Abbreviations: AP-1: activator protein-1, CaP: pancreas cancer, CFRs: cyclic reductions in coronary flow, COX: cyclo-oxygenase, CVD: cardiovascular disease, EGCG: epigallocatechin gallate, γ-GCS: gamma-glutamylcysteine synthetase, GSH: glutathione, GST: glutathione S-transferases, HCC: hepatocellular carcinoma, HDL-C: high-density lipoprotein-cholesterol, I3C: indole-3-carbinol, LDL-C: low-density lipoprotein-cholesterol, MAPK: mitogen-activated protein kinases, NF-κB: nuclear factor-kappaB, NO: nitric oxide, NOS: nitric oxide synthase, NQO1: NAD(P)H:quinone oxidoreductase, PK: protein kinase, ROS: reactive oxygen species, RSV: resveratrol, SOD: superoxide dismutase, TG: triglycerides, UV: ultraviolet, VEGF: vascular endothelial growth factor.
Since early in the history of medicine, an association between phytochemicals and disease has persisted. Galen of Pegamon (129–199 AD), a Greek physician and follower of Hippocrates’ teachings was said to have prescribed various foods, including peeled barley, and various vegetables for the treatment of cancer. The beneficial effects of fruits and vegetables have been attributed to, among other things, the high content of bioactive compounds that are non-nutrient constituents commonly present in food (Siddiqui et al., 2009). Natural dietary components, obtained from several fruits, vegetables, nuts and spices have drawn a considerable amount of attention due to their demonstrated ability to partially prevent cardiovascular disease (CVD) and suppress carcinogenesis in animal models, or delay cancer formation in humans. It has been ascribed in part to antioxidants in plant bionutrients inactivating reactive oxygen species (ROS) involved in initiation or progression of these diseases (Duthie et al., 2006). It is estimated that approximately 8000 phytochemicals are present in whole foods, and there are quite possibly many more (Liu, 2004). These compounds with much more complex scope, interaction, and magnitude may act on different targets with different mechanisms of action. Over the centuries, no fewer than 3000 plant species have been used for chemotherapy and chemoprevention. The World Health Organization (WHO) has estimated that approximately 80% of the population in some Asian and African countries still depends on complementary and traditional medicine for the prevention and treatment of diseases, most of which involves the use of plant extracts (Naczk and Shahidi, 2006).
Most natural products can be classified into three major groups: nitrogen-containing compounds, terpenoids and phenolic compounds. The major class of nitrogen-containing compounds is represented by alkaloids, synthesised principally from aspartic acid, tryptophan, arginine and tyrosine. More than 10 000 different alkaloids have been discovered in species from over 300 plant families. These compounds protect plants from a variety of herbivorous animals, and many possess pharmacologically important activities (Zenk and Juenger, 2007). Terpenoids (also called terpenes) are a large and diverse class of naturally occurring organic chemicals derived from five-carbonisopreneunits assembled and modified in thousands of ways. These chemically-different compounds are grouped in a unique class, since they are all derived from acetyl-CoA. Many plant terpenoids are toxins and feeding deterrents to herbivores or are attractants of various sorts. Plant terpenoids are used extensively for their aromatic qualities. They play a role in traditional herbal remedies and are under investigation for antibacterial, anti-neoplastic and other pharmaceutical functions. Phenolic compounds are widely distributed in the plant kingdom. Plant tissues may contain up to several grams per kilogram. Flavonoids are the most abundant, commonly known for their antioxidant activity and for their use in human diet, due to their widespread distribution, and their relatively low toxicity, compared to other active plant compounds, i.e. alkaloids (Le Marchand, 2002). Among plant chemopreventive agents we will highlight indoles, catechins, vitamins, isoflavonoids (silymarin) and phenols (resveratrol, and curcumin) (Gerhäuser et al., 2003).
Many phytochemicals of differing chemical structure have medicinal properties. They activate cytoprotective enzymes and inhibit DNA damage to block initiation in healthy cells, or modulate cell signalling to eliminate unhealthy cells at later stages in the carcinogenic process. In vitro results for several well-studied compounds indicate that each can affect many aspects of cell biochemistry (Manson et al., 2007). Many phytochemicals are poorly bioavailable and evidence suggests that combinations may be more effective than single agents. There may also be advantages in combining them with chemo- or radio-therapies (Manson et al., 2007). In this chapter we deal with the use and efficacy of phytochemicals as pharmaceuticals, i.e. as chemicals intended for the cure and treatment of disease. In addition, we will outline phytochemicals that have progressed to be used as therapeutic drugs. Finally, we will analalyse some active component of phytochemicals that has served as the basis for drug development.
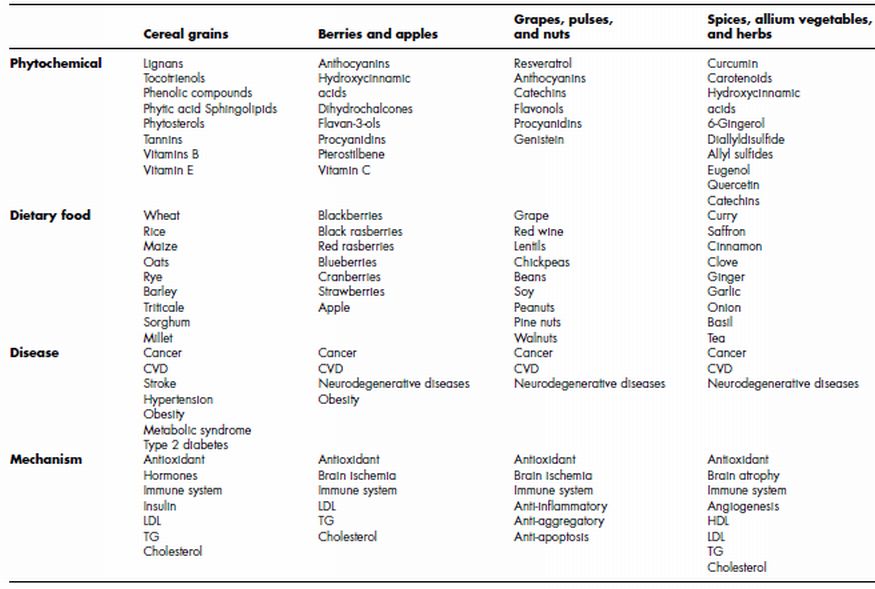
Nowadays, more than 600 functional non-nutrient food factors in vegetables and fruits are considered to be effective for health promotion and medicinal properties (Table 4.1). The optimal intake of various phytochemicals per capita has been calculated as more than 10 micromole per day; such as catechin, isoflavones, isothiocyanate, ferulic acid, quercetin, cinnamic acid and chlorogenic acid (Watanabe et al., 2004). Epidemiological studies find that whole grain intake is protective against cancer, CVD, diabetes and obesity. Whole grains are rich in nutrients and antioxidant phytochemicals with known health benefits. Published whole grain feeding studies report improvements in biomarkers with whole grain consumption, such as blood-lipid improvement, and antioxidant protection (Slavin et al., 2004). The major cereal grains include wheat, rice and maize, with others as minor grains (Table 4.1). Buckwheat, wild rice and amaranth are not botanically true grains but are typically associated with the grain family due to their similar composition (Slavin et al., 2003). Components in whole grains associated with improved health status include dietary fibre, starch, unsaturated fatty acids, minerals, phytochemicals and enzyme inhibitors (Table 4.1). In the grain-refining process the bran is removed, resulting in the loss of dietary fibre, vitamins, minerals, lignans, phyto-oestrogens, phenolic compounds and phytic acid (Slavin et al., 2004). Antinutrients found in grains include digestive enzyme (protease and amylase) inhibitors, phytic acid, haemagglutinins and phenolics and tannins. Protease inhibitors, phytic acid, phenolics and saponins have been shown to reduce the risk of cancer of the colon and breast in animals. Phytic acid, lectins, phenolics, amylase inhibitors and saponins have also been shown to lower plasma glucose, insulin and/or plasma cholesterol and triacylglycerols (Slavin et al., 2003). Phytic acid forms chelates with various metals, suppressing damaging Fe-catalysed redox reactions (Slavin et al., 2004). Hormonally active compounds called lignans may protect against hormonally mediated diseases (Adlercreutz et al., 1997). Lignans are compounds processing a 2,3-dibenzylbutane structure and exist as minor constituents of many plants where they form the building blocks for the formation of lignin in the plant cell wall. The plant lignans secoisolariciresinol and matairesinol are converted by human gut bacteria to the mammalian lignans enterolactone and enterodiol (Slavin et al., 2003). Plant sterols and stanols are found in oilseeds, grains, nuts and legumes. These compounds are known to reduce serum cholesterol (Yankah and Jones, 2001). It is believed that phytosterols inhibit dietary and biliary cholesterol absorption from the small intestine. Phytosterols displace cholesterol from micelles, which reduces cholesterol absorption and increases its excretion (Hallikainen et al., 2000).
Table 4.1 Medicinal properties of dietary foods and their phytochemicals
Whole grain intake is associated with reduced risk of chronic disease. Specifically, there is a decreased risk of obesity, coronary heart disease, hypertension, stroke, metabolic syndrome, type 2 diabetes and some cancers observed among the highest whole grain eaters compared with those eating little or no whole grains (Jones et al., 2008). Additional epidemiological studies have associated consumption of whole grains and whole grain products with reduced incidence of chronic diseases such as CVD, diabetes and cancer (Adom et al., 2005). The health beneficial phytochemicals of wheat are distributed as free, soluble-conjugated and bound forms in the endosperm, germ and bran fractions of whole grain (Adom and Liu, 2002). Health benefits of grains have been attributed in part to the unique phytochemical content and distribution of grains. Grain phytochemicals also include derivatives of benzoic and cinnamic acids, anthocyanidins, quinones, flavonols, chalcones, amino phenolics compounds, tocopherols and carotenoids (Adom et al., 2005). Grain phytochemicals exert their health benefits through multifactorial physiologic mechanisms, including antioxidant activity, mediation of hormones, enhancement of the immune system and facilitation of substance transit through the digestive tract, butyric acid production in the colon and absorption and/or dilution of substances in the gut (Adom and Liu, 2002). The bran/germ fraction of whole wheat may therefore impart greater health benefits when consumed as part of a diet and thus help reduce the risk of chronic diseases (Thompson et al., 1994). Nonetheless, the endosperm fraction also makes some significant contributions to the overall health benefits as outlined here(Adom et al., 2005).
Phenolic compounds fall into two major categories: phenolic acids and flavonoids. The phenolic acids are benzoic or cinnamic acid derivatives, whereas the flavonoids are largely tannins and anthocyanins (Dykes et al., 2006). In comparison with sorghum, other cereal brans examinated, such as oat, rice and wheat, had low phenolic contents and low antioxidant potential (Farrar et al., 2008). There are hundreds of phytochemical components in soybeans and soy-based foods. In recent years, accumulating evidence has suggested that the isoflavones or soy proteins stripped of phytochemicals only reflect certain aspects of health effects associated with soy consumption. Other phytochemicals, either alone or in combination with isoflavones or soy protein, may be involved in the health effects of soy (Kang et al., 2010). Polyphenols comprise a wide variety of compounds, divided into several classes (i.e. hydroxybenzoic acids, hydroxycinnamic acids, anthocyanins, proanthocyanindins, flavonols, flavones, flavanols, flavanones, isoflavones, stilbenes and lignans), that occur in fruits and vegetables, wine and tea, chocolate and other cocoa products (Manach et al., 2004). Epidemiological studies showed that increased intake of polyphenols were associated with reduced risk of CVD, cancer and neurodegenerative disorders. Several polyphenols have been demonstrated to have clear antioxidant properties in vitro as they can act as chain breakers or radical scavengers depending on their chemical structures, which also influence their antioxidant power (Rice-Evans, 2001). A hierarchy has been established for the different polyphenolic compounds within each class on the basis of their capability to protect lipids, proteins or DNA against oxidative injury (Heijnen et al., 2002). This concept, however, appears now to be a simplistic way to conceive their activity (Masella et al., 2005). First of all, pro-oxidant effects of polyphenols have also been described to have opposite effects on basic cell physiological processes (Elbling et al., 2005): for example, if as antioxidants they improve cell survival, as pro-oxidants they may indeed induce apoptosis, cell death and block cell proliferation (Lambert et al., 2005). It should be noted that intracellular redox status, which is influenced by antioxidants, can regulate different transcription factors, which in turn regulate various cell activities (Kwon et al., 2003).
Recent advances have been made in our scientific understanding of how berries promote human health and prevent chronic illnesses (Table 4.1). Berry bioactives encompass a wide diversity of phytochemicals ranging from fat-soluble/lipophilic to water-soluble/hydrophilic compounds (Seeram et al., 2010). Long-term feeding of blueberries to rats hindered and even reversed the onset of age-related neurologic dysfunctions, such as a decline in neuronal signal transduction, and cognitive, behavioral and motor deficits. In addition, Stoner and coworkers showed that supplementation with black raspberries in the diet reduced the multiplicity and incidence of esophageal tumours in N-nitrosomethylbenzylamine-treated rats (Stoner et al., 1999).
The tocopherols (α-, β-, γ- and δ-tocopherol) and resveratrol (RSV) are phytochemicals with alleged beneficial effects against atherosclerosis, vascular diseases and different cancers (Bishayee et al., 2010). Although they both can act as antioxidants, they also modulate signal transduction and gene expression by non-antioxidant mechanisms (Reiter et al., 2007). Apples are widely and commonly consumed and are one of the main contributors of phytochemicals in the human diet (Table 4.1), making them the largest source of dietary phenolics (Yang and Liu, 2009). It is believed that chemotherapeutic combination approaches have been used to reduce drug toxicity, to delay the development of cancer cells, and to reach a greater effect than with one active drug alone. Antioxidant synergism has been observed with different compounds such as vitamins E and C, vitamin E and β-carotene, catechin and malvidin 3-glucoside, flavonoids and urate, and tea polyphenols and vitamin E (Yang and Liu, 2009). The phytochemicals in fruits may act independently or in combination as anti-cancer agents. The additive and synergistic effects of phytochemicals in fruits may be responsible for their potent anti-cancer activities, and the benefit of a diet rich in fruits is attributed to the complex mixture of phytochemicals present in whole foods.
Among spices, saffron displayed the highest antioxidant capacity, whereas among dried fruits, prunes exhibited the highest value. Among cereal products, whole meal buckwheat and wheat bran had the greatest total antioxidant capacity. Among pulses and nuts, broad beans, lentils and walnuts had the highest antioxidant capacity, whereas chickpeas, pine nuts and peanuts were less effective. The contribution of bound phytochemicals to the overall antioxidant capacity was relevant in cereals as well as in nuts and pulses (Pellegrini et al., 2006). Of note, the polyphenol curcumin (1,7-bis(4-hydroxy-3-methoxyphenyl)-1,6-heptadiene-3,5-dione), a natural yellow pigment extracted from the rhizome of the turmeric plant Curcuma longa (von Metzler et al., 2009), was shown to inhibit the activation of the transcriptional factor nuclear factor-kappaB (NF-κB), and to inhibit the proteasome-ubiquitin system (Bharti et al., 2004). Curcumin modifies the invasive potential of breast cancer cells (Squires et al., 2003). Another polyphenol, (–)-epigallocatechin-3-gallate (EGCG), was found to inhibit neovascularisation in the chick chorioallantoic membrane assay and when given in drinking water could significantly suppress VEGF (vascular endothelial growth factor)-induced corneal neovascularisation (Manson et al., 2007). Such results suggest that EGCG may be a useful inhibitor of angiogenesis in vivo (Gilbert and Liu, 2010). A number of phytochemicals also affect expression of cadherins, catenins and matrix metalloproteinases (Manson et al., 2007). Also of increasing importance is the investigation of combinations of phytochemicals or their use in conjunction with other therapies, to increase efficacy or decrease unwanted side effects (Howells et al., 2007). It has been shown in breast cell lines that indole-3-carbinol (I3C) exhibits enhanced efficacy in combination with Src or EGFR kinase inhibitors, and in vivo I3C prevented the hepatotoxicity of trabectidin (ET743), an experimental antitumour drug with promising activity in sarcoma, breast and ovarian carcinomas, without compromising antitumour efficacy (Manson et al., 2007). Curcumin enhances the efficacy of oxaliplatin in both p53-positive and p53mutant colon cancer cells (Howells et al., 2007). However, caution is required, since it has also been reported to compromise the efficacy of some chemotherapeutic drugs in human breast cancer models (Somasundaram et al., 2002). Additionally, using a human osteoclast system, curcumin abrogated both osteoclast differentiation and bone resorbing activities, preventing the IκB phosphorylation (von Metzler, 2009).
Antioxidants work effectively as disease preventing species. The three major types of ROS are superoxide anion radical (O2•-), constitutively present in cells because of leakage from the respiratory chain in mitochondria, hydrogen peroxide (H2O2), resulting from the dismutation of O2•- or directly from the action of oxidase enzymes, and hydroxyl radical (•OH), a highly reactive species that can modify purine and pyrimidine bases and cause strand breaks that result in oxidatively damaged DNA (Matés et al., 2010). Free radical compounds result from normal metabolic activity as well as from the diet and environment (Matés et al., 2002), contributing to general inflammatory response and tissue damage (Matés et al., 2008). Antioxidants protect DNA from oxidative damage and mutation, leading to cancer (Matés et al., 2006). Antioxidants are considered as the most promising chemopreventive agents against various human cancers (Matés, 2000). However, some antioxidants play paradoxical roles, acting as double-edged swords. A primary property of effective and acceptable chemopreventive agents should be freedom from toxic effects in population (Kawanishi et al., 2005). In spite of identification, use of effective cancer chemopreventive agents has become an important issue in public health-related research; miscarriage of the intervention by some antioxidants makes necessary the evaluation of safety before recommending use of antioxidant supplements for chemoprevention (Calabrese et al., 2010).
