 CHAPTER 4
CHAPTER 4 
THE NATURE OF MUSCLE
CHAPTER 4 The human body contains three primary types of muscle tissue known as striated, smooth, and cardiac. These types are distinguished based on their cellular structure and anatomical locations. The organization of cells within the different types of muscle fibres influences how a muscle functions. For example, cellular organization determines the speed of muscle contraction, with those cells in skeletal muscle tissue being organized in a way that facilitates rapid contraction and those cells in smooth muscle tissue being designed in a way that limits contraction speed but enables involuntary contraction. Factors such as nutrient supply and utilization by individual muscle cells also play a major role in influencing muscle function.

Cross section of skeletal muscle highlighting the component elements of one muscle fibre. Nucleus Medical Art, Inc./Getty Images
Striated, or striped, muscle constitutes a large fraction of the total body weight in humans. Striated muscle contracts to move limbs and maintain posture. Both ends of most striated muscles articulate the skeleton and thus are often called skeletal muscles. They are attached to the bones by tendons, which have some elasticity provided by the proteins collagen and elastin, the major chemical components of tendons.
Each striated muscle has blood vessels and nerves associated with it. The vessels transport blood to and from the muscle, supplying oxygen and nutrients and removing carbon dioxide and other wastes. The signals that initiate contraction are sent from the central nervous system to the muscle via the motor nerves. Muscles also respond to hormones produced by various endocrine glands; hormones interact with complementary receptors on the surfaces of cells to initiate specific reactions. Each muscle also has important sensory structures called stretch receptors, which monitor the state of the muscle and return the information to the central nervous system. Stretch receptors are sensitive to the velocity of the movement of the muscle and the change in length of the muscle. They complete a feedback system that allows the central nervous system to assess muscular movement and to adjust motor signals in light of the movement.
Muscle is composed of many long cylindrical-shaped fibres from 0.02 to 0.08 mm in diameter. In some muscles the fibres run the entire length of the muscle (parallel fibres), up to several tens of centimetres long. In others a tendon extends along each edge, and the fibres run diagonally across the muscle between the tendons (pennate fibres). Considerable variation can be found among the different skeletal muscles, the actual arrangement of the fibres depending on the function of the muscle.
There is a high degree of organization within the fibre, a series of alternately dark and light bands. Each band extends perpendicular to the length of the fibre. Each fibre is surrounded by a complex multilayered structure called the sarcolemma. The outermost layer is a fine network of fibrils, which, at the ends of the muscle, extend into the tendons and form the structural link with them. The next layer of the sarcolemma is a foundation, or basement, membrane. The innermost layer is a plasma membrane similar to the ones that surround most cells. The plasma membrane consists of a lipid bilayer with proteins embedded in it. Some of the proteins are embedded entirely within the lipid layer, others extend to one or the other surface, and still others span the whole width of the two layers. These proteins represent enzymes, receptors, and various channels (such as those involved in the movement of ions between the exterior and interior of the cell). The plasma membrane maintains the electrical potential, which plays a major role in stimulating muscle contraction.
Sarcoplasm is the cytoplasm of a muscle fibre. It is a water solution containing ATP and phosphagens, as well as the enzymes and intermediate and product molecules involved in many metabolic reactions. The most abundant metal in the sarcoplasm is potassium. Sodium and magnesium are present in lower concentrations. Most of the calcium of muscle is bound to proteins or stored in the sarcoplasmic reticulum. Contraction is initiated by the release of calcium ions (Ca2+) upon the depolarization of the membrane, which is induced by nerve impulses.
Each striated muscle cell, or fibre, contains many nuclei. This is the result of the fusion of singly nucleated cells that occurs during the embryological development of striated muscle. After fusion, the cells never again divide.
Mitochondria in the sarcoplasm of the muscle fibre contain the enzymes involved in the Krebs cycle and in oxidative phosphorylation, processes by which living cells break down organic fuel molecules to harvest the energy they need to grow and divide. Granules in the sarcoplasm of muscle cells contain glycogen, the storage form of carbohydrate. The breakdown of glycogen and the metabolism of the individual units of the resulting carbohydrate through glycolysis, the Krebs cycle, and oxidative phosphorylation are important sources of adenosine triphosphate (ATP), the immediate source of energy for muscle contraction.
Muscles that contain many fibres that operate at a steady, low level of activity are red, due to the presence of cytochromes (molecules involved in oxidative phosphorylation) and myoglobin (an oxygen-carrying molecule in the sarcoplasm). Muscles that work in bursts of activity contain fibres that have fewer mitochondria and fewer molecules of cytochromes or myoglobin, are white, and depend more heavily on reactions that do not require oxygen to make ATP.
Electron micrographs of thin sections of muscle fibres reveal groups of filaments oriented with their axes parallel to the length of the fibre. There are two sizes of filaments, thick and thin. Each array of filaments, called a myofibril, is shaped like a cylindrical column. Along the length of each myofibril alternate sets of thick and thin filaments overlap, or interdigitate, presenting alternate bands of dark regions (with thick filaments and overlapping thin ones) and light regions (with only thin filaments). Within a fibre all the myofibrils are in register, so that the regions of similar density lie next to each other, giving the fibre the characteristic striated appearance it shows in the phase-contrast or polarized light microscope. Each light region is divided in two by a dark band. The unit between two dark bands is known as a sarcomere.
Each myofibril is about 1 or 2 micrometres (1 micrometre = 10-6 metre) in diameter and extends the entire length of the muscle fibre. The number of myofibrils per fibre varies. At the end of the fibre, the myofibrils are attached to the plasma membrane by the intervention of specialized proteins.
Forty to 80 nanometres (nm) usually separate adjacent myofibrils in a fibre. This space contains two distinct systems of membranes involved in the activation of muscle contraction. One system is a series of channels that open through the sarcolemma to the extra-fibre space. These channels are called the transverse tubules (T tubules) because they run across the fibre. The transverse tubular system is a network of interconnecting rings, each of which surrounds a myofibril. It provides an important communication pathway between the outside of the fibre and the myofibrils, some of which are located deep inside the fibre. The exact spatial relationship of the tubules to the filaments in the myofibril depends on the species of animal.
The other membrane system that surrounds each myofibril is the sarcoplasmic reticulum, a series of closed saclike membranes. Each segment of the sarcoplasmic reticulum forms a cufflike structure surrounding a myofibril. The portion in contact with the transverse tubule forms an enlarged sac called the terminal cisterna.
