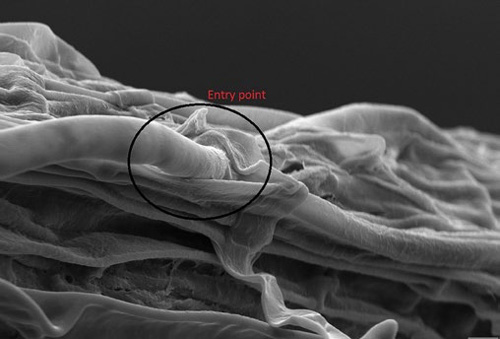
The soils of this hardwood forest are teeming with mycorrhizae.
NEARLY EVERY TREE species forms some type of mycorrhizal association. Some trees form arbuscular mycorrhizae, but others have evolved over time and are hosts to ectomycorrhizal fungi. Some trees associate with both forms of mycorrhizae, though usually at different periods of their lives. Members of the pine family (Pinaceae), for example, are mostly ectomycorrhizal hosts, but they can also be colonized by arbuscular fungi during the early phases of establishment, especially in disturbed sites where arbuscular fungal propagules are more abundant than ectomycorrhizal fungi.
Although arbuscular mycorrhizal fungi partner with many trees, and they are included in this discussion, when appropriate, the emphasis here is on ectomycorrhizal fungi, mostly members of phyla Basidiomycota and Ascomycota, with a few from Zygomycota. About 6000 species of ectomycorrhizal fungi associate with about 10 percent of plant families, including Pinaceae (pines and other conifers), Cupressaceae (cedars and cypress), Fagaceae (beeches and oaks), Betulaceae (birches and alders), Salicaceae (poplars and willows), Dipterocarpaceae (lowland rainforest trees), and Myrtaceae (myrtle, clove, guava, and eucalyptus). The plants and trees they support are large in size, grow in groups, and cover huge portions of the Earth.
The number and density of the mycorrhizal colonies formed is staggering, as many trees and shrubs associate with more than one species of mycorrhizal fungi and often more than one type. In one Swedish forest, for example, biologists discovered that 95 percent of all the root tips had mycorrhizal associations and as many as 1.2 million ectomycorrhizae were found in a mere 1 square meter of soil.
The benefits of ectomycorrhizae to trees and shrubs are, for the most part, the same as those imparted by endomycorrhizae formed by arbuscular mycorrhizal fungi and roots (some of which colonize tree and shrub roots). A quick review of what they do, however, doesn’t hurt. They increase root formation and mass, nutrient uptake, water uptake and storage, and stress and drought tolerance; they decrease nutrient runoff, moderate pH, and improve pathogen resistance.
Mycorrhizal fungi and plant roots access the same nutrients in the soil and use the same mechanisms to take up nutrients, but the fungi can also access minerals by growing into rock and dissolving them. This enables the fungi to capture mineral nutrients for the host plant that the plant would never get without the mycorrhizal association.
Mycorrhizal fungi also protect plants from damage by heavy metals. The fungi take up and retain arsenic, copper, zinc, iron, lead, cadmium, nickel, mercury, chromium, and aluminum, preventing their accumulation in the soil and consequent damage to the host plant and others. Some fungal tissues can even filter radioactive materials such as cesium.
A mycorrhizal fungus normally uses about 15 percent of the plant’s photosynthetic products. A 15-percent loss of energy from photosynthesis is a lot—imagine losing 15 percent of your energy and having to make energy to do it! In return, however, the plant can get 85 percent of its nitrogen through mycorrhizae. The transfer of carbon to the fungus and the cross-transfer of fungi-obtained nutrients to the plant host occur in the intercellular (apoplastic) space between the hypha and the plant cell wall. Molecules flow through the walls for transport across the respective organism’s membranes.
Monterey pine (Pinus radiata) seedlings germinate after a fire and provide host plants for mycorrhizae.
