 in order to increase the rate of photosynthesis and thus increase crop yields. When much work on single leaf photosynthesis was done in the 1970s, the normal atmospheric CO2 concentration was about 340 ppm. At that time, enrichment to 1000 ppm CO2 produced a 50% increase in the rate of leaf photosynthesis, but the proportional benefit is becoming less as the normal atmospheric concentration of CO2 rises (see Chapter 15).
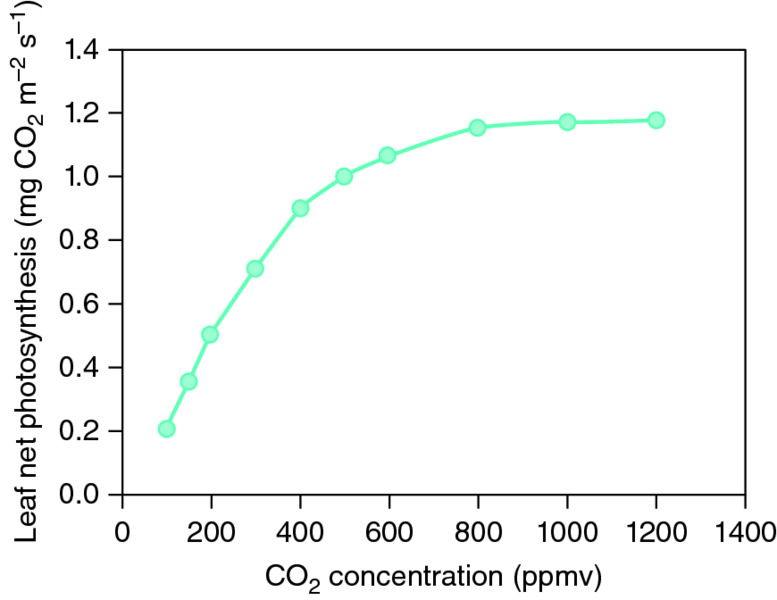
Figure 16.8 The effect of the concentration of CO2 in the glasshouse atmosphere on the photosynthetic rate of tomato leaves. ppmv, parts per million by volume.
It is quite feasible to add CO2 to the atmosphere of a greenhouse, either from cylinders of the compressed gas or by burning a hydrocarbon fuel such as paraffin or propane. Unfortunately, the main problem for gardeners is that it is also desirable to install a system for sampling and controlling the CO2 concentration that is achieved. The cost is worthwhile for commercial growers of tomato (Solanum lycopersicum) as fruit yield is closely linked to crop photosynthesis. Indeed, fruit yield can be increased by up to 35% if the CO2 concentration in the glasshouse is raised to 1000 ppm during daylight hours. Growers often obtain this CO2 by collecting the flue gases from a boiler or a CHP unit and then injecting them into the glasshouse. Unfortunately the by-products of burning a hydrocarbon fuel can also include other gases such as ethylene, carbon monoxide, sulphur dioxide and assorted oxides of nitrogen. Most of these can be harmful either to the plants or to humans and so they have to be removed, or at least reduced in concentration, before the flue gases can be injected into the glasshouse. Hence the choice of heating system for a commercial glasshouse is closely related to the cost of making the flue gases safe for injection into a glasshouse.
Because the rate of photosynthesis increases as the light intensity increases, there are times in summer when crop photosynthesis can be taking CO2 out of the greenhouse atmosphere faster than it can be replenished via ventilation. As a result, the CO2 concentration in the greenhouse atmosphere can be depleted below the normal atmospheric level. The gardener can minimise this problem in a small greenhouse by having the ventilators and doors open in summer. On the other hand, the commercial grower may have a serious problem, even with the ventilators wide open, especially if the crop is grown at high density. In such circumstances it can be economically worthwhile to add CO2 to the glasshouse atmosphere to bring its concentration back to the current concentration of 400 ppm or so. In certain circumstances it may even be desirable to burn a fuel just to produce CO2, in which case any hot water that is produced can be pumped into storage tanks (dump tanks) to be used later when heating is required.
Lighting
Although it is true that sunlight is free, the amount received each day is variable and there is not very much of it in the United Kingdom in winter. For these reasons, especially if growing commercial crops to a standard or for a specific occasion or market, it is worth considering augmenting sunlight by using an artificial light source. The one most commonly used to boost photosynthesis in commercial crop production is the high-pressure sodium lamp. Such lamps are expensive to install and operate, so their use has to be a compromise between what is desirable for the crop and what can be afforded by the grower. In one commercially viable installation, the light from each 600-watt high-pressure sodium lamp was spread over an area of about 15 m2 and provided some 12 W m−2 of photosynthetically useful radiation (c. 5 klux, see Table 8.1).
