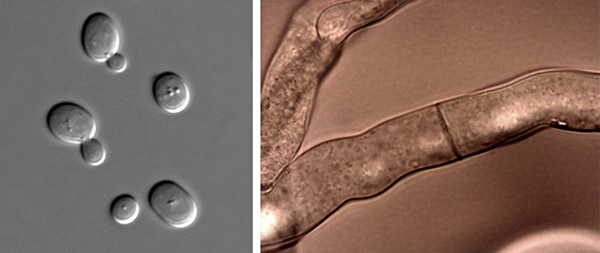
The two forms of fungi: unicellular yeast (left) and multicellular hyphae (right).
MOST OF THE more than 100,000 species of known fungi live on land, but they are truly ubiquitous organisms, with some species capable of living in any ecosystem and clime—from deserts, to tundra, to polar caps, to oceans and other saline environments. Yeasts, for example, which are single-celled fungi, have been found on both of the Earth’s polar ice caps. Lichens, which are formed by fungi and algae, grow throughout the Arctic as well as the temperate regions of the Earth. Fungi survive the extreme radiation of space. They exist throughout and cover your body (especially your feet). Some fungi appear to be specific to particular environments, such as the wood fungi that feast on the Antarctic huts built in the early 1900s by explorers Robert Scott and Ernest Shackleton, or the fungal mold that attacks the nopal cactus (Opuntia spp.) in the Mexican highlands.
Fungi play essential roles as decayers and nutrient recyclers (saprobes), parasites or pathogens, and symbiotic organisms (symbionts). Pathogenic fungi cause devastating crop losses and ravaging diseases in animals, ranging from ringworm to histoplasmosis. Fungi can cause death in humans, other animals, and plants. Many fungi can be beneficial, however, serving as biopesticides or contributing to the flavor of delicious foods such as cheese and all manner of intoxicating beverages. Other fungi have proven to be extremely important to human health: consider Tolypocladium inflatum, a fungus originally found in Norwegian soils, which produces cyclosporin, an immunosuppressant used in organ transplants to prevent rejection.
Fungi are fascinating organisms that sometimes resemble plants, sometimes resemble animals, and sometimes are uniquely fungal. Technically defined, fungi are eukaryotic organisms—that is, they consist of cells that contain membrane-bound nuclei, as do plants and animals. Scientists believe that fungi, plants, and animals once shared common ancestors, and all are grouped into the same kingdom: Eukaryota. Although they are related, these three types of organisms have enough distinct differences that each is placed in its own domain.
Not so long ago, in the 1950s, fungi were classified as plants. It is easy to understand why. Like plants, fungi have cell walls and many of the organelles present in plant cells: membrane-bound nuclei and vacuoles, ribosomes, mitochondria, and many others. In addition, fungi often live in soil, they are relatively immobile, and their morphology (structure) resembles that of plant roots and branches.
Unlike plants, however, fungi lack chlorophyll. They are heterotrophic, which means they are incapable of converting carbons into sugar to produce their own energy. They also lack the vascular systems found in animals and plants and can reproduce asexually through spores. Fungi cell walls are not full of cellulose like plant cell walls, and fungal walls contain the polysaccharide chitin, a main constituent in the exoskeletons of arthropods such as insects, lobsters, and crabs. In addition, fungi use glycogen, another polysaccharide, to store energy. This energy storage molecule is absent in plants, which store energy in starch molecules instead. Fungi also, at least initially, digest their food extracellularly—outside of their cells—which also differs from plants.
Fungi also share some common characteristics with animals. Both are heterotrophic, and both store their food as glycogen. Animals and fungi use enzymes to digest their food (as do plants, but not in the same way), though in animals, digestion occurs after ingestion. A fungus uses powerful enzymes to digest food outside of the organism before adsorption.
Of course, there are distinct differences between fungi and animals. Chief among them, animal membranes contain cholesterol, while fungal membranes contain a fungal-unique molecule, ergosterol, a steroid alcohol that converts to vitamin D when exposed to the sun’s ultraviolet light. (This is why it is a good idea to expose edible mushrooms to the sun for a day or so before you eat them. And, by the way, a raw mushroom’s chitin- and ergosterol-filled cell walls are nearly indigestible. Cooking mushrooms can not only avert a stomachache, but it helps release the nutrients locked inside their cells.) This alone is enough to separate the two organisms into separate domains.
Fungi can be complex multicellular or simple unicellular (single cell) organisms. Mycorrhizal fungi are multicellular, comprising masses of branching filaments, or hyphae (hypha, singular). Hyphal fungi thrive in myriad environments, including soil if oxygen is available; they are aerobic organisms and need oxygen to survive. Hyphal fungi can be lurking on spoiled food in a container tucked away in the refrigerator. They grow in some cheeses such as brie and Roquefort. And they are the mildew that crops up on bread.
Yeasts are single-cell fungi—the type found in packets used to make bread and beer. Yeasts can survive in anaerobic (oxygen-free) conditions such as pockets in compacted soil; however, none of these types of unicellular fungi form mycorrhizae, so they are not important in this discussion and I will ignore them here. Instead, let’s concentrate on the hyphal fungi, since all mycorrhizal fungi are of this type.
