
I WENT TO visit a local sawmill. It was late winter. The loggers had been busy, cutting and skidding butts of oak and maple, hemlock, ash, and pine off the steep slopes, selling them at the mill. The yard was full of thick butt logs, separated by species, roughly piled.
It was icy underfoot, but the ice was starting to melt in places, making for dangerous walking on the rutted ground. The trunks varied in diameter between eight and about thirty-six inches, and the sections were between six and twelve feet long. The oak pile was the largest. It was mainly red oak (Quercus rubra), with some white and chestnut oak (Q. alba and Q. prinus).
A big yellow payloader picked up a bucketful of oak logs, carried them to the mill, and dropped them onto a bed of rollers that stepped them down an incline until they reach a sluice.
A log shot forward and rotary grinders—enormous steel cylinders full of hobnails—ground off the bark. The naked log was then shunted onto another set of rollers, in line for the saw.
The log fell onto the saw carriage. Steel saw dogs gripped it. The operator set the dimensions, and the log began to rocket back and forth across the upright circular saw blade, turning with each pass.
First the sapwood came off. It shot down a conveyor belt where a man unloaded it into a pulp machine. Then the saw started to take off boards and fire them ahead. The same man shunted them onto a field of rollers where another man roughly graded the boards: #1 common, #2 common, #3 common, #3B common, or one-and-better, select-and-better. When the machine had reduced the log nearly to the pith, it cut and spit out a four-inch-square cant.
The log was gone.
The whole process took little more than a minute. The next log was already on the saw.
Number 2 and 3 common oak boards are 50 percent or less clear wood. The rest is knotty or has other flaws. It is sent to plants where it is made into flooring. Number 3B—even knottier—goes to separate plants, where it is made into flooring for box trucks and trailers. The cants go to plants that manufacture packing palettes.
Number 1 common oak boards are two-thirds clear. Select-and-better is 85 to 100 percent clear. These are for the cabinetmaker. They go to manufacturers of kitchen cabinets; the very best boards to custom cabinetmakers.
It does not matter to the high-speed saw whether the wood it cuts is oak or ash, maple or poplar, hemlock or pine. Given the chance, it would try to convert an icebox into boards.
Industry did to craft what Babylonian agriculture did to gathering. It caused a tremendous increase in output, but at high human and environmental cost. Industry often starts as a liberator but ends as a slaver because it cannot control itself. Its solution is to make more and to make it faster. When sooner or later it falls into the hands of the inept or the greedy, they run it full tilt, heedless of the consequences until the soil is salted, the Dust Bowl blows away, the forests decline under acid rain, the market crashes, Bhopal suffers under a poison cloud, Chernobyl melts down.
But who would want to do without the brilliant high-speed saw? It does away with so much backbreaking labor. The waste of energy a man encountered after half a day of cutting and splitting when a log turned out to have a ring shake or a black heart is no more. Though one might still want to make cleft oak boards, one might not want to do it for a living. It is tedious and hard.
Is there no way to have the fast, beautiful tool without getting the headlong rush, the anxiety, the ennui, the cruel trampling of surplus people?
Human beings show restraint when they value, worship, and respect what they encounter. Value comes from understanding, and understanding from intimacy. Humans in the age of oak had to confront the resistance of their materials every day. Memory, reason, and skill wove a world of oak. The people understood, valued, and indeed worshiped the tree that was their intimate companion and source of so much of their livelihood.
The bread, the tracks, the henges, the churches, the houses, the roofs, the ink, the casks, the leather, the ships of oak are mostly gone now. Oak is for truck floors and middle-market cabinets. There is a small but growing market for fine oak furniture in the arts and crafts style, which shows the beautiful wavy grain of quarter-sawn oak. The buyers are mainly cultivated urbanites, nostalgic for honest craft and simpler times.
But oak is not forgotten. Natural scientists, ecologists, and engineers remember it. They do not break down the oak tree into its genetic components, then recombine them and patent the results. Instead, they closely examine the tree in nature, to learn from its own life history and its own design.
In the 1970s it occurred to scientists that the annual rings in oak trees could tell time. Formed each year during the growing season, these rings vary in thickness according to environmental conditions. A cold year or a drought year, for example, will produce a much narrower annual ring than a warm year or one with abundant rainfall.
Archaeologists realized that the annual rings formed by different oaks in the same area would have similar patterns of wide and narrow rings, because they had experienced similar conditions. If they could learn to read the codes in the rings, they could in theory start from oaks today and—link by link, matching pattern to pattern—count back toward the beginning of humanity. The assembled patterns of the oak chronology would then allow the archaeologists to date precisely any oak artifact found at a site.
Chronologies are actively being created in Ireland, England, and Germany. Among them, they now allow dating that goes back almost ten thousand years, to the very beginning of the Holocene. And unlike radiocarbon dates—which at best have an accuracy of plus or minus ten years—these dates are exact. A good dendrochronologist, as the scientists in this new discipline are called, can even say in what season of the year the wood was cut.
What use is such exactness? For one thing, it will allow students of the past to know if neighboring archaeological sites were really neighbors or if one succeeded another. It has also begun to produce unanticipated help in interpreting the environmental history of the Holocene. Some narrow ring patterns found in many of the existing oak chronologies correlate with probable major volcanic eruptions and records of darkness, famine, and other portents, from as far away and as deep in time as the Shang dynasty in China, roughly 1750 B.C.
Recently, too, close observation of the natural laws of tree growth has begun to provide models for design. No tree is able to hold out heavier and longer branches for as long and as safely as an oak. Klaus Mattheck studied oaks and the other trees in German forests to learn how they could do it. From his observations, he postulated the Axiom of Uniform Stress, which states that trees actively grow to equalize the stresses over their entire surface. Wherever a dangerous stress point occurs, the tree grows new wood to control and reduce it.
Mattheck cleverly invented a way to simulate this adaptive growth on any object and modeled it. The method—called Computer Aided Optimization (CAO)—reveals the high-stress points in any designed object and treats them as though they were the parts of a living tree. Like a tree, the program “grows” new tissue for the component at the points of high stress until the dangerous weakness is eliminated. The product is then manufactured using the growth data. The sort of surgical screw used to mend complex bone fractures, for example, is now routinely made using this method. It lasts twenty times longer than did earlier products.
The study of oak itself, then, is a school of history, design, and society. There is no structure more supple and sustainable than nature’s, and oaks are among the most widely adapted and successful of all plants. Close attention to oak—the way it is built and the way it works—may lead to more sustainable designs for everything from clothes to transportation systems.
Just as students of early humans discovered that people did not in fact hunt big game to extinction in order to establish mankind, so those who study oak are finding that the tree’s success is owed not to the ruthless suppression of competitors but to its flexibility.
Diversity
 The history of our planet is the story of radical changes in landscape and climate. Continents split, moved, and collided. Temperatures, seasons, rainfalls, and ice sheets rose, fell, appeared, disappeared, shifted. The botanist Edgar Anderson referred to such changes as “hybridization of the habitat.”
The history of our planet is the story of radical changes in landscape and climate. Continents split, moved, and collided. Temperatures, seasons, rainfalls, and ice sheets rose, fell, appeared, disappeared, shifted. The botanist Edgar Anderson referred to such changes as “hybridization of the habitat.”
It was the great virtue of oaks that they responded not by specializing and narrowing their range, but by adapting, expanding, and radiating into more and wider-flung landscapes. There have seldom been creatures so tenacious as oaks, but their staying power is founded on their own ability to change.
