Introduction to Mycorrhizal Fungi
MYCORRHIZAL FUNGI HAVE existed and supported plants since terrestrial plants evolved more than 450 million years ago. In fact, mycorrhizal associations are believed to be a major factor that enabled plants to survive on land. The earliest fossil records of plant roots contain arbuscular mycorrhizae that look almost identical to arbuscular mycorrhizae growing in modern soils.
Today we know the true importance of mycorrhizal relationships, and we understand how they operate: the fungus colonizes the root system of a host plant, increasing the roots’ water and nutrient absorption capabilities, while the plant provides the fungus with carbon it obtains from photosynthesis. Both organisms thrive as a result of this symbiotic relationship.
At about 2 to 10 micrometers in diameter, mycorrhizal fungi hyphae are considerably thinner than root hairs, which average 15 micrometers. Some of the mycorrhizal hyphae are extraradical (or extramatrical), meaning they extend beyond the root into tiny soil pores that root hairs would be too large to penetrate. Mycorrhizal fungal hyphae can grow to several centimeters in length, which enables them to access nutrients and water that the roots cannot reach.
Although we now know that mycorrhizal fungi live on the roots of 80 to 95 percent of all Earth’s plant species, not long ago little was known about these specialized fungi, except that they existed. Very few appreciated the crucial role these fungi play, and many even questioned whether they had any benefit at all or were detrimental to plants.
DISCOVERY AND GROWTH OF UNDERSTANDING
In 1885, German botanist Albert Bernhard Frank (1839–1900) published an extraordinary paper about what would later become known as ectomycorrhizal fungi. It would take more than 50 years for Frank’s observations and analysis to be confirmed, especially his conclusion: that a beneficial, not a parasitic, relationship existed between the fungi and host plants. During that time, while the botanical community argued over this point, a thriving commercial fungicidal industry developed. Frank’s fungi were deemed to have little importance.
In his paper, whose title in English roughly translates to “On the nutritional dependence of certain trees on root symbiosis with belowground fungi,” Frank combined the Greek words for fungus (mykitas) and root (riza) to form the German word mykorhizen. He theorized that each partner provided nutrients to the other in a beneficial, not a detrimental, manner and that the fungi were not killing the trees he studied but supporting them, and in turn were being supported.
Frank also believed that the netlike structures that appeared around the roots of pines, as found by Theodor Hartig in 1840 (who believed that these were parasitic fungi), were in fact formed by the same mycorrhizal fungi, and that these fungi absorbed minerals from the soil and humus and brought them back to the plant in return for carbon. Previous observers had concluded that the netlike structures were fungal, but they all insisted that it was an invading fungus and not beneficial to the plant.
After continuing his research, Frank identified two types of mycorrhizal fungi: he called them endotrophic (in which the fungus penetrates the roots of the plant) and ectotrophic (in which the fungus is formed between the roots). One of his graduate students, Albert Schlicht, found both kinds of mycorrhizal fungi in a variety of environments throughout Germany, which led Frank to believe that mycorrhizae were the norm, not the exception. How right he proved to be.
Although Frank is often given credit for discovering mycorrhizae, endomycorrhizal fungi had been observed at least as early as 1842 by Swiss botanist Carl Wilhelm von Nägeli, though he may not have known exactly what he was looking at under his microscope. (You may not have heard of von Nägeli because he discouraged Gregor Mendel from continuing his genetic work with peas.)
For a long time after Frank’s paper was published, scientists believed that mycorrhizal fungi were parasites. They were lumped together into a group called Rhizophagus, for root-eater. (Although this name did not endear these fungi to plant growers, it has returned to use as a genus of mycorrhizal fungi.) In the mid-1900s, scientists believed endomycorrhizal fungi were the cause of diseases that affected strawberries, tobacco plants, and legumes. Because the treelike structures, or arbuscules, inside the host plant roots are digested by the plant, scientists believed that the plant was protecting itself from an invading pathogen.
Research continued, and in 1943, during World War II, a Japanese scientist published a paper that showed that plants that formed mycorrhizae grew faster than nonmycorrhizal plants. The paper was written in a nearly unintelligible form of German and released at an unfortunate time, however, so few, if any, researchers were aware of his work. It wasn’t until the mid-1950s, when a paper by scientist Barbara Mosse discussed improved apple seedling growth resulting from mycorrhizal fungi, that the true benefits of these fungi became clear.