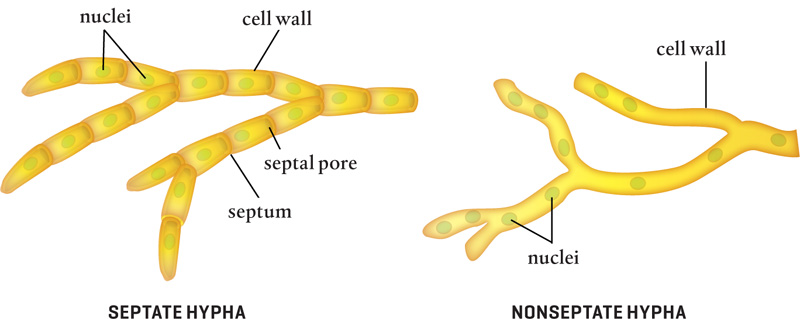
Each tiny mycorrhizal hyphal strand comprises connected cells, each filled with cytoplasm, organelles, and one or more nuclei. Tubular cell walls and septa (septum, singular) surround each cell and serve as structural supports. Septa are perforated by septal pores that allow cytoplasm to flow throughout a hypha’s cellular compartments, transporting minerals, enzymes, and other intracellular materials between cells.
Most hyphal tubes have diameters of only 2 to 10 micrometers, though they can grow larger in some fungi. For comparison purposes, the diameter of a human hair is about 100 micrometers (1.0 micrometer equals 0.001 millimeter). Fungal hyphae can grow from a few centimeters to several meters in length. They are so thin that it takes hundreds of thousands of individual hyphal strands to form a network thick enough for the human eye to see. In fact, a single teaspoon of good garden soil may contain several meters of fungal hyphae that are invisible to the human eye.
Some fungi are not divided by septa. In nonseptate (also known as aseptate or coenocytic) hyphae, one big cell contains many nuclei. These hyphae do, however, usually form a type of septum at branching points and to wall off damaged areas.
Most of the organelles inside a fungal hypha cell are also found in plants and animals and they function in much the same ways. Chloroplasts, which are used for photosynthesis in plants, are not present in fungal cells, however.
Hyphae are shaped like little trees, with branches that extend laterally. Septate hyphae are divided into compartments by septa.
Like plant cell walls, the outer cell walls of fungal hyphae protect the cells from changes in pressure and shield them from environmental stresses; they also contain some cellulose, a polysaccharide chain consisting of glucose, which makes up most of a plant’s cell walls. Unlike plant cell walls, however, the rigid hyphal cell walls also contain chitin, a long-chain molecule (polymer). Chitin helps strengthen the cell walls and offers flexibility. (That is why you need special tools to crack open lobsters and crab legs—their chitin-rich shells are hard, but they also bend.)
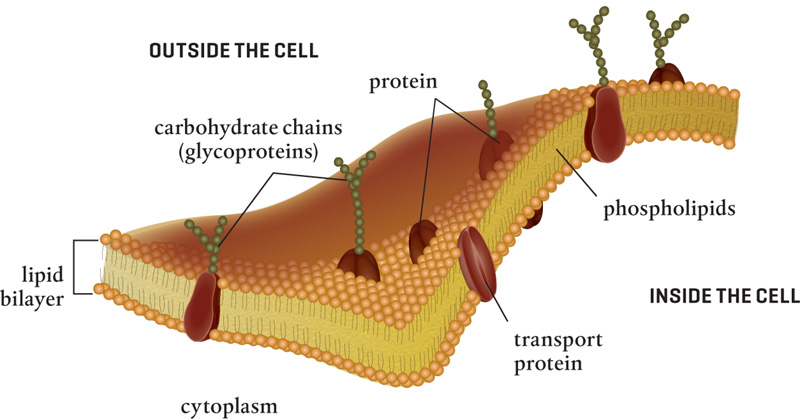
Chitin is produced by enzymes in the plasma membrane layer just inside the cell wall and intertwined with fiberlike strands (microfibrils) set in a base of glycoproteins. Almost 20 percent of the wall can comprise these glycoproteins, carbohydrate chains that play structural roles and are important for nutrient uptake. While some glycoproteins have structural roles, others are used in signaling, sensing, and recognition. Still others make up the tunnels and channels located in the membrane and used for nutrient uptake. Fungal cell walls also contain ions of phosphorus, calcium, and magnesium, important nutrients that can enter the cell and travel to the growing tip of the hypha via microfilaments and microtubules.
Up to 80 percent of a fungal cell wall consists of polysaccharides, the long chains of sugars; fibrils made up of chitin and chitosan; and glucans. These are all mixed in a gel that forms a matrix. In addition, the pigment melanin, which protects the cell from damaging ultraviolet rays, is produced in the cell wall. The exact composition of the wall depends on the specific fungus as well as the time of life of the particular hypha.