A number of epidemiological studies initially indicated utility of antioxidants in disease prevention, particularly for CVD and cancer. Regardless, recent conflicting results from intervention trials have identified negative consequences associated with antioxidant supplement use and a presumed reduction in ROS (Seifried et al., 2007). This apparent conundrum of antioxidant effects on ROS has recently been examined in light of molecular evidence for the role(s) of ROS in development and progression of cancer and CVD, especially since there has been a flurry of studies potentially linking some antioxidants with increased mortality and CVD (Seifried et al., 2006). Alternatively, compounds in a plant-based diet may increase the capacity of endogenous antioxidant defenses and modulate the cellular redox state. Changes in the cellular redox state, conveying physiologic stimuli through regulation of signaling pathways, may have wide-ranging consequences for cellular growth and differentiation (Haddad et al., 2002). In addition, it has been well documented that phytochemicals modulate protein kinase (PK) activities, serve as ligands for transcription factors and modulate protease activities (Moskaug, 2005).
Polyphenols are among the most abundant phytochemicals in human food items and, of these, flavonoids are probably the most deeply studied. Low concentrations of flavonoids stimulated transcription of a critical gene for glutathione (GSH) synthesis in cells. Both onion extracts and pure flavonoids transactivated human gamma-glutamylcysteine synthetase (γ-GCS) through antioxidant response elements in the promoter in both COS-1 cells and HepG2 cells, with quercetin being the most potent flavonoid. Structurally similar flavonoids were not as potent; myricetin, with only one hydroxyl group more than quercetin, was inactive, which emphasises the apparent specificity of human γ-GCS induction (Myhrstad et al., 2002). In vivo feeding experiments with polyphenol-rich diets revealed large differences in human γ-GCS promoter activity responses among individual animals. Some animals responded and some did not. One possible explanation for this phenomenon may be related to differences in bacterial populations in the gut microbial flora influencing the extent of enzymatic hydrolysis of polyphenol conjugates (Scalbert et al., 2000). On the other hand, flavonoid antioxidant scavenging of free radicals often involves formation of a radical of the flavonoid itself. Quercetin is oxidised to a quinone when serving as an antioxidant, and Boots et al. (2003) showed that such quinones react with thiols. Therefore, it could be speculated that free radical-oxidised quercetin reacts with thiols in Keap1, the key regulatory protein in transcriptional regulation of antioxidant-responsive genes through Nrf2. Quercetin and myricetin are known to auto-oxidise at physiologic pH, and subsequent reduction of glutathione concentrations can possibly explain transcriptional up-regulation of both γ-GCS subunits (Tian et al., 1997).
Although the redox potentials of most flavonoid radicals are lower than those of O2•- and peroxyl radicals (ROO•-) (Moskaug, 2005), the effectiveness of the radicals in generating lipid peroxidation, DNA adducts and mutations may still be significant in disease development (Skibola et al., 2000). Also of concern is the observation that some flavonoids inhibit enzymes (such as topoisomerases) involved in DNA structure and replication, and it has been suggested that high intake of flavonoids predisposes subjects to the development of certain childhood leukemias (Strick et al., 2000). Flavonoid supplementation as a general recommendation to increase cellular GSH concentrations may also be troublesome, because glutathione has a major role in overall redox regulation of cell functions and is not suitable as a therapeutic target for substances that alter cellular concentrations by orders of magnitudes (Moskaug, 2005). Interesting results add modulation of intracellular GSH concentrations to the list of possible disease-preventing effects of polyphenols, with the implication that they modulate GSH-dependent cellular processes, such as detoxification of xenobiotics, glutathionylation of proteins and regulation of redox switching of protein functions in major cellular processes (Carlsen et al., 2003).
Recent findings suggest that several heavily studied phytochemicals exhibit biphasic dose responses on cells with low doses activating signaling pathways that result in increased expression of genes encoding cytoprotective proteins including antioxidant enzymes, protein chaperones, growth factors and mitochondrial proteins. Examples include the transcription factor Nrf2, which binds the antioxidant response element (ARE) upstream of genes encoding cytoprotective antioxidant enzymes and Phase II proteins (Mattson, 2008). The latter pathway is activated by curcumin, sulforaphane (present in broccoli) and allicin (present in garlic). Other phytochemicals may activate the sirtuin-FOXO pathway resulting in increased expression of antioxidant enzymes and cell survival-promoting proteins; RSV has been shown to activate this pathway (Frescas et al., 2005). Ingestion of other phytochemicals may activate the hormetic transcription factors NF-κB and cAMP response element-binding (CREB) resulting in the induction of genes encoding growth factors and anti-apoptotic proteins (Mattson et al., 2006). Allicin and capsaicin activate transient receptor potential ion channels, and RSV activates sirtuin-1 (Mattson, 2008). Isothocyanates present at high levels in broccoli and watercress induced the expression of cytoprotective Phase IIproteins in liver, intestinal and stomach cells (McWalter et al., 2004); the curry spice curcumin has been reported to induce adaptive stress response genes and protect cells in animal models of cataract formation, pulmonary toxicity, multiple sclerosis and Alzheimer’s disease (Mattson, 2008); and RSV can activate stress response pathways and protect cells in models of myocardial infarction and stroke (Baur and Sinclair, 2006). Several epidemiological studies have shown beneficial effects of green tea in cancer and CVD (Kuriyama, 2008). Also, it has been demonstrated that coffee drinking may reduce the risk of liver cancer (Xu et al., 2009). Furthermore, dietary supplementation rich in polyphenols such as blueberries and apple juice showed neuroprotection for focal brain ischemia and Alzheimer’s disease (Ortiz and Shea, 2004). Individual grape compounds (Table 4.1) contain antioxidative, anti-inflammatory, antiapoptosis, antivirus, antiallergy, platelet antiaggregatory and/or anticarcinogenic properties (Aggarwal and Shishodia, 2006). Grape polyphenols reduced macrophage atherogenicity in mice, ameliorated cerebral ischemia-induced neuronal death in gerbils and exhibited a cardioprotective effect in humans (Zern et al., 2005). Freeze-dried grape powder contains anthocyanidins, catechin, epicatechin, quercetin, RSV and kaempferol (Xu et al., 2009). Anthocyanidin demonstrates cytotoxic effects in human breast, lung and gastric adenocarcinoma cells (Xu et al., 2007).
Herbal medicine has clearly recognisable therapeutic effects. The results obtained support prior observations and future pharmacologic uses concerning a huge amount of species (Table 4.2). We can outline tannins from Acacia catechu for respiratory diseases, flavonoids from Aesculus indica for joint pain, glycosides from Azadirachta indica as antipyretic, saponins, tannins, alkaloids and cardiac glycosides from Euphorbia hirta for asthma, alkaloids from Taxus wallichiana for anti-cancer theraphy and lignan glucosides from Tinospora sinensis for diabetes. Immunostimulant, antibacterial, analgesic and antiprotozoal characteristics of Andrographis paniculata extract have also been demonstrated. Crude root extract of Podophyllum hexandrum (Berberidaceae) was used as hepato-protective (Kunwar et al., 2010). The hepato-protective and hypoglycemic properties of Acacia catechu could be attributed to the quercetin and epicatechin respectively. Lectins of Cannabis sativa possess haema-gluttinating properties that corroborate the indigenous use of the leaf extract to control bleeding. Vegetale oil such as α-pinene obtained from crude leaf extract of Vitex negundo is recommended as antitussive and anti-asthma, antibacterial, antifungal, hypoglycemic, anti-cancer, acne control and inhibitor of edema to tracheal contraction (Kunwar et al., 2010). Linalool also possesses an anxiolytic effect, and this effect probably substantiates the folk uses of Skimmia anquetilia leaves as medicine for headache (Kunwar et al., 2010).
Table 4.2 Therapeutic properties of some exotic plants and their phytochemicals
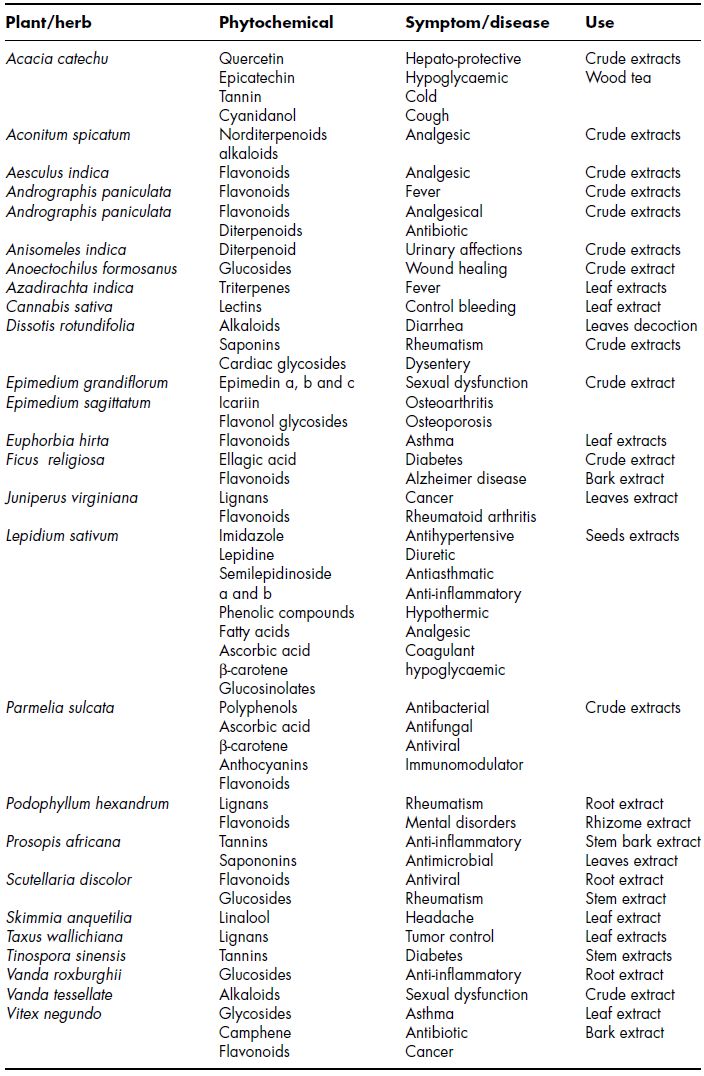
The results of pharmacological studies in Ficus religiosa will further expand the existing therapeutic activity of tannins, saponins, flavonoids, steroids and cardiac glycosides, and provide convincing support to its future clinical use in modern medicine (Singh et al., 2011). Lepidium sativum has been studied for its medicinal use in many diseases (Table 4.2), including as a bone fracture healing agent (Najeeb-Ur-Rehman et al., 2011). Orally, Epimedium has traditionally been used to treat impotence, involuntary ejaculation, weak backs and knees, postmenopausal bone loss, arthralgia, mental and physical fatigue, memory loss, hypertension, coronary heart disease, bronchitis, chronic hepatitis, HIV/AIDS, polio, chronic leukopenia and viral myocarditis. It is also used to arouse sexual desire. In clinics, Epimedium is used to treat osteoporosis, climacteric period syndrome, breast lumps, hyperpiesia and coronary heart disease (Ma et al., 2011). Taking into account their therapeutic efficiency and economical considerations, the total flavonoids and/or active ingredients might be developed into new drugs for the treatment of various diseases, especially sexual dysfunction, osteoporosis and immunity-related diseases (Ma et al., 2011).
Dissotis rotundifolia is used mainly for the treatment of rheumatism and painful swellings. The leaves decoction is used to relieve stomach ache, diarrhoea, dysentery, cough, prevent miscarriage/abortion, conjunctivitis, circulatory problems and venereal diseases (Abere et al., 2010). Extracts of D. rotundifolia have been found to possess antimicrobial and antispasmodic activities, which makes it a good candidate for further works in diarrhoea management (Abere et al., 2010). Lignans podophyllotoxin, deoxypodophyllotoxin, demethylpodophyllotoxin and podophyllotoxone are four therapeutically potent anti-cancer secondary metabolites found in Juniperus and Podophyllum species collected from natural populations in Himalayan environments and the botanical gardens of Rombergpark and Haltern (Germany). Juniperus virginiana has been used for treatmwent of genital warts, psoriasis and multiple sclerosis. Podophyllum hexandrum has been used to treat constipation, cold, fever and septic wounds (Kusari et al., 2011). Vanda tessellata is a potent aphrodisiac and fertility booster in mice. These results could be extrapolated to humans and preliminary tests could be done to see if researchers develop another drug like Viagra™.
A series of experiments were conducted on Anoectochilus formosanus, and accentuated the possibility of its commercial application for healing of different diseases. Wound healing properties of the extract of Vanda roxburghii is investigated, as it is reported in Ayurveda as a strong candidate of medicinal plant used in anti-inflammatory, antiarthritic, treatment of otitis externa and sciatica (Hossain, 2011). The combinations of sulfamethoxazole plus protocatechuic acid, sulfamethoxazole plus ellagic acid, sulfamethoxazole plus gallic acid and tetracycline plus gallic acid show synergistic mode of interaction. The identified synergistic combinations can be of potent therapeutic value against P. aeruginosa infections. These findings have potential implications in delaying the development of resistance as the antibacterial effect is achieved with lower concentrations of both drugs (antibiotics and phytochemicals). The present study clearly highlights the low toxic potential of phytochemicals as antibacterial compounds and makes suggestions on the possibility of use of the above shown synergistic drug and herb combinations for combating infections caused by this pathogen (Jayaraman et al., 2010). A large number of plant-derived triterpenoids are known to exhibit cytotoxicity against a variety of tumour cells as well as anti-cancer efficacy in preclinical animal models. Numerous triterpenoids have been synthesised by structural modification of natural compounds. Some of these analogues are considered to be the most potent anti-inflammatory and anticarcinogenic triterpenoids known in the prevention and therapy of human breast cancer (Bishayee et al., 2011).
Since ancient times, natural products, herbs and spices have been used for preventing several diseases. The term chemoprevention was coined in the late 1970s and referred to the prevention of cancer by selective use of phytochemicals or their analogues. The concept of using naturally derived chemicals as potential chemopreventive agents has advanced the field dramatically. Throughout the years, a vast number of chemopreventive agents present in natural products have been evaluated using various experimental models. A number of them have progressed to early clinical trials. More recently, the focus has been directed towards molecular targeting of chemopreventive agents to identify mechanism(s) of action of these newly discovered bioactive compounds. Moreover, it has been recognised that single agents may not always be sufficient to provide chemopreventive efficacy and, therefore, the new concept of combination chemoprevention by multiple agents or by the consumption of whole food has become an increasingly attractive area of study (Mehta et al., 2010).
Preventing many chronic diseases requires healthy dietary habits. Achieving a better balance of grain-based foods through the inclusion of whole grains is one scientifically supported dietary recommendation. Epidemiological and other types of research continue to document health benefits for diverse populations who have adequate intakes of both folic acid-fortified grain foods and whole grains. Folic acid fortification of grains is associated with reduced incidence of neural tube and other birth defects and may be related to decreased risk of other chronic disease and may contribute to specific health-maintaining and disease-preventing mechanisms (Jones et al., 2008). Despite the high levels of polyphenolic phytochemicals in grain cereals and their position as a major food staple, there has been a lack of research on the effects on both animal and human health and disease prevention. Cereal brans with a high phenolic content and high antioxidant properties inhibited protein glycation mediated by the reducing sugar fructose. These results suggest that certain varieties of cereal bran may affect critical biological processes that are important in diabetes and insulin resistance (Farrar et al., 2008). The consistent consumption of foods that contain significant levels of phytochemicals and dietary fibre correlates with tangible disease prevention. For example, whole grain comsumption is known to help in reducing the incidence of heart disease, metabolic syndrome, neuropathy, diabetes and other chronic diseases, partly due to components in cereal brans, especially dietary fibre and phytochemicals (Awika et al., 2005).
The combination of phytochemicals with relatively broad specificity on enzymes involved in signal transduction and gene expression may increase their activity in disease prevention by modulating several different molecular targets (Reiter et al., 2007). Moreover, with further development of nutrigenomics, on the basis of a simple gene test, physicians can personalise food medicine, which makes it possible for patients to control their weight, optimise their health and reduce the risk of cancer, diabetes and liver diseases (Xu et al., 2009). Dietary phytochemicals have the potential to moderate deregulated signalling or reinstate checkpoint pathways and apoptosis in damaged cells, while having minimal impact on healthy cells. These are ideal characteristics for chemopreventive and combination anti-cancer strategies, warranting substantial research effort into harnessing the biological activities of these agents in disease prevention and treatment (Manson et al., 2007). The absorption, metabolism, distribution and excretion profile of bioactive compounds is essential to assess the full potential of promising chemopreventive agents and may help guide in the design of novel synthetic analogues (Siddiqui et al., 2009). In order to optimise the chances of success in cancer chemoprevention trials, the ability to identify those individuals most likely to benefit is clearly important. In the case of primary prevention to inhibit the earliest stages of tumour development, selection has traditionally been based on known environmental and lifestyle risk factors, genetic predisposition and family history (Tsao et al., 2004). Secondary prevention is appropriate for those who have already developed pre-malignant lesions, such as intraepithelial neoplasia or intestinal polyps, the progress of which can be monitored in response to chemopreventive treatments (Manson et al., 2007). Several dietary compounds, including indoles and polyphenols, have shown promise in this respect, with regression of respiratory papillomatosis, cervical, vulvar and prostate intraepithelial neoplasia and oral leukoplakia (Thomasset et al., 2007). A third strategy is tertiary prevention, which focuses on patients who have been successfully treated for a primary tumour, in order to inhibit development of second primary tumours. Greatest success to date in this respect has resulted from the use of drugs such as tamoxifen and its analogues for breast cancer, and retinoids for skin, head and neck and liver cancer. If phytochemicals have a role at this stage, it is most likely to be as part of a combined therapy (Manson et al., 2007).