In managing a forest, a silviculturist focuses on the biggest trees, not only because they will be harvested for lumber, but also because they protect the health of the understory, provide shade, supply organic debris, and more. These large mother trees are colonized by mycorrhizal fungi, and the resulting mycorrhizae support all manner of other organisms, including other plants. They share their mycelial network with seedlings and young trees in the undercanopy that are not able to compete for sunlight. They also share the benefits of their sunlight-gathering capacity with their progeny via the underground extracellular network.
When big trees die in a forest, much of their nutrients are returned to the fungal network for reuse. This reuse also includes the decaying leaves that fall year after year, as evidenced by the mycorrhizal fungi, vesicles and all, which are found inside dead leaves, especially those close to colonized root systems.
Unlike arbuscular fungal spores, ectomycorrhizal fungal spores can survive forest fires by forming spore banks that reside in the soil until the spores are needed. Many shrub species rapidly sprout after a fire and keep mycorrhizal populations fed until regenerating trees can plug into the mycelial network. This enables mycorrhizal regeneration to occur immediately after a fire instead of having to wait for spores to blow in from the next forest over.
The formation of ectomycorrhizae involves a complicated but beautiful arrangement between fungus and plant. From 25 to 30 chemicals are produced by both participants, most of which will never be used again for any other purpose, which emphasizes the importance of this process.
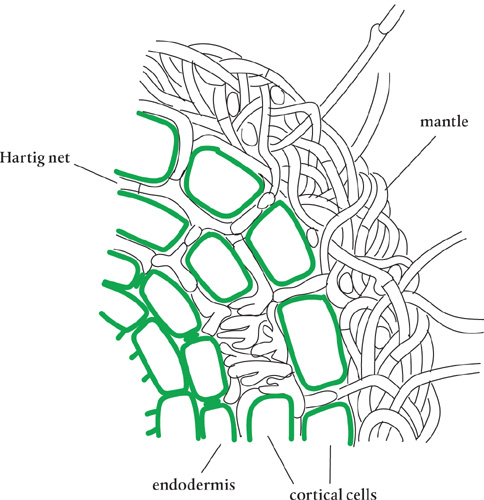
At the beginning of the process, the host plant roots send out signals via strigolactone hormones to encourage the mycorrhizal fungi in the area to extend their hyphal reach and seek out a partnership. When the growing ectomycorrhizal fungal hyphae reach the roots, they do not penetrate the cell wall as endomycorrhizal fungi do. Instead, they weave together to form a mantle around the root, and then they use cellulolytic enzymes to dissolve the middle lamellae (layer) of both epidermal and outer cortical cells. As a result, the hyphae surround the cell walls, forming the Hartig net. The plant cells are not penetrated, and the integrity of the protoplasm of each organism is kept totally intact. Within the Hartig net’s massive surface area, the exchange of nutrients with the symbiont occurs.
In the mantle, the hyphal sheath that surrounds plant roots, carbohydrates from the host plant are stored and retrieved. The mantle formation slows or stops the development of root hair cells and forces the plant to rely more on the fungi to obtain its nutrients. Mantles can be up to 40 micrometers thick—so thick, in fact, that they completely cover the roots. After the mantle is created, the host plant’s roots become short and thick, and their growth slows down.
Stained tissues on both sides of the vesicles show the Hartig net created by the ectomycorrhizal fungus.
Meanwhile, extraradical hyphae begin to develop in the soil. As with endomycorrhizal fungi, the ectomycorrhizal fungi excrete digestive enzymes and acids to help break down nutrients in the soil, which are transported through tunnels and channels in the plasma membrane. Cytoplasmic streaming in the fungal cell keeps things moving in the right direction.
Unlike arbuscular mycorrhizal fungi, most ectomycorrhizal fungi reproduce sexually, via large sporocarps, or fruiting bodies: mushrooms, puffballs, and truffles (though not all mushrooms and puffballs are ectomycorrhizal fungal fruits). Reproduction involves the transfer of genetic materials, development of spores, and then spore dispersal.