As in plants and animals, the plasma membrane layer of a hyphal cell is nestled against the inner cell wall and envelops the cytoplasm and organelles. The plasma membrane is the molecular gatekeeper for entry into the cell’s cytoplasm. This layer is so thin it would take 5000 to 8000 layers to equal the width of a single page of paper used in this book!
Some fungi in the phylum Glomeromycota—the arbuscular mycorrhizal fungi so important in agriculture and horticulture—produce glomalin, an essential glycoprotein that helps seal gaps and provides structural integrity to fungal cell walls. Sarah Wright, a soil research scientist for the U.S. Department of Agriculture (USDA), discovered glomalin in 1996. Before then, scientists couldn’t identify this recalcitrant and long-lasting part of the soil.
It turns out that the mysterious soil ingredient comes from arbuscular mycorrhizal fungi. This large molecule includes not one, but two carbon sites (along with lots of nitrogen). Glomalin is an extremely stable molecule with great longevity in the soil. Its molecular structure is sticky and contributes to soil structure. As arbuscular fungi grow and die, they are continually adding glomalin to soils. In fact, glomalin contributes 27 to 30 percent of the carbon in soil where mycorrhizal fungi are present.
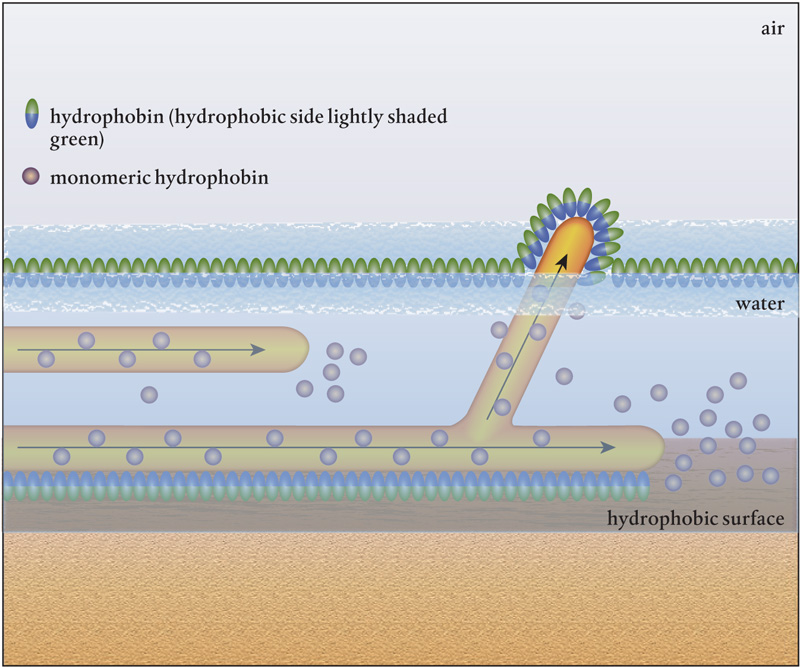
Hydrophobins are often left off the list of differences between plants, animals, and fungi. These cysteine-rich proteins are present only in fungi and are always embedded in fungal cell walls. They form a coating on the cell’s surface, which reduces water movement through the cell wall. Hydrophobins have both hydrophobic (or water repelling) sides and hydrophilic (or water attracting) sides. The hydrophilic side attaches to the fungal wall and keeps the cell from drying out. The hydrophobic side prevents water from rushing into the cell, which could dilute the cytoplasm and damage or drown the fungal hypha. The hydrophobic surfaces of these proteins also allow fungi to attach to other hyphae, bacteria, soil particles, plants, or fungal spores. These proteins are critical to the survival of hyphal fungi.
Monomers, the individual hydrophobin protein molecules, are assembled in the fungus and released from the growing tip of the hypha, where they stick to the outside of the cell wall. There they join together and form a solution that ensures that whenever a fungal hypha is out of the water, it is coated with molecules that keep it from drying out.
A fungus can contain ten or more different kinds of hydrophobins. Each has a specialized attribute that enables the fungus to function in diverse environments. Some help the fungal cell communicate inter- and intracellularly. Some control the flow to membranes, while others strengthen the hypha. These are complex molecules, and their full functions are under intense study because of their potential medical applications. Undoubtedly, there are agricultural and horticultural applications as well.
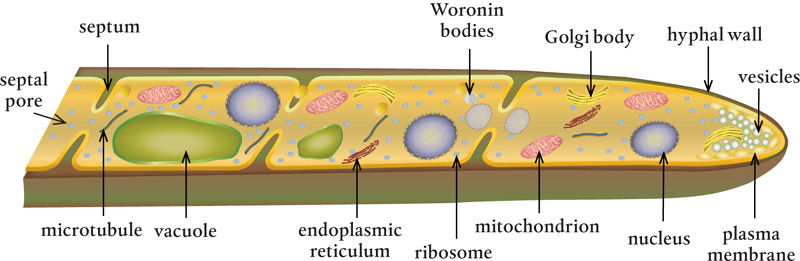
Septa divide a hypha into compartments. They can take several different structural forms, depending on the type of fungi. Some septa, particularly those in cells of Ascomycota fungi, have one central pore of 50 to 500 nanometers in diameter, which is large enough for organelles to move through as cytoplasm flows. Basidiomycota fungi have special structures that prevent nuclei from moving through the septa but allow passage of cytoplasm and small organelles. Other fungi have sievelike septa that are riddled with pores—each septum can contain as many as 50 tiny pores, each 9 or 10 nanometers in diameter.