WHO NEEDS MYCORRHIZAE?
Without mycorrhizal relationships, most plants would probably not exist. Most members of the plant kingdom have formed associations with mycorrhizal fungi, including bryophytes (such as mosses), angiosperms (most land plants), pteridophytes (such as ferns and club mosses), and many gymnosperms (such as conifers). These plant–fungal associations have remained pretty much the same despite all manner of other evolutionary changes that have occurred. Although mycorrhizae are important to most plants, not all plants depend on these relationships to the same degree. Some plants will not survive without mycorrhizal fungi, while a few others do not require mycorrhizae at all.
Obligatory mycorrhizal plants cannot survive without mycorrhizal associations. These plants often have thick, nonbranching roots that cannot effectively explore soil for nutrients. Their seeds may not germinate without mycorrhizal colonization, or if they germinate, seedlings will not survive.
Facultative mycorrhizal plants can survive without forming mycorrhizae until nutrients become scarce. When this happens, mycorrhizal associations form or the number of existing mycorrhizae increases. Roots of facultative mycorrhizal plants tend to be extensive and branching, and they are capable of reaching large volumes of soil and nutrients even without an extraradical mycorrhizal network.
BENEFITS OF MYCORRHIZAL FUNGI
Plants involved in a mycorrhizal relationship are better able to withstand drought and other environmental stresses, root pathogens, and even foliar diseases. Plant biomass is often improved, along with the timing and number of flowers and amount of fruit produced. Mycorrhizae also offer much to the soil by improving structure as the fungal hyphae explore for nutrients.
Mycorrhizae are sensitive to environmental conditions and can behave one way in a study under greenhouse conditions and another way outdoors in the field. To complicate the situation, mycorrhizal fungi can exchange DNA with other microorganisms and shift the benefits they confer. Nevertheless, much scientific research has confirmed the importance of mycorrhizae and the benefits of mycorrhizal associations to plants. Keep these benefits in mind as you read about the various types of and uses for mycorrhizal fungi.
Increased nutrient supply
As a root grows, it slowly takes up nutrients from the surrounding soil. The plant eventually uses up the available nutrients in this zone of depletion or takes them up at a rate that cannot be sustained at a high enough level for the plant to thrive. As a result, roots continue to grow to reach new areas of soil to mine for nutrients.
By associating with fungi, however, a plant’s roots can use the nutrients brought back by the fungal hyphae growing beyond the zone of depletion. Moreover, because the hyphae are considerably thinner than the roots with which they associate, they are able to reach smaller pores in the soil within and outside the depletion zone that were not available to the larger roots. Although the mycorrhizal hyphae are smaller than roots, they greatly add to the surface area and increase absorption. One study, for example, found that fungal hyphae associated with a pine tree added 20 percent more mass to the tree’s root system.

White masses of fungal hyphae surround the plant roots. Because they are so much smaller than roots, fungal hyphae can enter smaller soil pore spaces to reach more nutrients.