If you suggest to any two oak taxonomists today how many species there are, you will start a fight, because they will never agree. Some people think there are 450, others 250, and most think there are any number in between. Some say that species are really hybrids or hybrids really species. But even we casual observers can tell them that there are lots and lots.
All oaks have acorns, but sometimes the acorn is about the size of a .30-06 rifle shell, and when it falls from the tree on your head, it feels like one. Others are fat and round like cherry bombs. (A friend who lives in a red oak wood outfits his kids with batters’ helmets when they go out to play in autumn.) Some acorns are so small you could mistake them for martini olives; others look like dark brown Christmas lights.
The caps that attach acorns to their stems are equally various. Some are set on the seed like a beret two sizes too small; others are like modest grass skirts that cover practically the entire fat delicious pod; others are frilly and revealing. Some are hairy, some are scaly, some are knobbly, some are woolly. Some resemble the helms of kings, others peasants’ caps.
And what of the trees themselves? One’s crown is 120 feet wide and almost as high. Another is a single pillar rising ninety feet, with not one branch until sixty feet. One never grows higher than your knee but spreads like crabgrass; another throws its top branches skyward and its bottom branches earthward like a dancer caught in two poses at the same time. One lives in swamps where crawdads dig their burrows among the roots; another lives in uplands where there are prolonged dry spells. One has leaves as small as your thumbnail, another as large as a napkin. Some of the leaves are spiny and hard like the sloughed exoskeleton of a beetle. (If you sit on one, you get right up again.) Others are as fine and delicate as bible paper. Some hold their leaves for years, others hold them from spring to spring, and some lose them every fall.

A few acorns (Nora H. Logan)
But for all that there are many species of oak, each one differs comparatively little from its near relatives. Among the genus Quercus, only a few genes can make the difference between one species and another. Out of seven hundred genetic markers, for instance, only six distinguish Quercus grisea from its neighbor, Quercus gambelli. Oaks are very adaptable, because small genetic changes can make noticeable differences.
If you ask an experienced student of oak trees to go for a walk in the woods with you, you may end up as puzzled as he. He may see clearly here a pin oak, there a red oak, but he will likely stop before a tree and say, “Well, this seems to have some red and some black oak in it,” or “This is kind of a pin oak, but kind of a willow oak, or is it part scarlet oak?” It is always possible to distinguish the predominant type, but one often sees variations.
Oaks make frequent small genetic changes. Each change may be almost unnoticeable in itself, but over centuries, they can have major effects. Different species can pass genes back and forth, and through generations of crosses and backcrosses, new characteristics—both in the habit of the tree and in the habitat it prefers—come to be.
Often, this ability seems mere genetic doodling. A slightly scarlety pin oak seems to have no advantage in the landscape over a slightly willow-oaky pin oak. But the process is powerful over the long term. Many people think that when two species share genetic information the result will be a plant whose characteristics are halfway between the two. This is not necessarily so. A recombination of genes can also produce wild distinction, children with little resemblance to either parent. Most will not even be viable. They may want a nonexistent climate, or they may be sterile. But a few would be viable, if only they had the opportunity.
The few viable alternatives usually live out their lives and cross back into the next generation by sharing their pollen or their seeds with their parents’ species. This makes for little external change—just enough to annoy or delight oak walkers in the woods—but underneath, at the genetic level, a mosaic is building. Molecular-level studies of oak populations show that even apparently identical trees often have chloroplasts containing the DNA of different species. When major environmental changes happen around oak trees, this genetic richness allows them to adapt quickly. These genetic self-experiments perpetuate a fund of possibilities, waiting a place to be used.
Tenacity
 Around sixty-five million years ago, at the beginning of the Paleocene epoch, a nut fell to earth somewhere in what is now Thailand. A small hairy creature that looked like a cross between a squirrel and a rat worried the nut between its paws. It carried the nut away and buried it in a clearing. A long-lived member of the Fagaceae—the beech family, which includes all the oaks—will make at least three million acorns in its lifetime. This squirrel-rat had buried about two hundred that season. And it never again found that particular one.
Around sixty-five million years ago, at the beginning of the Paleocene epoch, a nut fell to earth somewhere in what is now Thailand. A small hairy creature that looked like a cross between a squirrel and a rat worried the nut between its paws. It carried the nut away and buried it in a clearing. A long-lived member of the Fagaceae—the beech family, which includes all the oaks—will make at least three million acorns in its lifetime. This squirrel-rat had buried about two hundred that season. And it never again found that particular one.
Water penetrated the seed coat and activated the miniature plant waiting within. Using the food stored in the seed, the radicle—the proto-root—penetrated downward through a fissure in the bottom of the nut. The radicle senses and follows gravity, while sensing and fleeing sunlight. Turn the nut upside down in the ground after the radicle has sprouted, and it will reverse direction to keep growing downward. This first root spiraled more than a foot into the soil during the plant’s first year. Already it was doing more in the way of roots than its ancestors did.
Otherwise, for about twenty years, the tree pretty much resembled its forebears. When it finally became a sexual adult, however, there was a startling change. When the male flowers came out, instead of standing up on the stems, ready to receive insect visitors, they hung, indeed they drooped, and waved in the wind. This tree, unlike its progenitors, had reverted to the more “primitive” wind pollination favored by the gymnosperms. And the nut, instead of being fully defended by a spiny husk, was open and smooth, with just a little cap to join it to the stem.
It was the first oak.
And in one regard, it was really disappointing.
Such a magnificent tree ought to have had a beautiful flower, at least the equal of the cherry or the magnolia or the tulip tree. But no. Fine flowers are for plants that need to attract, massage, and reward insects. The male oak flower looked like a few strands of kinky, limp thread. Today, most people don’t even know it is there. (When the flowers fall, a few of my clients complain about the “droppings.”) Even the pollen is lightweight and inconspicuous. And the female flower, hidden in the angle between twig and stem, looks like a pimple. It is hard to believe that something so large and long lasting could come from such a nondescript beginning.
Nevertheless, the male flowers made good use of the wind. They ripened, filled with yellow pollen grains, and then released a golden rain that floated for miles and miles through the countryside on the wind’s convective whorls. With luck, they would strike the stigmas of the tiny female flowers hidden in the leaf axils and new acorns would begin.
From this single beginning on the edge of a proto-continent that included present-day Thailand, the oaks began their travels through changing landscapes and through generally worsening weather.
Land masses spun apart and undersea ocean ridges—the sutures of tectonic plates—spread. Pressure built up on coastlines. Mountains rose. In the shadow of these mountains, it was drier and the winters were a little colder. It was not a good climate for dependable insect pollination. If it were too cold or too dry, there might be a poor hatch, then a poor crop of bugs, then poor fertilization of the flowers. Wind operated under any circumstances and assured that at least some fresh acorns would be made. The oaks spread across Asia within ten million years.
But to the west—where Europe and North America had once been connected to Asia—the forests were blocked. The ancient Turgai Sea stood in their way.
In the middle of the Eocene, a bridge opened across the Turgai when moving continents brought two peninsulas together. The oaks streamed across and into Europe. There, although the Atlantic was becoming an ocean, a mosaic of bridges still connected what would become Europe to what would become Scandinavia, Greenland, and North America. These bridges themselves opened and closed as the lands jostled, but the oaks moved into North America.
As late as forty-five million years ago, it was quite warm in the middle of the world. The zone of the tropical rain forests stretched as far north as the present northern boundaries of California, at forty degrees north latitude, and even into coastal Alaska. Broadleaf evergreens, including the oaks, were dominant.