Septal pores located between the septa are often accompanied by Woronin bodies, special membrane-bound organelles that can fill up septal pores when a hypha is damaged and leaking cytoplasm, too old, or full of vacuoles with unwanted toxins. If Woronin bodies are absent, the cells can produce protein crystals that clog septal pores to seal off a section of a hypha when it is damaged or under attack.
All hyphal fungi, whether septate or nonseptate, are homokaryotic—that is, all the nuclei in the hyphal cells are genetically identical and share the same cytoplasm. One of the biggest differences between nonseptate and septate hyphae is that nutrients flow much quicker through nonseptate hyphae so nutrient uptake can occur faster than it does in septate hyphae. As a result, nonseptate hyphae can outcompete septate hyphae.
The plasma membrane nestled inside the fungal cell wall is similar in structure, function, and composition to the plasmalemma that sits against the inside of a plant or animal cell wall. It comprises a double layer of lipids, in which individual phospholipid molecules with hydrophilic heads and hydrophobic tails move freely (like the film on a soap bubble). In addition to phospholipid molecules, there are concentrations of sterols and sphingolipids (waxes and fats) that pack and move together. Although the basic composition of the fungal plasma membrane can vary, one thing is constant: ergosterol is always found in fungal cell membranes and cholesterol is found in animal cell membranes.
The plasma membrane is embedded with signaling molecules and with proteins that act as carriers for nutrient molecules, selectively allowing molecules to move in or out of the cell. In fungal cells, embedded proteins include chitin synthase and glucan synthase, enzymes that act as pumps and carriers to move water, nutrients, and debris across the membrane actively or via diffusion or osmosis, just as they do in plants and animals—again suggesting the similar ancestral origin between plants and fungi. There are differences in the proteins embedded in various fungi membranes, probably because of the various metabolic processes that require different materials.
Vacuoles are intracellular storage bubbles filled with water and powerful enzymes that help destroy and build molecules. They are bound by double-layered membranes that separate their contents from the cell’s cytoplasm and allow vacuoles to merge with one another and with other cellular double membranes. These membranes resemble those of other eukaryotes in structure and function, suggesting that vacuoles were once independent, free-living organisms whose functions have been adapted. Vacuoles regulate the amount of water and the pH levels in the cell, they store and isolate harmful materials, and they allow nutrients and other substances to move in and out of the cell. More than one vacuole can exist in each fungal cell or compartment.
Vacuoles are formed by the fusion of multiple vesicles. Vesicles are similar to vacuoles, but they are smaller. Specialized vesicles, lysosomes, are used in lysis, the breakdown or death of cells. They form when smaller vesicles merge and serve to contain and neutralize harmful molecules that would damage or kill the fungi if released into the cellular cytoplasm. Peroxisomes are enzyme-filled vesicles used in breaking down long fatty acid chains, which is important for creating metabolic energy. They are also involved in breaking down amino acids and decomposing hydrogen peroxide (H2O2), a byproduct of digestion that is harmful to fungal organelles.
Each fungal cell contains one or more membraned nuclei. This is very different from plant and animal cells, which can have only one nucleus. In some fungi, free-floating nuclei can move through the septa between cells. These nuclei hold the genetic material, DNA, responsible for reproduction. Inside the nucleus is a nucleolus that produces ribosomes and contains proteins and RNA. The number of chromosomes in the cell, as well as the size and number of nuclei, depend on the individual fungus type.
The membrane surrounding the nucleus has large (for a fungus), tubelike extensions that create the endoplasmic reticulum. This tubular network transports proteins, synthesizes lipids, and stores calcium used in signaling and carbohydrate metabolism. It also produces new membrane material for the cell membranes, vacuoles, and vesicles, and it adds carbohydrates to glycoproteins.
As with plant and animal cells, the endoplasmic reticulum is divided into two parts—rough and smooth. The rough endoplasmic reticulum holds on to lots of ribosomes, protein-building machines, and also contain RNA and amino acids. Further along, as the endoplasmic reticulum becomes less populated by ribosomes, it smooths out; in this area, lipids are created to export or use in membrane creation.
Golgi bodies (named after Italian histologist Camillo Golgi), often called the Golgi apparatuses, are flat-membraned compartments that sort and package molecules for transport. They are stacked in plant and animal cells, but in some fungi this is not the case, which led some scientists to think that fungi did not have Golgi bodies. The electron microscope solved that mystery.
Golgi bodies finish up the jobs started in the endoplasmic reticulum. Specialized enzymes trim and cut up proteins and lipid chains that are synthesized in the endoplasmic reticulum to be routed inside or outside the cell. The modified molecules are loaded after labeling, so to speak, in the Golgi. The whole process is like the operations in a massive shipping hub that can pack and ship materials to ensure that they arrive at their destinations on time.