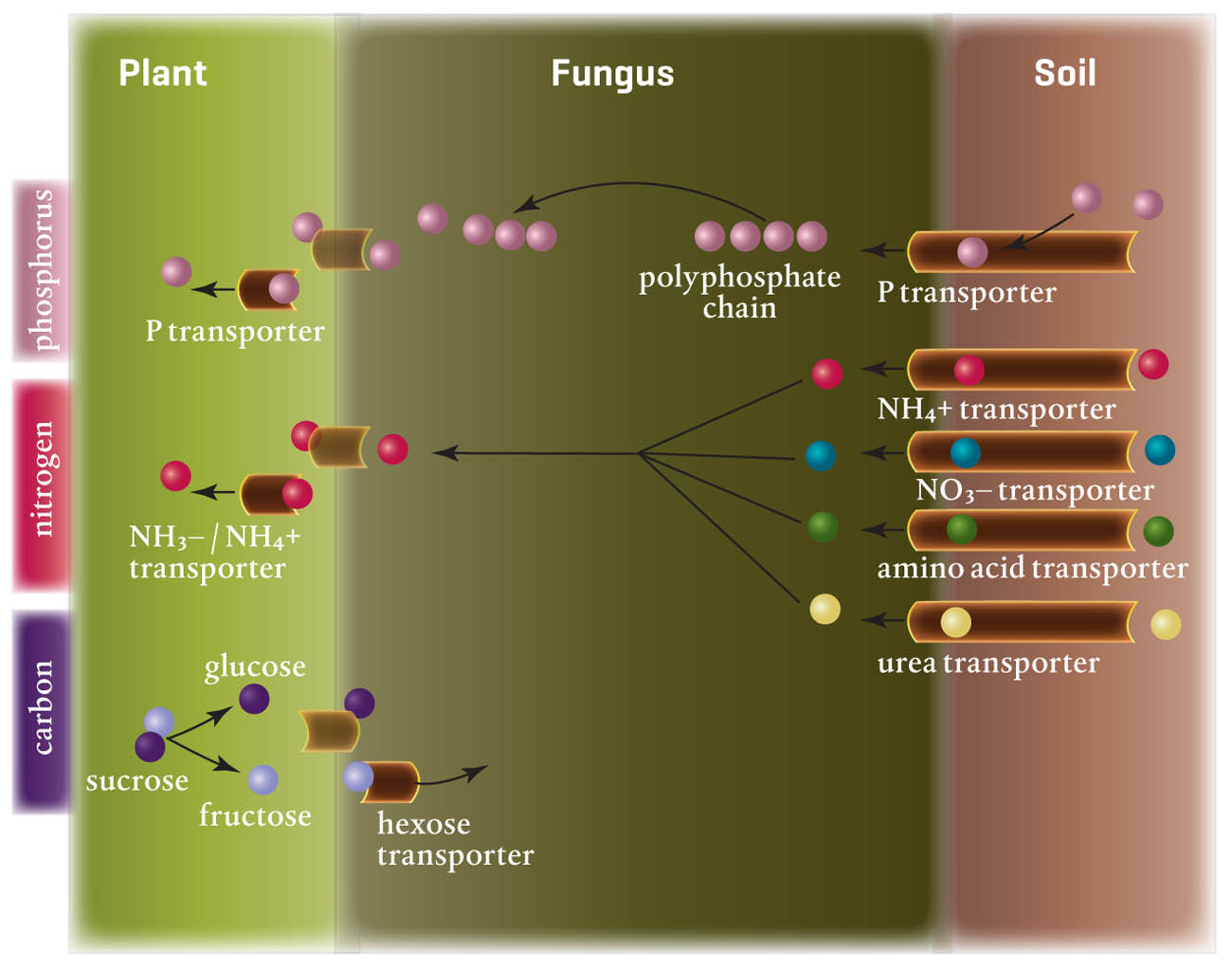
Mycorrhizal fungi transfer nutrients to the plant. In return, around 20 percent of the carbon produced by a plant can be transferred to its fungal partner. That is some sacrifice, but the returns are worth it. Phosphorus, an essential plant nutrient especially important because of its presence in ATP, the energy currency of life, is tied up chemically in the soil and difficult for most plants to absorb without the help of mycorrhizal fungi. Nitrogen, copper, zinc, iron, and nickel (all essential plant nutrients) are also taken up by mycorrhizal fungi and transferred to the host plant.
Mycorrhizal hyphae absorb the nutrients already in the soil, but their extracellular digestion can also result in decay that releases even more nutrients into the soil, especially nitrogen. In addition, as organisms in the mycorrhizosphere die, they release a variety of nutrients and produce all manner of metabolites that are absorbed by the fungi. And the fungi themselves are a food source for other soil food web organisms.
Drought tolerance
The existence of mycorrhizae can help host plants be more tolerant of drought conditions. Studies have demonstrated that from 50 to 100 times more water is available to plants when their roots associate with mycorrhizal fungi and form mycorrhizae.
Mycorrhizal fungi create water storage structures in the roots where they are needed. The net of mycorrhizal fungal hyphae branches surrounds plant roots and holds water, acting as a storage reservoir. The interface between some hyphae and the host plant root cells also holds water, as do vesicles, those special storage organelles inside the roots. This is a significant benefit of mycorrhizae. It is also important from an ecological standpoint, as agriculture accounts for about 70 percent of all water use worldwide, and mycorrhizal plants require less water during dry periods.