Modulation of detoxification enzymes is a main mechanism by which diet may influence risk of cancer and other diseases. However, genetic differences in taste preference, food tolerance, nutrient absorption, and metabolism and response of target tissues all potentially influence the effect of diet on disease risk. Thus, disease prevention at the individual and population level needs to be evaluated in the context of the totality of genetic background and exposures to both causative agents and chemopreventive compounds. Polymorphisms in the detoxification enzymes that alter protein expression and/or function can modify risk in individuals exposed to the relevant substrates. Genotypes associated with more favourable handling of carcinogens may be associated with less favourable handling of phytochemicals. For example, glutathione S-transferases (GST) detoxify polycyclic aromatic hydrocarbons present in grilled meats. GSTs also conjugate isothiocyanates, the chemopreventive compounds found in cruciferous vegetables. Polymorphisms in the GSTM1 and GSTT1 genes result in complete lack of GSTM1-1 and GSTT1-1 proteins, respectively. In some observational studies of cancer, cruciferous vegetable intake confers greater protection in individuals with these polymorphisms. A recent study of sulforaphane pharmacokinetics suggests that lack of the GSTM1 enzyme is associated with more rapid excretion of sulforaphane. Many phytochemicals are also conjugated with glucuronide and sulfate moieties, and are excreted in urine and bile. Polymorphisms in UDP-glucuronosyltransferases and sulfotransferases may contribute to the variability in phytochemical clearance and efficacy (Lampe, 2007).
The cancer chemopreventive activity of cruciferous vegetables such as cabbage, watercress and broccoli, Allium vegetables such as garlic and onion, green tea, citrus fruits, tomatoes, berries, ginger and ginseng, as well as some medicinal plants have been discussed. Several compounds, such as brassinin (from cruciferous vegetables like Chinese cabbage), sulforaphane (from broccoli) and its analogue sulforamate, withanolides (from tomatillos), and resveratrol (from grapes and peanuts among other foods), are in preclinical or clinical trials for cancer chemoprevention. Phytochemicals of these types have great potential in the fight against human cancer (Park and Pezzuto, 2002). Of particular significance, Indian habitual diets, which are based predominantly on plant foods like cereals, pulses, oils and spices, are all good sources of phytochemicals, particularly dietary fibre, vitamin E, carotenoids and phenolic compounds (Rao, 2003). According to the recent pharmacological findings, garlic is preventive rather than therapeutic. Epidemiological studies in China, Italy and the USA showed the inverse relationship between stomach and colon cancer incidences and dietary garlic intake. Anti-carcinogenic activities of garlic and its constituents including sulfides and S-allyl cysteine, have been demonstrated using several animal models. Garlic preparations has been also shown to lower serum cholesterol and triglyceride (TG) levels, which are major risk factors of CVD, through inhibition of their bio-synthesis in the liver, and to inhibit oxidation of low density lipoprotein (LDL). Furthermore, in vitro and in vivo studies have revealed that aged garlic extract stimulated immune functions, such as proliferation of lymphocyte, cytokine release and phagocytosis (Table 4.1). More recently, aged garlic extract has been demonstrated to prolong life span of senescence accelerated mice and prevent brain atrophy (Sumiyoshi, 1997). Besides, glucosinolates and eugenol (4-allyl-1-hydroxy-2-methylbenzene) are phytochemicals with cytochrome P-450 inducing activity. They have shown cholesterolemic effects in humans, increasing plasma high-density lipoprotein (HDL) concentrations (Hassel, 1998).
Functional foods are foods similar in appearance to a conventional food, consumed as part of the usual diet, with demonstrated physiological benefits, and/or to reduce the risk of chronic disease beyond basic nutritional functions (Hasler et al., 2004). Broccoli, carrots or tomatoes would be considered functional foods because they are rich in such physiologically active components as sulforaphane, β-carotene and lycopene, respectively (Sloan et al., 2002). Finally, there exists a growing selection of functional food components marketed under the umbrella of dietary supplements (Hasler et al., 2004). This category also includes a large number of herbal-enriched products that make a variety of structure/function claims. Examples include cereal fortified with ginkgo biloba, which is marketed as reducing symptoms of dementia, or juices with echinacea, which are marketed for boosting the immune system (Ernst and Pittler, 1999). Pharmaceutical companies have isolated many food components into supplement form to achieve disease prevention. These compounds include diallylsulfides (garlic), isoflavones (soy), anthocyanin (bilberry extract) and glycyrrhizin (licorice) (Fletcher and Fairfield, 2002).
Some alkaloids (aconitine, anisodamine, berberine, charantine, leurosine) show antidiabetic effects. Acacia catechu wood tea works as an expectorant. Additionally, the tannin and cyanidanol of the plant impart astringent activity, which helps to alleviate diarrhoea (Kunwar et al., 2010). Usnic acid and vulpunic acid of lichens are mitotic regulators and own antibiotic properties. Parmelia sulcata lichen manifests antibacterial and antifungal activities. Further pharmacological evaluation of the extracts of those species that reveal weak pharmacological validities are needed before they can be used as therapeutic potentials. The compounds that contribute to the antioxidative properties are polyphenols, vitamin C, β carotene, anthocyanins and flavonoids. Ellagic acid of Fragaria nubicola is also responsible for antioxidant activity. Wogonin of Scutellaria discolor is considered as a most potent antiviral and anxiolytic compound. Plant root extract is also useful for rheumatism (Kunwar et al., 2010). Fresh plant materials, crude extracts and isolated components of Ficus religiosa showed a wide spectrum of in vitro and in vivo pharmacological activities like, antidiabetic, cognitive enhancer, wound healing [in combination with other herbs like Ageratum conyzoides (root), C. longa (rhizome), Ficus religiosa (stem-bark) and Tamarindus indica (leaf)], anticonvulsant (modulation of glutamatergic and/or GABAergic functions), anti-inflammatory, anti-infectious diseases, hypolipidemic, antioxidant, immunomodulatory, parasympathetic, anti-tumour and hypotensive (Singh et al., 2011). Lepidium sativum Linn. is commonly known as ‘Common cress’, ‘Garden cress’ or ‘Halim’. Its seeds are popularly used as gastrointestinal stimulant, laxative, gastroprotective and digestive aid. In addition, the plant has been reported to have other properties, such as antibacterial, anti-asthmatic, diuretic, aphrodisiac and abortifacient. The plant has been reported to contain alkaloids (imidazole, lepidine, semilepidinoside A and B), β-carotenes, ascorbic, linoleic, oleic, palmitic and stearic acids, cucurbitacins and cardenolides. Moreover, a few phenolic constituents, such as sinapic acid and sinapin, were isolated from its seed extract (Najeeb-Ur-Rehman et al., 2011).
Epimedium, is a genus of about 52 species in the family Berberidaceae. Modern pharmacology studies and clinical practice demonstrated that Epimedium and its active compounds possess wide pharmacological actions, especially in hormone regulation, anti-osteoporosis, immunological function modulation, anti-oxidation and anti-tumour, anti-aging, anti-atherosclerosis and anti-depressant activities. Currently, effective monomeric compounds or active parts have been screened for pharmacological activity from Epimedium in vivo and in vitro. Epimedium pharmacological actions have attracted extensive attention (Ma et al., 2011). The major active constituents of Herba Epimedii are flavonoids, and among them epimedin A, B, C and icariin are considered major bioactive components that make up more than 52% of the total flavonoids in Herba Epimedii. A double-blind clinical trial relating to the effect of Epimedium Herbal Complex Supplement on sexual satisfaction in healthy men was compared with Viagra™ (Ma et al., 2011). Berberine, a traditional plant alkaloid, is used in Ayurvedic and Chinese medicine for its antimicrobial and antiprotozoal properties. Interestingly, current clinical research on berberine has revealed its various pharmacological properties and multi-spectrum therapeutic applications, including diabetes, cancer, depression, hypertension and hypercholesterolemia (Vuddanda et al., 2010). Dissotis rotundifolia revealed the presence of alkaloids, saponins and cardiac glycosides. The pharmacological effectiveness of glycosides is dependent on the aglycones, but the sugars render the compounds more soluble and increase the power of fixation of the glycosides. On the basis of the overall results from several investigations, the use of D. rotundifolia in the treatment of diarrhoea, veneral diseases, dysentery and relief of stomach ache is justified (Abere et al., 2010).
Orchid phytochemicals are generally categorised as alkaloids, flavonoids, carotenoids, anthocyanins and sterols. A few studies have been conducted on animal bodies, such as mice, rabbits, frogs and guinea pigs, which created optimism that life saving phytochemicals, like Taxol, Vinblastine or Quinine, will be proved. Organic compounds, called stilbenoids, inhibited aortic contractions provoked by noradrenaline and caused vasodialation, the relaxation and widening of blood vessels in the body. Again, the implications of these chemicals for usage in human models may be promising in cardiology, pending further examination (Hossain, 2011). In a recent study, the extract of the stem bark of Prosopis africana was evaluated for analgesic and anti-inflammatory activities in rats, comparable to that of piroxicam – the standard agent used. The preliminary phytochemical screening revealed the presence of flavonoids, saponins, carbohydrates, cardiac glycosides, tannins, terpenes and alkaloids (Ayanwuyi et al., 2010). Pseudomonas aeruginosa is a major nosocomial pathogen, particularly dangerous to cystic fibrosis patients and populations with weak immune system. In a recent study the in vitro activities of seven antibiotics (ciprofloxacin, ceftazidime, tetracycline, trimethoprim, sulfamethoxazole, polymyxin B and piperacillin) and six phytochemicals (protocatechuic acid, gallic acid, ellagic acid, rutin, berberine and myricetin) against five P. aeruginosa isolates, alone and in combination, have been evaluated (Jayaraman et al., 2010).
The heart is an aerobic organ, and most of the energy required for the contraction and maintenance of ion gradients comes from oxidative phosphorylation, generating a large amount of ROS (Matés et al., 2009b). Therefore, a great deal of attention has focused on the naturally occurring antioxidant phytochemicals as potential therapy for CVD. Until 500 generations ago, all humans consumed only wild and unprocessed food foraged and hunted from their environment. These circumstances provided a diet high in lean protein, polyunsaturated fats (especially omega-3 fatty acids), monounsaturated fats, fibre, vitamins, minerals, antioxidants and other beneficial phytochemicals. Historical and anthropological studies show hunter-gatherers generally to be healthy, fit and largely free of the degenerative CVD common in modern societies (O’Keefe and Cordain, 2004). CVD is the number one cause of death and disability of both men and women in the USA with a high impact on human health and community social costs (Anderson, 2002). Many compounds in grains, including antioxidants, phytic acid, lectins, phenolic compounds, amylase inhibitors and saponins, have been shown to alter risk factors for CVD. It is probable that the combination of compounds in grains, rather than any one component, explains their protective effects in CVD (Slavin et al., 2004). The phytochemical-rich diet included dried fruits, nuts, tea, whole grain products, fresh fruits and vegetables. The whole food diets significantly lowered serum cholesterol and LDL-cholesterol (LDL-C) and decreased measures of antioxidant defence, all biomarkers of decreased risk of chronic disease (Slavin et al., 2003). Refined diets that do not include whole grains were associated with higher serum cholesterol levels (Slavin et al., 2004). Recent studies find that serum enterolactone is associated with reduced CVD-related and all-cause death (Slavin et al., 2003). As already stated, whole grains are rich in compounds such as tocotrienols (a form of vitamin E) and other plant sterols (i.e. β-sitosterol), and short-chain fatty acids (i.e. acetate, butyrate and propionate), which can lower cholesterol (Slavin et al., 2004). Oxidative stress induced by ROS plays an important role in the aetiology of CVD. In particular, the LDL-oxidisation has a key role in the pathogenesis of atherosclerosis and cardiovascular heart diseases through the initiation of plaque formation process. Dietary phytochemical products such antioxidant vitamins (A, C and E) and bioactive food components (α- and β-carotene) have shown an antioxidant effect in reducing both oxidative marker stress and LDL-oxidisation process. Lycopene, an oxygenated carotenoid with great antioxidant properties, has shown both in epidemiological studies and supplementation human trials a reduction of cardiovascular risk (Riccioni et al., 2008).
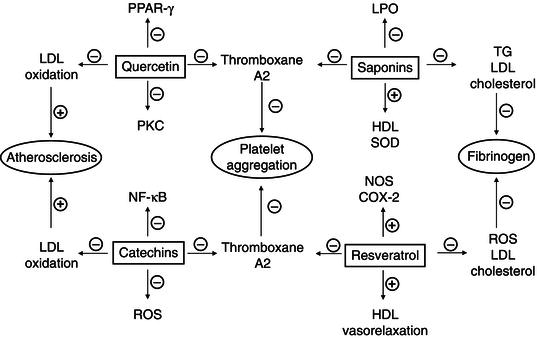
Epidemiological studies suggest that diets rich in polyphenols may be associated with reduced incidence of cardiovascular disorders (mainly coronary heart disease and myocardial infarction). Current evidence suggests that polyphenols, acting at the molecular level, improve endothelial function and inhibit platelet aggregation. In view of their antithrombotic, anti-inflammatory, and anti-aggregative properties, these compounds may play a role in the prevention and treatment of CVD. The antioxidant activity of several polyphenols positively correlated with the presence of a catechol ring in their molecular structure. Catechin, 3,4-dihydroxycinnamic acid, 3,4-dihydroxyhydrocinnamic acid, 3,4-dihydroxyphenylacetic acid, feluric acid, gallic acid and quercetin presented significant antioxidant capacity at concentrations commensurate with human plasma. The anti-atherosclerotic action of polyphenols is based on the removal of already formed ROS from the blood and on the inhibition of enzymes generating ROS, such as lipoxygenase, cyclo-oxygenase (COX), xanthine oxidase and NADPH oxidase by polyphenols. Polyphenols, particularly quercetin, are able to chelate pro-oxidant metals (mainly iron). Catechins and procyanidins block the enzymatic production of ROS, while quercetin displays a similar action of inhibiting the PKC-dependent NADPH oxidase (Figure 4.1). Moreover, polyphenols, as scavengers of the O2•-, limits its reaction with nitric oxide (NO), as a result of which highly toxic ONOO- is formed. Quercetin inhibits platelet reactivity through blocking collagen receptor (GPVI)-dependent activation. Molecular studies have also demonstrated an antagonistic action of flavones (apigenin) and isoflavones (genistein) on thromboxane A receptors. Flavonols and their derivatives, procyanidins, inhibit the expression of endothelial adhesion molecules VCAM-1, ICAM-1 and E-selectin and the activation of leukocytes and thus prevent the formation of platelet-leukocyte aggregates. Quercetin also reduces the activation of peroxisome proliferator-activated receptor gamma (PPAR-γ) (Michalska et al., 2010). Regarding this, Hayek et al. (1997) observed reduced susceptibility to LDL oxidation and an attenuation of the development of atherosclerotic lesions in the aortic arch in mice fed red wine or quercetin and, to a lesser extent, in mice fed catechins.
Figure 4.1 Summary of key modulatory effects on cardiovascular diseases of quercetin, resveratrol, saponins and catechins.
COX: cyclo-oxygenase, HDL: high-density lipoprotein, LDL: low-density lipoprotein, LPO: lipid peroxidation, NF-κB: nuclear factor kappaB, NOS: nitric oxide synthase, PKC: protein kinase C, PPAR-γ: proliferator-activated receptor gamma, ROS: reactive oxygen species, SOD: superoxide dismutase, TG: triglycerides.
Oxidation of LDL plays a crucial role in the initiation mechanism of atherosclerosis (Berliner et al., 1996). An epidemiologic study indicated that European populations with higher plasma concentrations of natural antioxidants, ascorbic acid and α-tocopherol have a lower incidence of coronary heart disease (Miura et al., 2001). Several epidemiologic investigations indicated that flavonoid intake is inversely associated with the mortality of coronary heart disease (Hertog et al., 1997). Apart from lowering cardiovascular risk factors associated with diabetes, phytosterols (β-sitosterol in particular) have been shown directly lowering fasting blood glucose levels by cortisol inhibition. Among the most abundant compounds from Aloe ferox leaf are protocatechuic, p-coumaric, p-toluic, benzoic, hydroxyphenylacetic, xanthine and β-sitosterol. Due to the occurrence of the polyphenols, phytosterols and perhaps the indoles present, A. ferox leaf gel may show promise in alleviating or preventing the symptoms associated with CVD (Loots et al., 2007). On the other hand, regular consumption of fruit Euphoria longana Lam. (longan), whose major components were identified as gallic acid, corilagin and ellagic acid, is associated with a lower risk of CVD (Rangkadilok et al., 2005). In addition, Scottish Heart Health Study and other studies all indicated an inverse correlation between black tea consumption and the risk of coronary heart disease (Miura et al., 2001). Green tea leaves (Camellia sinensis) contain antioxidative tea catechins consisting of various flavan 3-ols as follows: (+)-catechin, (–)-epicatechin, (–)-epicatechin gallate, (–)-epigallocatechin and (–)-epigallocatechin-3-gallate (EGCG). Green tea catechins exert potent inhibitory effects on Cu2+ -mediated oxidative modification of LDL in vitro. Daily consumption of catechins prevents the development of atherosclerosis (Figure 4.1). Green tea, and its principal component (EGCG), exerts a much stronger antioxidative effect than teaflavin and tealubidin, the major components of black tea. Several lines of investigation demonstrated that catechins have potent scavenging effects on O2•- and •OH (Miura et al., 2001). Lin and Lin (1997)reported that EGCG blocks the induction of nitric oxide synthase (NOS) by down-regulating lipopolysaccharide-induced activity of the transcription factor, NF-κB, which is a pleiotropic mediator for induction of genes, including genes relevant to atherogenesis.