Each transverse tubule has two cisternae closely associated with it, forming a three-element complex called a triad. The sarcoplasmic reticulum controls the level of calcium ions in the sarcoplasm. The terminal cisternae apparently are the sites from which the calcium ions are released when the muscle is stimulated, and the longitudinal tubules are the sites at which calcium ions are effectively removed from the sarcoplasm. The removal of calcium ions (Ca2+) from the sarcoplasm is accomplished by a protein that catalyzes the breakdown of ATP, making the free energy of hydrolysis available for the energy-requiring process of Ca2+ transport.
In a longitudinal section through a group of myofibrils, there is a light band of low density called the I band. In the centre of the I band there is a prominent dense line called the Z line, although in reality, considering the three-dimensional structure of the myofibril, it is more appropriate to speak of Z disks. The area between two Z lines, a sarcomere, can be considered the primary structural and functional unit directly responsible for muscle contraction. The myofibril can thus be thought of as a stack of sarcomeres. The A band, which contains thick filaments partly overlapped with thin filaments, appears dark.
At high magnification, small bridgelike structures can be seen on the thick filaments extending toward the thin filaments in the overlap region. They are called cross bridges and are believed to be responsible for the movement and force developed during contraction. In the middle of the A band, where only thick filaments are present, is a region called the H zone. The H zone looks somewhat lighter than the overlap region of the A band. Also in the A band is a narrow, lightly stained region that contains bare thick filaments without cross bridges and is called the pseudo-H zone. In the centre of the A band is a narrow, darkly stained region called the M band, in which occur fine bridges between the thick filaments. These bridges differ from the cross bridges between the thick and thin filaments and are in fact composed of an entirely different protein.
If cross sections of the myofibril at different levels of the sarcomere are examined by electron microscope, the filaments can be seen end-on, and the three-dimensional nature of the lattice of filaments can be appreciated. The I band contains only thin filaments, with a diameter of 6 to 8 nm. In the A band, in the overlap region, the thin filaments appear with thick ones (diameter of 12 nm) in an extremely regular pattern or lattice. In vertebrates the thick filaments are arranged in a hexagonal lattice, and the thin ones are located at the centre of the equilateral triangles formed by the thick filaments. Sections through the H zone contain only thick filaments arranged in the same hexagonal pattern they form in the overlap region. In the M band the hexagonal array of thick filaments can be seen with M bridges running between them.
The discovery that during contraction the filaments do not shorten but that the two sets—thick and thin—merely move relative to each other is crucial for understanding muscle physiology. During contraction the thin filaments move deeper into the A band, and the overlap of the thick and thin filaments increases. If a longitudinal section of the sarcomere is considered, the thin filaments on the left side of the A band would move to the right into the A band, and the filaments on the right of the A band would move to the left into the A band. Directionality of the motion partly results from the structural polarity of both the thick filaments, since in the two halves of the filament the myosin molecules are oriented in opposite directions, and the actin filaments, in which the actin molecules are oriented with respect to the Z bands.
To understand the finer structural details of the myofilaments and the mechanism by which sliding, and ultimately muscle contraction, occurs, one must understand the molecular components of the filaments and of the structures associated with them. The myofilaments are composed of several different proteins, constituting about 50 percent of the total protein in muscle. The other 50 percent consists of the proteins in the Z line and M band, the enzymes in the sarcoplasm and mitochondria, collagen, and the proteins in membrane structures. Of the myofilament proteins, myosin and actin are known to play a direct part in the contractile event. Troponin and tropomyosin, which are located in the thin filaments together with calcium ions, regulate contraction by controlling the interaction of myosin and actin.
The main constituent of the thick filaments is myosin. Each thick filament is composed of about 250 molecules of myosin. Myosin has two important roles: a structural one, as the building block for the thick filaments, and a functional one, as the catalyst of the breakdown of ATP during contraction and in its interaction with actin as part of the force generator of muscle. The individual myosin molecule contains two major protein chains and four small ones, the entire molecule being about 160 nm in length and asymmetrically shaped. The rodlike tail region, about 120 nm long, consists of two chains of protein, each wound into what is known as an α-helix, together forming a coiled-coil structure. At the other end of the molecule, the two protein chains form two globular headlike regions that have the ability to combine with the protein actin and carry the enzymatic sites for ATP hydrolysis.
In the middle portion of the thick filament, the myosin molecules are assembled in a tail-to-tail fashion. Along the rest of the filament, they are arranged head to tail. The tail parts of the molecules form the core of the filament, while the head portions project out from the filament. The cross bridges are actually the globular head regions of myosin molecules extending outward from the filament, and the smooth pseudo-H zone is the region of tail-to-tail aggregation, in which there are only tails and no heads.
The precise three-dimensional arrangement of the cross bridges projecting from the thick filament cannot be seen easily in electron micrographs but can be determined from X-ray diffraction study of living muscle. The three bridges project 120 degrees from the opposite sides of the filament every 14.3 nm along the length of the filament. Each successive set of bridges is located in a position rotated 40 degrees farther around the filament. The pattern of nine bridges (three sets of three bridges) repeats itself every 42.9 nm along the thick filament. Some variation may exist from species to species and muscle to muscle.
The thin filaments contain three different proteins—actin, tropomyosin, and troponin. The latter is actually a complex of three proteins.
Actin, which constitutes about 25 percent of the protein of myofilaments, is the major component of the thin filaments in muscle. An individual molecule of actin is a single protein chain coiled to form a roughly egg-shaped unit. Actin in this form, called globular actin or G-actin, has one calcium or magnesium ion and one molecule of ATP bound to it. Under the proper conditions, G-actin is transformed into the fibrous form, or F-actin, that exists in the thin filament in muscle. When the G-to-F transformation takes place, the ATP bound to G-actin breaks down, releasing inorganic phosphate (PO43-) and leaving an adenosine diphosphate (ADP) molecule bound to each actin unit. Actin molecules repeat every 2.75 nm along the thin filament. They give rise to a helical structure that can be viewed as a double or single helix. The apparent half-pitch is about 40 nm long. Actin is believed to be directly involved in the process of contraction because the cross bridges can become attached to it.
Tropomyosin is a rod-shaped molecule about 40 nm long. Two strands of tropomyosin molecules run diametrically opposed along the actin filaments. Tropomyosin has a structure similar to that of the myosin tail, being a coiled unit of two protein chains. Each tropomyosin molecule is in contact with seven actin units.
Troponin is a complex of three different protein subunits. One troponin complex is bound to every tropomyosin molecule. A troponin molecule is located approximately every 40 nm along the filament. Troponin and tropomyosin are both involved in the regulation of the contraction and relaxation of muscles. One of the subunits (TnC) is the receptor for Ca2+ released from the sarcoplasmic reticulum on activation of the muscle. It is thought that calcium binding then causes further structural changes in the interaction of actin, tropomyosin, and another troponin subunit (TnI) that lead to contraction by activating the actin-myosin interaction.