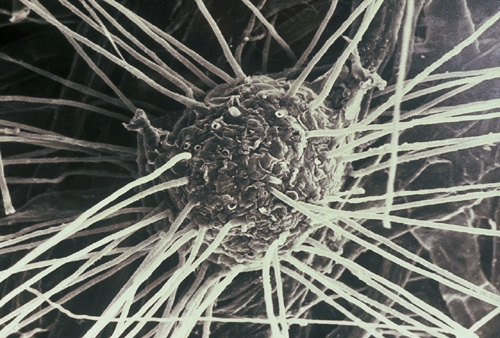
The ectomycorrhizal fungal hyphae on this pine root move out into the soil and explore for and mine nutrients to bring back to the plant in exchange for carbon.
Protection from pathogens
Mycorrhizal fungi create physical barriers around, and even within, roots, which can prevent some pathogenic organisms from successfully infiltrating the host plant. Mantles, layers of ectomycorrhizal hyphae surrounding some roots, can also make roots difficult to penetrate. If a pathogen does penetrate a root, it is often stopped by the internal structure of mycorrhizae.
Pathogens also have a much more difficult time foraging when they are competing for nutrients with mycorrhizal fungi. These fungi are intense competitors for nutrients in the soil and thus limit access to vital resources needed by pathogens. In addition, as fungal hyphae digest and ingest, they can produce metabolites, chemicals that decay and act as deterrents to other organisms’ foraging, and some can even produce their predator’s specific defensive and offensive chemicals.
Improved soil structure and carbon storage
As mycorrhizal hyphae weave through the soil, they bind particles together and create soil structure. Mycorrhizal fungi produce sticky exudates that help unite individual soil grains into water-stable aggregates surrounded by pore spaces, which are essential for the movement of air and water. In addition, the fungi produce glomalin, a carbon-laden compound that remains in the soil even after the fungi die. Glomalin molecules are a major source of carbon in the soil.
One of the most amazing things about mycorrhizal fungi is their ability to associate with more than one host plant at the same time—in other words, their networks can be shared among plants, even plants of different species. As a result of this feat, mycorrhizae can benefit entire forests, as the larger trees literally feed and protect the smaller trees through an interconnected mycelial network. And when one plant dies, many of its nutrients are returned to the network and flow toward other plants.
Protectionism
Some mycorrhizal fungi can make it difficult or impossible for plants other than their hosts to grow, which slows down or stops successional changes. The fungi use several mechanisms for this—some produce chemicals that weaken or counteract the chemical components necessary for the invaders’ survival, while others influence the microbes in their vicinity with metabolites that attract or repel other plants. They also compete for or provide food sources to protect their host plants.

Root cross-section with several mycorrhizal types.
MYCORRHIZAL TYPES
Mycorrhizal fungi are commonly divided into two groups according to how the fungal cells associate with plant cells. The hyphae of endomycorrhizal fungi penetrate the cell wall, but they do not enter the cell beyond the plasma membrane; these mycorrhizal types are most often associated with the roots of vegetables, grasses, flowers, shrubs, and fruit and ornamental trees. The hyphae of ectomycorrhizal fungi do not penetrate all the way through the cell wall; they form ectomycorrhizae mainly with conifers and some deciduous trees such as oaks.
Endomycorrhizal fungi fall into three main subgroups, each of which has adapted to particular types of host plants, after which the relationship is named: arbuscular, ericoid, and orchid mycorrhizae. Of these, arbuscular mycorrhizae are by far the most common and widespread type. They are of great importance to growers.
ARBUSCULAR MYCORRHIZAE
About 70 percent of all terrestrial plants form the same arbuscular mycorrhizal associations found in fossils from the Devonian period, about 420 million years ago. All arbuscular mycorrhizal fungi are members of the phylum Glomeromycota and form the dominant type of mycorrhizae. Although there are only about 230 species of glomeromycetes, they form mycorrhizae with more than 400,000 different plants. These fungi are not very host-specific and form other associations in nature, including those with many liverworts and mosses. They are not discriminating.