Mitochondria (mitochondrion, singular) serve as little generators that supply energy throughout the fungal cells. These double-membrane–bound organelles are essential to the existence of all higher life forms on Earth. They create adenosine triphosphate (ATP), the source of energy used throughout living cells, and serve as the seat of aerobic respiration in the fungi. They also power the transport of material to the Golgi bodies, create intracellular membranes and associated embedded proteins, and control cell growth and death. Mitochondria produce proteins for their own use and contain their own DNA, suggesting that their ancestors were free-living organisms that entered into a mutual endosymbiotic relationship with eukaryotes.
The cytoskeletal system in a fungus supports the structure of the cell from the inside. It comprises a complex transportation network of fibrils, microtubules, and tubules. Molecular motors in microtubules transport materials, organelles, nuclei, and other important cargo throughout the fungal hyphae and are so important to how fungi function that you can kill a fungus by interfering with the synthesis of microtubules. The cytoskeleton’s microtubules also play a vital role in cell growth. You may remember coming across diagrams of mitosis (cell division in which pairs of chromosomes split and are aligned) in high school biology. The movement of chromosomes during mitosis is also aided by microtubules, which are visible under a common microscope. In addition, the chitinous material made in the cytosol (the liquid part of the cytoplasm) is transported to the cell wall via microtubules, which are important to the growth of fungal hyphae tips.
Hyphae can grow as much as 40 micrometers a minute. Cells grow apically, at the tips, in response to water, gases, and nutrients in the soil, moving toward favorable and away from unfavorable food sources. Hyphal growth gives a fungus the ability to move relatively long distances.
New molecules are constantly being pushed into the hyphal tip and along the cell wall, extending the wall outward and elongating the hyphal tube. The fungal cytoplasm transports vesicles containing the building materials necessary for wall construction. As the hyphae grow, they form new branches of hyphal tubes that explore and exploit new areas. Each tip of every newly formed hyphal branch performs exactly the same function as the original hypha.
Before electron microscopes, scientists using ordinary light microscopes identified dark spots in the tips of actively growing hyphae. When hyphal growth stopped, the spots disappeared. Since then, scientists have learned that the presence of this organelle, the Spitzenkörper, indicates not only an actively growing fungus but the direction of growth. All that a fungus is starts at the Spitzenkörper. The vesicles in the Spitzenkörper travel from the Golgi bodies through the microtubules and actin microfilaments. The protein actin is a major component of the fungal cytoskeleton and plays a crucial role in cell growth and intracellular transport.
Growing in an outward direction enables a hyphal tip to exert tremendous force. It has been reported that some fungal hyphae can produce 1200 pounds of force per square inch (218 kilos per centimeter), which is amazing when you consider that all this happens because of a few enzymatic processes.
While growth is occurring in the apical zone, at the other end of the hypha, in the older parts, things are deteriorating. The cells undergo autolysis, or self-destruction. What can be reused is moved toward the hyphal growing tip, while the nutrients contained in the rest of the decaying fungus become available to other organisms in the soil food web. In addition, hyphae leave behind a system of microscopic tunnels in the soil, through which air and water can flow, contributing to soil structure and health.
The Spitzenkörper is the dark area located at the hyphal tip. The microtubules (shown in red) and actin microfilaments (shown in brown) help transport vesicles produced in Golgi bodies to the tip.
Unlike plants, fungi are heterotrophic, incapable of making their own nutrients via photosynthesis. Like animals, fungi must obtain food from other organisms. Heterotrophs are categorized in several ways, based on how they derive their food.
Saprophytic fungi, or saprobes, are nature’s recyclers. They absorb nutrients from dead or decaying organic material, such as fallen trees, dead insects and animals, and even animal wastes. These fungi release minerals for plant uptake and in the process play a key role in the operations of soil food webs. Without saprobes, forest organic material would accumulate ad infinitum.
Parasitic fungi obtain their food from the cells of living organisms, attacking and sometimes even killing their hosts; for example, the Ophiocordyceps unilateralis fungus infects and kills certain species of ants. They cause skin problems such as ringworm and athlete’s foot and plant diseases such as apple scab, mildew, and rust.
Some facultative fungi can function as parasites or as saprobes, depending on the available food sources. Facultative saprobes can turn parasitic and invade a living host to obtain food. The opposite is also true: if a facultative parasite’s host happens to die, the fungi will continue feeding as a saprobe. Unlike parasitic fungi, which usually don’t kill their host lest they endanger their own well-being, facultative fungi have less reason to care if their partner lives or dies, because they can change to take advantage of either situation.
A red-banded polypore, sometimes called a conk, is a saprobe fungus that absorbs nutrients from dead or decaying organic material.
Symbiont fungi are mutualistic, deriving food from hosts in a way that mutually benefits both organisms. Mutualistic fungi are partners in all lichens, which consist of a fungal symbiont and an algal or cyanobacterial partner.
Mycorrhizal fungi are clearly mutualistic. The fungal symbiont provides its botanical symbiont (host plant) with nutrients and receives carbon in return. Mycorrhizal fungi are also saprobes, capable of breaking down organic materials as well as mineralizing inorganic matter. They are not the best decomposers of lignin and cellulose, but they can nibble at them. They are weak saprobes and decompose material that is already in an advanced stage of decay.