Mixtures of myosin and actin in test tubes are used to study the relationship between the ATP breakdown reaction and the interaction of myosin and actin. The ATPase reaction can be followed by measuring the change in the amount of phosphate present in the solution. The myosin-actin interaction also changes the physical properties of the mixture. If the concentration of ions in the solution is low, myosin molecules aggregate into filaments. As myosin and actin interact in the presence of ATP, they form a tight compact gel mass; the process is called superprecipitation. Actin-myosin interaction can also be studied in muscle fibres whose membrane is destroyed by glycerol treatment; these fibres still develop tension when ATP is added. A form of ATP that is inactive unless irradiated with a laser beam is useful in the study of the precise time course underlying contraction.
If troponin and tropomyosin are also present, however, the actin and myosin do not interact, and ATP is not broken down. This inhibitory effect corresponds to the state of relaxation in the intact muscle. When calcium ions are added, they combine with troponin, inhibition is released, actin and myosin interact, and ATP is broken down. This corresponds to the state of contraction in intact muscle. The exact mechanism by which troponin, tropomyosin, and calcium ions regulate the myosin-actin interaction is not fully agreed upon. In the thin filament there are one troponin and one tropomyosin molecule for every seven actin units. According to one view, Ca2+ binding to troponin (actually the TnC subunit) induces a change in the position of tropomyosin, moving it away from the site where myosin also binds (steric blocking). Alternatively, the calcium-induced movement of tropomyosin in turn induces changes in the structure of actin, permitting its interaction with myosin (allosteric model). In smooth muscles, Ca2+ activates an enzyme (kinase) that catalyzes the transfer of phosphate from ATP to myosin, and the phosphorylated form is then activated by actin.
In skeletal muscle, most ATP is produced in metabolic pathways involving reactions of the sugar glucose or some other carbohydrate derived from glucose. During contraction, for example, glucose is made available for these reactions by the breakdown of glycogen, the storage form of carbohydrate in animal cells. The concentration of Ca2+ is transiently increased on activation of muscle. The ions are also activators of the process of glycogen breakdown. During the recovery period, the glycogen supply is replenished by synthesizing glycogen from glucose supplied to the muscle tissue by the blood.
In a resting muscle, the available supply of ATP can sustain maximal muscle work for less than one second. The muscle, therefore, must continuously replenish its ATP store, and this is done in many different ways. One mechanism for the formation of ATP operates so rapidly that for a long time scientists were unable to detect any change in the amount of ATP in the muscle as a result of contraction. This immediate rebuilding of ATP is accomplished by the reactions of compounds called phosphagens. All of these compounds contain phosphorus in a chemical unit called a phosphoryl group, which they transfer to ADP (adenosine diphosphate) to produce ATP (these compounds are also referred to as high-energy phosphates).
During rapid and intense contraction, phosphagen can be utilized to rebuild ATP rapidly and maintain its level as long as the phosphagen lasts, which in a maximally working human muscle is just a few seconds. After contraction, ATP is utilized to form phosphagen from creatine. ADP is also formed.
The amount of phosphagen is higher in skeletal muscle than it is in cardiac or smooth muscle. This correlates with the type of activity of the muscles. Skeletal muscle operates in bursts of activity, whereas cardiac and smooth muscle contract in a regular pattern. Skeletal muscle needs an immediate supply of a large amount of ATP, which is provided by the phosphagen reaction; cardiac and smooth muscle, which use ATP at a lower rate, rely on slower reactions to fill their energy requirements.
The nerve impulse that ultimately results in muscle contraction appears as an action potential at the sarcolemma, the membrane that surrounds the muscle fibre. This electrical signal is communicated to the myofilaments inside the fibre in the following way. When the action potential reaches the opening of the transverse tubules (channels that open through the sarcolemma to the space outside the fibre) in the surface of the fibre, it travels down into the fibre along the tubular membranes, which are continuous with the surface membrane, to within a fraction of a micrometre of each functional contractile unit. In mammalian muscles the transverse tubules are located at the edge of the A bands and I bands. At the triads (the three-element complex consisting of one transverse tubule and two cisternae, which are enlarged saclike membranes), the transverse tubule walls are close to the membranes of the terminal cisternae of the sarcoplasmic reticulum.
By some as-yet-unknown mechanism, the change in the electrical properties of the transverse tubules during an action potential causes the rapid release by the terminal cisternae of relatively large amounts of calcium ions into the sarcoplasm. As the concentration of calcium ions increases in the sarcoplasm, they become bound to the troponin in the thin filaments. This releases (or removes) the troponin-tropomyosin-mediated inhibition of the myosin-actin interaction. As the stimulation of the muscle continues, the terminal cisternae continue to release calcium ions. At the same time, however, some of the calcium ions are being removed from the sarcoplasm by another portion of the sarcoplasmic reticulum, the longitudinal tubules. Once the calcium ions are inside the lumen (cavity) of the longitudinal tubules, many of them slowly diffuse back to the terminal cisternae, where they are bound to a protein, calsequestrin, as a storage site. The removal of calcium ions from the sarcoplasm by the sarcoplasmic reticulum is energy-requiring. The breakdown of ATP is the chemical reaction that supplies the energy, and two calcium ions are apparently removed from the sarcoplasm for each ATP molecule that is split.
Because vertebrate smooth muscle is located in the walls of many hollow organs, the normal functioning of the cardiovascular, respiratory, gastrointestinal, and reproductive systems depends on the constrictive capabilities of smooth muscle cells. Smooth muscle is distinguished from the striated muscles of the skeleton and heart by its structure and its functional capabilities.
As the name implies, smooth muscle presents a uniform appearance that lacks the obvious striping characteristic of striated muscle. Vascular smooth muscle shortens 50 times slower than fast skeletal muscle but generates comparable force using 300 times less chemical energy in the process. These differences in the mechanical properties of smooth versus striated muscle relate to differences in the basic mechanism responsible for muscle shortening and force production. As in striated muscle, smooth muscle contraction results from the cyclic interaction of the contractile protein myosin (i.e., the myosin cross bridge) with the contractile protein actin. The arrangement of these contractile proteins and the nature of their cyclic interaction account for the unique contractile capabilities of smooth muscle.
Smooth muscle contains spindle-shaped cells 50 to 250 μm (micrometres) in length by 5 to 10 μm in diameter. These cells possess a single, central nucleus. Surrounding the nucleus and throughout most of the cytoplasm are the thick (myosin) and thin (actin) filaments. Tiny projections that originate from the myosin filament are believed to be cross bridges. The ratio of actin to myosin filaments (approximately 12 to 1) is twice that observed in striated muscle and thus may provide a greater opportunity for a cross bridge to attach and generate force in smooth muscle. This may in part account for the ability of smooth muscle to generate, with far less myosin, comparable or greater force than striated muscle.