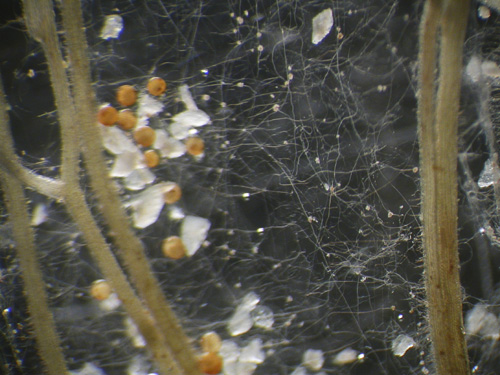
Strands of Gigaspora margarita, a glomeromycete, surround a plant’s roots.
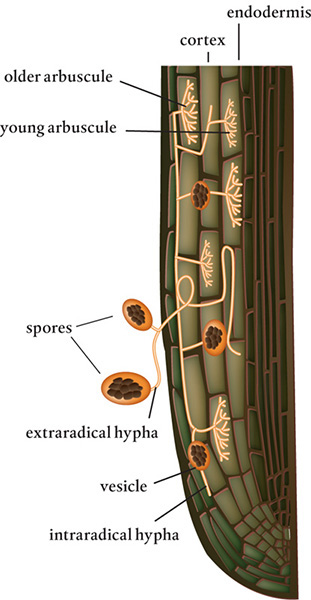
Although an arbuscular mycorrhizal fungus penetrates the plant root’s cell wall, it never penetrates the plasmalemma. Instead, the cell forms a membrane that surrounds the fungal hypha, enclosing it in an envelope of sorts. The plant continually builds this membrane as the fungus grows, branches, and produces arbuscules, the tiny, finely branched clusters of hyphae. The enveloping structure creates a cavity into which the two partners deposit their molecular payloads. Many arbuscular mycorrhizal fungi form temporary intercellular vesicles that store water and nutrients and sometimes produce spores.
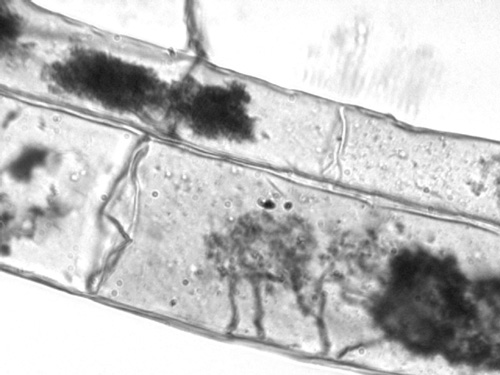
The microscopic arbuscular system; the dark smudges are arbuscules.
How arbuscular mycorrhizae form
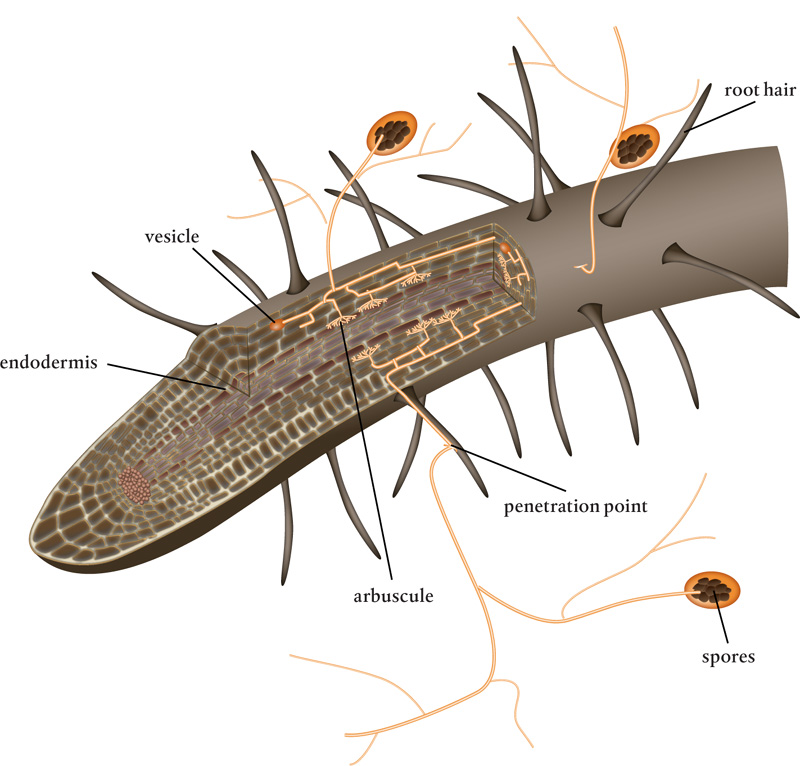
The process of forming arbuscular mycorrhizae is initiated by the plant when soil nutrient conditions, specifically phosphorus levels, are low. Such conditions increase the production and release of strigolactones from the host plant’s roots. These specialized hormones attract fungal spores or hyphae in the soil. Once the strigolactones are discovered by the fungus, they guide the fungal hyphae to the host plant’s roots. An arbuscular mycorrhizal spore has about seven to ten days to reach a root to get carbon before its own on-board supply runs out and it dies.