Today, there are more than 130 species of oak in mountainous northern Mexico. It is a world full of oak. If you stand in the middle of them, you will get a good idea of what the world was like in the Eocene epoch, for all these oaks or their near relatives once lived far abroad across every continent. In fact, the fossil leaves of oaks found as late as the Eocene in Italy are precisely the same as the leaves of oaks that live today in southern California and northern Mexico.
As the Eocene ended, about thirty-seven million years ago, pieces of the world started banging into each other again. The Pacific Rim pushed against North America, the number of volcanoes increased, and the Rocky Mountains began to rise. The sea that had divided one half of North America from the other drained away and dried. Meanwhile, halfway around the globe, the Indian subcontinent at last slammed into mainland Asia and pushed up the Himalaya. In southern Europe, the Alps began to rise.
Suddenly, about thirty million years ago, the most dramatic cooling that the earth had known in the past one hundred million years occurred. It was a disaster for much of what had come before. It was also the foundation of the modern world. When it was done, the mean temperature of the globe had declined by almost thirteen degrees Celsius.
A permanent ice cap began to cover the earth’s high latitudes. The oceans for the first time acquired cold-water depths. Cold and dry seasons became longer, more pronounced, and more unpredictable. The cold flowed quickly down the troughs made by north-south mountain ranges, such as the Appalachians and the Rockies. It was slowed where it met east-west ranges, such as those of Asia and southern Europe. The Turgai Sea at last disappeared for good, and Europe was firmly connected to Asia.
Insect pollination, which had seemed such a sure thing, now was less secure. A lot of pollinators died out altogether, while the survivors depended on the mercy of the weather and tended to remain where the climate stayed most like it had been in the old days. Some plants—including the oldest oaks—took refuge in the places where the climate changed little. But new oaks appeared.
In North America, where the cold penetrated farther south, the oaks evolved a new strategy. If the climate were too unpredictable to make sure that an acorn would mature in one season, why not give it two seasons?
The red oak tribe came into being. Today, all true oaks belong either to the white oak or the red oak group. The white is older. Its acorns are fertilized and mature in one season. The leaves of its member species are smooth at the tips of the lobes. Red oak species have leaves with spiny tips and take two seasons to mature an acorn. In the first year, the wind-borne pollen grains reach the stigmas of the tiny female flowers in early spring. Each grain bores down the stigma toward one of the six ovules hidden deep inside. But as the first autumn sets in, the pollen stops partway down the tube. Next spring, soon after it warms, the winning grain reaches its ovule immediately and the acorn begins to ripen. It would be hard to imagine a better way to beat a climate that was progressively less predictable.
After the great cooling, the weather briefly warmed, but soon it cooled again. In the north, it got too cold for a tree to be a broadleaf evergreen anymore. The winter drying of leaves and the burning caused by cycles of freeze and thaw made evergreen leaves (not needles) inefficient. So the oaks changed again. For the first time, some became deciduous, losing their leaves and hardening off their tissues in preparation for the cold season of each year.
This was a risky change. It meant an energy economy that had to be strictly regulated. Now there was a period of the year during which no photosynthesis occurred, and the stored energy of the previous year would have to be sufficient to pump out a whole new crop of leaves each spring. During the time immediately following leaf-out, the tree would be severely energy depleted. An attack of pests or disease at this time, or the failure of the sun to shine, could have disastrous results.
In the last million years, the climate has had at least two dozen sharply defined pulses from warm to cold and back again. Within these, perhaps six have been deep and prolonged, lasting from forty thousand to one hundred thousand years each. At the low points, glacial ice could form at elevations more than three thousand feet lower than it does now, and masses of ice swallowed the entire landscape of the Northern Hemisphere down to about the level of Seattle, St. Louis, and New York in the United States; Amsterdam, Warsaw, and Berlin in Europe; and Kiev in West Asia. At the high points, the climate was more or less as it is now, sometimes even a bit warmer.
With each advance of ice, the oaks retreated south into places like the Mediterranean basin, the valleys south of the European Alps, Southeast Asia, and southeastern and southwestern North America. With each warming, they returned northward. The peaks of warmth in the interglacial eras were steep and narrow, and so were the troughs of cold in the ice times. No interglacial era in the last million years has lasted more than twenty thousand years.
The depth of the most recent ice age was nineteen thousand years ago. The rolling glaciers scored rock, broke it to pieces, and rolled the hunks beneath its weight until the rock was ground to the texture of flour. Even beyond the reach of the glaciers proper, repeated freeze-thaw episodes shattered stone and left fresh shards exposed to weathering.
Twelve thousand years ago, the climate began quite suddenly to warm. Fresh, completely unleached soils liquefied beneath the melting glaciers and flowed over the landscape. Everywhere that there was a slope water flowed, bearing new soils in its wake. Swirling winds lifted up the lighter of the dusty stuff and carried it hundreds of miles, spreading it over the landscape. This was the stage onto which humanity emerged, among the tenacious oaks.
Cooperation
 A jay appears out of nowhere and lights on the branch of an oak tree. It preens a little, looks about. It is October, a windy day with long tattered clouds streaming overhead, and a great meal of thousands of acorns is ripened on the branches and scattered on the ground beneath the tree. The jay looks over its shoulder, then picks an acorn. It shakes the nut, turns its head a moment as though reflecting, then drops that acorn. (A weevil had already eaten most of the seed inside. It was too light and it rattled.) The jay looks around once more, hops, seizes another. Holding the acorn securely in its long, bendable toes, the jay pushes its hooked upper bill through the shell. It clamps down with the lower jaw, then presses, twists, and tears until the shell comes off. Then it chomps and swallows the rich seed.
A jay appears out of nowhere and lights on the branch of an oak tree. It preens a little, looks about. It is October, a windy day with long tattered clouds streaming overhead, and a great meal of thousands of acorns is ripened on the branches and scattered on the ground beneath the tree. The jay looks over its shoulder, then picks an acorn. It shakes the nut, turns its head a moment as though reflecting, then drops that acorn. (A weevil had already eaten most of the seed inside. It was too light and it rattled.) The jay looks around once more, hops, seizes another. Holding the acorn securely in its long, bendable toes, the jay pushes its hooked upper bill through the shell. It clamps down with the lower jaw, then presses, twists, and tears until the shell comes off. Then it chomps and swallows the rich seed.
As if on cue, more jays appear in the area and make for this wonderful tree. As soon as he sees them, the first jay changes his mind about eating. He explores the branches, testing for sound acorns. When he finds a good one, however, he swallows it whole. This is a startling thing to see for the first time, since no one would think that such a small bird could swallow such a big thing without choking to death. In fact, he is not swallowing it all the way down. He is storing it in the expandable throat that all jays have evolved for this purpose. A jay can store up to five acorns—depending on their size and his—in this container. He stuffs in as many as he can fit, takes a final acorn in his beak, and flies away.
The other dozen jays who have found the tree begin doing just the same thing. Test, select, swallow, select, swallow, test, yecch, select, swallow, get one more, hold, fly. Each jay makes many trips to collect the fruits, harvesting both in the tree and on the ground.
While many acorns remain, the gathering is amicable and practically silent. Each bird is occupied with his or her task. When only a few sound acorns remain, however, mayhem ensues. Jays fight over the last acorns, screaming, and when the victor finally flies away in triumph, he is often pursued by more than one bird, intent on later theft.