There is also considerable interest in the use of light-emitting diodes (LEDs) to boost plant growth. Their advantages are that although individual LEDs are of low wattage and their output is usually restricted to a specific wavelength (i.e. colour) they have a very long life. Furthermore they can be combined to form lamps having very specific properties, and they can be mounted very close to the plants as their output is accompanied by almost no IR radiation. As yet, however, they cannot generate very high irradiances with the same efficiency as the high-pressure sodium lamp and so are more suited to the raising of young seedlings in growth rooms (see ‘Special cases’, below).
The time of flowering of many plants, as well as leaf-fall, the onset of dormancy and the formation of storage organs all respond to day-length. These responses to day-length are called ‘photoperiodic’ (see Chapter 9). In essence, they are usually regulated by the number of hours of uninterrupted darkness that the plant receives in each 24-hour period. This can easily be controlled in a protected environment by covering the plants with a light-tight cover from late afternoon until early morning to produce long nights, or by switching on electric lights at night to produce long days and short nights. Furthermore, switching the light on in the middle of a long night (a night-break treatment) allows many plants to respond as if they had received two short nights. Examples of these techniques are described below (see ‘Crops grown under protection’).
Irrigation
It is usually necessary to supply water to plants growing under protection as they are shielded from rain, hail and snow. In many situations it is feasible to set up semi-automatic or even fully automatic systems that provide water at fixed intervals or when the plants need it. With plants growing in pots on benches it is possible to supply water by means of capillary matting, or by an ebb and flood system. Water can also be supplied via an overhead spray line, through seepage tubes, or via drippers, and all these systems can be used for plants growing in beds as well as for those standing on benches. It is generally beneficial to use a dilute nutrient solution rather than water alone as this allows water and nutrients to be supplied at the same time.
On commercial nurseries, nutrients are almost always added to the irrigation water, and it is not uncommon to find crops being grown in some form of hydroponics, especially relatively long-lived crops such as tomato (Solanum lycopersicum) and cucumber (Cucumis sativus). These crops can be grown in an inert material such as perlite or rockwool (a synthetic mineral fibre) and the dilute nutrient is supplied via drippers. These drippers are attached to a plastic supply tube, and the flow of dilute nutrient solution can then be triggered by demand. An alternative arrangement is for the plants to be grown in troughs down which flows a thin film of nutrient solution, that is the nutrient film technique (NFT). Under this system, a plant’s need for water is satisfied at all times. The merit of these systems is that they isolate the plants from soil-borne diseases and also reduce water stress.
Control systems
All mechanical and electrical systems need a control system to ensure that best use is made of the resource. An average small greenhouse may utilise thermostats to control heating and perhaps time switches to control irrigation. However, in commercial glasshouses the control of heating and ventilation, as well as irrigation, will almost certainly be exercised by one or more computers. These can be programmed to control the speed with which the temperature changes from night to day, and vice versa, and to produce a given average temperature over some specified period of time. They will also control when the change to night or day temperature should begin as well as the differential between night and day temperatures. They can also control the CO2 concentration in the glasshouse atmosphere, as well as its water vapour content (i.e. humidity), and regulate the time and duration of irrigation. Complete control of this kind is essential to maximise crop yields.
Biological control
It is feasible to use biological agents to control insect and fungal pests within a heated glasshouse (see Chapter 18). One criterion for the introduction of an exotic insect as a biological control agent is that, if it were to escape, it should not be able to survive the cold of winter. The biological agents that are normally used include a variety of parasites and predators as well as fungal and bacterial diseases. Biological control agents have become the standard method of controlling most insect pests of commercial crops, and their use enables tomatoes, for example, to be promoted as having been grown without any pesticide sprays. An added bonus of not using pesticide sprays is that hives of bees can be introduced into glasshouses so that the bees can help with pollination. Experiments have shown that bees are more effective than using other methods for pollinating tomato flowers.
Mechanisation
One advantage of the complete protection afforded by a greenhouse is that it allows mechanisation to be introduced. There is not much scope for doing this in small greenhouses but mechanisation is employed in a host of different ways in large commercial glasshouses. One simple application is using electric motors to operate the ridge ventilators. Mechanisation also allows seeds to be sown very precisely into the cells of multi-celled trays that have been filled with compost by machine. Other machines can move the trays onto mobile benches and these can then be moved into germination chambers. Later, a vision system can be used to detect those cells without a seedling in them and a robot can fill the empty cell with an appropriate seedling taken from a tray of spare plants. If the object is to produce pot plants, other robots can then lift the plugs, complete with seedlings, and plant them into pots of compost before placing the pots onto benches. The benches can then be moved around the glasshouse by remote control while their positions are continually tracked by computer (Fig. 16.9).

Figure 16.9 A modern commercial glasshouse equipped with mobile benches and supplementary lighting. Reproduced by permission of K.E. Cockshull, courtesy of Double H Nurseries, Hampshire.