Strigolactone molecules are sesquiterpenes, a subset of the terpene group of chemicals, which are found in plants and insects and provide defense against plant–insect and plant–fungus interactions. (Sesquiterpene molecules are also present in the essential oils of products such as myrrh, sandalwood, and cedarwood.) These hormones are released in very low concentrations, which indicates a high susceptibility on the part of the fungus for detecting the signal.
Once discovered by germinating arbuscular fungi, strigolactones cause fungal hyphae to undergo extensive branching. This increases the number of hyphal tips and thus improves the chances of timely contact with the plant roots. Strigolactones also guide the fungi to grow toward the roots and aid them in the formation of an appressorium, the penetrating mechanism used by fungi to enter the roots. But this is not a one-way dance. The fungi also produce and send chemical signals necessary to prevent the plant from turning its defenses against the incoming mycorrhizal fungi.
These fungal-produced signals, or Myc factors, have been identified as lipo-chitooligosaccharides, a big word that represents an important chemical group that facilitates the close interaction of rhizobia bacteria and leguminous plants. Even low concentrations of lipo-chitooligosaccharides can be detected by plants. When they are detected by plant roots, root growth is stimulated. In fact, this stimulation is important enough to encourage scientific research targeting the development of synthetic molecules that will do the same thing.
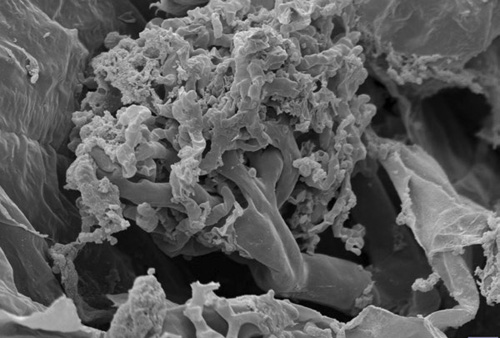
An image from a scanning electron microscope shows an individual arbuscule, which resembles a tiny tree.
The hypha of an arbuscular mycorrhiza.
A plant root tip colonized by arbuscular fungi.
When one of the hyphal tips comes into contact with a root as a result of the signaling, an appressorium forms and the epidermis in the root is penetrated. The appressorium is a specialized cell that produces a peg, which is pressed into and then grows into the root. What follows is a coordinated dance between the fungal hyphal membrane and the plant’s cell membrane. As the arbuscule develops, its cell wall thins and loosens. In the meantime, vacuoles in the plant cell break up, and the resulting pieces of double membrane concentrate around the invading fungi. These membrane bits merge with the existing plant cell plasmalemma. The hyphal tip is completely surrounded, so it never penetrates the membrane. What forms is an apoplastic cavity that is not directly connected to the inside of the plant cell, where nutrients can be transferred in both directions.
At 10 micrometers wide, a hypha can end up supporting an arbuscule with branches less than 1 micrometer in diameter. These structures are short-lived, and after a few days they start to fall apart. The hypha remains, however, sometimes for months or even years, as do the vesicles formed with the arbuscular features. Vesicles start to form at the same time as the arbuscules, between and inside the host cell walls. They can remain in the root long after the arbuscules are gone.
Meanwhile, as the fungi grow into the soil, they create extraradical hyphae, the fungal network outside the root that increases the surface area for absorption. This mycelial network can enable a plant root to access 100,000 times more soil than a root can access without a mycorrhizal association.
Arbuscular fungal hyphae are almost always nonseptate. If there are septa, they are few and far between. Nuclei and cytoplasm flow throughout the fungal network, which under a microscope can look at lot like the Los Angeles freeway system at rush hour.