The tree is left alone, the rich supply of thousands of acorns reduced to a few hundred, almost all of them unripe or already parasitized by weevils or fungi or both. It is quiet again, except for the sound of the wind rattling the drying brown leaves.
The oaks evolved about sixty-five million years ago. So did the jays. There are still more different species of oaks in places like Mexico’s Sierra Madre than in any other. Ditto for the jays. Jays and oaks are a couple. Essentially, the oaks have domesticated the jays, or vice versa.
The list of animals and fungi that feed on one part or another of the oak would fill a small phonebook, but most just take what they need. The oak is their host, accustomed to bearing a lot of hungry guests. Indeed, every part of the oak is filled with a bitter substance, tannin, meant to control the number of guests or at least reduce their appetites. Some give as good as they get—certain fungi give water and nutrients in exchange for the food they take, and squirrels act on oaks much as jays do, spreading and planting acorns, eating maybe one for every three they bury and lose. But oaks and jays have been key to each other’s survival since their beginnings and so to each other’s spread around the globe.
Jays and oaks make a strange couple. We are accustomed to admire the oaks, but what about the jays? In one place, they are gray blue and look permanently dusty. In another, they are blue black with pointed crests that make them look in flight like animated arrowheads. A third group have shoulder stripes like lance corporals, and still others are a nice orangey-tan color decorated with blue sleeves. But wherever they may be and however they may appear, jays always make too much noise, an insistent, raucous, grating caw. On the edge of a wood, it is almost impossible not to stray into jay territory, and whenever you do, you will certainly hear about it. Jays screaming is the aural equivalent of mosquitoes biting. Indeed, the European jay has the genus name Garrulus, that is, “talks too much.” But if we are going to admire the oaks, we had better learn to admire jays too, for between them they have changed the surface of the earth.
Jays do the one thing oaks can’t. They move. All the time, it seems, they move. When we are passing through their territory, their flights look flitting and purposeless, though likely they are meant to distract and annoy us, to make us go away. But if you settle down and stay awhile, you may begin to see what they are doing.
The jays carry acorns away, usually to areas near where they are nesting. There, on the edge of the woods or in any case on some edge—it may be between wood and field, between tall grass and low, between garden and lawn—they bury their found acorns, not in groups of nuts, but one at a time.
The jay pushes each acorn into the soil as deep as it will go. Then it hammers with its bill until the nut is fully covered. Sweeping side to side with its bill, it covers the nut with soil and sand. It then picks up a few leaves, twigs, or pebbles and artfully scatters these over the hoard.
The average jay will bury more than forty-five hundred acorns each autumn. They are the adults staple food through the winter months, and serve both adults and fledglings in the spring. But how easy can it be to find thousands of nuts that were hidden weeks or months before?
I. Bossema studied the European jay intensively, concluding that jays buried on edges largely so as to have landmarks by which to locate their caches. Furthermore, they buried the acorns one by one—not in a single cache—so that if a scavenger found one, it would not necessarily get them all. That they could indeed find the nuts again was first demonstrated to Bossema by chance in his own garden as he reported in his 1979 study.
In the middle of October I observed a marked jay hiding food in my garden. This bird had three acorns in the aesophagus and carried a fourth one in the bill. It concealed the fruits on four different sites: the first one close to the trunk of a tree and the others along the edges of the lawn. I did not observe this jay at this place again until 72 days later. It landed in front of the hiding place close to the tree trunk and located the acorn precisely without hesitation. It flew into the tree and dehusked and ate the acorn completely. Thereafter the bird flew to the lawn, approached the edge, dug briefly twice within [a few inches] of a hiding place and found the acorn at the third attempt. After dehusking and eating it in the same tree, the bird disappeared and was not seen again in this part of the garden. In May the remaining hidden acorns produced seedlings.
Even the young seedlings are of use to some jays. When the sprouts emerge, jays may lift them, shaking them in their bills and dislodging the food-rich cotyledons that persist underground in what remains of the acorn. This procedure often kills the seedling, but as often it does not. In the spring, what’s more, jays often feed their newborn on a nice pulp made from the larvae of oak leaf–eating insects.
The oaks make staple foods for the jays and have done so for epochs. Jays have changed both physically and mentally to adapt to this way of being. The V-shaped hooked upper bill is adapted to tearing husks, the expanding esophagus to storing acorns for transport. Jays have evolved a lower bill that is attached to their headbones so that the bill cannot be dislocated when hammering. And jays’ breeding is timed to coincide with periods when food will be available from the oak. Mentally, jays have developed an extraordinary ability to remember landscape features to be able to find again the acorns that they have hidden.
The jays do well in this deal, as they loudly proclaim. But what about the oaks? It’s very nice to have a predator eat the larvae that would otherwise eat your leaves, but not so nice when the predator seizes your fruit and eats it, or manhandles your seedlings. Yet the fact is that the jays are the world’s great cultivators of oak and a principal tool in the oak’s spread and dominance.
Jays carry acorns out of the deep shade of their adult trees, where they could never thrive, and plant them on forest edges, where the acorns have plenty of sun but some protection from wind. They plant the acorns in soils that are neither compacted nor waterlogged, since both resist a jay’s effort to place the nut. The acorns have a good soil in which to grow, one that will not subject them to fungal infection or to asphyxiation. The jays cover and conceal the acorns, making them a less easy prey for the deer, mice, voles, turkeys, bears, porcupines, badgers, and others that love to feast on them.
On average, for every four acorns that the jay hides, it recovers only one. Even if many more are found by various other predators, or become infected and rot, or fall prey to jay foraging after they sprout, it is thought that at least a couple hundred out of the average jay’s annual hoard of acorns survive to become viable seedlings. It doesn’t take higher mathematics to see what this can mean for the propagation of oak forests.
Students of the postglacial landscapes of North America and Europe have long been puzzled by how fast the oaks recolonized the areas left bare by retreating glaciers. The ordinary route of spreading by wind pollination—however adaptable the oak genome—could not account for the one- to two-mile-per-year dispersal rates indicated by the distribution of fossil oak pollen. Neither could squirrels, who compete with the jays to collect and hoard acorns and so also help to propagate oaks, but who only take an acorn about one hundred feet from the source.
But a jay can go almost a full mile before it decides to bury its throatful of acorns. With the rain of light oak pollen following, it just might happen that oaks would appear—on the scale of plant migration—to be running at top speed northward. They have probably done this again and again, ever since oaks and jays left Laurasia.
So it is hard to admire those strong, stable oaks in the park or street or woods or chaparral without also admiring the jays. Those things that scream at you, dart overhead, and occasionally drop an acorn or something worse on your head give the oaks legs.
Flexibility
 There are no deciduous pines, no evergreen maples. Nor are there any deciduous spruce, nor evergreen ash, no deciduous junipers, no evergreen lindens, no deciduous cedars, no evergreen dogwoods. But there are both deciduous and evergreen oaks.
There are no deciduous pines, no evergreen maples. Nor are there any deciduous spruce, nor evergreen ash, no deciduous junipers, no evergreen lindens, no deciduous cedars, no evergreen dogwoods. But there are both deciduous and evergreen oaks.
Where I grew up, on the coast range of northern California, it snows once in fifty years or so. Most, but not all, of our oaks were evergreen. An unclothed oak was a dead oak, a very sad thing in my childhood. I assumed therefore that most oaks were eternally green, but the few fallen dead leaves interested me. They might appear on the ground, a dozen at a time, usually in autumn but at other times too. Most were from coast live oak (Quercus agrifolia), and they were cupped, waxy, stiff, and sharp-tipped. Usually they had turned a pale yellow, with the black spots of some fungus that had helped to kill them. If they fell tips down, they could scud along the ground in the wind like scooting insects. If they fell tips up, they pricked bare feet or surprised the unwary who sat down on a chair set beneath the oaks.