Special cases
For certain specialised applications, highly insulated structures with opaque walls are required. These ‘growth rooms’ can be used for the reliable and predictable germination of seeds, for the production of mustard and cress, and for the production of mushrooms and other fungi. If light is required, it is usual to install tubular fluorescent lamps over the benches, allowing the benches and their lamps to be arranged in tiers. This arrangement enables a very large number of young plants to be raised in each room, and is particularly well suited to culturing orchid seedlings in glass flasks. The system has the advantage that light can be provided at the same level every day and for any desired duration so that productivity can be controlled and predicted very accurately. One application is in the production of very small, high-value crops, such as ‘micro-leaves’ or ‘baby leaf’ salads.
There is currently much interest in the possibility of using light-emitting diodes (LEDs) for some of the above applications (see ‘Lighting’, above). The advantages of LEDs are that they are very long-lived and the colour range of the light they emit is normally very restricted. This latter quality is useful if either red or blue light only is required to influence a specific developmental response (see Chapters 8 and 9). In addition, plants can be grown quite satisfactorily under the light from combinations of red and blue LEDs, and such combinations are being trialled by commercial growers. At present, white LEDs are unlikely to be used as they are not yet as efficient as other LEDs.
Plant growth regulators
Plant growth regulators (PGRs) are widely used in protected cultivation, mainly by commercial growers. Plant development is controlled by hormones, and most of the synthetic PGRs used in protected cultivation are either identical to a natural hormone or mimic its action. Growth retardants form a different group as they either block the synthesis or the mode of action of a group of natural hormones such as the gibberellins. Some PGRs can be sprayed onto a plant’s foliage, whereas others have to be incorporated into the potting compost or applied as a soil drench.
The auxins form one class of natural hormones (see Chapters 2 and 8), and one of their synthetic equivalents is indole-3-butyric acid (IBA), which encourages root formation. Cell extension is promoted by a variety of natural gibberellins, of which two (GA3 and GA4+7) are widely used in a number of different applications. Cell division is often controlled by cytokinins, of which zeatin is available commercially. Other hormones include abscisic acid (ABA), which inhibits gibberellins in seed germination and is also produced in the roots in response to environmental stress. Cell division and also fruit ripening can be inhibited by ABA in some plants. Another class of hormones is the jasmonates, or ‘wound hormones’, which are released by cells when tissues are attacked, by insects for example, and they seem able to stop further attacks.
Growth retardants inhibit stem growth and so are particularly important in the cultivation of pot plants, which are normally required to be compact. The most widely used retardants are paclobutrazol (e.g. Bonzi®) and daminozide (e.g. B-Nine®), which inhibit steps in the synthesis of gibberellins. Other retardants include the ethylene generator ethephon (e.g. Ethrel®), which also inhibits flowering in some plants. Yet other growth retardants, including ancymidol, chlormequat chloride (e.g. Cycocel®) and 2,4-dichlorobenzyl-tributylphosphonium chloride (Phosfon), are still available but are not now widely used in protected cultivation.
Does protection have disadvantages?
The main disadvantage of protection is that it inevitably causes some loss of light and so a plant’s rate of photosynthesis will be lower than it would otherwise have been. Additional light loss may occur if sand and dust accumulate on the outer surface of the cladding, or if algae and lichens grow on it, or the cladding itself becomes less transparent over time. For most gardeners, the benefits of protection are so great that they are prepared to tolerate at least some loss of light. For the commercial grower, however, the problem is more serious as the tomato, for example, gives 1% less yield for every 1% loss of light. For such crops there is merit in trying to maximise light transmission by regularly washing the outer surface of the protective cladding and by minimising any shading caused by obstructions within the glasshouse.
As protective structures limit air exchange, there is also a real risk that air and tissue temperatures will increase to harmful levels under very sunny conditions. This can be overcome by reducing the amount of radiant energy entering the greenhouse, either by temporarily coating the outside of the greenhouse with some reflective material or by placing opaque objects over part of the roof. When sunny conditions are associated with limited air exchange, the concentration of CO2 in the greenhouse air can be depleted by the photosynthetic activity of the plants and this depletion will limit their growth. A lack of air exchange will also encourage a rise in humidity, which will encourage fungal diseases. Finally, depending upon the source of any added CO2, harmful atmospheric pollutants may be added and accumulate in the greenhouse atmosphere. Almost all of these potential problems can be overcome by taking sensible and appropriate precautions.
Crops grown under protection
Chrysanthemum
Growing chrysanthemums (Chrysanthemum x morifolium) under even very rudimentary protection confers some advantages as the flowers can be damaged by hail and by rain. Chrysanthemums are usually grown in the ground or in pots, and when grown in pots the plants can readily be moved into the protection of a greenhouse, if required. Late-flowering types, especially, are usually moved into a greenhouse in the autumn for protection. Once the plants have flowered and the leaves have started to die, the stems are cut down to within a few inches of the ground and the resulting stools are usually kept outdoors over winter. They will then be exposed to cold, which should assist flower production, but they may need to be protected against extreme cold. New shoots will arise from the base of the stools in spring and these can be removed as cuttings and rooted.