Many fruiting bodies of ectomycorrhizal fungi are safe to eat and have commercial value. Many are poisonous, however, such as the Alice-in-Wonderland toadstool Amanita muscaria (fly agaric), which associates with birch trees where I live in Alaska. Whether edible or not, all ectomycorrhizal fungal fruits produce small spores of usually less than 10 micrometers, which are dispersed primarily by wind. However, small mammals, birds, insects, slugs, worms, bacteria, and even other fungi eat spores and mushrooms or otherwise transport them. These ectomycorrhizal spores are more easily dispersed than the much larger and heavier endomycorrhizal fungal spores.
Some ectomycorrhizal tree hosts show specificity for particular fungal associations. The extent of this host specificity varies not only in the forest but in the lab as well. A set of chemical cues inform the plant that initiates the formation of mycorrhizae. Sometimes this is the only way that fungal spores will germinate. This protective measure has evolved to prevent other organisms from affecting germination, which would be to the detriment of both the fungi and plants, neither of which can survive without the other.
Some fungi are specialists and associate with only one host species or a limited number of plant hosts, while others are generalists and can associate with a wide range of hosts. In monocolonizations, one fungus invades one root. But an individual mycorrhizal fungus can be simultaneously associated with not only more than one host plant but with more than one species of plant. The host plant is also free to form associations with other organisms. A mature tree can be associated with ten or more different fungal species forming mycorrhizae. Things can get pretty complicated.
Modern techniques using DNA and RNA for identification enable scientists to determine the best host plant associations for particular mycorrhizae. A problem, however, is that some associations that occur in the lab do not occur in the wild, and some field-observed associations cannot be replicated in a lab setting. This makes matching tree to fungi very tricky.
Trees and shrubs without the proper mycorrhizal associations struggle and usually die because they cannot obtain enough nutrients and/or water. (A friend, for example, once planted 10,000 trees in Alaska without a mycorrhizal fungal inoculation. They all died.) A well-known example of the importance of these associations occurred in Puerto Rico in the 1950s, when attempts to grow pine trees using native island soils failed. The seedlings turned yellow because of lack of nutrients, even if extra fertilizers were added. Within two years after being planted, every tree died. In 1955, soils taken from around some pines in North Carolina were used to inoculate the roots of the pines on the island. These trees thrived because of the association with the appropriate ectomycorrhizal fungi.
Specific strains of ectomycorrhizal fungi, tested with specific hosts, are commercially available. Not only do these fungi help the plants grow in the nursery, the plants are healthier after being transplanted in the forest or field.
Good general-purpose ectomycorrhizal fungi for tree nurseries include Thelephora terrestris, Laccaria laccata, and species of Inocybe. These are found in natural forests and can be used to inoculate outdoor nursery-grown stock. Ectomycorrhizal Pisolithus arhizus and Scleroderma citrinum have been used for oak inoculation, and the latter is the best fungus for European beech. Hebeloma arenosum has been used on pine seedlings, and Leccinum scabrum is a good inoculant for birch seedlings. Arbuscular mycorrhizal Rhizophagus intraradices is used to inoculate seedlings of red cedar, redwood, and giant sequoia.
Silviculturists and arboriculturists can use mycorrhizal fungi to speed the growth process when reestablishing trees in disturbed areas, such as chemical spill sites, borrow pits, and heavily eroded terrain. Studies have shown that mycorrhizae also improve both growth and survival of seedlings in disturbed soils and during times of drought.
Mycorrhizae can take up and isolate toxins—including heavy metals and radioactive elements. According to research, some mycorrhizal fungi bind and isolate radioactive elements, which can remain immobilized within the fungal tissue for years.
Consider several studies involving mushrooms related to the 1986 Chernobyl nuclear disaster. In 2002, a robot sent inside the destroyed nuclear power plant took samples of fungi growing on the walls. Researchers determined that these fungi used radioactivity as an energy source to break down food and increase growth. Studies have also shown that native deciduous trees and conifers can be inoculated with mycorrhizal fungi such as Gomphidius glutinosus, Craterellus tubaeformis, and Laccaria amethystina, all of which absorb radioactive cesium.
Other studies demonstrate the importance of mycorrhizal inoculation in reseeding and reclaiming forested areas damaged by mining operations. The goal is to return the mine site to natural conditions, in part by inoculating the soils with mycorrhizal fungi.