Now I live in the northeastern United States, where all of the oaks are deciduous. But here I notice the ones that refuse to shed their leaves. Most are pin oaks, young ones, but older ones and even other oak species sometimes retain their dry, dead, withered, stiff, tan-brown leaves. They are my winter companions. No one who has lived all his life in a place like coastal California can understand the variety of noises in winter woods where deciduous and evergreen trees are mixed. You can tell where you are by the distinct winter sounds.
Wind awakens the snowy woods. A grove of hemlock or spruce make a whishing sound in the wind, and snags and stems creak. Deciduous forests make deeper rising and falling sounds. If a tree has a crack in it, the rubbing parts may make sounds like gunshots, enough to make a walker start and turn.
The homeliest winter sounds, to my ears, are made by the dead, persistent leaves of the oaks. Not happy, but homely. They rattle and hiss, like a kid sliding on gravel or kicking it. I can look and see which tree is making the sounds. Each tree sounds different. I find I am proud of their persistence, and I always promise myself to notice when the leaves finally fall. (If they didn’t, the new ones couldn’t come.) But I have not caught them at it yet. I turn around one April morning, and there are the dangling flowers and the mouse-ear beginnings of the new leaves.
The evergreen oaks evolved for warm climates, whether wet or dry. Photosynthesis continues all year in these climates, so there is a continual need for water to climb the stems, carrying minerals and water to the sites of food and energy manufacture. The leaf surfaces tend to be waxy, preventing sudden water loss and wilting; the leaves are usually whole, not lobed, meaning that they both heat and cool more slowly; and the air resistance that impedes transpiration tends to be greater over these whole leaves than it is over thinner or lobed leaves. Often, the leaves may be slightly cupped, protecting the water-transmitting stomates on their undersides from drying winds and steadying the rate of water loss.
This outer difference has a corresponding inner difference. The evergreen oak, unlike its deciduous relations, does not show a pronounced shift between the large-diameter vessels of early wood, the wood produced at the beginning of the growing season, and the small, thick-walled vessels of late wood, the wood produced during the latter part of the growing season. The vessels that serve as the tree’s circulation system are scattered more evenly through each annual ring.
Less water moves through these smaller pores, and it moves more slowly. Yet also it moves more surely. The smaller vessels are less likely to develop gas bubbles that break the water column, rendering that channel useless. Most of the evergreen’s vessels function not only during the current year, but for several years after as well. The evergreen system creates a slow and steady pulse of life.
The deciduous oak represents the opposite choice. Though evergreen and deciduous oaks occur together in many climates, the deciduous comes into its own where the weather has a distinct cold season. When a deciduous oak’s leaves first unfurl in the spring, they signal the cambium, the tree’s growing layer, as far away as the root, and very quickly, a new ring of xylem cells, those which will conduct water from the ground to the leaves, blow up to more than five hundred times their original size. At the same time, xylem tissue starts expanding from each leaf bud’s base downward, until the two meet and a new year’s circulation system is inaugurated. Each xylem cell dies soon after it expands. Evacuated of a cell’s usual internal contents, the xylem cells join end to end and in millions of combinations—vertically, horizontally, and spirally—to create a labyrinth of rising pipes.
The suction generated in an oak’s xylem can be four times greater than the minimum needed to lift water from the root system to the top of the tree. The system makes a vacuum more perfect than any that can be made in a laboratory. When the sun is warm and water is available, more than four gallons of water per hour may pass through a single eight-inch diameter trunk. In high spring, the rising sap reaches velocities of two hunded feet per hour, or better than three feet per minute.
As the season progresses, though, most of these enormous sap superhighways suffer embolisms—bubbles of gas that block the water’s flow. Sometimes this is caused by variation in pressure from day to day and season to season, but often it may be caused simply by too much tossing of the branches in the wind. When one vessel is blocked, of course, another path will be sought, but by summer, a large proportion of paths are likely blocked.
In a seeming anticipation of this, the deciduous oaks start making the much smaller, thick-walled vessels of late wood in late spring. After the spring binge, the deciduous trees become more like their cousins, the evergreens. Though the smaller vessels are slower to transmit the materials from the soil, they are surer to be passable, since their small size keeps pressures high and discourages bubble formation.
The size and shape of deciduous oak leaves helps to regulate their water economy. Slender leaves, like those of the southern live oak (Quercus virginiana) or of the aptly named willow oak (Q. phellos), experience less air resistance and so are able to rapidly give off water vapor. At the same time, however, their comparatively small size makes them quicker to cool, so that their rate of flow slows rapidly in the evening. The large, lobed leaves of red, black, or white oak (Q. rubra, Q. alba, and Q. velutina) experience more air resistance, but they heat up more and are capable of transpiring phenomenal amounts of water. But because the deeply dissected lobes expose more surface directly to the air than would unlobed leaves, these comparatively large leaves also cool quickly at evening.
If you had to characterize the difference between evergreen and deciduous oaks, you might say that the former were adapted to the marathon and the latter to wind sprints. And between these two pillars exists every possible oak intermediate. There are small-, whole-leaved deciduous oaks, and huge-, lobed-leaved evergreen oaks. There are oaks like the southern live oak or the pin oak (Q. palustris), whose leaves are not quite evergreen but are “persistent,” remaining on the tree until the new leaves appear. There are oaks like the deciduous black oak, with immense, lobed leaves, whose pubescent hairs are thought to help cool the leaves’ engines. There are oaks like the swamp white oak (Q. bicolor), which appear undecided as to whether to have lobes or not, and others like the post oak (Q. stallata), which appear to be trying to turn their leaves into small crosses. There are oaks like the English oak (Q. robur), with slender, shallow-lobed leaves that seem ready to wilt if you simply looked crossly at them, while the leaves of the bear oak (Q. ilicifolia) are so tough, spiny, and arch-backed they look like thousands of tiny reptiles about to attack.
One can’t help thinking that there is an element of play in these large variations, but it must be adaptive, directed play. In the very same habitat, there seem to be oaks that bet one way and oaks that bet the other. Generally, tree architecture tends to specialize either in outracing others for dominance in the forest canopy or in spreading fast to crowd the others out. The oak does both.

A few oak leaves (Nora H. Logan)
Up in the folded hills of the northern Sonoma wine country, I found very tall deciduous California white oaks (Q. lobata) living alongside wide-spreading evergreen canyon live oaks (Q. chrysolepis). On one homestead on a Pomo Indian reservation, the tallest tree in the neighborhood—a white oak—lived across the street from the widest tree in the neighborhood—a live oak. The latter was so broad that the owner had built a fence around the drip line and made the enclosure into a hen coop and pigsty.
Prudence
 In the tale of Sir Gawain and the Green Knight, the evergreen holly giant does not need saving. It is Gawain, the deciduous oak, who has to be saved from death. This is neither a quaint legend nor an archaic understanding. It tells truly and shortly the reality of leaves.
In the tale of Sir Gawain and the Green Knight, the evergreen holly giant does not need saving. It is Gawain, the deciduous oak, who has to be saved from death. This is neither a quaint legend nor an archaic understanding. It tells truly and shortly the reality of leaves.
Leaves are not decorative. Not for trees and not for us. Little but microorganisms would be alive on earth were it not for leaves. When the leaves of deciduous trees appear and expand each spring, they signal a new season of harvesting energy from the sun. None but they can do it. Without them, we all die.