Smooth muscle differs from striated muscle in its lack of any apparent organization of the actin and myosin contractile filaments into the discrete contractile units called sarcomeres. Research has shown that a sarcomere-like structure may nonetheless exist in smooth muscle. Such a sarcomere-like unit would be composed of the actin filaments that are anchored to dense amorphous bodies in the cytoplasm as well as dense plaques on the cell membrane. These dense areas are composed of the protein α-actinin, found in the Z lines of striated muscle, to which actin filaments are known to be attached. Thus, force generated by myosin cross bridges attached to actin is transmitted through actin filaments to dense bodies and then through neighbouring contractile units, which ultimately terminate on the cell membrane.
Relaxed smooth muscle cells possess a smooth cell membrane appearance, but upon contraction, large membrane blebs (or eruptions) form as a result of inwardly directed contractile forces that are applied at discrete points on the muscle membrane. These points are presumably the dense plaques on the cell membrane to which the actin filaments attach. As an isolated cell shortens, it does so in a corkscrewlike manner. It has been hypothesized that in order for a single cell to shorten in such a unique fashion, the contractile proteins in smooth muscle are helically oriented within the muscle cell. This helical arrangement agrees with earlier speculation that the contractile apparatus in smooth muscle may be arranged at slight angles relative to the long axis of the cell. Such an arrangement of contractile proteins could contribute to the slower shortening velocity and enhanced force-generating ability of smooth muscle.
The contractile proteins interact to generate a force that must be transmitted to the tissue in which the individual smooth muscle cells are embedded. Smooth muscle cells do not have the tendons present in striated muscles that allow for transfer of muscular force to operate the skeleton. Smooth muscles, however, are generally embedded in a dense connective tissue matrix that connects the smooth muscle cells within the tissue into a larger functional unit.
Other organelles of the cell interior are related to energy production and calcium storage. Mitochondria are located most frequently near the cell nucleus and at the periphery of the cell. As in striated muscles, these mitochondria are linked to ATP production. The sarcoplasmic reticulum is involved in the storage of intracellular calcium. As in striated muscle, this intracellular membrane system plays an important role in determining whether or not contraction occurs by regulating the concentration of intracellular calcium.
Smooth muscle cells contract in response to neuronal or hormonal stimulation, either of which results in an increase in intracellular calcium as calcium enters through membrane channels or is released from intracellular storage sites. The elevated level of calcium in the cell cytoplasm results in force generation. The rise in the level of intracellular calcium, however, initiates contraction through a mechanism that differs substantially from that in striated muscle. In striated muscle, myosin cross bridges are prevented from attaching to actin by the presence of the troponin-tropomyosin system molecules on the actin filament. In smooth muscle, although tropomyosin is present, troponin is not, which means that an entirely different regulatory scheme operates in smooth muscle. Regulation of the contractile system in smooth muscle is linked to the myosin filament; regulation in striated muscle is linked to the actin filament.
In order for the smooth muscle myosin cross bridge to interact cyclically with actin, a small protein on the myosin molecule called the light chain must be phosphorylated (receive a phosphate group). This phosphorylation is the result of a series of interdependent biochemical reactions that are initiated by the rise in intracellular calcium. For the cell to relax, the concentration of intracellular calcium falls, thus inactivating these biochemical processes associated with light chain phosphorylation. The phosphate molecule that was added in the previous steps, however, still must be removed from the light chain so that attachment of the cross bridge to actin is prevented. Phosphatases are enzymes in the muscle cell that cleave the phosphate group from the myosin light chain.
Smooth muscle contraction requires the release of chemical energy stored in ATP molecules. The release of this chemical energy by the myosin cross bridge and the resultant mechanical work is commonly referred to as the cross-bridge cycle, which in smooth muscle is believed to be a multistep process similar to that in striated muscle. Therefore, the mechanical properties of smooth muscle, as of striated muscles, are intimately linked to this multistate cross-bridge cycle. For instance, there is a correlation between the rate at which the cross bridges cycle and the maximum shortening velocity of the muscle. Since the actomyosin ATPase cross-bridge cycle in smooth muscle is considerably slower than that in striated muscle, the slower shortening velocity in smooth muscle must be partly due to the reduced turnover rate of the cross bridge. The slower cycling rate could also account for the high economy of ATP utilization that characterizes smooth muscle force production, since fewer cycles are required and less energy is consumed in the generation of force.
The relationship between smooth muscle’s ability to shorten and to generate force is characterized by the force-velocity relationship. The form of this relationship is qualitatively similar to that in striated muscle. However, the smooth muscle force-velocity relationship differs from that of striated muscle in having a slower maximum shortening velocity and a greater force per cross-sectional area of muscle. As mentioned above, the slower shortening velocity may relate to the slower cycling rate of the cross bridge as well as the orientation of the contractile proteins within the muscle cell.
The force-generating capabilities of smooth muscle are greater than those of striated muscle despite the fact that there is considerably less myosin in smooth muscle. Possible explanations for this relate to the arrangement of the contractile apparatus within the cell, which gives rise to more cross bridges effectively operating in conjunction with one another. Also, enhanced force production could be related to the greater amount of time that the cross bridge spends in the attached, high force-producing state (i.e., duty cycle). Evidence for such an increase in the duty cycle does exist in smooth muscle.
When fully contracted, the amount of force that smooth muscle can generate depends on the muscle length. Therefore, as in striated muscle, an optimal length for force production exists, with force being reduced at both lesser and greater lengths. The similarity in shape for the force-velocity and length-tension relationships between smooth and striated muscle suggests that in smooth muscle both a cross-bridge mechanism and a sliding of contractile filaments must occur.
Smooth muscle cells often must generate constant force for prolonged periods of time. In order to do this without depleting the muscle’s energy supply, smooth muscle appears to have adapted by altering the cross-bridge cycling rate during the time course of a single contraction. Thus, the modulation of cross-bridge cycling rate represents a highly economical means of generating force in a muscle that often exists in a tonic state of contraction.
The heart is the pump that keeps blood circulating throughout the body and thereby transports nutrients, breakdown products, antibodies, hormones, and gases to and from the tissues. The heart consists mostly of muscle. The myocardial cells (collectively termed the myocardium) are arranged in ways that set it apart from other types of muscle. The outstanding characteristics of the action of the heart are its contractility, which is the basis for its pumping action, and the rhythmicity of the contraction.