Reforestation efforts usually require planting commercially grown seedlings or shrubs. Commercial nurseries sometimes use sterile growing media and fumigate to destroy pathogens. These conditions, however, can destroy or slow the growth of existing mycorrhizae. Although such practices may outweigh the cost benefits of fostering mycorrhizae while the plants are in the nursery, reforestation planting always benefits from the addition of mycorrhizal fungi propagules. Although ectomycorrhizal spores are light enough to be distributed by the wind from an area where the fungi are growing, the spores of arbuscular fungi are heavier and must be distributed via water, animals, or inoculation. To ensure that sufficient numbers of spores are available to the growing trees, inoculation is required.
Workers replant red spruce (Picea rubens) in North Carolina with seedlings inoculated with ectomycorrhizal propagules.
In nurseries, inoculation of tree stock has met with mixed results—usually good, but sometimes there has been no visible difference between inoculated and uninoculated plants. It is clear, however, that when the growing conditions are not conducive to mycorrhizal health, such as when overfertilization is the rule, mycorrhizal fungi suffer.
The appropriate mycorrhizal association should be established as early as possible on any tree or shrub and maintained for the life of the plant. Because the appropriate fungi need to be used as inoculants, using a broad mix of mycorrhizal fungi can help ensure colonization; each fungus in the mix imparts its own particular benefit to a host plant.
Many commercial planting soils contain propagules, and liquid formulations can be applied via drip or other watering systems. Seedling roots can be dipped in granular or liquid formulations at the time of transplanting. Granular formulas can be placed in holes in the soil around drip lines, and liquid inoculants can be applied at any time to existing stock.
Growers must carefully monitor the use of fertilizers and chemicals when plants are inoculated with mycorrhizal fungi. Overfertilization, particularly with phosphorus and nitrogen, can lead to a reduction of mycorrhizal associations or even death of the fungi. Because pesticide and fungicide applications can also be detrimental to mycorrhizae, growers must apply only compatible chemicals at the appropriate times.
The amount of organic matter in the forest soil can also impact how mycorrhizae fare. In one study, mycorrhizal formation was reduced by up to 90 percent when Douglas fir seedlings were grown in soils where timber operations greatly reduced the organic material. Slash and debris should remain on site after harvest to help maintain mycorrhizal populations.
Thousands of studies demonstrate the efficacy of inoculating with ectomycorrhizal and, when appropriate, arbuscular mycorrhizal fungi when seeding, rooting, and transplanting shrub and tree seedlings. Here are a few summaries of the results of studies on trees and shrubs.
Almond (Prunus spp.) and walnut (Juglans spp.) One study showed that regardless of the planting medium—peat, sandy soil, or bark compost—after two years, inoculated almond and walnut seedlings showed the same amount of mycorrhizal colonization, and all inoculated seedlings grew larger than uninoculated plants. A study of the rootstocks of several almond cultivars indicated that inoculation with Rhizophagus intraradices resulted in larger plants and root masses. Inoculation with R. intraradices, Funneliformis mosseae, and Claroideoglomus etunicatum also suppressed root-knot nematodes.
Apple (Malus spp.) Five months after Rhizophagus intraradices was used to inoculate apple seedlings, the plants showed an increase in phosphorus, stem length, number of nitrogen-fixing nodules, and dry weight. When Diversispora versiformis and Glomus macrocarpum were used as inoculants, the host trees’ ability to withstand drought stress was improved. Inoculated trees grew more leaves and increased their uptake of zinc, copper, and other minerals. In addition, inoculated plants fared as well after application of low levels of phosphorus as control plants fared with higher amounts. Results were similar after inoculations of Claroideoglomus etunicatum and Gigaspora margarita.
Black walnut (Juglans nigra) Black walnut trees that formed arbuscular mycorrhizae with Glomus deserticola and Claroideoglomus etunicatum showed high levels of root colonization. Studies showed a strong relationship between where the seeds were harvested and which Glomus species was the best host. This might explain why walnuts inoculated with Rhizophagus intraradices produced taller seedlings than trees inoculated with C. etunicatum. These studies also demonstrate the importance of using the appropriate fungal symbiont for each seed source.