The sprouting of the new leaves is called “flushing” or a “flush.” Some trees have just one flush of new leaves, in the spring. Others manage two flushes. Some add leaves continuously from spring through fall.
All of these methods have advantages and disadvantages. One flush may be fine if spring is dependable, but bad weather just at that time may result in many damaged leaves and so seriously reduce the plant’s ability to make food for the rest of the year. Two flushes provide some insurance against bad weather, but bad luck is still too likely. You would think that continuous growth would solve the problem, but the trouble is that it costs a great deal of energy to keep pumping out new growth. If bad weather is concentrated in one season, repeated efforts to send out leaves can seriously reduce the tree’s energy reserve.
The deciduous oaks have evolved a compromise that takes advantage of the energy economy of discrete flushes and the long time frame of continuous growth. The deciduous oak may flush up to four times: in early spring, early summer, high summer, and sometimes one in the fall. In other words, an oak may sprout up to four times from its buds each year. The discrete flushes do not depend on the success of the previous flush, but the tree rests between flushes so that it does not waste all its energy against a climatic anomaly. In this way, even in a bad-weather year, the oaks have a chance at growth and new life. And if conditions change for a number of years in a row, they can adjust the number of flushes to fit the new climate.
At each flush, a new set of tip and lateral buds is formed in the terminal bud of the previous flush. When the flush hits, the terminal buds open and leaf out at the expense of the lateral buds, which usually remain dormant. This is why oaks may seem to change color in midsummer: The suddenly expanding new growth may be reddish or yellowish in color, and because it is held out stiffly on the terminals, it seems to be a whole fresh set of leaves covering the trees. The laterals behind offer no competition.
What is happening behind this bright show, however, accounts for a lot of the oak’s adaptability and its wide, space-filling growth. The strongest lateral buds of the previous flush—which had done nothing when their tip buds were expanding—now sprout and expand. Behind the out-thrust scouts of the new terminals, then, phalanxes of lateral twigs appear all around the branch. And what’s more, these laterals may outgrow the terminals, the main shoots. So behind the up and outward thrust of each succeeding flush, the oak completely occupies the recently conquered space.

Oak buds (from H. Marshall Ward, The Oak)
The oak is profligate not only in the number of lateral buds that grow, but also in the number of lateral buds that do not. Typically, only the outermost and strongest laterals grow, leaving more than half of them dormant. This means that in the average mature oak, perhaps a million buds will never sprout at all. Liberty Hyde Bailey, the American horticulturist whose fine observations and careful illustrations of growing stems alone are worth the price of his books, thought this a fine demonstration of the fact that nature was red in tooth and claw. The buds, he said, compete for the right to grow, but only the strongest and fittest actually get to.
But this is not exactly so. The dormant buds are not the losers; they are part of the reserves. They may remain viable for more than a year, and if necessary—say, after the loss of part of a branch in a storm—they sprout. And where dormant buds fall, the tree can produce new buds called adventitious buds, from scratch.
This reserve was crucial not only to the oak’s survival, but to the health of human communities. The wooden world depended on oak for everything from firewood to structural timber, and much of it was obtained by cutting the tree back—either to major branches or right to the ground. You could count on the oak to regenerate strongly from these cut ends and create fine timber within a decade.
Dormant and adventitious buds are also part of the oak’s persistence and its flexibility. Humans are not the only agents that damage or destroy branches. There are also caterpillars, beetles, twig girdlers, borers, cankers, rots, thunderstorms, ice loads, hurricanes, goats, cattle, and sheep. And who knows what else feasted on the tender leaves and twigs of oaks in prehuman times? The buds are the oak’s answer to destruction. They are the sites from which, no matter what, growth can be renewed.
They are also a gift to its decline. No tree grows old so beautifully as the oak. If you could compress nine hundred years of time into a few minutes, you would see an oak as a firework, shooting up, expanding, and then like the best rockets, seeming to sink back into itself while a rain of green drops all around.
In its sinking, the oak becomes stag-headed, its dying leaders sticking out like the horns of a buck. The branch ends begin to die and to fall away. With every death, a few new buds sprout and keep up the show, together with the lower branches. The sprouts often go straight up and then curl over like water in a fountain. Sometimes there are many on a branch, sometimes few. The oak does not go quickly and quietly. It celebrates its own demise. This is why the poet John Dryden said of it, “three hundred years growing, three hundred years living, three hundred years dying.”
Persistence
 The ratio of roots to shoots in a plant is a standard measure among botanists and a rough way to express the staying power of a young tree. If a tree puts on a lot of top growth and few roots, it is liable to be weak wooded and short lived. Fast growers mature young and reproduce in haste. The birch, for example, dwarfs the oak in its capacity to produce seed, making orders of magnitude more seeds in a life that it is only a quarter as long. The early death of a few thousand individuals is of little consequence to such a tree. If a tree puts down a great deal of roots and adds shoots more slowly, however, it is liable to be long lived and more resistant to stress and strain.
The ratio of roots to shoots in a plant is a standard measure among botanists and a rough way to express the staying power of a young tree. If a tree puts on a lot of top growth and few roots, it is liable to be weak wooded and short lived. Fast growers mature young and reproduce in haste. The birch, for example, dwarfs the oak in its capacity to produce seed, making orders of magnitude more seeds in a life that it is only a quarter as long. The early death of a few thousand individuals is of little consequence to such a tree. If a tree puts down a great deal of roots and adds shoots more slowly, however, it is liable to be long lived and more resistant to stress and strain.
Few seedling trees have a root-to-shoot ratio that is greater than one, and in most trees it is less than one. This means that in most trees there is more shoot mass aboveground than there is root mass belowground. But the average ratio for a seedling oak is between four and six, and there have been oaks whose ratio was greater than ten. A seedling oak may have ten times as much root mass as it does stem and branch mass.
When the acorn first ruptures, surface dwellers often don’t even know about it. Likely, the lucky acorn is hidden beneath a layer of leaf and litter. The future trunk has not yet lifted above the surface. But the root is already heading down fast.
Oaks make lots and lots of roots. No matter what, they stand a chance of survival. The British botanist M. W. Shaw reported finding oak seedlings less than two feet tall that were twenty-five years old. Within the first year, the taproot of an oak can reach more than a foot in depth, thickening to about the width of a pencil and sending out its first lateral roots all along the stem. Though the taproot later declines in importance, it spends the first few years thickening and storing as much food as it can, for until the oak’s canopy is well above the reach of everything from turkey to deer, it is liable to be chewed down again and again.

The root going down (from H. Marshall Ward, The Oak)
Some root tips circumnutate, that is, they “swim around.” The meristem cells—that is, the growing tips—divide, then stretch, then divide again. The fact that they do this at varying rates according to their position around the circumference of the root accounts for the twisting motion. The root is like an auger, opening the way in the soil. Roots can travel pretty fast, up to twenty-five millimeters in a day. That is fast enough that were you patient (and able to see through soil), you could watch them grow.
Half the total food that the tree makes each year goes to grow new roots. One student laments this inefficiency and is at a loss to explain why oaks should be so profligate. I think that the oak is practicing, the same way a person plays the same succession of musical notes again and again to know them by heart. By putting out far more roots than are strictly needed, the oak is more likely to have enough to prosper.