Part of the mystique of growing chrysanthemums is to know when to remove the growing point at the tip of the main shoot; a process called stopping, or pinching, that eliminates the apical dominance of the terminal bud and therefore encourages branching. Different varieties form flower buds at different times, and the object of ‘stopping’ is to do it when there is still time for the new branches to create an adequate length of stem below the flower. Once a flower bud has been formed, it stops the growing point at the tip of the shoot from making any more leaves or stem internodes. The full details of when to ‘stop’ different varieties can be found in books published by the National Chrysanthemum Society. Once flower buds have formed, the next decision is whether to remove the terminal bud (disbudding), which encourages the axillary flowers to develop and create a ‘spray’. Alternatively, removing all the axillary flowers concentrates resources into the developing terminal flower and produces a ‘disbudded standard’ bloom.
Each flower of chrysanthemum is actually a composite inflorescence, or capitulum (see Chapter 3), consisting of many small florets mounted on a receptacle that is surrounded by small, simple leaves called bracts. The central, disc florets are tubular in shape and have stamens and ovaries, whereas the outer, ray florets have what seems to be a single tongue-shaped, highly coloured petal but no stamens.
The natural time of flowering for autumn-flowering chrysanthemums is controlled mainly by day-length. Chrysanthemum leaves are able to detect that the nights are lengthening between July and October, and they respond by producing a message or flowering hormone (see Chapter 9). When this reaches the growing points at the end of each shoot it causes them to initiate flower buds, but their development into open flowers requires that the leaves be exposed to even longer nights. It thus appears that the chrysanthemum has evolved to flower naturally in late autumn as the nights continue to lengthen. The actual length of night that leads to flower initiation must vary between varieties as their flower buds appear at different times. Flower development is evidently not controlled exclusively by night length as the process is also strongly influenced by temperature (see ‘Development’ above).
The chrysanthemum provides one of the best examples of how the control of flowering by day-length has been exploited to enable commercial growers to produce flowers all-year-round (AYR). Stock plants are ‘stopped’ to encourage the production of young vegetative shoots, which are then removed as stem cuttings. The stock plants are now grown in areas with good natural light, such as South Africa, Kenya and the Mediterranean region, and this enables cuttings of good quality to be produced during the UK winter. The stock plants and the cuttings are kept from flowering by switching on night-break lighting around the middle of every night, so that plants respond as if they have received two short nights (see ‘Lighting’ above). The irradiance required is relatively low (c. ≤ 0.5 W m−2) but if given for at least 4 hours around the middle of the night is usually effective. The main requirement is that any period of continuous darkness should not exceed 8 hours in length. Until recently, conventional incandescent lamps were used for night-break lighting, but as these are now being phased out there is a need to find an alternative that will be as effective.
When grown for cut-flower production, the chrysanthemum cuttings are planted in beds in a heated glasshouse if flowers are to be produced to a specific time schedule. They continue to receive a night-break treatment until they have formed sufficient leaves and internodes to create the length of stem that the market requires. The night-break lighting is then turned off and the plants are given long nights (short days) to encourage flower formation. They will receive these naturally in winter but at other times of year a light-tight cover has to be drawn over the plants in the early evening (Fig. 16.10) and removed again in the morning once they have received at least 12 hours of continuous darkness.

Figure 16.10 Chrysanthemum pot plants under a blackout cover to produce long-nights. Photograph by K.E. Cockshull, courtesy of Double H Nurseries, Hampshire.
To produce a flowering pot plant, up to five cuttings are planted in each pot of a soil-less compost. The cuttings are ‘stopped’ when very young to encourage branching, and will start to receive long nights much earlier than with cut-flower production, to keep the stems short. Indeed, as very compact plants are required for pot-plant production, this is generally achieved by applying a growth retardant (see ‘Plant growth regulators’ above).
As well as flower initiation being encouraged by long periods of darkness each night, all varieties also have a mechanism whereby they will eventually initiate a terminal flower bud even when receiving short nights (i.e. long days) and even in continuous light. This ‘autonomous induction’ can be measured in terms of how many leaves the growing point of the shoot forms in long days prior to initiating a flower (the ‘long-day leaf number’). Indeed, it is as though the growing point is moved closer to forming a flower bud each time a leaf is initiated. The long-day leaf numbers of different varieties (Table 16.1) show that whereas some varieties cannot form many leaves before they initiate a flower in long days others, such as ‘May Shoesmith’, are able to form a great number of leaves. An estimate of how long a variety will grow vegetatively can be made from their long-day leaf number and their leaf initiation rate (Table 16.1). Varieties with a low long-day leaf number are more likely to form flowers prematurely while receiving long-day treatment in AYR chrysanthemum production.