Accumulating evidence has suggested that the isoflavones or soy protein only reflected certain aspects of health effects associated with soy consumption. For example, using primate models of atherosclerosis, the intact soy protein has been shown to be effective in lowering cholesterol. Studies have shown that non-isoflavone compounds, such as soyasaponins, phytic acid or plant sterols, display a wide range of bioactivities, including cardiovascular protective effects (Kang et al., 2010). Soyasaponins showed their cardiovascular protective effects through several different mechanisms (Oakenfull and Sidhu, 1990). In animal models, soyasaponins were found to significantly reduce the serum total cholesterol, LDL-C and TG concentrations and to increase the HDL-cholesterol (HDL-C) levels (Xiao et al., 2005). 24-Methylenecycloartanol, in combination with soysterol, greatly reduced plasma cholesterol and enhanced cholesterol excretion in rats (Kang et al., 2010). Total soyasaponins prevented the decrease of blood platelets and fibrinogen, and the increase of fibrin degradation products in the disseminated intravascular coagulation (Figure 4.1). In vitro experiments, total soyasaponins, soyasaponins I, II, A1, and A2 inhibited the conversion of fibrinogen to fibrin (Kang et al., 2010). Total soyasaponins decreased elevated blood sugar and lipid peroxidation levels and increased the decreased levels of superoxide dismutase (SOD) in diabetic rats (Wang et al., 1993). Phytosterols (not restricted to soy-based sources), which have long been known to reduce intestinal cholesterol absorption, lead to decreased blood LDL-C levels and lower CVD risk (Kang et al., 2010).
There has been great deal of focus on the naturally occurring antispasmodic phytochemicals as potential therapy for CVD. Diterpenes exert several biological activities such as anti-inflammatory action, antimicrobial and antispasmodic activities. Several diterpenes have been shown to have pronounced cardiovascular effects, for example, grayanotoxin I produces positive inotropic responses, forskolin is a well-known activator of adenylate cyclase, eleganolone and 14-deoxyandrographolide exhibit vasorelaxant properties and marrubenol inhibits smooth muscle contraction by blocking L-type calcium channels. In the last few years, the biological activity of kaurane and pimarane-type diterpenes, which are the main secondary metabolites isolated from the roots of Viguiera robusta and V. arenaria, respectively, has been investigated. These diterpenoids exhibit vasorelaxant action and inhibit the vascular contractility mainly by blocking extracellular Ca2+ influx. Moreover, kaurane and pimarane-type diterpenes decreased mean arterial blood pressure in normotensive rats. Diterpenes likely fulfil the definition of a pharmacological preconditioning class of compounds and give hope for the therapeutic use in CVD (Tirapelli et al., 2008). The aqueous extracts from three popular Thai dietary and herbal plants, Cratoxylum formosum, Syzygium gratum and Limnophila aromatica possessed high free radical scavenging and antioxidant activities. Vascular responsiveness to bradykinin, acetylcholine and phenylephrine in phenylhydrazine-control rats was markedly impaired. Moreover, the plant extracts prevented loss of blood GSH and suppressed formation of plasma malondialdehyde, plasma NO metabolites and blood O2•-. It was concluded that the plant extracts possess antioxidants and have potential roles in protection of vascular dysfunction (Kukongviriyapan et al., 2007).
Recent studies have highlighted the role of dietary fibre, particularly water-soluble varieties, in decreasing the risk of CVD. Several types of soluble fibre, including psyllium, β-glucan, pectin and guar gum, have been shown to decrease LDL-C in well-controlled intervention studies, whereas the soluble fibre content of legumes and vegetables has also been shown to decrease LDL-C (Bazzano, 2008). Surprisingly, the consumption of insoluble fibre from whole grains, though metabolically inert, has been associated with a reduction in the risk of developing coronary heart disease in epidemiological studies. The likely reason is that whole grains, like nuts, legumes and other edible seeds, contain many bioactive phytochemicals and various antioxidants. After cereals, nuts are the vegetable foods that are richest in fibre, which may partly explain their benefit on the lipid profile and cardiovascular health (Salas-Salvadó et al., 2006). On the other hand, men who had low concentrations of β-caroteno and vitamin C in their blood had a significantly increased risk of subsequent ischemic and coronary heart disease, suggesting that carotenoid-containing diets are protective against CVD. In conclusion, the consumption of β-carotene-rich foods has been associated consistently with a decreased risk of CVD. In contrast, supplementation with β-carotene in major intervention trials generally has failed to reduce the incidence of CVD (Mayne, 1996).
Stilbenes have been shown to protect lipoproteins from oxidative damage and to have chemopreventive activity (Vitrac et al., 2005). Resveratrol (3,4′,5-trihydroxy-trans-stilbene), or (E)-5-(p-hydroxystyryl)resorcinol, is a naturally occurring polyphenolic compound abundant in grapes, peanuts, red wine, pines and other leguminosae family plants in response to injury, ultraviolet irradiation and fungal attack (Matés et al., 2009a). Epidemiological evidence has shown that CVD is less prevalent in the French population than expected in light of their saturated fat intake and serum cholesterol concentrations (Zenebe et al., 2001). The protective effect of moderate consumption (two to three units) of red wine on the risk of CVD morbidity and mortality, however, has been consistently shown in many epidemiological studies (Zenebe and Pechanova, 2002). Phenolic compounds and especially a group of flavonoids seem to be responsible for the majority of protective effects of red wine on CVD, particularly their antithrombic, antioxidant, anti-ischemic, vasorelaxant and antihypertensive properties (Zenebe et al., 2001). Polyphenols have been shown to be able to modulate the process of thrombosis in several systems. Fuster et al. (1992) reported that a reduced rate of development of atherosclerosis and coronary artery disease caused by daily intake of flavonoids was based mainly on the possibility of flavonoids to inhibit acute thrombus formation. One of the most recognised and widely studied compounds is RSV, a phytoalexin member of a family of polyphenols called viniferins. Although RSV was first isolated in 1940 from the roots of white hellebore (Veratrum grandiflorum), the importance of RSV was recognised only after the widely publicised historic French paradox associated with drinking of red wine. Both epidemiological and experimental studies have revealed that drinking wine, particularly red wine, in moderation protects cardiovascular health. A growing body of evidence supports the role of RSV as evidence based cardiovascular medicine. RSV protects the cardiovascular system in multidimensional ways. The most important point about RSV is that, at a very low concentration, it inhibits apoptotic cell death, thereby providing protection from various diseases including myocardial ischemic reperfusion injury, atherosclerosis and ventricular arrhythmias (Figure 4.1). Both in acute and in chronic models, RSV-mediated cardioprotection is achieved through the preconditioning effect, rather than direct effect as found in conventional medicine. The same RSV when used in higher doses facilitates apoptotic cell death and behaves as a chemopreventive alternative. RSV likely fulfils the definition of a pharmacological preconditioning compound and gives hope for the therapeutic promise of alternative medicine (Das and Das, 2007).
Evidence indicates that some polyphenols modulate specific pathways regulating the expression and activation of genes involved in the control of the cardiovascular system (Zenebe et al., 2001). It is possible that red wine polyphenols decrease degradation of basal levels of NO, preventing its destruction by superoxides, or stimulate NO synthase in endothelial cells. It is conceivable that both mechanisms are active in vivo (Andriambeloson et al., 1997). Red wine polyphenols reduced the level of thromboxane A2 similarly to acetylsalicylic acid. Polyphenols, in contrast to acetylsalicylic acid, had a shorter-term effect on coronary blood flow but interfered with glycoprotein receptors on endothelial cells. Several polyphenols have been also shown to interfere with several enzyme systems critically involved in cellular responses, such as tyrosine and serine-threonine PKs, phospholipases and COXs (Middleton et al., 2000). Adhesion of platelets to the subendothelial matrix, after vessel damage, is a triggering mechanism of thrombus formation, and thus platelet inhibition by red wine may partially explain the prevention of thrombus growth (Zenebe et al., 2001). In humans, Pace-Asciak et al. (1995) showed that polyphenolic compounds from red wine, especially quercetin, catechin and RSV, inhibited the synthesis of thromboxane in platelets and of leukotriene in neutrophils. In their experiments, RSV and quercetin exhibited a dose-dependent inhibition of thromboxane-induced and ADP-induced platelet aggregation, while epicatechin, α-tocopherol and butylated hydroxytoluene were inactive. RSV also inhibited synthesis of thromboxane B2 and hydroxyheptadecatrienoate, and slightly inhibited synthesis of 12-hydroxyeicosatetraenoate. Alcohol-free red wine only inhibited the synthesis of thromboxane B2. Interestingly, cyclic reductions in coronary flow (CFRs) were eliminated by red wine and grape juice when given intravenously or intragastrically; however, a 2.5-fold greater amount of grape juice than red wine was needed for the elimination of CFRs. In the case of white wine, the elimination of CFRs was not significant (Zenebe et al., 2001). Quercetin and rutin were also found to eliminate CFRs in the same model. Measurement of quercetin, rutin and RSV content of red wine, white wine and grape juice indicated that flavonoid content was several-fold higher in red wine and grape juice than in white wine (Wollny et al., 1999). Red wine consumption was also found to increase plasma HDL concentrations characterised by their antiatherogenic effects (Zenebe et al., 2001). All the mechanisms by which red wine polyphenols exert their antiatherogenic effect appear to be crucial in the prevention and treatment of CVD. Sato et al. (2000) found that an ethanol-free red wine extract as well as RSV protected the heart from detrimental effects of ischemia-reperfusion injury, as seen by improved postischemic ventricular function and reduced myocardial infarction. Both the red wine extracts and RSV reduced oxidative stress in the heart, as indicated by decreasing malondyaldehyde formation. A reduction effect of several flavonoids on acute regional myocardial ischemia in isolated rabbit hearts was also reported (Zenebe et al., 2001). Ning et al. (1993) showed that flavone administration improved functional recovery in the reperfused heart after a bout of global ischemia. The effect of flavone on postischemic recovery was proposed to be caused by its stimulation of the cytochrome P450 system. Quercetin was reported to exert a protective effect by preventing the decrease in the xanthine dehydrogenase to oxidase ratio observed during ischemia-reperfusion in rats (Zenebe et al., 2001). The protective effects of flavonoids in cardiac ischemia are also associated with their ability to inhibit mast cell secretion, which may be involved in cardiovascular inflammation, at present considered one of the key factors in coronary artery disease (Ridker et al., 1998).
Polyphenolic compounds have also the ability to relax precontracted smooth muscle of aortic rings with intact endothelium; moreover, some of them are able to relax endothelium-denuded arteries (Andriambeloson et al., 1997). Because red wine polyphenols consist of hydroxycinnamic acid, proanthocyanidins, anthocyanins, flavanes and flavonols, the question of which substance(s) may be responsible for increased NO synthesis had to be addressed (Zenebe et al., 2001). From anthocyanin-enriched wine extracts, aglycone-, monoglycoside- and diglycoside-enriched fractions induced endothelium-dependent vasorelaxation, similar to that elicited by the original red wine polyphenolic extract. The representative derivatives of phenolic acid (benzoic, vanillic and gallic acid), hydroxycinnamic acid (p-coumaric and caffeic acid), flavanols (catechine and epicatechine) and the higher polymerenriched fraction of condensed tannins failed to induce endothelium-dependent vasorelaxation (Stoclet et al., 2000). Mechanisms implicated in the vasorelaxant effects of flavonoids may also include inhibition of cyclic nucleotide phosphodiesterases and activation of Ca2+- activated K+ channels (Zenebe et al., 2001). Both an increase in NO synthase activity and a decrease in phosphodiesterase activity may lead to increased cyclic guanosine monophosphate (cGMP) concentration, resulting in vasorelaxation and inhibition of platelet aggregation. The ability of polyphenolic compounds to activate the NO-cGMP system seems to be associated also with their antihypertensive effect. Mizutani et al. (1999) reported that in vivo administration of an extract of polyphenolic compounds from wine attenuated elevated blood pressure in spontaneously hypertensive rats, and Hara (1992) found that in vivo administration of an extract of polyphenolic compounds from tea reduced blood pressure and decreased risk of stroke in susceptible rats. Improved biomechanical properties of aorta, lowering of cholesterol concentrations and inhibition of LDL oxidation were suggested as the mechanisms responsible for blood pressure reduction (Zenebe et al., 2001). This hemodynamic effect of red wine polyphenolic compounds was associated with augmented endothelium-dependent relaxation and a modest induction of gene expression of inducible NO synthase and COX-2 within the arterial wall, which together maintained unchanged agonist-induced contractility (Diebolt et al., 2001).
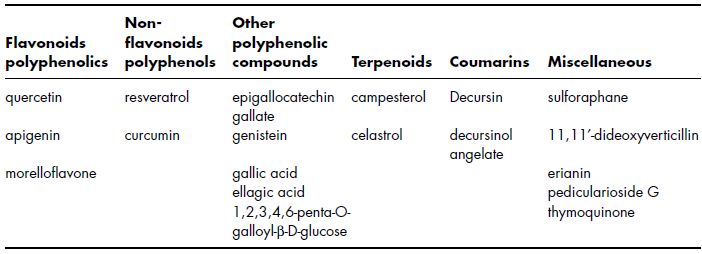
Cancer induction, growth and progression are multi-step events and numerous studies have demonstrated that various dietary agents interfere with these stages of cancer. Fruits and vegetables represent an untapped reservoir of various nutritive and nonnutritive phytochemicals with potential cancer chemopreventive activity. Overall, completed studies from various scientific groups conclude that a large number of phytochemicals are excellent sources of various anti-cancer agents and their regular consumption should thus be beneficial to the general population (Kaur et al., 2009). Evidence has shown that dietary polyphenolic compounds including anthocyanidins from berries, catechins from green tea, curcumin from turmeric, genistein from soy, lycopene from tomatoes and quercetin from red onions and apples are phytochemicals with significant anti-cancer properties (Bishayee et al., 2010). Identification of dietary phytochemicals that target key molecules regulating apoptosis, invasion and angiogenesis has become a major focus of cancer chemoprevention in recent years (Yang et al., 2009). Many compounds have also shown a high efficacy on tumour angiogenesis (Jeong et al., 2011), i.e. polyphenolic compounds (flavonols, flavones, flavanols, isoflavones, phenolic acids), non-flavonoids polyphenols (stilbenes and pterostilbenes), terpenoids (terpenes, sesquiterpenes) and indoles (sulforaphane) (Table 4.3) (El-Najjar et al., 2010).
In recent years, the effects of phytochemicals on cell transformation and suppression of transformed cells during the different phases of carcinogenesis have been a topic of interest to many laboratories (Kang, 2010). Among specific groups, Allium vegetables and garlic were considered to offer probable protection against stomach and colorectal cancers, respectively. Other groups, i.e. cruciferous vegetables (source of isothiocyanates and indoles) or tea (source of polyphenols), received little attention in this report. Overall, the link between diet and health seems to be much more complicated than previously anticipated. Further investigations into the potential of phytochemicals, which take account of modifying factors, a potential threshold effect and cancer subgroups, are essential to establish their effective use in chemoprevention (Moiseeva and Manson, 2009). Where phytochemicals (I3C, tea polyphenols and curcumin) have been investigated in extended trials, they have been associated with very few side effects (Rosen and Brison, 2004). In this regard, I3C has shown great promise as a chemopreventive agent for several types of cancer, yet enthusiasm for this compound has been somewhat diminished due to its unstable characteristics upon exposure to acids in the stomach (Mehta et al., 2010). Resveratrol also merits further clinical evaluation as a potential colorectal cancer chemopreventive agent. Recent results suggest that daily doses of resveratrol produce levels in the human gastrointestinal tract of an order of magnitude sufficient to elicit anticarcinogenic effects (Patel et al., 2010).