Roots may spin, but they try to grow outward in straight lines. They branch frequently, but the growing tip of any one root keeps going as it began. The lateral roots of a large, open-grown oak stretch hundreds of yards from their origin. At first, they may be as thick as a man’s torso, but they quickly dwindle until six feet from the trunk they are only as thick as his wrist, at thirty feet they are as slender as a pencil lead, and at fifty feet as thin as grass, and at one hundred feet thinner than a human hair.
Roots do not simply swim. They sense and respond to irritants—rocks, water-saturated zones, poisons—as finely as the human tongue, and they are stronger and more intelligent than a human finger or arm. If a growing root encounters a stone, it may compress slightly, deflect to one side, and return to its straight line once the obstacle has been passed. Or, if it senses the slenderest crack in the rock, it may splay out thin and grow right through the fissure to the other side.
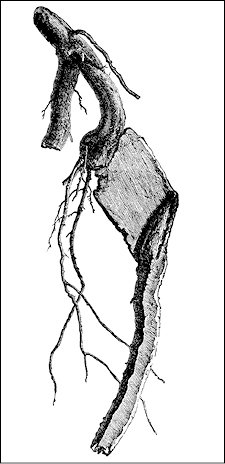
A root that squeezed itself flat to pass through the crack in a rock (from H. Marshall Ward, The Oak)
Roots know where they are. They know how deep they are in the soil, because they measure the amount of oxygen around them. Where there is more oxygen, there is more growth. This is why the cambium—the layer of cells just below the bark that is responsible for the thickening of stems—usually puts on more new tissue on the upper side of a root growing parallel to the ground than on its lower side. If a root has to twist out of the way of a stone, it has the means and knowledge to return from the deviation, since it will seek to reestablish its preferred oxygen environment.
It was once thought that the tree’s branches and its roots were mirrors of each other. If you took the tree out of the ground and held it by the middle of the trunk, you would have something that resembled a hairy dumbbell. But this is not so. Botanists found this out when some extremely patient Scandinavians painstakingly, with the care of great painters or archaeologists, scraped away the soil from the root systems of several mature trees.
Imagine their excitement and exasperation. They had thought they would run out of roots at about the same distance from the bole as they ran out of branches overhead. No, the roots kept going. Five feet, ten feet, twenty-five, fifty . . . As the roots got smaller and more numerous toward the tips, the scientists must have been giddy with fatigue, eyestrain, and the sense of discovering something very important while doing something virtually impossible and completely ridiculous. They gave up when the distance of the root mass from the trunk was nearly three times as wide as the maximum spread of the tree’s crown. And there were still more roots beyond.
The smaller the roots get, the shorter they live. Most smaller roots live four years or less, some for as little as a season or even a week. Many of the pencil-lead and thinner roots die and are replaced several times in one year. Root hairs—the almost microscopic hairs that work with beneficial fungi to bring nutrients into the tree—may be replaced every few hours. Yet the average mature red oak has better than five hundred million living root tips.
Root growth in oaks is doubly persistent. While the leaves sprout three or four times each year, the roots grow continuously from March to October, and in some climates into November. The biggest spike of growth occurs between May and late June in the temperate Northern Hemisphere, with a small spike between late August and early October.
It is not a good thing to risk damaging the roots, for it is these upon which the oak’s life depends. Leaves and branches are the beauty of the tree, and one tends to think that they must be the most important part. Not so. You can cut the oak back to the ground again and again, and it will resprout from the energy contained in the roots. But once you seriously damage the roots there is a good chance that the tree will die.
Unseen, a cloud of hundreds of millions of root tips swim through the soil. The mystery writer Wilkie Collins was not just waxing poetic when he wrote, “Fancy and imagination, grace and beauty, all those qualities which are to the work of art what scent and beauty are to the flower, can only grow towards heaven by taking root in the earth.”
Community
 Many individual oaks aren’t. The live oaks of central Texas, for example, for all that they form groves stretching over fifty square miles, are really each composed of a small number of individual oaks that have made many auxiliary trees by sprouting from their own roots. In other words, a grove that appears to comprise hundreds of trees may really only consist of four or five genetic individuals.
Many individual oaks aren’t. The live oaks of central Texas, for example, for all that they form groves stretching over fifty square miles, are really each composed of a small number of individual oaks that have made many auxiliary trees by sprouting from their own roots. In other words, a grove that appears to comprise hundreds of trees may really only consist of four or five genetic individuals.
But even where oaks are born as individuals, they often do not remain so. As an oak matures, its roots reach an area four to seven times as wide as the width of the tree’s crown, the diameter of their spread about twice as wide as the tree is tall. This means that in a mixed forest dominated by oak the soil is densely packed with winding roots. Roots of different species of oak, or of different genera, may wind tightly about each other and may hold their fans and mats of infinitesimal, mycorrhizally infected feeder roots cheek by jowl in the same parcels of decaying leaf litter. Nevertheless, they remain independent of each other. But within the same species of oak—particularly for species in the red oak group—when roots meet, they very often graft, that is, their vascular systems join and in essence, they become one flesh.
The old saw “You can’t see the forest for the trees” is truer than metaphor. Because we cannot see beneath the soil, we do not know that these oaks are not simply neighbors, providing mutual shade, protection from the wind, and sharing predator pressures, they are physically united beneath the ground.
In any forest, some trees are dominant. They reach the light first, spread fastest, occupy territory both above- and belowground. Other trees—foresters call them “suppressed”—have trouble reaching the canopy at all and their trunks may be only half the diameter of the dominant trees, though they are the same age. Still others may have started much later and so have little chance to reach the crown. Other species, too, may grow faster than young oaks and so deprive them of light.
In a world where nature is said always to be red in tooth and claw, one might well expect this to be the norm: If you can’t compete, then just die and get out of the way. The fact is, however, that the dominant oaks in an oak forest sometimes support the suppressed ones. Food travels through the grafted roots from the trees with extra food to those that do not have enough. The forest supports its weaker members. This turns out to be a good thing for the entity as a whole, since when the largest tree succumbs to a storm, to old age, or to axes and saws, the suppressed trees may be healthy enough to take over the dominant position.
The forest supports its sick trees too. Say an oak is suffering from a partial girdling, and nutrition cannot flow down from the photosynthesizing leaves to the base of the plant and to the roots. Instead of dying of starvation, the tree is fed by sugars that are imported from other, healthy oaks through the root grafts. The borrowed sugars ascend the stem in the healthy xylem and buy time so that the wounded tree can rebuild its own circulation.
In fact, the forest even supports its dead. Stumps in an oak forest are very slow to decay, and they will often sprout again. This is not simply because they are “vigorous” in themselves, but because their roots may be supplied with nutrients by the neighbor roots to which they are grafted. If you look closely at some forests of pin or red oak in America’s upper Midwest, you will see that the majority of the trees are stump sprouts, regenerated from a forest logged decades ago.
Even where a stump does not regenerate, its roots may go on serving the forest. The grafted roots may go on acquiring water and nutrients for the surviving trees long after their parent tree has rotted away. These “adopted roots” are common in oak forests.
All this mutual help, however, comes with one grave disadvantage. The roots are an open pipeline for food, but they might also be a conduit for disease to spread quickly and without barrier. If the forest were merely a community, there might be hope: The physically separated trees might be spared, just as the storytellers of Boccaccio’s Decameron hoped to be spared from the plague by locking themselves up in a castle and avoiding all contact with the diseased. But where the members are connected underground, there is no hope of isolation.