Table 16.1 The ‘long-day leaf numbers’ of some chrysanthemum cultivars (i.e. the number of leaves produced by the terminal growing point of different cultivars when grown only in long days). Leaf production on any shoot is eventually stopped by the formation of a terminal flower bud, even in long days. The duration of vegetative growth before a flower bud is initiated has been estimated from the rate of leaf initiation for that cultivar and its long-day leaf number
| Cultivar | Long-day leaf number | Estimated duration of vegetative (number of leaves) growth (days) |
| Bright Golden Anne | 20 | 24 |
| Polaris | 32 | 36 |
| Snowdon | 39 | 49 |
| Robeam | 47 | 66 |
| Orange Westland | 56 | 100 |
| Pink Gin | 81 | 109 |
| May Shoesmith | 94 | 118 |
Data from Langton, F.A., Royce, H.M. & & Cockshull, K.E. (1982) Leaf numbers of chrysanthemum cultivars grown in long days. Tests of Agrochemicals and Cultivars. Annals of Applied Biology, 100 (Suppl. 3), 11011.
Cucumber
Modern cucumber (Cucumis sativus) varieties are generally F1 hybrids (see Chapter 5) and have been selected because they almost always produce only female flowers. Furthermore, they are selected to produce fruit without pollination, that is they do so parthenocarpically. Indeed, any male flowers that appear should be removed as soon as possible, as pollinated fruit are often distorted in shape and of poor quality.
For a commercial grower of cucumber, the propagator will germinate seed in small cubes of rockwool before transplanting them into larger cubes. These are delivered to the grower, who then places them on bags of an inert medium such as rockwool, coir or perlite and allows the plants to root into it. The rooting medium is usually irrigated by drippers. Cucumbers produce very large leaves, so the commercial planting density may be as low as 1 or 1.5 plants per square metre, and the optimum temperature for growth and fruit production is around 26–30°C. Cucumbers used to be grown on straw, which produced CO2 as it decomposed, but today the CO2 is added to the atmosphere to produce a concentration of about 1000 ppm. Cucumbers can be grown as single-stem cordons in what are called ‘high-wire’ systems, or they can be allowed to grow vertically until they reach the required height, at which time the shoot growing point is removed. Branches then grow out but just two are usually retained, trained to hang down and allowed to carry just one fruit at each node. The fruit are cut and marketed when they are about 30 cm long; they have to be shrink-wrapped because they are thin-skinned and lose water rapidly. In commercial production it is usual to replace the crop with fresh plants every few months.
Lettuce, leafy salads and other salad vegetables
Lettuce (Lactuca sativa) is a typical salad vegetable. The range of types that can be grown in the garden is considerable but the types most commonly grown in the field include ‘Lollo Rosso’, ‘Romaine’ and ‘Iceberg’. Some of these form hearts but all should be cut just above ground level rather than being pulled from the ground. Many are also grown as ‘baby leaf’ vegetables as their young leaves can be picked off by hand or cut off by machine for use in salads. Other species that are grown as salad vegetables include radicchio (Cichorium intybus), radish (Raphanus sativus) and spinach (Spinacia oleracea), whereas pak choi (Brassica rapa) is grown both as a salad vegetable and as a baby leaf. Most plants of the above salads are hardy and suited to growing in a garden, but their quality and speed of production will be enhanced by protecting them under a simple cover such as horticultural fleece. They will mature even earlier if grown under cloches or in tunnels, or even in unheated greenhouses, but they are prone to form flowers and ‘bolt’ if the conditions are too warm. A recent development is the production of ‘micro-leaf’ vegetables, which are harvested at a very young stage and produced in highly automated systems.
Poinsettia
Very few gardeners attempt to grow the poinsettia (Euphorbia pulcherrima), perhaps because its requirements are quite demanding. The flowers are relatively insignificant and form cyathia, that is collections of reduced male and female flowers typical of the family Euphorbiaceae. The poinsettia is grown for the numerous highly coloured bracts and leaves that are formed immediately below these flowers. Cyathia are initiated in response to lengthening nights and so the production system has many similarities with that of the chrysanthemum, the main difference being that the poinsettia is almost always produced only as a pot plant and just for Christmas. For that reason, most commercially grown varieties have bright red bracts, although varieties with pink or cream bracts are available (Fig. 16.11).

Figure 16.11 Poinsettias in a trial showing the range of colours available.
Stock plants of poinsettia are maintained by commercial propagators, who supply growers with rooted cuttings in August. Three cuttings or fewer are then planted per pot of soil-less compost and ‘pinched’ about a week after planting, to encourage branching. Night-break lighting is not usually required as the natural day-length through this period is long enough to stop the plants flowering. However, to be sure of having plants in full flower by Christmas they are covered with a light-tight, blackout material each evening from the beginning of October, the object being to give the plants at least 13 hours of continuous darkness each night. The poinsettia is very sensitive to light, and there have been many reports where flowering has been delayed by ‘light spill’ from adjacent street lighting or from lighting used on adjacent crops. Flowering is also delayed if the night temperature falls below 18°C, which may be another reason why few gardeners try to grow them. Some commercial growers lower the night temperature once the coloured bracts are fully developed as this helps to keep the plants in a state of readiness for marketing; also it conditions them to the sort of environment they are likely to encounter in the home.