Arbuscular mycorrhizal spores
Arbuscular mycorrhizal fungi produce very distinctive spores on the roots of the plant and in the soil. For most fungi, a spore 5 micrometers in diameter is big, but arbuscular mycorrhizal spores are gigantic by comparison: they can measure up to 1000 micrometers in diameter, though most are between 50 and 250 micrometers. You can find many images of arbuscular fungal spores on the Internet to help you to distinguish them by their colors and shapes—they are quite beautiful.
The spores’ thick and hard outer shells result in longer shelf life, so to speak, and the ability to withstand abuse, such as passing thorough the digestive track of an insect or a rodent. Because they are so large, arbuscular fungal spores don’t travel far in the wind, so catching a ride inside an animal (and being passed through the digestive tract) helps disperse them greater distances.
ERICOID MYCORRHIZAE
Ericoid mycorrhizal fungi form mutualistic symbiotic relationships with members of the plant family Ericaceae, which includes rhododendrons and azaleas (Rhododendron spp.) and blueberries and cranberries (Vaccinium spp.). These plants grow in acidic, peat-rich soils and make up about 5 percent of terrestrial plant species. The largest fungal group that enters into these relationships are the ascomycetes, which have adapted to the fine root system of ericaceous plants. These roots have an epidermis and a one- or two-cell–layered cortex. The ericoid mycorrhizal fungal hypha penetrates the cortical cell wall and forms a dense coil structure, or peleton.
A veil of hyphae grows over the surface of the roots, but it is finer than and not as thick as an arbuscular mantle. Ericoid mycorrhizal fungi do not penetrate as deeply into the soil as other mycorrhizal fungi. They specialize in obtaining nitrogen from organic matter in environments where nitrogen is a limiting factor for plants, producing strong acids that break it up. This has two implications: the fungi free up a lot of hydrogen ions, and the soil environment becomes acidic. Fortunately, ericoid fungi help the plants with which they associate survive in acidic soils that would not be suitable for other plants.

The distinctive spores of glomeromycetes come in many colors and textures.
ORCHID MYCORRHIZAE
About 10 percent of the Earth’s plant species are orchids (family Orchidaceae), and most of them depend on specialized endomycorrhizal fungi from the Basidiomycota at some point in their lives. Orchid seeds are tiny and do not contain sufficient nutrients to support the growing embryonic plant; they get what they need from the mycorrhizal association.
The orchid seed germinates and sends out a few hairs that are immediately colonized by fungi. The hyphae grow into the root cell epidermis and the cortex, which is thicker than in ericaceous plants, allowing for a complex network of peletons. The peletons live for only a few days and are then absorbed by the plant. These associations can be temporary, disappearing later in the plant host’s life or being replaced by another kind of symbiotic fungi. Some orchids are heterotrophic and do not contain chlorophyll, so they rely on their fungal partners to obtain nutrients throughout their lives.
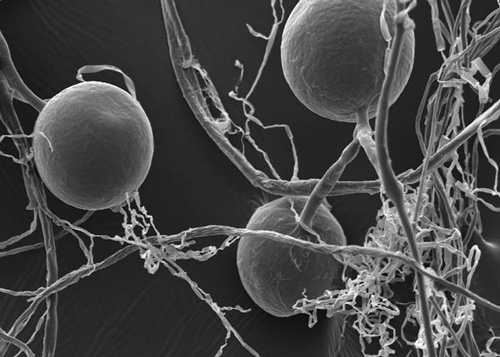
Although some are visible to the naked eye, most arbuscular fungal spores are visible only under magnification.
ARBUTOID MYCORRHIZAE
The fungi that form arbutoid mycorrhizal relationships are the Basidiomycota. The most important host plants in these mycorrhizal relationships are the Pacific Northwest madrone tree (Arbutus menziesii), from which the arbutoid name is derived, and manzanitas and bearberries (Arctostaphylos spp.). In arbutoid mycorrhizal associations, a mantle surrounds the roots of the host plant, and intercellular Hartig nets are formed. Sometimes, however, arbutoid mycorrhizal fungi will penetrate the root’s cell wall, as in endomycorrhizae. The hyphae penetrate the outer cortical cells of the plant roots, forming tiny coils that allow for the transfer of nutrients.