Heart muscle differs from its counterpart, skeletal muscle, in that it exhibits rhythmic contractions. The amount of blood pumped by the heart per minute (the cardiac output) varies to meet the metabolic needs of the peripheral tissues (muscle, kidney, brain, skin, liver, heart, and gastrointestinal tract). The cardiac output is determined by the contractile force developed by the muscle cells of the heart (myocytes), as well as by the frequency at which they are activated (rhythmicity). The factors affecting the frequency and force of heart muscle contraction are critical in determining the normal pumping performance of the heart and its response to changes in demand.
The heart is a network of highly branched cardiac cells 110 μm in length and 15 μm in width, which are connected end to end by intercalated disks. The cells are organized into layers of myocardial tissue that are wrapped around the chambers of the heart. The contraction of the individual heart cells produces force and shortening in these bands of muscle, with a resultant decrease in the heart chamber size and the consequent ejection of the blood into the pulmonary and systemic vessels. Important components of each heart cell involved in excitation and metabolic recovery processes are the plasma membrane and transverse tubules in registration with the Z lines, the longitudinal sarcoplasmic reticulum and terminal cisternae, and the mitochondria. The thick (myosin) and thin (actin, troponin, and tropomyosin) protein filaments are arranged into contractile units (that is, the sarcomere extending from Z line to Z line) that have a characteristic cross-striated pattern similar to that seen in skeletal muscle.
The human heart. Shutterstock.com
The rate at which the heart contracts and the synchronization of atrial and ventricular contraction required for the efficient pumping of blood depend on the electrical properties of the myocardial cells and on the conduction of electrical information from one region of the heart to another. The action potential (activation of the muscle) is divided into five phases (0–4). Each of the phases of the action potential is caused by time-dependent changes in the permeability of the plasma membrane to potassium ions (K+), sodium ions (Na+), and calcium ions (Ca2+). The resting potential of the myocytes of the ventricle (phase 4) begins with the outside of the cell being positive—i.e., having a greater concentration of positive ions. Atrial and ventricular myocytes are normally quiescent (nonrhythmic). However, when the resting membrane potential is depolarized to a critical potential (Ecrit), a self-generating action potential follows, leading to muscle contraction. Phase 0, the upstroke, is associated with a sudden increase in membrane permeability to Na+. Phases 1, 2, and 3 result from changes in membrane permeability and conductance to Na+, K+, and Ca2+.
The electrical activity of heart muscle cells differs substantially from that of skeletal muscle cells in that phases 1, 2, and 3 are considerably prolonged (200 milliseconds versus 5 milliseconds, respectively). Another significant difference in excitability is that heart muscle cannot be tetanized (i.e., induced to spasm) by the application of repetitive stimuli, thus ensuring the completion of the contraction/relaxation cycle and the effective pumping of blood.
Because atrial and ventricular cells are normally quiescent, exhibiting action potentials only after the muscle is depolarized to the critical membrane potential (Ecrit), the source of the rhythmic contractions of the heart must be sought elsewhere. In contrast to atrial and ventricular myocytes, the myocytes of the sinoatrial (SA) node, the atrioventricular (AV) node, the bundle branches, and the Purkinje fibre system are made up of specialized cardiac muscle cells that exhibit a spontaneous upward drift in the resting potential toward Ecrit, resulting in the generation of the action potential with all of its phases. The normal rhythmicity of cells from each of these regions depends on the rate at which spontaneous depolarization occurs and the resting membrane potential from which it starts. The region with the fastest intrinsic rate, the SA node, sets the pace for the whole heart. The pacemaker activity is propagated to the rest of the heart by means of the low electrical resistance pathways through the muscle cells (e.g., intercalated disks) and the presence of specialized conducting tissue (e.g., bundle branches and the Purkinje system). The time course of activation and the shape of the action potentials in different parts of the heart are responsible for the synchronous activation and contraction of the muscles of the atrium followed by those of the ventricle.
The normal rhythm of the heart (i.e., the heart rate) can be altered by neural activity. The heart is innervated by sympathetic and parasympathetic nerves, which have a profound effect on the resting potential and the rate of diastolic depolarization in the SA nodal region. The activity of the sympathetic nervous system may be increased by the activation of the sympathetic nerves innervating the heart or by the secretion of epinephrine and norepinephrine from the adrenal gland. This decreases the resting potential of the myocytes of the SA node while increasing the rate of diastolic depolarization. The result is an increase in the heart rate. Conversely, stimulating the parasympathetic nervous system (vagal nerves to the heart) increases the resting potential and decreases the rate of diastolic depolarization; under these circumstances the heart rate slows. The sympathetic nervous system is activated under conditions of fright or vigorous activity (the so-called fight-or-flight reaction), where the increase in force and rate of heart contraction are easily felt, while the parasympathetic system exerts its influence during periods of rest.
Immediately following depolarization of the plasma membrane and the ensuing action potential, the heart muscle develops force and then relaxes. The surface action potential is transmitted to the interior of the muscle by means of the transverse tubular system. Calcium ions enter the muscle cell during the plateau phase of the action potential (phase 2), triggering the release of calcium from the terminal cisternae of the sarcoplasmic reticulum. Calcium diffuses to the myofilaments and combines with the troponin-tropomyosin system (associated with the thin actin filaments), producing a conformational change that allows actin and myosin to interact. This interaction in the presence of ATP results in cross-bridge cycling and ATP hydrolysis.
The force developed in the whole muscle is the sum of all the forces developed by each of the millions of cycling cross bridges of the muscle. The free calcium ions in the cytosol are removed by an energy-dependent calcium uptake system involving calcium ion pumps located in the longitudinal sarcoplasmic reticulum. These calcium pumps lower the concentration of free calcium in the cytosol, resulting in the dissociation (release) of calcium from the troponin-tropomyosin system. The troponin-tropomyosin system is then transformed back to its original state, preventing myosin and actin from interacting and thus causing relaxation of the muscle. At the same time, calcium is extruded from the cell into the surrounding medium.
There are a number of factors that change the force developed by heart muscle cells. In a manner similar to that seen in skeletal muscle, there is a relationship between the muscle length and the isometric force developed. As the muscle length is increased, the active force developed reaches a maximum and then decreases. This maximum point is the length at which the heart normally functions. As with skeletal muscle, changes in length alter the active force by varying the degree of overlap of the thick myosin and thin actin filaments.
The force developed by heart muscle also depends on the frequency at which the muscle is stimulated. As the stimulus frequency is increased, the force is increased until the maximum is reached, at which point it begins to decrease. An increase in the level of circulating epinephrine and norepinephrine from the sympathetic nervous system also increases the force of contraction. All these factors can combine to allow the heart to develop more force when required. At any given length the velocity of contraction is a function of the load lifted, with the velocity decreasing as the load is increased.