Poinsettias make remarkably long-lived house plants when kept in a centrally heated home but they will eventually lose their red bracts. Hence the perennial question ‘How can I get them to form red bracts again?’ The answer is first to rejuvenate them by cutting off the old bracts and flowers, which will encourage fresh branches to grow out and produce green leaves. By September, however, it will be necessary to cover them with a light-tight, blackout cover so that they get at least 13 hours of continuous darkness each night. This is still necessary even in the home as lights are often switched on, albeit briefly, in most rooms at some time in the evening. If ‘blacking out’ sounds too tedious, the plants can be left to initiate flowers in response to the naturally lengthening nights of late autumn but they are unlikely to have red bracts in time for Christmas.
Many modern poinsettia cultivars have been bred to produce compact plants but older cultivars have to be treated with a growth retardant (see ‘Plant growth regulators’ above). An alternative to applying a growth retardant is to give the plants a DROP treatment (see ‘Stem growth’ above). In this treatment, the temperature is dropped at dawn and kept there for one or more hours before it is allowed to rise to its daytime setting.
Strawberry
The strawberry (Fragaria x ananassa) has been grown outdoors both by gardeners and commercial growers for many years but its cultivation under protection has recently become an extremely important commercial activity. The main factor that has contributed to this is the use of tunnels clad with polythene, popularly known as polytunnels (see Fig. 16.12). These protect the plants and their fruit from rain, which is important because the presence of free water encourages the germination of spores of the grey mould of fruit (Botrytis cinerea). The protection also enables pickers to harvest fruit in all weathers, and the warmer conditions allow fruit to develop more quickly and be harvested earlier. The use of polytunnels has enabled almost 90% of the harvested fruit to be classed as grade 1.

Figure 16.12 A ‘walk-in’ polytunnel for strawberry production. Reproduced by permission of Bellis Brothers, Wrexham.
Polytunnels can be of various sizes and heights but it is very convenient to use tunnels that are high enough to walk into comfortably. Such tunnels can be free-standing (Fig. 16.12) or they can be joined together at their gutters to produce multi-tunnel arrays with a skirt around the periphery. Within each polytunnel, early crops can receive additional protection from smaller polytunnels that straddle individual rows, or they can be covered with horticultural fleece. Various types of polythene film are available as cladding, but one that has considerable potential is a film that blocks the transmission of the UV light that is required for spore formation by Botrytis cinerea. Protection also enables the grower to control pests and diseases by biological methods so that fruit can be produced with minimal use of pesticides. The development of protected cropping of strawberry has also benefited from an improved understanding of the control of flowering and fruiting as well as from the breeding of new varieties that are better suited to growing under protection.
In general, strawberry varieties fall into one of two, possibly three, groups depending upon the environmental factors that control flower formation and hence the time of year at which flowering and fruiting occur. The traditional ‘June-bearing’ varieties that are grown in the garden in the United Kingdom initiate flowers in response to long nights (short days) and to temperatures lower than 15°C. For these varieties, it is the shortening day-length of autumn that is the main control of flower formation and the main cause of the onset of dormancy. The low temperatures of winter then slowly break this dormancy and allow the flowers that were initiated in the autumn to develop and open. Although it is called the June-bearing group, it contains a spread of flowering times as the group comprises ‘early’, ‘mid-season’ and ‘late-flowering’ varieties.
A second group of strawberries contains the ‘day-neutral’ varieties that form flowers in any day-length provided that the temperature is between about 4 and 30°C. More of these are now being bred, and although they are sometimes referred to as ‘everbearing’ many people regard everbearing strawberries as yet a third group.
The flower of strawberry is typical of the family Rosaceae and consists of five sepals and five petals surrounding a large number of stamens, which in turn surround a large number of ovaries. All of these organs are born on a receptacle and it is this that swells and forms the strawberry fruit once the ovaries are fertilised. Many modern varieties can be self-pollinated but fruit yields and fruit shape are usually better if the varieties are pollinated by bees or bumblebees. Each ovary develops into a one-seeded fruit or achene and they are all carried on the external surface of the receptacle. Commercially, the fruit are still picked by hand and picked with the calyx present.
In addition to encouraging flower development, the warm temperatures of spring also encourage stolons to grow out from the leaf axils and form ‘runners’ that can either be rooted near the mother plants or cut off and rooted elsewhere. The young plants are allowed to initiate flowers as the day-length shortens in autumn, after which they are lifted and transferred to a cold-store. The overwintered, cold-stored plants with flowers already present are bare rooted and can be planted in polytunnels in early spring to produce fruit ready for picking about 60 days later. This technique provides another means by which the strawberry season can be extended, especially if the plants are taken out of the cold-store at intervals.
Certified virus-free runners are planted with the crown of the plant just above the soil surface. They can be planted directly into moist well-prepared soil, but it is more usual to plant them through a film-plastic covering, especially for outdoor crops, as the plastic protects the developing fruit from contact with the soil. Many growers prefer to shape the soil into ridges and place the plants at the top of the ridge.