Generosity
 The list of guests at the great oak feast is longer than the Catalogue of Ships in the Iliad. Among the creatures that feed at the table are the great prominent, the oak hook tip, the thorn moth, the oak tree pug, the brindled and marbled pugs, the oak beauty, the great oak beauty, the green oak tortrix, the blotched emerald, the purple emperor, the wood worm, the oak bark beetle, the red oak roller, the nut weevil, the leafhopper, the buzzing and jumping spiders, the sawflies, the deathwatch beetle, the tent caterpillar, and the tussock moth, the oakworm and the oak twig pruner, the two-lined chestnut borer and the Asiatic oak weevil, the pigeon tremex, the periodical cicada, the greedy and golden-oak scales, the obscure scale, the oak leucanium, the lactarius, the boletes, the russellas, the armillarias and the ceratocistus, the truffles and the ganodermas, the mice and the voles, the shrews and the badger, the squirrels, the turkey, the wood pigeon, the fallow deer, the mule deer, the red deer, the white-tailed deer, the porcupine . . . There are many more. But none is so intimately related to the oak as the gall-making wasps, the cynipines.
The list of guests at the great oak feast is longer than the Catalogue of Ships in the Iliad. Among the creatures that feed at the table are the great prominent, the oak hook tip, the thorn moth, the oak tree pug, the brindled and marbled pugs, the oak beauty, the great oak beauty, the green oak tortrix, the blotched emerald, the purple emperor, the wood worm, the oak bark beetle, the red oak roller, the nut weevil, the leafhopper, the buzzing and jumping spiders, the sawflies, the deathwatch beetle, the tent caterpillar, and the tussock moth, the oakworm and the oak twig pruner, the two-lined chestnut borer and the Asiatic oak weevil, the pigeon tremex, the periodical cicada, the greedy and golden-oak scales, the obscure scale, the oak leucanium, the lactarius, the boletes, the russellas, the armillarias and the ceratocistus, the truffles and the ganodermas, the mice and the voles, the shrews and the badger, the squirrels, the turkey, the wood pigeon, the fallow deer, the mule deer, the red deer, the white-tailed deer, the porcupine . . . There are many more. But none is so intimately related to the oak as the gall-making wasps, the cynipines.
“The oak,” wrote Theophrastus in about 300 B.C. in his Enquiry into Plants, “bears more things besides its fruit than any other tree.” He noticed a dozen different galls: one a scarlet berry; another a “black resinous gall”; one like a phallus; another like a bull’s head; one soft, woolly, and spherical, which could be used as a fire starter; another a hairy ball that in spring sweated sweet juice. He recorded spot galls and hollow galls, button galls and horned galls, red, yellow, black, and transparent galls. Not only that, he said, the oak bore many fungi that grew from its roots. And he cited Hesiod as claiming that both honey and bees were also produced by the oak tree, though being perhaps a better observer than the poet, Theophrastus noted that Hesiod may have mistaken the honey-sweet droplets often found on oak leaves—the aphid drippings, as we know now—for the precursor and origin of honey.


A few oak galls (Nora H. Logan)
Theophrastus was as good an observer as humankind has ever produced. The favorite student of Aristotle, he succeeded his master as head of the Peripatetic School. He recounted the uses of the galls. One was used for tanning, another for a black dye, another for a red dye. Theophrastus properly observed that sometimes little flies would emerge from the galls—he was lucky to see them since they are only about an eighth of an inch long and frequent the galls only for a short time—but he did not draw the conclusion that these visitors were in fact most likely the galls’ architects.
Who would? It is clear that insects and their larvae may eat leaves or bore into wood, lay eggs under bark or inside leaves, or suck sap, but who would believe that an entire family of tiny wasps—almost four hundred species in all—could induce the oak to make elaborate homes for their larvae? The oaks and the cynipine wasps, often called gall wasps, have kept company like this for thirty million years.
Every gall wasp has two different lives. The impregnated female lays her eggs in some part of the freshest and most undifferentiated tissue of an oak tree, be it in the twigs, the buds, the leaves, the tiny male or female oak flowers, or even on the rootlets. The newly hatched larvae secrete something—no one is sure just what—that causes that oak to provide it with its distinctive home, the gall. These larvae grow into adult self-fertile females, who promptly go lay their own eggs in a different part of the oak. This generation of larvae makes its own distinct gall. In short, every cynipine wasp makes two different galls.
The oak derives no benefit from these galls, but neither is it usually harmed by them. Occasionally, an aggressive and numerous maker of twig galls—like the wasps that make the so-called horned and gouty oak galls—create so many galls that branches are girdled and die. (Such oaks appear to have sprouted black golf balls.) But this is the exception and only occurs to oaks under stress for other reasons. Mainly, the oak acts as the bricks and mortar that are deployed according to the larvae’s plans.
The cynipine galls are extraordinary, many-layered and labyrinthine. They exist not simply to feed growing larvae but also to defend it against the weather and especially against the numerous parasitoids that would either eat the larvae or take over the house for their own broods. Every species of cynipine has up to twenty other species of arthropods and countless species of fungi that, given the chance, would devour its larvae and move into the vacated space.
A gall is a castle. Usually, the inner layer is a rich sphere dripping with food that the larvae graze upon with their chewing mouthparts. It may contain one or numerous larvae, depending upon the species. Surrounding this sphere is usually a hardened layer, resistant to the piercing mouthparts or egg-laying tubes of parasitoid mothers. Outside this is a spongy layer very rich in tannins and in other acids that the gall’s tissue somehow concentrates to as much as ten times their level in normal oak tissue.
Still, the contest between larvae and would-be predators is a serious one. Some of the invaders can even digest tannins, and in fact hollow out a little space in the tannic layer where their own young are born and grow, sharing quarters with the gall wasp larvae. The cynipines are not new to this situation. They have had thirty million years to prepare and elaborate further defenses in the outer layers of their gall homes. One solution is to make the shape a puzzle: The pink to purple gall of one wasp looks rather like a sea urchin, but the larval chamber is at the base of only one of the many spikes. The warty and horny twig galls of another are a labyrinth of larval chambers, only a few of which contain the living treasure.
A second solution is to outrun your enemy’s firepower. The chalcid wasp’s weapon against the cynipine is the ovipositor: a penetrating lance that holds on its tip an egg that is deposited at the uttermost limit of its extension. The cynipines, in response, engineer a mature gall just a little bit thicker than the ovipositor is long, so the frustrated chalcid is left with its egg just shy of the juicy larval chamber. Many galls, upon examination, turn out to have ovipositors broken off in them. The predator’s only chance is to attack when the gall is still young and the diameter of the protective tissue is less.
A third solution is to cover the gall’s surface with something unappealing to attackers. Some galls are quite sticky and threaten to retain the would-be mother like a fly on flypaper. Others are hairy and difficult to alight upon at all, or spiny and hard.
But perhaps the most novel and effective idea is to hire protection. Ants will furiously attack anything that threatens their herds. Ants, as is well known, farm aphids, scales, and other invertebrates, milking these creatures for the sweet honeydew that they draw from the plant’s sap. Certain galls themselves have developed a means of synthesizing and secreting a similar honeydew, so that ants gather around the galls, feed upon the honeydew, and fiercely defend the galls.
The average oak, then, is like a country dotted with gall towns. Each town has its weather, its streets and walls, its inhabitants, its attackers, and a life span. Once the cynipines move out—leaving holes in the walls of their homes—fungi immediately enter and begin to digest the walls. I have a collection of old galls—big hollow balls, horned tunnels, red spots, odd clown’s hats—and each of them reminds me of ruins. To feel one and turn it over in my hand is like visiting an abandoned village.