Demands on the heart vary from moment to moment and from day to day. In moving from rest to exercise, the cardiac output may be increased tenfold. Other increases in demand are seen when the heart must pump blood against a high pressure such as that seen in hypertensive heart disease. Each of these stresses requires special adjustments. Short-term increases in demand on the heart (e.g., exercise) are met by increases in the force and frequency of contraction. These changes are mediated by increases in sympathetic nervous system activity, an increase in the frequency of contraction, and changes in muscle length. The response to long-term stress (such as high blood pressure) results in an increase in the mass of the heart (hypertrophy), providing more heart muscle to pump the blood, which helps meet the increase in demand. In addition, subtle intracellular changes affect the performance of the muscle cells.
In the pressure-overload type of hypertrophy (hypertensive heart disease), the pumping system of the sarcoplasmic reticulum responsible for calcium removal is slowed while the contractile protein myosin shifts toward slower cross-bridge cycling. The outcome is a slower, more economical heart that can meet the demand for pumping against an increase in pressure. At the molecular level the slowing of calcium uptake is caused by a reduction in the number of calcium pumps in the sarcoplasmic reticulum. The change in the maximum velocity of shortening and economy of force development occur because each myosin cross-bridge head cycles more slowly and remains in the attached force-producing state for a longer period of time.
In the thyrotoxic type of hypertrophy, calcium is removed more quickly while there is a shift in myosin. At the molecular level there are more sarcoplasmic reticular calcium pumps, while the myosin cross-bridge head cycles more rapidly and remains attached in the force-producing state for a shorter period of time. The result is a heart that contracts much faster but less economically than normal and can meet the peripheral need for large volumes of blood at normal pressures.
Abductor muscles cause movement of a limb away from the midplane of the body or away from a neighbouring part or limb, as in raising the arms to the side (effected by the deltoideus muscle) or spreading the fingers or toes. In humans certain muscles of the hands and feet are named for performing this function. In the hand, the abductor digiti minimi manus acts upon the little finger, and both the abductor pollicis longus and abductor pollicis brevis act upon the thumb. The corresponding foot muscles are the abductor digiti minimi pedis and the abductor hallucis, which act on the little and great toes, respectively.
The human biceps muscle contracted (l) and extended. Shutterstock.com
Adductor muscles draw a part of the body toward its median line or toward the axis of an extremity, particularly three powerful muscles of the human thigh—adductor longus, adductor brevis, and adductor magnus. Originating at the pubis and the ischium (lower portions of the pelvis—the hip bone), these ribbonlike muscles are attached along the femur. Their primary action is adduction of the thigh, as in squeezing the thighs together. They also aid in rotation and flexion of the thigh.
Other muscles named for this function include the adductor pollicis, which draws in and opposes the thumb, and the adductor hallucis, which acts on the great toe.
A muscle that increases the angle between members of a limb, as by straightening the elbow or knee or bending the wrist or spine backward, is known as an extensor muscle. The movement is usually directed backward, with the notable exception of the knee joint.
In humans, certain muscles of the hand, and foot are named for this function. In the hand these include the extensor carpi radialis brevis, extensor carpi radialis longus, and extensor carpi ulnaris, which run from the humerus (bone of the upper arm) along the back of the forearm to the metacarpal bones at the back of the hand and which extend the wrist. Other extensor muscles in the hand include the extensor digitorum, which runs from the humerus to a common tendon attached to all of the fingers and which extends the fingers; the extensor indicis, which acts upon the index finger; and the extensor pollicis brevis and extensor pollicis longus, which run from the radius and ulna (bones of the forearm), respectively, and act upon the thumb.
Extensors in the foot include the extensor digitorum longus and extensor digitorum brevis, which originate at the upper and lower parts of the lower leg and act through long tendons upon the toes, and the extensor hallucis brevis and extensor hallucis longus, which act upon the great toe. The longus muscles of the foot also aid upward flexion of the foot at the ankle.
Flexor muscles work in an opposite manner from extensor muscles, decreasing the angle between bones on two sides of a joint, as in bending the elbow or knee. Several of the muscles of the hands and feet are named for this function. The flexor carpi radialis and flexor carpi ulnaris stretch from the humerus (upper-arm bone) along the inside of the forearm to the metacarpal bones of the hand and flex the wrist. The flexor digitorum profundus is a deep muscle that originates at the ulna (bone of the forearm) and acts to bend the fingers near their tips. The flexor digitorum superficialis is closer to the surface. It originates at two points, one at the junction of the humerus and ulna and the other along the radius (bone of the forearm), and acts upon the midsections of the fingers. Also in the hand are the flexor pollicis longus and flexor pollicis brevis, long and short flexors of the thumb, originating in the forearm and base of the hand, respectively. The flexor digiti minimi brevis manus acts upon the little finger.
In the foot are the flexor digitorum longus and flexor digitorum brevis, originating at the tibia (shin) and calcaneus (heel bone), respectively, and acting upon the four smaller toes. The flexor hallucis longus and flexor hallucis brevis originate in the calf and near the heel, respectively, and flex the great toe. The flexor digiti minimi brevis pedis acts upon the smallest toe.
The ringlike muscles that surround and contract or close a bodily passage or opening are called sphincter muscles. One of the most important human sphincter muscles is the sphincter pylori, a thickening of the middle layer of stomach muscle around the pylorus (opening into the small intestine) that holds food in the stomach until it is thoroughly mixed with gastric juices. Other sphincters are involved in excretion of waste: the sphincter ani externus keeps the anal opening closed by its normal contraction, and the sphincter urethrae is the most important voluntary control of urination. There is also a sphincter in the eye, the sphincter pupillae, a ring of fibres in the iris that contracts the pupil in the presence of bright light.

The human male urinary bladder and urethra. Encyclopædia Britannica, Inc.
Levator muscles act to raise a body part. For example, the levator anguli oris raises the corner of the mouth, and the levator ani, the collective name for a thin sheet of muscle that stretches across the pelvic cavity, helps hold the pelvic viscera in position, forming a kind of sphincter around the vagina in the female and the anal canal in both sexes. Other examples include the levatores costarum, which help raise the ribs during respiration; the levator palpebrae superioris, which raises the upper eyelid; the levator scapulae, a straplike muscle of the shoulder that helps raise and rotate the shoulder blade; and the levator veli palatini, which raises the soft palate of the mouth.
The spinalis muscles form the deep muscles of the back near the vertebral column that, as part of the erector spinae (sacrospinalis) muscle group, assist in extension (e.g., bending backward), lateral flexion (bending to the side), and rotation of the spine. The spinalis thoracis is the major spinalis muscle, arising from the bones of the lower thoracic and upper lumbar vertebral spine and inserted into the bones of the upper thoracic vertebral spine. It may be prolonged into the neck (spinalis cervicis) and head (spinalis capitis).