Strawberry crops growing in polytunnels are commonly irrigated by trickle irrigation, which can be laid under any film-plastic mulch. Other systems include growing the plants in raised troughs, which has the advantage that the fruit hang down over the sides of the trough and are then more easily picked when ripe. This system is frequently referred to as ‘table-top’ production (Fig. 16.13). The trough can be filled with coir, perlite, vermiculite, pebbles, expanded clay pellets (e.g. Leca®), a soil-less compost or even rockwool, all of which allow the plants to be grown in a hydroponic system. In gardens, strawberry plants are usually grown in the soil, where they can be protected by mulching with straw, which also helps to keep the fruit from making contact with the soil. This can also be achieved by using tall pots with planting holes in the side that allow the fruit to hang in the air.

Figure 16.13 ‘Table-top’ strawberry production in a polytunnel. Reproduced by permission of Bellis Brothers, Wrexham.
Tomato
The tomato (Solanum lycopersicum) is one of the most popular vegetables grown by gardeners. The advantages of growing them in greenhouses are that the fruit will be ready to pick well before those on plants grown outdoors, and there is less chance of the plants being killed by frost. Tomatoes can be grown in border soil, in pots, or in grow-bags filled with a sustainable rooting medium such as bark, coir or perlite. Gardeners in the United Kingdom are discouraged from using peat because its use is no longer regarded as sustainable (but see Chapters 20–22). Some tomato varieties (indeterminate types) can be left to branch and form a mass of leaves, stems and fruits whereas others perform better if all the side-shoots are removed at the earliest opportunity, so leaving a single main shoot (determinate types).
The flowers of tomato are formed by the growing point at the tip of the main shoot, and it creates a cluster of flowers on a leafless stalk or truss. Almost as soon as flower initiation begins a side shoot grows out from the axil of the uppermost primordial leaf and overgrows the truss to become the main shoot. As a result, the flowering truss later appears to grow out from the side of the stem. Once the new main shoot has produced three leaves (sometimes fewer), it too will begin to initiate an inflorescence and the sequence is repeated. The number of flowers formed by each truss depends largely on the variety, but the number that become fruits depends on various factors. Whole flowers may abort if the light is very poor and the temperature too warm, but inadequate pollination may also be a problem for the stamens do not readily release pollen when the light is poor. At such times, the flowers should be gently shaken to encourage pollen to fall onto the stigmas. The shaking is usually done commercially by mechanically shaking the truss or even the whole plant. However, because chemical sprays are now used so rarely in commercial fruit production, it is usually possible to place hives of bumblebees in the glasshouse so that the bumblebees can pollinate the flowers.
The fruit of tomato is divided into a number of chambers (locules) in each of which there are numerous seeds immersed in a jelly-like material. The size and shape of the fruits vary considerably according to variety. The smallest fruits are formed by the cocktail and cherry types, followed in size by plum tomatoes, then the traditional round tomatoes (now called ‘classic’). The largest fruits are found on beefsteak tomatoes, which are mainly used for slicing. The main physiological problem for all varieties is ‘blossom-end rot’, when tissues at the blossom end of the fruit die and turn black due to a localised lack of calcium.
Tomato plants are usually raised from seed by specialist propagators and may be grafted onto seedlings of a rootstock variety growing in small cubes of rockwool, especially if they are to be supplied to a commercial grower. The rockwool cubes are then placed onto bigger slabs of rockwool so that the roots of the young plants can grow into it. Each slab is wrapped in a film-plastic sleeve, which is slit along its sides to allow excess liquid to escape when the slabs are watered. The slabs can be laid out in rows on the floor of the glasshouse or suspended in gutters above the floor and are set either side of a path that has a looped heating pipe running on either side of it (Fig. 16.14). The glasshouse floor is covered with a white polythene sheet, which isolates the slabs from the soil and also helps to reflect light back up towards the crop.

Figure 16.14 The layout of heating pipes and rockwool slabs for growing tomatoes in a commercial glasshouse. Reproduced by permission of G. Hayman, courtesy of Cornerways Nursery, Norfolk.
In commercial production, the young plants will probably be planted in the glasshouse in December or January and are kept in place through to the following October or November. The tomatoes are grown as cordons, that is where all the side shoots are removed to leave just one main shoot, which is supported by twisting a vertical string around it. The string is wound onto a bobbin which is attached to a hook suspended from a wire running along the side of the path at eaves height. As the plant grows, some string can be let out from the bobbin so that the stem can be lowered towards the floor. The hook and its bobbin are then moved along the wire, as required, with all the plants on one side of the row being moved in the same direction. When they reach the end of the row, their hooks and bobbins are taken around the end and attached to the wire on the other side of the row so that the plants then go back along the row.