Table 4.3 Phytochemicals showing a high efficacy on tumour angiogenesis
The lower rates of several chronic diseases in Asia, including certain types of cancer, have been partly attributed to consumption of large quantities of soy foods (Kang, 2010). Genistein from soy has demonstrated breast and prostate cancer preventive activities (Mage and Rowland, 2004). Conversely, the tumour-promoting effects of high doses of genistein have been confirmed by the USA National Toxicology Program. Moreover, combining EGCG and genistein in the diet enhanced intestinal tumourigenesis (Moiseeva and Manson, 2009). Increasing evidence suggests the potential toxicity of some dietary phytochemicals. For instance, overdose of flavonoids could increase the risk of leukemia in offspring (Ross et al., 1994). It was also reported that EGCG can reduce cell viability, which was associated with increased production of ROS and depletion of GSH. Therefore, it is required to assess the adverse effects of certain diet-derived compounds (Yang et al., 2010). The purported benefits of healthy dietary agents are challenged by the uncertain results regarding the lowering of cancer risk that were obtained in large-scale intervention studies using specific single dietary ingredients at supraphysiological doses (Hsieh and Wu, 2009). The concept of functional synergy was tested by investigating the combination of EGCG and genistein, derived from tea and soy products commonly found in a traditional Asian diet, with quercetin, present in abundance in fruits and vegetables, for efficacy against CaP (Conte et al., 2004). Each chosen agent reportedly has shown anti-CaP activities, with overlapping and distinct molecular actions and targets. For example, EGCG acts at G1/S whereas genistein affects the G2/M checkpoint of the cell cycle (Hsieh and Wu, 2009). EGCG exerts epigenetic control by inhibiting DNA methyl-transferases (Fang et al., 2003). Synergy was observed between EGCG, genistein and quercetin in regards to the control of androgen receptor, p53 and NAD(P)H:quinone oxidoreductase (NQO1) (Hsieh and Wu, 2009). As stated before, a combination of agents is more effective than any single constituent in achieving chemopreventive effects (Nakamura et al., 2009). For this reason, studies on synergistic effects of different phytochemicals might contribute to the chemopreventive strategies against malignant tumours.
Genistein is a soy-derived isoflavone with multiple biochemical effects, including the alteration of cell cycle-regulatory kinase activities (Banerjee et al., 2008). Previous studies indicated that genistein enhanced the induction of apoptosis by chemotherapeutic agents, and increased radiosensitivity in several cancer cell lines (Sarkar and Li, 2006). Genistein is also known as an estrogen receptor agonist and it can antagonise the proliferation of breast cancer cells by estradiol (Figure 4.2). I3C and genistein synergistically induces apoptosis in human colon cancer HT-29 cells by inhibiting Akt phosphorylation and progression of autophagy (Nakamura et al., 2009). It is believed that phenolics can exert their effects on the different signaling pathways such as mitogen-activated protein kinases (MAPK), activator protein-1 (AP-1) or NF-κB either separately or sequentially, as well as possibly interacting between/among these pathways, which can offer complementary and overlapping mechanisms of action. Bioactive compounds can offer additive or synergistic interaction through different biochemical targets (Yang and Liu, 2009). For example, quercetin could enhance the action of carboxyamidotriazole in human breast carcinoma MDA-MB-435 cells (Liu, 2004). Other anti-cancer polyphenols are found in tea, in particular catechins. In green tea, EGCG, (−)-epicatechin-3-gallate and (−)-epicatechin are the major compounds. EGCG reduces the growth of gastric cancer (Zhu et al., 2007) and inhibits the progression of human pancreatic cancer cells, inducing apoptosis. Several observations have indicated that a number of anti-neoplastic effects could be found among several well-studied anthraquinones, including the phenolic compounds emodin, aloe-emodin and rhein (Lentini et al., 2010).
Figure 4.2 Summary of key modulatory effects on cancer of genistein, silymarin, curcumin and resveratrol
Akt: protein kinase B, Cdc: cell division cycle, Cdk: cyclin-dependent kinases, COX: cyclo-oxygenase, GSH: glutathione, ER: estrogen receptor, NF-κB: nuclear factor kappaB, ODC: ornithine decarboxylase, ROS: reactive oxygen species, SOD: superoxide dismutase.

Ginger rhizome (Zingiber officinale) is consumed worldwide as a spice and herbal medicine. It contains pungent phenolic substances collectively known as gingerols (Lee et al., 2008). One of the gingerols, 6-gingerol, was found to be a major pharmacologically active component of ginger. It has anti-inflammatory, antioxidant and anti-cancer activities (Kim et al., 2005). Genotoxic effects of 6-gingerol in HepG2 cells can be used as a suitable system for the prediction of toxicity, carcinogenicity and cell genotoxicity in humans (Yang et al., 2010). A decrease of GSH was observed in HepG2 cells exposed to 6-gingerol, which indicates GSH, as a main intracellular antioxidant, plays a vital role in defence against genotoxic effects induced by 6-gingerol. As a natural product of ginger, 6-gingerol is recommended for prevention of cancer and other diseases (Kim et al., 2005). 6-Gingerol is anti-mutagenic as well as mutagenic depending on the tested dose, and its active part is the aliphatic chain moiety containing a hydroxy group. Some studies have also found that both cinnamaldehyde and vanillin are anti-mutagens and DNA-damaging agents (King et al., 2007). The motility and invasive potential of many metastatic cancer cell lines has been inhibited by phytochemicals such as 6-gingerol, genistein, apigenin, ganoderic acid from the mushrooms Ganoderma lucidum and Phellinus linteus (Adams et al., 2010).
Blueberry decreased cell proliferation in HCC38, HCC1937 and MDA-MB-231 cells with no effect on the nontumuorigenic MCF-10A cell line. Additionally, black raspberries inhibited esophageal tumours in rats and modulated NFκB, AP-1, nuclear factor of activated T cells and the expression of a number of genes associated with cellular matrix, cell signaling and apoptosis (Li et al., 2008). Oral intake of blueberries could be a key component of long-term breast cancer prevention strategies. It was suggested that the antiproliferative activity of fruit extracts against cancer cell lines is due to the production of H2O2 and resultant oxidative stress. A single serving of fresh blueberries could be an important part of dietary cancer prevention strategies (Adams et al., 2010). An extract of Mangifera pajang kernel has been previously found to contain a high content of antioxidant phytochemicals (Naczk and Shahidi, 2006). Consistent with this, M. pajang kernel extract has been shown to inhibit the proliferation of liver carcinoma (HepG2), ovarian carcinoma (Caov3) and colon carcinoma (HT-29) cell lines in vitro (Abu Bakar et al., 2010a); however, the extract did not inhibit the proliferation of normal human fibroblasts, suggesting a selective action of the kernel extract on tumour cells. Extract of the kernel of M. pajang contains high levels of phenolic compounds, including phenolic acids (gallic, p-coumaric, sinapic, caffeic, ferulic and chlorogenic) and flavonoids (naringin, hesperidin, rutin, luteolin and diosmin) (Abu Bakar et al., 2010a). Many of these are known to inhibit growth of breast cancer cells, and so it is likely that at least some of the growth inhibitory effect of the extract can be ascribed to these phenolic compounds (Abu Bakar et al., 2010b). On the other hand, similar extracts of a related Mangifera species, M. indica (mango), have been demonstrated to contain a xanthine glycoside, mangiferin, in addition to other phenolic compounds, which is a highly active cytotoxic agent (Masibo and He, 2008). To emphasise, luteolin potentiates the cytotoxicity of cisplatin in LNM35 cells and decreases the growth of LNM35 tumour xenografts in athymic mice after intraperitoneal injection. Thus, luteolin, in combination with standard anti-cancer drugs such as cisplatin, may be a promising phytochemical for the treatment of lung cancer (Attoub et al., 2011).
The diferuloylmethane curcumin has been consumed for centuries in Asian countries as a dietary spice in amounts in excess of 100 mg/day without any side effects. In Southeast Asia, up to 4 g per adult/day appears to lower the incidence rate of colorectal cancer. This spice and food-colouring agent has been considered as nutraceutical because of its strong anti-inflammatory, antitumour, antibacterial, antiviral, antifungal, antispasmodic and hepato-protective roles (Montopoli et al., 2009). Increasingly, preclinical and clinical evidence supports curcumin’s chemopreventive and antitumour progression properties against human malignancies (Matés et al., 2009a). Curcumin acts on cell signaling pathways, modulates transcription factor activities, induces apoptosis, modulates the cell cycle and cell adhesion, and inhibits angiogenesis and metastasis (Fong et al., 2010). Curcumin inhibits cell proliferation, arrestes the cell-cycle progression and induces cell apoptosis in rat aortic smooth muscle cell line (A7r5). It produces similar effect on human leukemia HL-60, mouse leukemia WEHI-3 cells, and on the population of B cells from murine leukemia in vivo (Su et al., 2008). While men residing in Asia show a lower incidence of CaP compared to Caucasian males, Asian men who move to and live in the United States and adopt a Western lifestyle have CaP rates indistinguishable from Caucasian males. These findings suggest that Asian diets contain ingredients that might protect against the development of CaP. Here, whether or not a combination of EGCG, genistein and quercetin, phytochemicals present in a traditional Asian diet, might exert synergy in controlling proliferation and gene expression of cancer cells (Hsieh and Wu, 2009). In vitro, curcumin causes cell-cycle arrest in many different tumour cells by affecting various molecular targets, such as up-regulating Cdk inhibitors and p53 and down-regulating cyclin D1, Cdk-1, cdc2, NFκB (Aggarwal et al., 2006). In vivo, curcumin reduces incidence and multiplicity of both epithelial invasive and non-invasive adenocarcinomas (Sharma et al., 2004). Curcumin causes suppression, retardation or inversion of carcinogenesis, i.e. decreasing the frequency of tongue carcinoma, and oral carcinogenesis. Curcumin combined with cisplatin or oxaliplatin cause best dose- and time-dependent increases in cell-cycle arrest and apoptosis (Figure 4.2). Curcumin is cytotoxic to ovarian cancer cells (Montopoli et al., 2009). In experimental models with mice using a soluble formulation of curcumin (dimethyl sulfoxide), curcumin injected at the tail vein crosses the blood-brain barrier and kills tumour cells inside the brain (Purkayastha et al., 2009). This investigation becomes the first in vivo demonstration of the anti-carcinogenic and anti-metastatic activity of curcumin in the brain. In summary, curcumin acts as antioxidant (scavenging free radicals), detoxifying agent (inducing Phase II enzymes) and like a multi drug resistance compound (inhibiting ABC transporters) (Fong et al., 2010). Courmarins, tannins, ketones weakly moderated tumour specific cytotoxicity against human oral squamous cell carcinomas, whereas anthracyclines, nocobactins and cyclic α,β-unsaturated compounds showed much higher tumour specific cytotoxicity (Shin et al., 2010).
Silibinin is the primary active constituent of crude extract (known as silymarin) from the seeds of milk thistle plant Silybum marianum (Matés et al., 2009a). Chemically, the active constituent of this extract is a flavolignan, silymarin, which in itself represents the mixture of four isomeric flavonoids: silibinin, isosilibinin, silydianin and silychristin. Silibinin is the major component (70–80%) found in silymarin and is thought to be the most biologically active (Ramakrishnan et al., 2009). Several studies have clearly shown the preclinical efficacy of both silibinin and its crude extract source, silymarin, against various epithelial cancers, and at least silibinin efficacy is currently being evaluated in cancer patients (Deep and Agarwal, 2007). Silibinin treatment strongly inhibits the growth of colorectal cancer LoVo cells and induces apoptotic death, which was associated with increased levels of cleaved caspases and cleaved poly(ADP-ribose) polymerase. Analyses of xenograft tissue showed that silibinin treatment inhibits proliferation and increases apoptosis (Figure 4.2). Together, these results suggest the potential use of silibinin against advanced human colorectal cancer (Kaur et al., 2009). Pharmacological studies have revealed that silymarin is nontoxic even at relatively high physiological doses, which suggests that it is safe to use for treatment of various diseases (Matés et al., 2009a). Some studies have shown that silymarin is a strong antioxidant and hypolipidaemic agent with a potent anticarcinogenic effect (Lah et al., 2007). Silymarin has been shown to have an inhibitory effect on ornithine decarboxylase activity (Katiyar et al., 2007). Silymarin suppresses proliferation of hepatocellular carcinoma (HCC) by inhibiting β-catenin accumulation and, hence, its target genes for cyclin D1 and c-Myc; and this may be the underlying mechanism of its antiproliferative effect. In conclusion, it has been found that the mechanism by which silymarin exhibits its growth inhibitory effect on human hepatocellular carcinoma cells in vitro is inhibiting cell proliferation and inducing apoptosis (Ramakrishnan et al., 2009).
Rosemary phytochemicals, such as carnosic acid, have inhibitory effects on anti-cancer drug efflux transporter P-glycoprotein and may become useful to enhance the efficacy of cancer chemotherapy (Nabekura et al., 2010). The leaves of rosemary (Rosmarinus officinalis) are commonly used as a spice in cooking. However, because of the presence of phenolic diterpenes and triterpenes with strong antioxidative activity, interest has grown in using rosemary as a natural antioxidant in foods. In addition to antioxidative activities, rosemary phytochemicals are reported to have antimicrobial, anti-inflammatory, and anticancer properties (Nabekura et al., 2010). Interestingly, inhibitory effects of several dietary chemopreventive and antioxidative phytochemicals, such as curcumin, RSV, tannic acid, quercetin, kaempferol, EGCG and other tea catechins, on the function of P-glycoprotein have been reported (Kitagawa et al., 2007). Natural antioxidative and chemopreventive rosemary phytochemicals, carnosic acid, carnosol and ursolic acid, have inhibitory effects on P-glycoprotein and the potential to cause food–drug interactions (Nabekura et al., 2010).
Triptolide/PG490, an extract of the Chinese herb Tripterygium wilfordii Hook F, is a potent anti-inflammatory agent that also possesses anti-cancer activity. Triptolide is a potent inhibitor of colon cancer proliferation and migration in vitro. The down-regulation of multiple cytokine receptors, in combination with inhibition of COX-2 and VEGF and positive cell-cycle regulators, may contribute to the antimetastatic action of this herbal extract (Johnson et al., 2011). Herbal extracts may modify biologic responses to classic chemotherapy agents and influence multiple signaling pathways; their actions likely include antiproliferative, antiangiogenic, proapoptotic and/or antimetastatic effects (HemaIswarya and Doble, 2006). Very recently, antioxidative and antiproliferative activity of different horsetail (Equisetum arvense L.) extracts has been investigated. Extracts inhibited cell growth that was dependent on cell line, type of extract and extract concentration. Ethyl acetate extract exhibited the most prominent antiproliferative effect, without inducing any cell growth stimulation on human tumour cell lines (Cetojević-Simin et al., 2010).
RSV functions as a fungicide produced by the plant itself to ward off potentially lethal organisms and counteract environmental stress. RSV exhibits antioxidant, anti-inflammatory and antiaging action and displays chemopreventive effects in a number of biological systems (Zamin et al., 2009). In spite of the anti-cancer efficacy of RSV in preclinical models, its low bioavailability remains enigmatic and elusive. In order to explore whether RSV metabolites exert antitumour properties, some major human sulfated conjugates have recently been tested against human breast cancer cells (Matés et al., 2009a). RSV affected the growth of a human cholangiocarcinoma cell line (Lentini et al., 2010) and reduced formation of preneoplastic lesions (aberrant crypt foci) in rats, to decrease the incidence and size of tumours in the 1,2-dimethylhydrazine-induced model of colon cancer in rats and to prevent the formation of colon and small intestinal tumours in mice (Paul et al., 2010). The anticarcinogenesis activity of RSV was first shown in a pioneering study by Jang and Pezzuto (1999), who reported that RSV was effective in all the three major stages (initiation, promotion and progression) of carcinogenesis. RSV suppresses the proliferation of a variety of human cancer cells in vitro, including glioma cells (Kundu and Surh, 2008) and HCC (Bishayee et al., 2010). RSV is a potent antioxidant because of its ability to scavenge free radicals, spare and/or regenerate endogenous antioxidants, that is, GSH and α-tocopherol, and to promote the activities of a variety of antioxidant enzymes (Figure 4.2). The antioxidant property of RSV is also attributed to its ability to promote the activities of a variety of antioxidant enzymes. When intraperitoneally administered in rats, it was found to dose-dependently increase SOD, catalase and peroxidase activities in the brains of healthy rats (Mokni et al., 2007). The long-term exposure of human lung fibroblasts to RSV results in a highly specific upregulation of MnSOD (Matés et al., 2009a). In human lymphocytes, RSV increased GSH levels and the activity of glutathione peroxidase, GST and glutathione reductase. Depending on the concentration and cell type, RSV can also act as a prooxidant molecule, this effect being Cu(II)-dependent (De la Lastra and Villegas, 2007). Such a prooxidant effect could be an important action mechanism for its anti-cancer and proapoptotic properties. Compared with normal cells, cancer cells have been shown to contain elevated levels of copper and, hence, might be more sensitive to the prooxidant and cell-damaging effects of RSV. Therefore, DNA damage induced by RSV in the presence of Cu(II) might be an important pathway through which cancer cells can be killed while normal cells survive. In rats and mice, RSV toxicity is minimal, and even actively proliferating tissues are not adversely affected. The high systemic levels of RSV conjugate metabolites would warrant investigation of their potential cancer chemopreventive properties. Interestingly, RSV and quercetin, chronically administered, presented a strong synergism in inducing senescence-like growth arrest (Zamin et al., 2009). These results suggest that the combination of polyphenols can potentialise their antitumoural activity. Gliomas are the most malignant of primary tumours that affect the brain and nervous system and carry the worst clinical prognosis in both adults and children. RSV-induced inhibition of catalase might represent an additional tool for the current clinical armamentarium that might increase glioma cell kill, leading to improved survival time for patients. At present, RSV is undergoing various Phase I and Phase II interventional trials. Results from the most recent studies performed in rat and human glioma cell lines suggest that the use of RSV in combination with other bioactive food components, such as quercetin and sulforaphane, might be a viable approach for the treatment of human glioma (Gagliano et al., 2010).