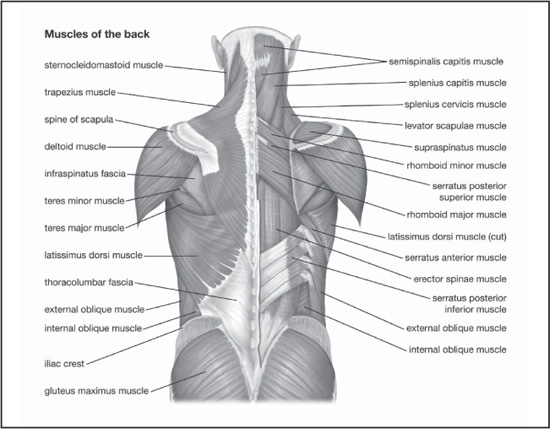
Muscles of the back. Encyclopædia Britannica, Inc.
The semispinalis muscles are deep muscles, located just to either side of the spine, that arise from the transverse processes (side projections) of the lower vertebrae and reach upward across several vertebrae to insert at the spines of vertebrae farther up, except for the upper segment (semispinalis capitis), which inserts at the occipital bone of the skull. The lower and middle segments (semispinalis dorsi and semispinalis cervicis) help extend (bend backward) and rotate the spinal column; the upper segment also helps bend the head backward.
A deep muscle of the back, the erector spinae arises from a tendon attached to the crest along the centre of the sacrum (the part of the backbone at the level of the pelvis, formed of five vertebrae fused together). When it reaches the level of the small of the back, the erector divides into three columns, each of which has three parts. The muscle system extends the length of the back and functions to straighten the back and to rotate it to one side or the other.
Iliocostalis muscles are deep muscles of the back that, as part of the erector spinae (sacrospinalis) muscle group, aid in extension (bending backward), lateral flexion (bending to the side), and rotation of the spinal column. The iliocostalis group consists of a lower part (iliocostalis lumborum) that extends from the ilium (upper part of the hip bone) to the lower ribs, a middle part (iliocostalis dorsi, or thoracis) that extends from the lower to the upper ribs, and an upper part (iliocostalis cervicis) that extends from the upper ribs to the transverse processes (side projections) of the cervical vertebrae (in the neck).
The coccygeus muscle of the lower back arises from the ischium (lower, rear portion of the hip bone) and from the ligaments that join the spinal column and the sacrum (triangular bone at the base of the spine). It is attached to the lower sacrum and the coccyx (tailbone). In humans its major function is to support the pelvic viscera, especially in the female. It also raises the coccyx.
The widest and most powerful muscle of the back is the latissimus dorsi. It is a large, flat, triangular muscle covering the lower back. It arises from the lower half of the vertebral column and iliac crest (hip bone) and tapers to a rounded tendon inserted at (attached to) the front of the upper part of the humerus (upper-arm bone).
The action of the latissimus dorsi draws the upper arm downward and backward and rotates it inward, as exemplified in the downstroke in swimming the crawl. In climbing it joins with the abdominal and pectoral muscles to pull the trunk upward. The two latissimus dorsi muscles also assist in forced respiration by raising the lower ribs.
The trapezius muscle is a large, superficial muscle at the back of the neck and the upper part of the thorax, or chest. The right and left trapezius together form a trapezium, an irregular four-sided figure. It originates at the occipital bone at the base of the skull, the ligaments on either side of the seven cervical (neck) vertebrae (ligamentum nuchae), and the seventh cervical and all thoracic vertebrae. It is inserted on the posterior of the clavicle (collarbone) and on the spine of the scapula (shoulder blade). Its chief action is support of the shoulders and limbs and rotation of the scapula necessary to raise the arms above shoulder level.
The pectoralis muscles connect the front walls of the chest with the bones of the upper arm and shoulder. There are two such muscles on each side of the sternum (breastbone) in the human body: pectoralis major and pectoralis minor.
The pectoralis major, the larger and more superficial, originates at the clavicle (collarbone), the sternum, the ribs, and a tendinous extension of the external oblique abdominal muscle. The pectoralis major extends across the upper part of the chest and is attached to a ridge at the rear of the humerus (the bone of the upper arm). Its major actions are adduction, or depression, of the arm (in opposition to the action of the deltoideus muscle) and rotation of the arm forward about the axis of the body. When the raised arms are fixed (as in mountain climbing), it assists the latissimus dorsi and teres major muscles in pulling the trunk up.
The pectoralis minor lies, for the most part, beneath the pectoralis major, arising from the middle ribs and inserting into (attaching to) the scapula (shoulder blade). It aids in drawing the shoulder forward and downward (in opposition to the trapezius muscle).
The intercostalis muscles form a series of short muscles that extend between the ribs and serve to draw them together during inspiration and forced expiration or expulsive actions. A set of external and internal intercostalis muscles is found between each vertical pair of ribs on each side of the chest.
The abdominal muscles include the muscles of the anterolateral walls of the abdominal cavity. They are composed of three flat muscular sheets, from outside in: external oblique, internal oblique, and transverse abdominis, supplemented in front on each side of the midline by rectus abdominis.
The first three muscle layers extend between the vertebral column behind, the lower ribs above, and the iliac crest and pubis of the hip bone below. Their fibres all merge toward the midline, where they surround the rectus abdominis in a sheath before they meet the fibres from the opposite side at the linea alba. Strength is developed in these rather thin walls by the crisscrossing of fibres. Thus, the fibres of the external oblique are directed downward and forward, those of the internal oblique upward and forward, and those of the transverse horizontally forward.
Around the rectus abdominis, which extends from the pubis upward to the ribs, the above muscles are all fibrous. In the region of the groin, between the pubic bone and the anterior superior iliac spine, a specialized arrangement of these fibres permits the formation of the inguinal canal, a passage through the muscular layers. This develops at birth as the testes descend out of the abdominal cavity through its wall into the scrotum. In the female this is replaced by a fibrous cord from the uterus. This gap is a potentially weak area where inguinal hernias can occur.
The muscles of the abdominal walls perform a variety of functions: (1) They provide a tonic, elastic muscular support for the viscera and, by their recoil, pull down the rib cage in expiration. (2) They contract against blows to form a rigid protective wall for the viscera. (3) When the glottis is closed and the thorax and pelvis are fixed, these muscles take part in the expulsive efforts of urination, defecation, childbirth, vomiting, and of singing and coughing. (4) When the pelvis is fixed, they initiate the movement of bending the trunk forward. Thereafter, gravity comes into play, the abdominal muscles relax, and the muscles of the back then take on the strain. (5) Conversely, the abdominal muscles come into play in preventing hyperextension. (6) When the thorax is fixed, the abdominal muscles can pull up the pelvis and lower limbs. (7) The muscles of one side can bend the vertebral column sideways and assist in its rotation.