Spores from the parasitic fungus Ophiocordyceps unilateralis infect and kill ants (here Camponotus leonardi). The stoma of the fungus emerges from the back of the ant’s head, and the perithecium, from which spores are produced, grows from one side of the stoma.
An enormous range of food is available to fungi, and it seems that no matter what the material, a fungus has evolved to digest it (consider, for example, Amorphotheca resinae, the kerosene fungus, which eats creosote). Some generalist fungi can digest a wide range of materials, others have specific nutrient requirements, and many are in between. Many fungal species have developed the ability to break down and digest certain kinds of food, producing and releasing particular enzymes according to the environment in which they are usually found.
Although digestion is always extracellular, or outside the cell, fungi must internally produce the enzymes needed to break down their food sources. Because digestion occurs just outside the hyphal tip, most of the enzymes for digestion are released there after being assembled in the Spitzenkörper.
The vesicles in the constantly moving cytoplasm also include enzymes for activating growth and taking apart the cell walls at the hyphal tips, which is necessary so that new sections can be inserted. Several powerful enzymes capable of dissolving all but the most recalcitrant carbon compounds are released as the new cell walls are put in place. All this requires tremendous amounts of energy, which is ultimately supplied by the host or the fungal symbiont.
A range of isozymes, complex specialized enzymes, work together to break down foods. These powerful enzymes convert lignin, cellulose, and other tough organic matter into simple sugars and amino acids that are small enough to be absorbed through the plasma membrane in the hyphal tip. Each isozyme has some small variation in how it does its job, breaking down different parts of the material synergistically and rapidly to ensure complete decay.
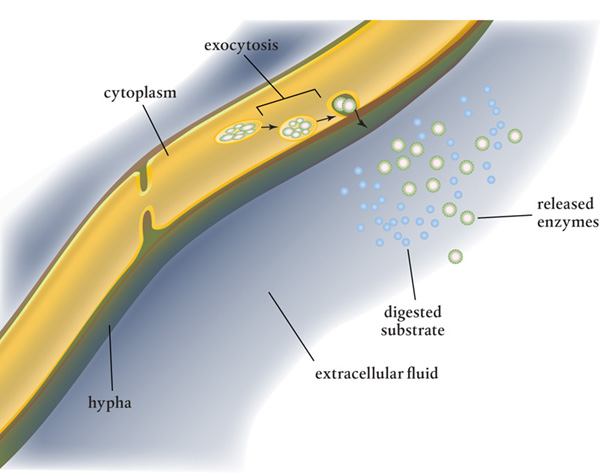
Enzymes are released through the plasma membrane and are delivered by membrane-bound intracellular vesicles that merge with the membrane, spilling or releasing their contents into the cell. In this exocytosis process, some of these enzymes work directly within the cell wall while others are diffused through to the extracellular environment. Many of these remain in the soil as the hyphae move on, breaking down organic matter and making it available to plants and animals in the soil food web.
Two biological processes move oxygen, water, and nutrients across the cell wall and plasma membrane, into and out of the cell. In passive transport, molecules pass directly through the plasma membrane from areas of higher concentration to areas of lower concentration to maintain equilibrium inside and outside the cell. This kind of transport involves diffusion and osmosis. Active transport, on the other hand, uses a chemical energy source, ATP, the high-energy molecule found in every cell, to pump larger molecules against the concentration gradient and through the plasma membrane.
Extracellular digestion occurs when enzymes created inside the fungal cells are exported outside the hypha by exocytosis.
The fungal plasma membrane is quite the gatekeeper. The molecules absorbed passively through the membrane are very small, about 4000 to 6000 Daltons in size. (Abbreviated Da, 1 Dalton is approximately the mass of a single proton or neutron. For comparison, an average DNA strand is huge, at about 900,000 Da.) Only water, oxygen, carbon dioxide, and very simple sugar molecules are small enough to make the grade. Molecules too large to be absorbed use specialized transport proteins made by the fungus and embedded within the plasma membrane that control what enters and exits the cell. These proteins operate exactly like the embedded protein transporters in the plasmalemma in plants and animals. Each protein is constructed within the cell and then embedded into the membrane. With very few exceptions, only charged ion particles pass through the channels formed by these proteins. Each embedded protein is monoparticular, allowing only one specific kind of molecule to pass through—so, for example, phosphate ions cannot move through an embedded protein designed for calcium ions. Each to its own.
As with plants and animals, each of the several nutrient molecules, such as potassium and phosphorus, requires a different carrier protein to transport it across the plasma membrane into the fungal cell. These nutrient molecules attach themselves to the carrier, which ferries them across the membrane via carrier-facilitated active transport. Very often, larger sugars enter fungal cells in the same way, using specific carrier proteins. Sugars are critical to the heterotrophic fungi and enter the fungal cytoplasm both passively and actively, emphasizing their importance as nutrients.