In addition, Funneliformis mosseae in soil was an effective control for stunt disease. In another study, plants inoculated with F. mosseae showed resistance to nematodes. Inoculated trees also showed higher amounts of several nutrients than control plants.
Chokecherry (Prunus virginiana) Chokecherry saplings responded to Funneliformis mosseae and Rhizophagus intraradices with greater mass above and below ground than control plants.
Douglas fir (Pseudotsuga menziesii) Inoculation with Laccaria proxima greatly improved growth of pot-grown Douglas fir. Inoculation with Hebeloma longicaudum, Paxillus involutus, and Pisolithus arhizus increased growth of seedlings compared to controls with increasing application of fertilizer. This suggests that if mycorrhizae are established, less fertilizer may be required.
Dwarf willow (Salix reinii) Dwarf willow shrubs are colonized by several species of ectomycorrhizal fungi. Improved growth and nitrogen content in seedlings grown near willows showed that the ectomycorrhizal fungi from established willows are critical to the succession of others that grow in the area.
European beech (Fagus sylvatica) When colonized by Lactarius subdulcis and Cenococcum geophilum, beech tree roots better withstood the impacts of drought than control plants.
Incense cedar (Calocedrus decurrens) Inoculation with Rhizophagus intraradices significantly increased the survival rates of incense cedar on a disturbed site.
Oak (Quercus spp.) Inoculation with ectomycorrhizal Pisolithus arhizus improved seedling growth, shoot diameter, and height, and inoculation with naturally occurring mycorrhizal species showed the same effect. In one study, forest soils and agricultural soils were used as media in which to grow oaks. The forest soils showed the growth of two more mycorrhizal species when compared to the agricultural soils, with better overall root and shoot growth and survival rates.
Palms (Arecaceae) Inoculations with Funneliformis mosseae, Rhizophagus intraradices, and Glomus deserticola were effective in increasing plant growth. Funneliformis mosseae–inoculated plants were hardier than uninoculated fertilized plants, suggesting that inoculating plants can result in less fertilizer use with better growth. In addition, inoculated nursery plants showed a better survival rate when transplanted to the field.
Pear (Pyrus spp.) Pear trees inoculated with Rhizophagus intraradices and Glomus deserticola showed increased growth, but the phosphorus levels and the species affected the outcome, suggesting the importance of experimenting with inoculants to find the most effective mixture.
Pine (Pinus spp.) Inoculation with Laccaria proxima greatly improved growth of pot-grown Japanese black pine (Pinus thunbergii), jack pine (P. banksiana), and mugo pine (P. mugo). Inoculation with Hebeloma longicaudum, Paxillus involutus, and Pisolithus arhizus increased growth of seedlings compared to controls with increasing application of fertilizer. When competing with Sitka spruce (Picea sitchensis) in the field, inoculated pine trees were healthier after 11 years of growth. Pines with mycorrhizae also contained more nitrogen. In experiments to determine whether mycorrhizal fungi would speed and improve rooting, biologists found no significant difference between controls and the inoculated cuttings; however, when the cuttings were potted up, they showed 50 percent better growth the first year. Ponderosa pine (Pinus ponderosa) seedlings inoculated with Rhizopogon roseolus showed significantly higher survival rates when planted in the field, with a 93 percent survival rate versus 37 percent for uninoculated seedlings. Red pine (P. resinosa) seedlings inoculated with H. arenosum while still in the nursery showed greater survival rates when planted in the field.
Spruce (Picea spp.) Inoculation with Laccaria proxima improved growth of black spruce (Picea mariana), red spruce (P. rubens), and especially pot-grown white spruce (P. glauca). Inoculation with Hebeloma longicaudum, Paxillus involutus, and Pisolithus arhizus increased growth of seedlings compared to controls with increasing application of fertilizer.
Yew (Taxus spp.) When Rhizophagus intraradices was used to inoculate yew cuttings, root production was significantly higher than production in the control group, with results as good as or better than using a rooting hormone.