The term arm is sometimes restricted to the proximal part, from shoulder to elbow (the distal part is then called the forearm). However, it is more commonly used to refer to both the proximal part and the distal part.
The bones of the human arm, like those of other primates, consist of one long bone, the humerus, in the arm proper; two thinner bones, the radius and ulna, in the forearm; and sets of carpal and metacarpal bones in the hand and digits in the fingers. The muscle that extends, or straightens, the arm is the triceps, which arises on the humerus and attaches to the ulna at the elbow; the brachialis and biceps muscles act to bend the arm at the elbow. A number of smaller muscles cover the radius and ulna and act to move the hand and fingers in various ways. The pectoralis muscle, anchored in the chest, is important in the downward motion of the entire arm.
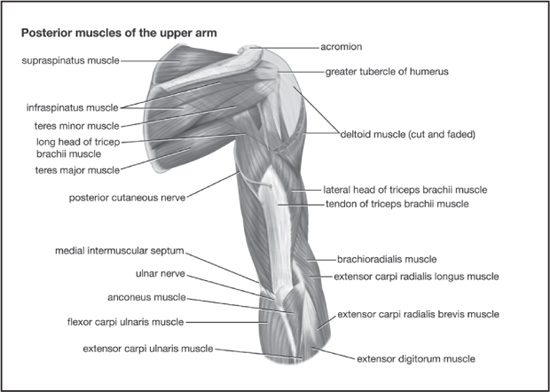
Muscles of the upper arm (posterior view). Encyclopædia Britannica, Inc.
The deltoideus muscle, or deltoid as it is commonly known, is a large, triangular muscle that covers the shoulder and serves mainly to raise the arm laterally. The deltoid originates on the outer front third of the clavicle and the lower margin of the spine of the scapula. Its fibres unite to form a thick tendon that inserts at the deltoid tuberosity, a rough spot above the middle of the outer surface of the humerus.
The biceps muscle is so named because it has two heads, or points of origin (from Latin bis, “two,” and caput, “head”). In humans, there are the biceps brachii and biceps femoris.
The biceps brachii is a prominent muscle on the front side of the upper arm. It originates in two places: the coracoid process, a protrusion of the scapula; and the upper glenoid cavity, the hollow for the shoulder joint. The tendon of this muscle is attached to the inner protrusion near the head of the radius, in the forearm. The biceps brachii bends the forearm toward the upper arm and is thus used in lifting and pulling movements. It also supinates the forearm (turns the palm forward or upward). The size of the biceps brachii is a conventional symbol of bodily strength.

Muscles of the forearm (posterior view). Encyclopædia Britannica, Inc.
The biceps femoris is one of the hamstring muscles at the back of the thigh. It originates in two places: the ischium (lower, rear portion of the pelvis, or hip bone) and the back of the femur. The fibres of these two origins join and are attached at the head of the fibula and tibia, the bones of the lower leg. This muscle extends the thigh, rotates it outward, and flexes the leg at the knee.
The triceps muscle is so named because it has three heads, or points of origin, particularly the large extensor along the back of the upper arm in humans. It originates just below the socket of the scapula and at two distinct areas of the humerus, the bone of the upper arm. It extends downward and inserts on the upper part of the ulna, in the forearm. Its major action, as noted above, is extension of the forearm upon the elbow joint, in opposition to the biceps brachii.
The gastrocnemius muscle and soleus muscle are sometimes considered the triceps of the lower leg (triceps surae).
The function of the leg is to support the body and to provide locomotion. The bones of the human leg consist of the femur, the tibia and the smaller fibula, and the foot, consisting of tarsals, metatarsals, and phalanges (toes).
The gluteus muscles are the large, fleshy muscles of the buttocks, stretching from the back portion of the pelvic girdle (hip bone) down to the greater trochanter, the bony protuberance at the top of the femur. The gluteus muscles include the gluteus maximus, gluteus medius, and gluteus minimus.
The gluteus maximus is the large, wide, thick muscle at the surface of the buttocks. It originates at the ilium (the crest of the pelvic girdle) and at portions of the sacrum and coccyx, bones at the base of the spine. It stretches across and attaches to the iliotibial tract, a band of fibrous tissue extending from the ilium to the tibia, and to the upper portion of the femur. Its major action is extension of the thigh, as in rising from a sitting position, running, or climbing. It also rotates the thigh outward.
The gluteus medius is located directly under the gluteus maximus. It originates at the back of the ilium below its crest and stretches downward to the greater trochanter of the femur. The gluteus minimus is situated under the gluteus medius; it also originates at the ilium and attaches to the femur. Both these muscles abduct the thigh—i.e., pull it laterally away from the midline of the body. In addition, their front portions help rotate the thigh inward, while their rear fibres aid in its extension and outward rotation. When one leg is raised off the ground (e.g., in walking or running), the gluteus medius and minimus of the opposite, fixed side act from below and exert a strong pull on the hip bone, even tilting up the unsupported side, which tends to sag when the limb is raised.
The quadriceps femoris muscle forms the large fleshy group of muscles covering the front and sides of the thigh. The quadriceps has four parts: rectus femoris, vastus lateralis, vastus medialis, and vastus intermedius. They originate at the ilium and femur, come together in a tendon surrounding the patella (kneecap), and insert at the tibia. These muscles extend the legs at the knee and are important for standing, walking, and almost all activities involving the legs.
The sartorius (from the Latin sartor, “mender”) is a long, narrow, ribbonlike thigh muscle that begins at the front of the crest of the pelvic girdle, extends obliquely down the front and side of the thigh, and inserts at the inner and upper portion of the tibia. It received its name because it is especially useful in assuming the cross-legged position that ancient tailors used in their work.
Actions of the sartorius include flexion of the thigh and of the leg at the knee and outward rotation of the femur.
The gastrocnemius muscle, also called the leg triceps, is a large posterior muscle in the calf of the leg. It originates at the back of the femur and patella and, joining the soleus (another muscle of the calf), is attached to the Achilles tendon at the heel. The gastrocnemius muscle works to pull the heel up, thereby extending the foot downward. This muscle provides the propelling force in running and jumping.
The soleus muscle is a flat, broad muscle of the calf of the leg that lies just beneath the gastrocnemius muscle. It arises from the upper portions of the tibia and fibula, the bones of the lower leg, and then joins with the gastrocnemius to attach via the Achilles tendon at the heel. Its major action is flexion of the ankle joint, particularly when the leg is bent at the knee, thereby extending the foot downward.