Pterostilbene (trans-3,5-dimethoxy-4′-hydroxystilbene), a structurally similar compound to RSV found in blueberries, has been also shown to exhibit significant effects against cell proliferation, invasion and metastasis (Pan et al., 2007). Overall analyses indicated that pterostilbene reduced colon tumour multiplicity of non-invasive adenocarcinomas, lowered proliferating cell nuclear antigen and downregulated the expression of β-catenin and cyclin D1. In HT-29 cells, pterostilbene reduced the protein levels of β-catenin, cyclin D1 and c-Myc, altered the cellular localisation of β-catenin and inhibited the phosphorylation of p65 (Paul et al., 2010). Cyclin D1 is a very well-known cell-cycle protein targeted by β-catenin and is known to be overexpressed in colonic tumours. c-Myc is yet another important protein for cell proliferation regulated by β-catenin and Wnt pathway. Pterostilbene alone or in combination with other known chemopreventive agents can be of great importance for colon cancer prevention (Paul et al., 2010). In this regard, eugenol, a natural phenolic constituent of clove oil, cinnamon, basil and nutmeg, used primarily as a food flavouring agent has been documented to exhibit antiproliferative effects in diverse cancer cell lines as well as in B16 melanoma xenograft model. Recent studies have indicated that some of these phytochemicals are substrates and modulators of specific members of the superfamily of ABC transporting proteins. Such interactions may have implications on the pharmacokinetics of xenobiotics and the possible role of phytochemicals in the reversal of multi-drug resistance in cancer chemotherapy (Li et al., 2010).
The plant-derived anti-cancer agents are commonly classified into one of four major classes: vinca alkeloids (vinblastin, vincristine and vindesine), epipodophyllotoxins (etoposide and teniposides), texanes (paclitaxel and docetaxel) and camtothecins (camptothecin and irinotecan) (Siddiqui et al., 2009). Plant alkaloids are used as chemotherapeutic agents due to their capability to depolymerise the microtubules, inhibiting cell division. Vinca alkaloids are isolated from Vinca rosea L. and they are potent microtubule destabilising agents, first recognised for their myelosuppressive effects. Another alkaloid used as a chemotherapy drug for some types of cancer is taxol. It is mainly used to treat ovarian, breast and non-small cell lung cancer. Moreover, capsaicin, the major pungent ingredient in red peppers, has a profound antiproliferative effect on prostate cancer (Mori et al., 2006). Tylophorine downregulates cyclin A2, which plays an important role in G1 arrest in carcinoma cells. In addition, camptothecin, a pentacyclic alkaloid isolated from Camptotheca acuminata Decne, was reported to possess an interesting antitumour activity (Staker et al., 2002).
Finally, combination of carotenoids and myo-inositol was found to prevent HCC development in patients with chronic viral hepatitis and cirrhosis (Nishino, 2009). Supplemental β-caroteno has been shown to reduce precancerous lesions of the oral cavity and cervix, but not of the lung (Mayne, 1996). Agents that suppress tumour formation, such as the vitamin A metabolite all-trans retinoic acid and the isothiocyanate sulforaphane in an ultraviolet (UV) model, can block AP-1 signaling (Dickinson et al., 2009). Another naturally occurring chemical, perillyl alcohol, also suppressed tumour formation in the UV model, and this effect correlated with AP-1 suppression. A study including combination of dietary elagic acid or calcium D-glucarate plus topical RSV, or grape seed extract and dietary grape seed extract plus topical RSV showed synergistic effects in reducing DMBA-induced hyperplasia (Clifford and DiGiovanni, 2010). A previous study showed that oral green tea polyphenols can suppress both UV-induced and chemically induced skin tumours in mice (Kowalczyk et al., 2010).
Fruit and vegetable consumption has been inversely associated with the risk of many pathological diseases, with the beneficial effects attributed to a variety of protective phytonutrients (Kawashima et al., 2007). The mechanisms explaining this correlation have not been fully elucidated. Regardless, a consensus exists that a diet rich in fruit and vegetables is beneficial for health in preventing coronary heart disease and some forms of cancer. The nutrients responsible for the protective action are not known, but vitamins, antioxidants and flavonoids are among the likely candidates (Matés et al., 2009a). Dietary supplements are generally used to increase plasma levels of these compounds. Similar results can be achieved also by increasing the proportion of vegetables and fruit in the diet. Disease prevention by dietary factors or foods consumed in small quantities and exotic plants is likely to represent one of the strategies to reduce the risk of development of malignancy in humans. The identification of new molecules able to reduce proliferative and metastatic potential of cancer cells is the goal of the newborn differentiation therapy. Noteworthy examples of diet-derived substances that have been shown to reduce experimental carcinogenesis are I3C from cruciferous vegetables, curcumin from the root of curcuma, EGCG from tea and RSV from red wine. Vegetables and fruits contain fibre, vitamins, minerals and a variety of bioactive compounds, such as carotenoids, flavonoids, indoles and sterols, all of which could account for this protective effect. On the other hand, spices, herbs and rare plants are gaining uses as pharmacologic and therapeutic agents. A better understanding of the actions of phytochemicals will facilitate its use in more specific clinical trials, may allow for synthetic derivatives to be engineered (equally effective but less toxic) and potentially offer insight into additional therapeutic uses for their antioxidant potential (Johnson et al., 2011).
The chemopreventive effects of dietary phytochemicals on malignant tumours have been studied extensively because of a relative lack of toxicity. To achieve desirable effects, however, treatment with a single agent mostly requires high doses. Therefore, studies on effective combinations of phytochemicals at relatively low concentrations might contribute to chemopreventive strategies. The field of chemoprevention has expanded to include nanotechnology as a novel approach to deliver packaged chemopreventive agents in a manner which allows them to be delivered selectively to the target tissues. For example, Mukhtar and colleagues recently reported on the bioavailability of EGCG which had been packaged into nanoparticles. Results showed that EGCG delivered by nanoparticles maintained the efficacy of EGCG both as an antiangiogenic and proapoptotic agent (Siddiqui et al., 2009). These results indicated that nano-chemoprevention can provide a new approach to avoiding systemic toxicity and increasing bioavailability. Similarly, packaging RSV into solid lipid nanoparticles has been reported to improve intracellular delivery of RSV and to reduce RSV toxicity. The application of lipid- or polymer-based nano-particles or nano-shells for improved delivery of chemopreventive or chemotherapeutic agents has facilitated the delivery of agents to selective tissues and may serve to lessen systemic toxicity by reducing the amount of agent needed and/or limiting the exposure to the body (Teskac and Kristl, 2010).
Of particular significance, heyneanol, a tetramer of RSV, has comparable or better antitumour efficacy than RSV in a mouse lung cancer model (Jeong et al., 2011). The current interest in phytochemicals has been driven primarily by epidemiological studies. However, to establish conclusive evidence for the effectiveness of dietary phytochemicals in disease prevention, it is useful to better define the bioavailability of these bioactive compounds, so that their biological activity can be evaluated. The bioavailability appears to differ greatly among the various plant compounds, and the most abundant ones in our diet are not necessarily those that have the best bioavailability profile. The evaluation of the bioavailability of phytochemicals has recently been gaining increasing interest as the food industries are continually involved in developing new products, defined as functional food, by virtue of the presence of specific compounds. Despite the increasing amount of data available, definitive conclusions on bioavailability of most bioactive copmpounds are difficult to obtain and further studies are necessary. At least four critical lines of research should be explored to gain a clear understanding of the health beneficial effects of dietary phytochemicals (D’Archivio et al., 2010):
Abere, T.A., Okoto, P.E. and Agoreyo, F.O. (2010) Antidiarrhoea and toxicological evaluation of the leaf extract of Dissotis rotundifolia Triana (Melastomataceae). BMC Complementary and Alternative Medicine, 10, 71.
Abu Bakar, M.F., Mohamad, M., Rahmat, A., Burr, S.A. and Fry, J.R. (2010) Cytotoxicity, cell cycle arrest, and apoptosis in breast cancer cell lines exposed to an extract of the seed kernel of Mangifera pajang (bambangan). Food Chemistry and Toxicology, 48, 1688–1697.
Abu Bakar, M.F., Mohamed, M., Rahmat, A., Burr, S.A. and Fry, J.R. (2010) Cytotoxicity and polyphenol diversity in selected parts of Mangifera pajang and Artocarpus odoratissimus fruits. Nutrition and Food Science, 40, 29–38.
Adams, L.S., Phung, S., Yee, N., Seeram, N.P., Li, L. and Chen, S. (2010) Blueberry phytochemicals inhibit growth and metastatic potential of MDA-MB-231 breast cancer cells through modulation of the phosphatidylinositol 3-kinase pathway. Cancer Research, 70, 3594–35605.
Adlercreutz, H. and Mazur, W. (1997) Phyto-oestrogens and western diseases. Annals of Internal Medicine, 29, 95–120.
Adom, K.K. and Liu, R.H. (2002) Antioxidant activity of grains. Journal of Agriculture and Food Chemistry, 50, 6182–6187.
Adom, K.K., Sorrells, M.E. and Liu, R.H. (2005) Phytochemicals and antioxidant activity of milled fractions of different wheat varieties. Journal of Agriculture and Food Chemistry, 53, 2297–2306.
Aggarwal, B.B. and Shishodia, S. (2006) Molecular targets of dietary agents for prevention and therapy of cancer. Biochemical Pharmacology, 71, 1397–1421.
Aggarwal, S., Ichikawa, H., Takada, Y., Sandur, S.K., Shishodia, S. and Aggarwal, B.B. (2006) Curcumin (diferuloylmethane) down-regulates expression of cell proliferation and antiapoptotic and metastatic gene products trough suppression of IB kinase and Akt activation. Molecular Pharmacology, 69, 1195–1206.
Anderson, J.W. (2002) Whole-grains intake and risk for coronary heart disease. In L. Marquart, J.L. Slavin and R.G. Fulcher (eds) Whole-Grain Foods in Health and Disease, Eagan Press, St Paul (MN), pp. 187–200.
Andriambeloson, E., Kleschyov, A.L., Muller, B., Beretz, A., Stoclet, J.C. and Andriantsitohaina, R. (1997) Nitric oxide production and endothelium-dependent vasorelaxation induced by wine polyphenols in rat aorta. British Journal of Pharmacology, 120, 1053–1058.
Attoub, S., Hassan, A.H., Vanhoecke, B., Iratni, R., Takahashi, T., Gaben, A.M., Bracke, M., Awad, S., John, A., Kamalboor, H.A., Al, Sultan, M.A., Arafat, K., Gespach, C. and Petroianu, G. (2011) Inhibition of cell survival, invasion, tumor growth and histone deacetylase activity by the dietary flavonoid luteolin in human epithelioid cancer cells. European Journal of Pharmacology, 651, 18–25.
Awika, J.M., McDonough, C.M., Rooney, L.W. (2005) Decorticating sorghum to concentrate healthy phytochemicals. Journal of Agriculture and Food Chemistry, 53, 6230–6234.
Ayanwuyi, L.O., Yaro, A.H. and Abodunde, O.M. (2010) Analgesic and anti-inflammatory effects of the methanol stem bark extract of Prosopis africana. Pharmacuetical Biolology, 48, 296–299.
Banerjee, S., Li, Y., Wang, Z. and Sarkar, F.H. (2008) Multi-targeted therapy of cancer by genistein. Cancer Letters, 269, 226–242.
Baur, J.A. and Sinclair, D.A. (2006) Therapeutic potential of resveratrol: the in vivo evidence. Nature Reviews Drug Discovery, 5, 493–506.
Bazzano, L.A. (2008) Effects of soluble dietary fiber on low-density lipoprotein cholesterol and coronary heart disease risk. Current Atherosclerosis Reports, 10, 473–477.
Berliner, J.A. and Heinecke, J.W. (2006) The role of oxidized lipoproteins in atherogenesis. Free Radical Biology and Medicine, 20, 707–727.
Bharti, A.C., Takada, Y. and Aggarwal, B.B. (2004) Curcumin (diferuloylmethane) inhibits receptor activator of NF-kappaB ligand-induced NF-kappaB activation in osteoclast precursors and suppresses osteoclastogenesis. Journal of Immunology, 172, 5940–5947.
Bishayee, A., Ahmed, S., Brankov, N. and Perloff, M. (2011) Triterpenoids as potential agents for the chemoprevention and therapy of breast cancer. Frontiers in Bioscience, 16, 980–996.
Bishayee, A., Politis, T. and Darvesh, A.S. (2010) Resveratrol in the chemoprevention and treatment of hepatocellular carcinoma. Cancer Treatment Reviews, 36, 43–53.
Boots, A.W., Kubben, N., Haenen, G.R. and Bast, A. (2003) Oxidized quercetin reacts with thiols rather than with ascorbate: implication for quercetin supplementation. Biochemical and Biophysical Research Communications, 308, 560–565.
Calabrese, V., Cornelius, C., Mancuso, C., Lentile, R., Stella, A.M. and Butterfield, D.A. (2010) Redox homeostasis and cellular stress response in aging and neurodegeneration. Methods in Molecular Biology, 610, 285–308.
Carlsen, H., Myhrstad, M.C., Thoresen, M., Moskaug, J.O. and Blomhoff, R. (2003) Berry intake increases the activity of the gamma-glutamylcysteine synthetase promoter in transgenic reporter mice. Journal of Nutrition, 133, 2137–2140.
Cetojević-Simin, D.D., Canadanović-Brunet, J.M., Bogdanović, G.M., Djilas, S.M., Cetković, G.S., Tumbas, V.T. and Stojiljković, B.T. (2010) Antioxidative and antiproliferative activities of different horsetail (Equisetum arvense L.) extracts. Journal of Medicinal Food, 13, 452–459.
Clifford, J.L. and DiGiovanni, J. (2010) The promise of natural products for blocking early events in skin carcinogenesis. Cancer Prevention Research, 3, 132–135.
Conte, C., Floridi, A., Aisa, C., Piroddi, M. and Galli, F. (2004) Gamma-tocotrienol metabolism and antiproliferative effect in prostate cancer cells. Annals of the New York Academy of Sciences, 1031, 391–394.
D’Archivio, M., Filesi, C., Varì, R., Scazzocchio, B. and Masella, R. (2010) Bioavailability of the polyphenols: status and controversies. International Journal of Molecular Science, 11, 1321–1342.
Das, S. and Das, D.K. (2007) Resveratrol: a therapeutic promise for cardiovascular diseases. Recent Patents on Cardiovascular Drug Discovery, 2, 133–138.
De la Lastra, C.A and Villegas, I. (2007) Resveratrol as an antioxidant and pro-oxidant agent: mechanisms and clinical implications. Biochemical Society Transactions, 35, 1156–1160.
Deep, G. and Agarwal, R. (2007) Chemopreventive efficacy of silymarin in skin and prostate cancer. Integrative Cancer Therapies, 6, 130–145.
Dickinson, S.E., Melton, T.F., Olson, E.R., Zhang, J., Saboda, K. and Bowden, G.T. Inhibition of activator protein-1 by sulforaphane involves interaction with cysteine in the cFos DNA-binding domain: implications for chemoprevention of UVB-induced skin cancer. Cancer Research, 69, 7103–7110.
Diebolt, M., Bucher, B. and Andriantsitohaina, R. Wine polyhenols decrease blood pressure, improve NO vasodilatation and induce gene expression. Hypertension, 38, 159–165.
Duthie, S.J., Jenkinson, A.M., Crozier, A., Mullen, W., Pirie, L., Kyle, J., Yap, L.S., Christen, P. and Duthie, G.G. (2006) The effects of cranberry juice consumption on antioxidant status and biomarkers relating to heart disease and cancer in healthy human volunteers. European Journal of Nutrition, 45, 113–122.
Dykes, L. and Rooney, L.W. (2006) Sorghum and millet phenols and antioxidants. Journal of Cereal Science, 44, 236–251.
Elbling, L., Weiss, R.M., Teufelhofer, O., Uhl, M., Knasmueller, S., Schulte-Hermann, R., Berger, W. and Micksche, M. (2005) Green tea extract and (−)-epigallocatechin-3-gallate, the major tea catechin, exert oxidant but lack antioxidant activities. FASEB Journal, 19, 807–809.
El-Najjar, N., Chatila, M., Moukadem, H., Vuorela, H., Ocker, M., Gandesiri, M., Schneider-Stock, R. and Gali-Muhtasib, H. (2010) Reactive oxygen species mediate thymoquinone-induced apoptosis and activate ERK and JNK signaling. Apoptosis, 15, 183–195.
Ernst, E. and Pittler, M.H. (1999) Ginkgo biloba for dementia: A systematic review of double-blind placebo-controlled trials. Clinical Drug Investigation, 17, 301–308.
Fang, M.Z., Wang, Y., Ai, N., Hou, Z., Sun, Y., Lu, H., Welsh, W. and Yang, C.S. (2003) Tea polyphenol (-)-epigallocatechin-3-gallate inhibits DNA methyltransferase and reactivates methylation-silenced genes in cancer cell lines. Cancer Research, 63, 7563–7570.
Farrar, J.L., Hartle, D.K., Hargrove, J.L. and Greenspan, P. (2008) A novel nutraceutical property of select sorghum (Sorghum bicolor) brans: inhibition of protein glycation. Phytotherapy Research, 22, 1052–1056.
Fletcher, R.H. and Fairfield, K.M. (2002) Vitamins for chronic disease prevention in adults: Clinical applications. Journal fo the Aerican Medical Association (JAMA), 187, 3127–3129.
Fong, D., Yeh, A., Naftalovich, R., Choi, T.H. and Chan, M.M. (2010) Curcumin inhibits the side population (SP) phenotype of the rat C6 glioma cell line: towards targeting of cancer stem cells with phytochemicals. Cancer Letters, 293, 65–72.