A conventional planting density in January might be approximately two plants per square metre; this can be increased to three plants per square metre by allowing every other plant to keep one side shoot as the light improves. The plants are supplied with a dilute nutrient solution at a frequency related to the amount of solar radiation received; the solution is delivered to the plants through pipes ending in drippers mounted on the rockwool slabs. The glasshouse air temperature is kept at about 18°C.
As mentioned earlier, growers aim to enrich the CO2 concentration to 1000 ppm while the glasshouse ventilators are closed, but once they are opened the control level may be lowered to c. 400 ppm CO2. Fruit production is closely related to the amount of light received by the foliage each day – i.e. the daily light integral – which is the product of the duration and the intensity of sunlight. Because light is so important in determining fruit yield, glass is the most commonly used cladding material due to its high light transmission. Fruit that are destined for market have to be picked as they begin to change from yellow to red, but the gardener can leave them on the plants until the fruit are ripe. Increasingly this is also done commercially, and ‘vine-ripened’ fruit are now available in most retail outlets. Pest control in commercial production principally employs biological control based upon a mix of predators and parasites of the pests.
Pot plants
Almost any tender plant can be grown as a pot plant in a heated greenhouse, including many orchids, and there is now a remarkable range of flowering and foliage pot plants available for sale in garden centres. In essence, the only restrictions on what can be grown in a heated greenhouse are the temperature the heating system is able to sustain, regardless of external conditions, and the price the owner is prepared to pay for the pleasure of growing very tender, conservatory or hot-house plants.
Conclusions
Growing plants under protection confers many advantages, including protection from frost, rain, hail, sleet and snow, which enables plants to be overwintered safely as well as enabling them to produce flowers or fruits of better quality, earlier than unprotected plants. Although the heated greenhouse is probably the ultimate form of protection for growing plants in natural light rather than under lamps in a growth room, much can be achieved with simpler arrangements such as unheated tunnels, cloches and mulches. A wide range of crops are now grown under protection, and many of the methods employed by commercial growers can be adapted for use by gardeners. A basic understanding of how plants respond to changes in their environment enables growers and gardeners to avoid many potential problems.
Further reading
- Bakker, J.C., Bot, G.P.A., Challa, H. & Van de Groot, H.J. (1995) Greenhouse Climate Control: An Integrated Approach. Wageningen Academic Publishers, Wageningen, The Netherlands.
- Benton Jones Jr, J. (2008) Tomato Plant Culture, 2nd edn. CRC Press, Boca Raton, FL.
- Cockshull, K.E. (1985) Chrysanthemum morifolium. In: Handbook of Flowering, Halevy, A.H. (ed.), Vol. II, pp. 238–57. CRC Press, Boca Raton, FL.
- Cockshull, K.E. (2002) Greenhouse food production (agriculture intensive). In: Encyclopedia of Global Environmental Change, Munn, T. (ed.), Vol. 3, pp. 352–6. John Wiley & Sons, Ltd., Chichester.
- Cockshull, K.E. (2003) Production systems and agronomy – horticultural crops, protected. In: Encyclopedia of Applied Plant Sciences, Thomas, B., Murphy, D. & Murray, B. (eds), Vol. 2, pp. 917–24. Academic Press.
- Cockshull, K.E., Langton, F.A. & Lumsden, P.J. (eds) (1997) II Workshop on Environmental Regulation of Plant Morphogenesis. Acta Horticulturae, 435.
- Colborn, N. & Perry, C. (2007) A Garden under Glass: The Glasshouse, RHS Garden, Wisley. Royal Horticultural Society, UK.
- Frediani K. (2011) High rise food. The Horticulturist, 20(4), 18–20.
- Grange, R.I. & Hand, D.W. (1987). A review of the effects of atmospheric humidity on the growth of horticultural crops. Journal of Horticultural Science, 62, 125–34.
- Grant, F. (2013) Glasshouses. Shire Publications Ltd., Oxford.
- Hanan, J.J. (1998) Greenhouses: Advanced Technology for Protected Horticulture. CRC Press, Boca Raton, FL.
- Hand, D.W. (1982) CO2 enrichment, the benefits and problems. Scientific Horticulture, 33, 14–43.
- Hix, J. (2005) The Glasshouse, 2nd edn. Phaidon Press, London.
- Holder, R. & Cockshull, K.E. (1990) The effects of humidity on the growth and yield of glasshouse tomatoes. Journal of Horticultural Science, 65, 31–9.
- Langton, A. & Cockshull, K. (2013) All change in the glasshouse. The Horticulturist, 22(4), 17–20.
- Larson, R.A. (ed.) (1992) Introduction to Floriculture. Academic Press, San Diego, CA.
- Woods, M. & Warren, A.S. (1988) Glass Houses: History of Greenhouses, Conservatories and Orangeries. Aurum Press, London.
Authors and affiliations
Written for the third edition by Ken Cockshull, Associate Fellow, University of Warwick (to replace Chapter 10 in the first edition/Chapter 14 in the second edition).