Water too is critical to all life, and without the ability to absorb some water, a fungus, like any living organism, would die. Because of water’s small molecular size and bipolar nature (with positive and negative charges on opposite ends of each molecule), water molecules can enter a fungal cell directly though the fungal membrane by diffusion. But if a fungal cell needs more water, it brings it in through aquaporin proteins that form little channels through the plasma membrane.
Nitrogen molecules, which build proteins and nucleic acids, are derived by fungi from the decay of organic and inorganic sources. Without nitrogen, there would be no enzymes. The molecules enter the cell as nitrates (NO3–) and ammonium (NH4+) via diffusion. In some fungi, in order to take up nitrates, glucose must be present in the cytoplasm.
Celluloses are digested by the cellulase enzyme, which results in glucose and other sugars. These molecules require active transport carrier proteins to help move them across the plasma membrane. Some fungal cells use more than one carrier system to transport the same kind of sugar; this stresses the importance of sugar to fungal organisms that cannot create their own because they do not photosynthesize.
Finally, amino acids freed up by extracellular digestion are also needed to create proteins. These large molecules require active transport via a carrier protein to move into a fungal cell. Clearly, extracellular digestion and the absorption of nutrients into the fungi is a complicated dance worthy of its own book.
In addition to releasing extracellular digestive enzymes, fungi produce and release secondary metabolites, which are organic chemicals unrelated to digestion. Some of these products control or impact other microbes in the vicinity, some speed up or slow down the growth of host plants, and others can kill other plants’ roots. These metabolites include well-known antibiotics such as penicillin, while others, such as aflatoxins, are deadly to humans and other animals.
Metabolites produced by mycorrhizal fungi are used in the mycorrhizosphere, the area surrounding the fungal hypha, which corresponds to the rhizosphere, the area immediately surrounding a plant root. These metabolites act just like the exudates from plant roots in the rhizosphere by attracting bacteria, other fungi, and microorganisms. Some of the benefits of mycorrhizae probably result from these organisms. Fungi can adjust their metabolites to attract, sustain, or repel them. One group of organisms in particular, phosphorus-solubilizing bacteria, are known to help with the digestion of phosphorus.
Fungi are often placed into one of two categories: perfect fungi are capable of making spores and can reproduce either asexually (with only one parent) or sexually (with two parents). Imperfect fungi reproduce only by asexual spores, or at least they have never been seen to reproduce sexually.
The most common type of fungal asexual reproduction is through the formation of spores that are dispersed from the parent organism. Some fungi develop sporangia, budlike cells that hold onto young sporangiospores until they mature and separate from the parent cell. Other fungi develop spores outside the reproductive cells. Still others create sclerotia, storage structures made up of compact fungal tissue, some of which can survive dormancy for long periods in adverse environmental conditions.
Fungi can also be asexually cloned from hyphal cuttings. Some mycorrhizal fungi form vesicles that can reproduce or clone the fungi. Still, nothing beats sporulation in terms of numbers. It is the reason fungi are so prolific and successful.
Sexual reproduction requires two mating types and can occur in several ways. In some fungal species, meiosis (nuclear division) occurs in the asci, single-celled spore structures, which results in the formation of exactly eight spores. These fungi shoot out their spores to distances of up to several centimeters. Given the size of the organism, this is comparable to tossing an American football a couple of miles (or kilometers). Consider the osmotic pressure that must be involved. In other species, basidia structures develop to produce and hold or eject spores. The pressure in this case is decidedly weaker. These tiny spores are dispersed by wind currents and can travel long distances.
One other point is important to mention. Scientists have discovered that fungi exchange DNA with other fungi and with other microorganisms. (Don’t be surprised, because human bacteria do it all the time.) They seem to do this based on environmental conditions that require some sort of adaptation. This has all manner of ramifications, many of which are being sorted out by lots of further study.
Some mycorrhizal fungi types (specifically, ectomycorrhizal fungi) form mushrooms, the specialized fruiting bodies that play a role similar to that of flowers in plants. Mushrooms can grow below the soil, as do truffles, or above, as do morels. Their caps can be gilled or nongilled. Some have economic and gastronomic value, such as porcinis and chanterelles. Not all mycorrhizal fungi produce mushrooms, although all mushrooms are fungi.