Frescas, D., Valenti, L. and Accili, D. (2005) Nuclear trapping of the forkhead transcription factor FoxO1 via Sirt-dependent deacetylation promotes expression of glucogenetic genes. Journal of Biological Chemistry, 280, 20589–20595.
Fuster, V., Badimon, J.J. and Chesebro, J.H. The pathogenesis of coronary artery disease and the acute coronary syndromes. New England Journal of Medicine, 326, 242–250.
Gagliano, N., Aldini, G., Colombo, G., Rossi, R., Colombo, R., Gioia, M., Milzani, A. and Dalle-Donne, I. (2010) The potential of resveratrol against human gliomas. Anticancer Drugs, 21, 140–150.
Gerhäuser, C., Klimo, K., Heiss, E., Neumann, I., Gamal-Eldeen, A., Knauft, J., Liu, G.Y., Sitthimonchai, S. and Frank, N. (2003) Mechanism-based in vitro screening of potential cancer chemopreventive agents. Mutatation Research, 523–524, 163–172.
Gilbert, E.R. and Liu, D. (2010) Flavonoids influence epigenetic-modifying enzyme activity: structure – function relationships and the therapeutic potential for cancer. Current Medicinal Chemistry, 17, 1756–1768.
Haddad, J.J. (2002) Antioxidant and prooxidant mechanisms in the regulation of redox(y)-sensitive transcription factors. Cellular Signalling, 14, 879–897.
Hallikainen, M.A., Sarkkinen, E.S. and Uusitupa, M.I.J. (2000) Plant stanols esters affect serum cholesterol concentrations of hypercholesterolemic men and women in a dose-dependent manner. Journal of Nutrition, 130, 767–776.
Hara, Y. (1992) The effect of tea polyphenols on cardiovacular disease. Preventive Medicine, 21, 333.
Hasler, C.M., Bloch, A.S., Thomson, C.A., Enrione, E. and Manning, C. (2004) Position of the American Dietetic Association: Functional foods. Journal of the American Dietetetic Association, 104, 814–826.
Hassel, C.A. (1998) Animal models: new cholesterol raising and lowering nutrients. Current Opinion in Lipidology, 9, 7–10.
Hayek, T., Fuhrman, B., Vaya, J., Rosenblat, M., Belinky, P., Coleman, R., Elis, A. and Aviram, M. (1997) Reduced progression of atherosclerosis in apolipoprotein E-deficient mice following consumption of red wine, or its polyphenols quercetin or catechin, is associated with reduced susceptibility of LDL to oxidation and aggregation. Arteriosclerosis, Thrombosis, and Vascular Biology, 17, 2744–2752.
Heijnen, C.G.M., Haenen, G.R.M.M., Oostveen, R.M., Stalpers, E.M. and Bas, A. (2002) Protection of flavonoids against lipid peroxidation: structure activity relationship revisited. Free Radicical Research, 36, 575–581.
HemaIswarya, S. and Doble, M. (2006) Potential synergism of natural products in the treatment of cancer. Phytotherapy Research, 20, 239–249.
Hertog, M.G.L, Feskens, E.J.M. and Kromhout, D. (1997) Antioxidant flavonols and coronary heart disease risk. The Lancet, 349, 699.
Hossain, M.M. (2011) Therapeutic orchids: traditional uses and recent advances--an overview. Fitoterapia, 82, 102–140.
Howells, L.M., Mitra, A. and Manson, M.M. (2007) Comparison of oxaliplatin- and curcumin-mediated antiproliferative effects in colorectal cell lines. International Journal of Cancer, 121, 175–183.
Hsieh, T.C. and Wu, J.M. (2009) Targeting CWR22Rv1 prostate cancer cell proliferation and gene expression by combinations of the phytochemicals EGCG, genistein and quercetin. Anticancer Research, 29, 4025–4032.
Jang, M. and Pezzuto, JM. Cancer chemopreventive activity of resveratrol. (1999) Drugs Under Experimental and Clinical Research, 25, 65–77.
Jayaraman, P., Sakharkar, M.K., Lim, C.S., Tang, T.H. and Sakharkar, K.R. (2010) Activity and interactions of antibiotic and phytochemical combinations against Pseudomonas aeruginosa in vitro. International Journal of Biolological Sciences, 6, 556–568.
Jeong, S.J., Koh, W., Lee, E.O., Lee, H.J., Lee, H.J., Bae, H., Lü, J. and Kim, S.H. (2011) Antiangiogenic phytochemicals and medicinal herbs. Phytotherapy Research, 25, 1–10.
Johnson, S.M., Wang, X. and Evers, B.M. (2011) Triptolide inhibits proliferation and migration of colon cancer cells by inhibition of cell cycle regulators and cytokine receptors. Journal of Surgical Research, doi:10.1016/j.jss.2009.07.002.
Jones, J.M. and Anderson, J.W. (2008) Grain foods and health: a primer for clinicians. Physician and Sportsmedicine, 36, 18–33.
Kang, J., Badger, T.M., Ronis, M.J. and Wu, X. (2010) non-isoflavone phytochemicals in soy and their health effects. Journal of Agricultural Food Chemistry, 58, 8119–8133.
Katiyar, S.K., Korman, N.J., Mukhtar, H. and Agarwal, R. (1997) Protective effects of silymarin against photocarcinogenesis in a mouse skin model. Journal of the National Cancer Institute, 89, 556–565.
Kaur, M., Agarwal, C. and Agarwal, R. (2009) Anticancer and cancer chemopreventive potential of grape seed extract and other grape-based products. Journal of Nutrition, 139, 1806 S–1812 S.
Kawanishi, S., Oikawa, S. and Murata, M. (2005) Evaluation for safety of antioxidant chemopreventive agents. Antioxidants and Redox Signaling, 7, 1728–1739.
Kawashima, A., Madarame, T., Koike, H., Komatsu, Y. and Wise, J.A. (2007) Four week supplementation with mixed fruit and vegetable juice concentrates increased protective serum antioxidants and folate and decreased plasma homocysteine in Japanese subjects. Asia Pacific Journal of Clinical Nutrition, 16, 411–421.
Kim, E.C., Min, J.K., Kim, T.Y., Lee, S.J., Yang, H.O., Han, S., Kim, Y.M. and Kwon, Y.G. (2005) [6]-Gingerol, a pungent ingredient of ginger, inhibits angiogenesis in vitro and in vivo. Biochemical and Biophysical Research Communications, 335, 300–308.
King, A.A., Shaughnessy, D.T., Mure, K., Leszczynska, J., Ward, W.O., Umbach, D.M., Xu, Z., Ducharme, D., Taylor, J.A., Demarini, D.M. and Klein, C.B. (2007) Antimutagenicity of cinnamaldehyde and vanillin in human cells: global gene expression and possible role of DNA damage and repair. Mutation Research, 616, 60–69.
Kitagawa, S., Nabekura, T., Nakamura, Y., Takahashi, T. and Kashiwada, Y. (2007) Inhibition of P-glycoprotein function by tannic acid and pentagalloylglucose. Journal of Pharmacy and Pharmacology, 59, 965–969.
Kowalczyk, M.C., Kowalczyk, P., Tolstykh, O., Hanausek, M., Walaszek, Z. and Slaga, T.J. (2010) Synergistic effects of combined phytochemicals and skin cancer prevention in SENCAR mice. Cancer Prevention Research, 3, 170–178.
Kukongviriyapan, U., Luangaram, S., Leekhaosoong, K., Kukongviriyapan, V. and Preeprame, S. (2007) Antioxidant and vascular protective activities of Cratoxylum formosum, Syzygium gratum and Limnophila aromatica. Biolological and Pharmacuetical Bulletin, 30, 661–666.
Kundu, J.K. and Surh, Y.J. Cancer chemopreventive and therapeutic potential of resveratrol: mechanistic perspectives. Cancer Letters, 269, 243–261.
Kunwar, R.M., Shrestha, K.P. and Bussmann, R.W. (2010) Traditional herbal medicine in far-west Nepal: a pharmacological appraisal. Journal of Ethnobiolology and Ethnomedicine, 6, 35.
Kuriyama, S. (2008) The relation between green tea consumption and cardiovascular disease as evidenced by epidemiological studies. Journal of Nutrition, 138, 1548S–1553S.
Kusari, S., Zühlke, S. and Spiteller, M. (2011) Chemometric evaluation of the anti–cancer pro-drug podophyllotoxin and potential therapeutic analogues in juniperus and podophyllum species. Phytochemical Analysis, 22, 128–143.
Kwon, Y.W., Masutani, H., Nakamura, H., Ishii, Y. and Yodoi, J. (2003) Redox regulation of cell growth and cell death. Biological Chemistry, 384, 991–996.
Lah, J.L., Cui, W. and Hu, K.Q. (2007) Effects and mechanisms of silibinin on human hepatoma cell lines. World Journal of Gastroenterology, 13, 5299–5305.
Lambert, J.D., Hong, J., Yang, G., Liao, J. and Yang, C.S. (2005) Inhibition of carcinogenesis by polyphenols: evidence from laboratory investigations. American Journal of Clinical Nutrition, 81, 284S–291S.
Lampe, J.W. (2007) Diet, genetic polymorphisms, detoxification, and health risks. Alternative Therapies in Health and Medicine, 13, S108–111.
Le Marchand, L. (2002) Cancer preventive effects of flavonoids. A review. Biomedicine and Pharmacotherapy, 56, 296–301.
Lee, H.S., Seo, E.Y., Kang, N.E. and Kim, W.K. (2008) [6]-Gingerol inhibits metastasis of MDA-MB-231 human breast cancer cells. Journal of Nutritional Biochemistry, 19, 313–319.
Lee, J., Shim, Y. and Zhu, B.T. Mechanisms for the inhibition of DNA methyltransferases by tea catechins and bioflavonoids. Molecular Pharmacology, 68 (4), 1018–1030.
Lentini, A., Tabolacci, C., Provenzano, B., Rossi, S. and Beninati, S. (2010) Phytochemicals and protein-polyamine conjugates by transglutaminase as chemopreventive and chemotherapeutic tools in cancer. Plant Physiology and Biochemistry, 48, 627–633.
Li, J., Zhang, D., Stoner, G.D. and Huang, C. (2008) Differential effects of black raspberry and strawberry extracts on BaPDE-induced activation of transcription factors and their target genes. Molecular Carcinogenesis, 47, 286–294.
Li, Y., Revalde, J.L., Reid, G. and Paxton, J.W. (2010) Interactions of dietary phytochemicals with ABC transporters: possible implications for drug disposition and multidrug resistance in cancer. Drug Metabolism Reviews, 42: 590–611.
Lin, Y. and Lin, J. (1997) (–)-Epigallocatechin-3-gallate blocks the induction of nitric oxide synthase by down-regulating lipopolysaccharide-induced activity of transcription factor nuclear factor-kappaB. Molecular Pharmacology, 52, 465–472.
Liu, R.H. (2004) Potential synergy of phytochemicals in cancer prevention: mechanism of action. Journal of Nutrition, 134, 3479S–3485S.
Loots, du T., van der Westhuizen, F.H. and Botes, L. (2007) Aloe ferox leaf gel phytochemical content, antioxidant capacity, and possible health benefits. Journal of Agricultural and Food Chemistry, 55, 6891–6896.
Ma, H., He, X., Yang, Y., Li, M., Hao, D. and Jia, Z. (2011) The genus Epimedium: An ethnopharmacological and phytochemical review. Journal of Ethnopharmacology, doi:10.1016/j.jep.2011.01.001.
Magee, P.J. and Rowland, I.R. (2004) Phyto-oestrogens, their mechanism of action: current evidence for a role in breast and prostate cancer. British Journal of Nutrition, 91, 513–531.
Manach, C., Scalbert, A., Morand, C., Rémésy, C. and Jimenez, L. (2004) Polyphenols: food sources and bioavailability. American Journal of Clinical Nutrition, 79, 727–747.
Manikandan, P., Murugan, R.S., Priyadarsini, R.V., Vinothini, G. and Nagini, S. (2010) Eugenol induces apoptosis and inhibits invasion and angiogenesis in a rat model of gastric carcinogenesis induced by MNNG. Life Sciences, 86, 936–941.
Manson, M.M., Foreman, B.E., Howells, L.M. and Moiseeva, E.P. (2007) Determining the efficacy of dietary phytochemicals in cancer prevention. Biochemical Society Transactions, 35, 1358–1363.
Masella, R., Di Benedetto, R., Varì, R., Filesi, C. and Giovannini, C. (2005) Novel mechanisms of natural antioxidant compounds in biological systems: involvement of glutathione and glutathione-related enzymes. Journal of Nutrional Biochemistry, 16, 577–586.
Masibo, M. and He, Q. Major mango polyphenols and their potential significance to human health. Comprehensive Reviews in Food Science and Food Safety, 7, 309–319.
Matés, J.M., Pérez-Gómez, C., Núñez de Castro, I., Asenjo, M. and Márquez, J. (2002) Glutamine and its relationship with intracellular redox status, oxidative stress and cell proliferation/death. International Journal of Biochemistry and Cell Biology, 34, 439–458.
Matés, J.M., Segura, J.A., Alonso, F.J. and Márquez, J. (2008) Intracellular redox status and oxidative stress: implications for cell proliferation, apoptosis, and carcinogenesis. Archives of Toxicology, 82, 273–299.
Matés, J.M., Segura, J.A., Alonso, F.J. and Márquez, J. (2009) Natural antioxidants, therapeutic prospects for cancer and neurological diseases. Mini-Reviews in Medicinal Chemistry, 9, 1202–1214.
Matés, J.M., Segura, J.A., Alonso, F.J. and Márquez, J. (2006) Pathways from glutamine to apoptosis. Frontiers in Bioscience, 11, 3164–3180.
Matés, J.M., Segura, J.A., Alonso, F.J. and Márquez, J. (2010) Roles of dioxins and heavy metals in cancer and neurological diseases using ROS-mediated mechanisms. Free Radical Biology and Medicine, 49, 1328–1341.
Matés, J.M., Segura, J.A., Campos-Sandoval, J.A., Lobo, C., Alonso, L., Alonso, F.J. and Márquez, J. (2009) Glutamine homeostasis and mitochondrial dynamics. International Journal of Biochemistry and Cell Biology, 41, 2051–2061.
Matés, J.M. (2000) Effects of antioxidant enzymes in the molecular control of reactive oxygen species toxicology. Toxicology, 153, 83–104.
Mattson, M.P. and Meffert, M.K. (2006) Roles for NF-kappaB in nerve cell survival, plasticity, and disease. Cell Death and Differentiaion, 13, 852–860.
Mattson, M.P. (2008) Dietary factors, hormesis and health. Ageing Research Review, 7, 43–48.
Mayne, S.T. (1996) Beta-carotene, carotenoids, and disease prevention in humans. FASEB Journal, 10, 690–701.
McWalter, G.K., Higgins, L.G., McLellanm L.I., Henderson, C.J., Song, L., Thornalley, P.J., Itoh, K., Yamamoto, M. and Hayes, J.D. (2004) Transcription factor Nrf2 is essential for induction of NAD(P)H:quinone oxidoreductase 1, glutathione S-transferases, and glutamate cysteine ligase by broccoli seeds and isothiocyanates. Journal of Nutrition, 134, 3499S–3506S.
Mehta, R.G., Murillo, G., Naithani, R. and Peng, X. (2010) Cancer chemoprevention by natural products: how far have we come? Pharmaceutical Research, 27, 950–961.
Michalska, M., Gluba, A., Mikhailidis, D.P., Nowak, P., Bielecka-Dabrowa, A., Rysz, J. and Banach, M. The role of polyphenols in cardiovascular disease. Medical Science Monitor, 16, RA110–119.
Middleton, E.J.R., Kandaswami, C. and Theoharides, T.C. (2000) The effect of plant flavonoids on mammalian cells: implications for inflammation, heart disease and cancer. Pharmacology Review, 52, 673–751.
Miura, Y., Chiba, T., Tomita, I., Koizumi, H., Miura, S., Umegaki, K., Hara, Y., Ikeda, M. and Tomita, T. (2001) Tea catechins prevent the development of atherosclerosis in apoprotein E-deficient mice. Journal of Nutrition, 131, 27–32.
Mizutani, K., Ikeda, K., Kawai, Y. and Yamori, Y. (1999) Extract of wine phenolics improves aortic biomechanical properties in stroke-prone spontaneously hypertensive rats (SHRSP). Journal of Nutritional Science and Vitaminology, 45, 95–106.
Moiseeva, E.P., Manson, M.M. (2009) Dietary chemopreventive phytochemicals: too little or too much? Cancer Prevention Research, 2, 611–616.
Mokni, M., Elkahoui, S., Limam, F., Amri, M. and Aouani, E. (2007) Effect of resveratrol on antioxidant enzyme activities in the brain of healthy rat. Neurochemical Research, 32, 981–987.
Montopoli, M., Ragazzi, E., Froldi, G. and Caparrotta, L. (2009) Cell-cycle inhibition and apoptosis induced by curcumin and cisplatin or oxaliplatin in human ovarian carcinoma cells. Cell Proliferation, 42, 195–206.
Mori, A., Lehmann, S., O’Kelly, J., Kumagai, T., Desmond, J.C., Pervan, M., McBride, W.H., Kizaki, M. and Koeffler, H.P. (2006) Capsaicin, a component of red peppers, inhibits the growth of androgen-independent, p53 mutant prostate cancer cells. Cancer Research, 66, 3222–3229.
Moskaug, J.Ø., Carlsen, H., Myhrstad, M.C. and Blomhoff, R. (2005) Polyphenols and glutathione synthesis regulation. American Journal of Clinical Nutrition, 81, 277S–283S.