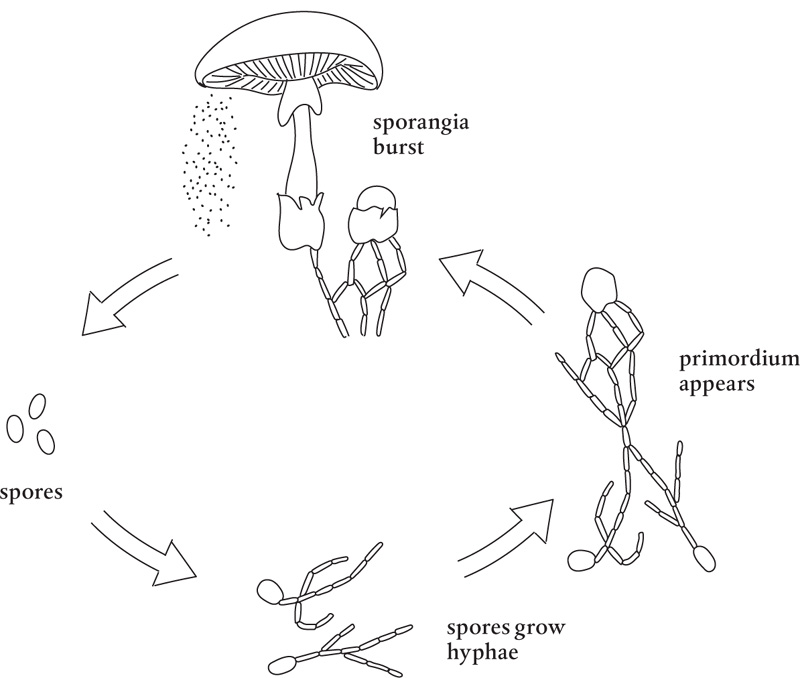
Mushrooms are typically the part of the fungus that carries and disperses fungal spores, usually through sporangia (sporangium, singular), the enclosures in which the spores are formed. Each tiny spore released from a sporangium gives rise to a hypha, which branches and grows into hyphae, which then continue to grow and interweave to form a mycelial web.
Sometimes the hyphae form rhizomorphs, structures that transport water and nutrients throughout the network. This occurs when parallel hyphae merge. The inner hyphae lose their nuclei and cytoplasm, becoming hollow.
Merging hyphae also form fruiting bodies as a result of hyphae growing together. Specifically, two homokaryotic fungal hyphae (each with genetically identical nuclei) merge cytoplasm, but keep their nuclei separate, to form a dikaryotic hypha. Every time the cell divides, the new cells keep the two sets of haploid nuclei separate.
At the appropriate time, given some cue from nature, some environmental condition stimulates the fungus, and signals are created in the circulating cytoplasm, which cause the hyphae to merge and form a primordium, a tiny knot in the mycelia. This primordium includes all the cells the mushroom will ever have. It enlarges into a tiny budlike structure, or button, which will grow into a full-size mushroom.
All a mushroom requires is water to fill the cells. The mushroom can rapidly pull in water from its mycelium and expand the primordium. It can take less than 24 hours for a primordium to fill and grow; this is why mushrooms can pop up so quickly. Mushrooms have outer membranes, or veils, that dehydrate rapidly, so they usually appear only when temperature and moisture conditions are right. Those hydrophobins are working full time.
The formal classification of fungi, whether perfect fungi or imperfect fungi, is a complicated subject and an ever-changing landscape, as new fungi are discovered and new classification systems developed. No one knows exactly how many different species of fungi exist, but about 100,000 have been identified and studied thus far. Scientists using modeling methods have estimated that as many as 5.1 million fungal species may exist (not surprising, considering that nearly 200 different species of fungi live on your feet). Fungal diversity is huge, to say the least, and scientists have developed a number of ways to subdivide and otherwise classify them.
Science has determined a system of classification of all living things, a hierarchical order based on the genetic characteristics of organisms. Fungi are classified as members of the Eukaryote domain at the top of the order. Next in line, the kingdom Fungi includes heterotrophic organisms that obtain food through absorption. Until the late 1960s, fungi were considered part of the plant kingdom (Plantae), but they were given their own kingdom when scientists finally concluded that the differences between plants and fungi were too great. The kingdom Fungi is now recognized as one of the oldest and largest groups of living organisms on the planet.
The fungal phylum groupings have been based primarily on the types of spores produced for reproduction and the reproductive structures that create them. DNA sequencing, however, is suddenly ending a lot of debate about what is a fungus and what is not and making it easier to make phylum assignments. In fact, all that is needed to determine a fungus’s phyla is some of its DNA. Even better, RNA can be used to identify individual species. Most scientists now recognize seven phyla of fungi, but only three of these include mycorrhizal fungi.
Ascomycota, also called sac fungi, is a diverse and large phylum that includes more than 30,000 species. All ascomycetes develop an ascus, a saclike reproductive structure that contains tiny ascospores. Many ascomycetes are plant or animal pathogens, and others are edible mushrooms. It is the phylum to which morels, truffles, brewer’s and baker’s yeasts, and penicillin belong. Less helpful ascomycetes include powdery mildews and the fungi responsible for apple scab, Dutch elm disease, and chestnut blight. These fungi break down cellulose in plants and collagen in animals, two extremely resistant materials.
Members of this phylum of about 30,000 filamentous fungi reproduce via a microscopic, spore-producing basidium, which is attached to a fruiting body. The word basidium means little pedestal and is derived from the way the fruiting body supports the spores. This phylum includes mushrooms, chanterelles, puffballs, stinkhorns, bird’s nest fungi, shelf fungi, rusts, and smuts.
The phylum Glomeromycota includes fewer species than the other two phyla, at about 230, but they are among the most abundant and widespread of all fungi. Arbuscular mycorrhizal fungi are glomeromycetes that form mutualistic relationships with the roots of most herbaceous plants and many trees. Most species reproduce asexually. These fungi play a large role in this book’s story.