Myhrstad, M.C., Carlsen, H., Nordstrom, O., Blomhoff, R. and Moskaug, J.Ø. Flavonoids increase the intracellular glutathione level by transactivation of the gamma-glutamylcysteine synthetase catalytical subunit promoter. Free Radical Biology and Medicine, 32, 386–393.
Nabekura, T., Yamaki, T., Hiroi, T., Ueno, K. and Kitagawa, S. (2010) Inhibition of anticancer drug efflux transporter P-glycoprotein by rosemary phytochemicals. Pharmacology Research, 61, 259–263.
Naczk, M. and Shahidi, F. (2006) Phenolics in cereals, fruits and vegetables: occurrence, extraction and analysis. Journal of Pharmaceutical Research and Biomedical Analysis, 41, 1523–1542.
Najeeb-Ur-Rehman, Mehmood, M.H., Alkharfy, K.M. and Gilani, A.U. (2011) Prokinetic and laxative activities of Lepidium sativum seed extract with species and tissue selective gut stimulatory actions. Journal of Ethnopharmacology, doi:10.1016/j.jep.2011.01.047.
Nakamura, Y., Yogosawa, S., Izutani, Y., Watanabe, H., Otsuji, E. and Sakai, T. (2009) Molecular Cancer, 8, 100.
Ning, X-H, Ding, X., Childs, K.F., Bolling, S.F. and Gallagher, K.P. (1993) Flavone improves functional recovery after ischemia in isolated reperfused rabbit hearts. Journal of Thoracic and Cardiovascular Surgery, 105, 541–549.
Nishino, H. (1990) Phytochemicals in hepatocellular cancer prevention. Nutrition and Cancer, 61, 789–791.
Oakenfull, D. and Sidhu, G.S. (1990) Could saponins be a useful treatment for hypercholesterolaemia? European Journal of Clinical Nutrition, 44, 79–88.
O’Keefe, J.H. Jr and Cordain, L. (2004) Cardiovascular disease resulting from a diet and lifestyle at odds with our Paleolithic genome: how to become a 21st-century hunter-gatherer. Mayo Clinic Proceedings, 79, 101–108.
Ortiz, D. and Shea, T.B. (2004) Apple juice prevents oxidative stress induced by amyloid-beta in culture. Journal of Alzheimer’s Disease, 6, 27–30.
Pace-Asciak, C.R., Hahn, S., Diamandis, E.P., Soleas, G. and Goldberg, D.M. (1995) The red wine phenolics trans-resveratrol and quercetin block human platelet aggregation and eicosanoid synthesis: implications for protection against coronary heart disease. Clinica Chimica Acta, 31, 207–219.
Pan, M.H., Chang, Y.H., Badmaev, V., Nagabhushanam, K. and Ho, C.T. (2007) Pterostilbene induces apoptosis and cell cycle arrest in human gastric carcinoma cells. Journal of Agricultural and Food Chemistry, 55, 7777–7785.
Park, E.J. and Pezzuto, J.M. (2002) Botanicals in cancer chemoprevention. Cancer and Metastasis Reviews, 21, 231–255.
Patel, K.R., Brown, V.A., Jones, D.J., Britton, R.G., Hemingway, D., Miller, A.S., West, K.P., Booth, T.D., Perloff, M., Crowell, J.A., Brenner, D.E., Steward, W.P., Gescher, A.J. and Brown, K. (2010) Clinical pharmacology of resveratrol and its metabolites in colorectal cancer patients. Cancer Research, 70, 7392–7399.
Paul, S., DeCastro, A.J., Lee, H.J., Smolarek, A.K., So, J.Y., Simi, B., Wang, C.X., Zhou, R., Rimando, A.M. and Suh, N. (2010) Carcinogenesis, 31, 1272–1278.
Pellegrini, N., Serafini, M., Salvatore, S., Del Rio, D., Bianchi, M. and Brighenti, F. (2006)Total antioxidant capacity of spices, dried fruits, nuts, pulses, cereals and sweets consumed in Italy assessed by three different in vitro assays. Molecular Nutrition and Food Research, 50, 1030–1038.
Purkayastha, S., Berliner, A., Fernando, S.S., Ranasinghe, B., Ray, I., Tarig, H. and Banarjee, P. (2009) Curcumin blocks brain tumor formation. Brain Research, 1266, 130–8.
Ramakrishnan, G., Lo Muzio, L., Elinos-Báez, C.M., Jagan, S., Augustine, T.A., Kamaraj, S., Anandakumar, P. and Devaki, T. (2009) Silymarin inhibited proliferation and induced apoptosis in hepatic cancer cells. Cell Proliferation, 42, 229–240.
Rangkadilok, N., Worasuttayangkurn, L., Bennett, R.N. and Satayavivad, J. (2005) Identification and quantification of polyphenolic compounds in Longan (Euphoria longana Lam.) fruit. Journal of Agricultural and Food Chemistry, 53, 1387–92.
Rao, B.N. (2003) Bioactive phytochemicals in Indian foods and their potential in health promotion and disease prevention. Asia Pacific Journal of Clinical Nutrition, 12, 9–22.
Reiter, E., Azzi, A. and Zingg, J.M. (2007) Enhanced anti-proliferative effects of combinatorial treatment of delta-tocopherol and resveratrol in human HMC-1 cells. Biofactors, 30, 67–77.
Riccioni, G., Mancini, B., Di Ilio, E., Bucciarelli, T. and D’Orazio, N. (2008) Protective effect of lycopene in cardiovascular disease. European Review for Medical and Pharmacolical Sciences, 12,183–90.
Rice-Evans, (2001) C. Flavonoid antioxidants. Current Medicinal Chemistry, 8, 797–807.
Ridker, P.M., Hennekens, C.H., Roitman-Johnson, B., Stampfer, M.J. and Allen, J. (1998) Plasma concentration of soluble intercellular adhesion molecule 1 and risks of future myocardial infarction in apparently healthy men. Lancet, 351, 88–92.
Rosen, C.A. and Bryson, P.C. (2004) Indole-3-carbinol for recurrent respiratory papillomatosis: long-term results. Journal of Voice, 18, 248–253.
Ross, J.A., Potter, J.D. and Robison, L.L. (1994) Infant leukemia, topoisomerase II inhibitors, and the MLL gene. Journal of the National Cancer Institute, 86, 1678–1680.
Salas-Salvadó, J., Bulló, M., Pérez-Heras, A. and Ros, E. (2006) Dietary fibre, nuts and cardiovascular diseases. British Journal of Nutrition, 96, S46–S51.
Sarkar, F.H. and Li, Y. Using chemopreventive agents to enhance the efficacy of cancer therapy. Cancer Research, 66, 3347–3350.
Sato, M., Maulik, G., Ray, P.S., Bagchi, D. and Das, D.K. (2000) Cardioprotective effect of grape seed proanthocyanidin against ischemic reperfusion injury. Journal of Mollecular and Cellular Cardiology, 31, 1289–1297.
Scalbert, A. and Williamson, G. (2000) Dietary intake and bioavailability of polyphenols. J Nutr 130:2073S–85S.
Seeram NP. Berry fruits: compositional elements, biochemical activities, and the impact of their intake on human health, performance, and disease. J Agric Food Chem 56:627–9 (2008).
Seeram NP. Recent trends and advances in berry health benefits research. Journal of Agricultural and Food Chemistry, 58:3869–70 (2010).
Seifried HE, Anderson DE, Fisher EI, Milner JA. A review of the interaction among dietary antioxidants and reactive oxygen species. Journal of Nutritional Biochemistry, 18:567–79 (2007).
Seifried, H.E., Anderson, D.E., Milner, J.A. and Greenwald, P. (2006) Reactive oxygen species and dietary antioxidants: double-edged swords?. In: H. Panglossi (ed.) New Developments in Antioxidant Research, Hauppauge (NY), Nova Science Publishers Inc, pp. 1–25.
Sharma, R.A., Euden, S.A. and Platton, S. (2004) Phase I clinical trial of oral curcumin: biomarkers of systemic activity and compliance. Clinical Cancer Research, 10, 6847–5684.
Shin, H.K., Kim, J., Lee, E.J. and Kim, S.H. Inhibitory effect of curcumin on motility of human oral squamous carcinoma YD-10B cells via suppression of ERK and NF-kappaB activations. Phytotherapy Research, 24:577–82 (2010).
Siddiqui, I.A., Adhami, V.M., Bharali, D.J., Hafeez, B.B., Asim, M., Khwaja, S.I., Ahmad, N., Cui, H., Mousa, S.A. and Mukhtar, H. (2009) Introducing nanochemoprevention as a novel approach for cancer control: proof of principle with green tea polyphenol epigallocatechin-3-gallate. Cancer Research, 69, 1712–1716.
Singh, D., Singh, B. and Goel, R.K. (2011) Traditional uses, phytochemistry and pharmacology of Ficus religiosa: A review. Journal of Ethnopharmacology, doi:10.1016/j.jep.2011.01.046.
Skibola, C.F. and Smith, M.T. (2000) Potential health impacts of excessive flavonoid intake. Free Radicical Biology and Medicine, 29, 375–383.
Slavin, J. (2004) Whole grains and human health. Nutritional Research Review, 17, 99–110.
Slavin, J. (2003) Why whole grains are protective: biological mechanisms. Proceedings of the Nutrition Society, 62, 129–134.
Sloan, A.E. (2002) The top 10 functional food trends: The next generation. Food Technology, 56, 32–57.
Somasundaram, S., Edmund, N.A., Moore, D.T., Small, G.W., Shi, Y.Y. and Orlowski, R.Z. (2002) Dietary curcumin inhibits chemotherapy-induced apoptosis in models of human breast cancer. Cancer Research, 62, 3868–3875.
Squires, M.S., Hudson, E.A., Howells, L., Sale, S., Houghton, C.E., Jones, J.L., Fox, L.H., Dickens, M., Prigent, S.A. and Manson, M.M. Relevance of mitogen activated protein kinase (MAPK) and phosphotidylinositol-3-kinase/protein kinase B (PI3K/PKB) pathways to induction of apoptosis by curcumin in breast cells. Biochemical Pharmacology, 65, 361–376.
Staker, B.L., Hjerrild, K., Feese, M.D., Behnke, C.A., Burgin, Jr, A.B. and Stewart, L. (2002) The mechanism of topoisomerase I poisoning by a camptothecin analog. Proceedings of the National Academy of Science USA, 99, 5387–5392.
Stoclet, J.C., Andriambeloson, E. and Andriantsitohaina, R. (2000) Endothelial NO release caused by red wine polyphenols with specific structure. Experimental and Clinical Cardiology, 5, 24–27.
Stoner, G.D., Kresty, L.A., Carlton, P.S., Siglin, J.C. and Morse, M.A. (1999) Isothiocyanates and freeze-dried strawberries as inhibitors of esophageal cancer. Toxicological Sciences, 52, 95–100.
Strick, R., Strissel, P.L., Borgers, S., Smith, S.L. and Rowley, J.D. (2000) Dietary bioflavonoids induce cleavage in the MLL gene and may contribute to infant leukemia. Proceedings of the National Academy of Science USA, 97, 4790–4795.
Su, C.C., Yang, J.S., Lin, S.Y., Lu, H.F., Lin, S.S., Chang, Y.H., Huang, W.W., Li, Y.C., Chang, S.J. and Chung, J.G. (2008) Curcumin inhibits WEHI-3 leukemia cells in BALB/c mice in vivo. In Vivo 22, 63–68.
Sumiyoshi, H. (1997) New pharmacological activities of garlic and its constituents. Nippon Yakurigaku Zasshi, 110, 93P–97P.
Teskac, K. and Kristl, J. The evidence for solid lipid nanoparticles mediated cell uptake of resveratrol. International Journal of Pharmacology, 390, 61–69.
Thomasset, S.C., Berry, D.B., Garcea, G., Marczylo, T., Steward, W.P. and Gescher, A.J. Dietary polyphenolic phytochemicals--promising cancer chemopreventive agents in humans? A review of their clinical properties. International Journal of Cancer, 120, 451–458.
Thompson, L.U. (1994) Antioxidant and hormone-mediated health benefits of whole grains. Critical Reviews in Food Science and Nutrition, 34, 473–497.
Tian, L., Shi, M.M. and Forman, H.J. (1997) Increased transcription of the regulatory subunit of gamma-glutamylcysteine synthetase in rat lung epithelial L2 cells exposed to oxidative stress or glutathione depletion. Archives of Biochemistry and Biophysics, 342:126–133.
Tirapelli, C.R., Ambrosio, S.R., da Costa, F.B. and de Oliveira, A.M. (2008) Diterpenes: a therapeutic promise for cardiovascular diseases. Recent Patents on Cardiovascular Drug Discovery, 3, 1–8.
Tsao, A.S., Kim, E.S. and Hong, W.K. (2004) Chemoprevention of cancer. Cancer Journal for Clinicians, 54, 150–180.
Vitrac, X., Bornet, A., Vanderlinde, R., Valls, J., Richard, T., Delaunay, J.C., Mérillon, J.M. and Teissédre, P.L. (2005) Determination of stilbenes (delta-viniferin, trans-astringin, trans-piceid, cis- and trans-resveratrol, epsilon-viniferin) in Brazilian wines. Journal of Agricultural and Food Chemistry, 53, 5664–5669.
von Metzler, I., Krebbel, H., Kuckelkorn, U., Heider, U., Jakob, C., Kaiser, M., Fleissner, C., Terpos, E. and Sezer, O. (2009) Curcumin diminishes human osteoclastogenesis by inhibition of the signalosome-associated I kappaB kinase. Journal of Cancer Research and Clinical Oncology, 135, 173–179.
Vuddanda, P.R., Chakraborty, S. and Singh, S. (2010) Berberine: a potential phytochemical with multispectrum therapeutic activities. Expert Opinion on Investigational Drugs, 19, 1297–1307.
Wang, Y.P., Wu, J.X., Zhang, F.L. and Wang, X.R. (1993) Effect of soyasaponin and ginsenoside (stem-leave saponin) on SOD and LPO in diabetic rats. Journal of Baiqiuen Medical University, 19, 122–123.
Watanabe, S., Zhuo, X.G. and Kimira, M. (2004) Food safety and epidemiology: new database of functional food factors. Biofactors, 22, 213–219.
Wollny, T., Aiello, L., Di Tomasso, D. et al. (1999) Modulation of haemostatic function and prevention of experimental thrombosis by red wine in rats: a role for increased nitric oxide production. British Journal of Pharmacology, 127, 747–755.
Xiao, J.X., Peng, G.H. and Zhang, S.H. (2005) Prevention effects of soyasaponins on hyperlipidemia mice and its molecular mechanism. Acta Nutrimenta Sinica, 27, 147–150.
Xu, Y., Khaoustov, V.I., Wang, H., Yu, J., Tabassam, F. and Yoffe, B. (2009) Freeze-dried grape powder attenuates mitochondria- and oxidative stress-mediated apoptosis in liver cells. Journal of Agricultural and Food Chemistry, 57, 9324–9331.
Yang, C.S., Wang, X., Lu, G. and Picinich, S.C. Cancer prevention by tea: animal studies, molecular mechanisms and human relevance. Nature Reviews Cancer, 9, 429–439.
Yang, G., Zhong, L., Jiang, L., Geng, C., Cao, J., Sun, X. and Ma, Y. (2010) 6-Gingerol caused DNA strand breaks in HepG2 cells. These genotoxic effects are probably associated with oxidative stress. Genotoxic effect of 6-gingerol on human hepatoma G2 cells. Chemico-biological Interactions, 185, 12–17.
Yang, J. and Liu, R.H. (2009) Synergistic effect of apple extracts and quercetin 3-beta-d-glucoside combination on antiproliferative activity in MCF-7 human breast cancer cells in vitro. Journal of Agricultural and Food Chemistry, 57, 8581–8586.
Yankah, V.V. and Jones, P.J.H. (2001) Phytosterols and health implications-efficacy and nutritional aspects. Inform, 12, 899–903.
Zamin, L.L., Filippi-Chiela, E.C., Dillenburg-Pilla, P., Horn, F., Salbego, C. and Lenz, G. Resveratrol and quercetin cooperate to induce senescence-like growth arrest in C6 rat glioma cells. Cancer Science, 100, 1655–1662.
Zenebe, W., Pechánová, O. and Bernátová, I. (2001)_Protective effects of red wine polyphenolic compounds on the cardiovascular system. Experimental and Clinical Cardiology, 6, 153–158.
Zenebe, W. and Pechanova, O. (2002) Effects of red wine polyphenolic compounds on the cardiovascular system. Bratislavske Lekarske Listy, 103, 159–165.
Zenk, M.H. and Juenger, M. Evolution and current status of the phytochemistry of nitrogenous compounds. Phytochemistry, 68, 2757–2772.
Zern, T.L., Wood, R.J., Greene, C., West, K.L., Liu, Y., Aggarwal, D., Shachter, N.S. and Fernandez, M.L. (2005) Grape polyphenols exert a cardioprotective effect in pre- and postmenopausal women by lowering plasma lipids and reducing oxidative stress. Journal of Nutrition, 135, 1911–1917.
Zhu, B.H., Zhan, W.H., Li, Z.R., Wang, Z., He, Y.L., Peng, J.S., Cai, S.R., Ma, J.P. and Zhang, C.H. (2007) Epigallocatechin-3-gallate inhibits growth of gastric cancer by reducing VEGF production and angiogenesis. World Journal of Gastroenterology, 13, 1162–1169.