MONOTROPOID MYCORRHIZAE
Monotropoid mycorrhizal fungi were once thought to be part of the arbutoid group, but they do not penetrate the plant cell walls. They colonize plants of the family Monotropaceae, which includes Indian pipes (Monotropa spp.), a woodland plant that lacks chlorophyll. In fact, all plant hosts in this group lack chlorophyll. The fungi form an ectomycorrhizal association with trees such as beech (Fagus spp.), oak (Quercus spp.), and cedar (Cedrus spp.) and then form a monotropoid association and transfer some of the trees’ carbon to the plants.
Monotropoid mycorrhizal fungi form a dense sheath, or mantle, around the root. These fungi also form Hartig nets, which surround but do not penetrate the root cells. Some individual hyphae form fungal pegs, which do penetrate the cortex. These live inside the host plant for a couple of weeks and then die. The host plant absorbs the fungal hyphae after they die to obtain the carbon it needs.
ECTOMYCORRHIZAE
Ectomycorrhizal fungi are more modern than endomycorrhizal fungi. They evolved to associate with plants about 250 million years ago. Although there are several thousand different types of ectomycorrhizal fungi, only about 5 percent of terrestrial plants form ectomycorrhizal associations with ascomycetes or basidiomycetes. In addition, many of these fungi produce mushrooms next to their plant hosts and are not only recognizable but often of economic and culinary value. Ectomycorrhizal fungi are more acidophilic than other mycorrhizal fungi—that is, they prefer acidic soils with a low pH.
The plant hosts of ectomycorrhizae are always woody plants, generally trees, including pine (Pinus spp.), beech (Fagus spp.), birch (Betula spp.), and myrtle (Myrtus spp.). Although there are fewer plant hosts than exist for endomycorrhizal fungi, the plants that are colonized by ectomycorrhizal fungi include some 25,000 tree species found across the globe. This may have to do with the diversity of their fungal symbionts, of which there are a few thousand.
Ectomycorrhizae formation begins with fungi–root contact below the apex of young, actively growing roots. The hyphae first grow on the surface of the root. After a day or two, a mantle is formed around the root, followed by penetration between the host cortex cells, branching, and growth, which results in the formation of the Hartig net between the cells. In angiosperms (herbaceous plants, shrubs, and most trees), this is usually a single cell layer and it does not move out of the root’s epidermal cells. In gymnosperms (conifers, ginkgo, and cycads) that associate with ectomycorrhizae, the Hartig net can penetrate the cortex.

A fairy ring of mushrooms around this white spruce may indicate ectomycorrhizae formation below ground.
As the ectomycorrhizal fungus covers the roots, only a small portion that sits about 1 to 3 millimeters from the growing tip is active. As the root grows, this active zone continues to move outward, and older parts of the Hartig net die. Meanwhile, the fungal hypha that spread into the soil from the mantle will grow parallel to one another and branch. They can also merge to form rhizomorphs, hollow structures that store and transport water, or sclerotia, which store nutrients. Rhizomorphs, in particular, can look like roots to the naked eye, and they perform some of the same functions. They can carry nutrients and water, sometimes very long distances.
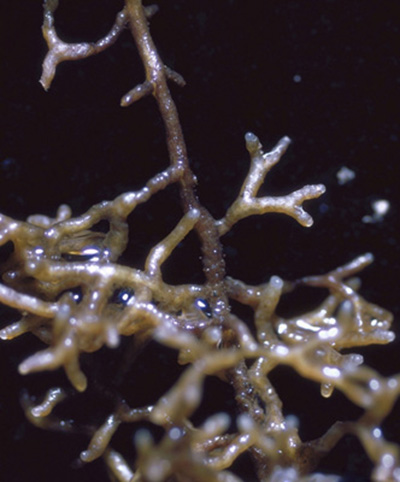
Classic ectomycorrhizae with red pine as the host.

Amanita muscaria (fly agaric) is a fruiting body of an ectomycorrhizal fungus that colonizes birch trees.
ECTENDOMYCORRHIZAE
Though rare, some mycorrhizae have the characteristics of both endo- and ectomycorrhizae. This seems to be confined to a few species of deuteromycetes, which are imperfect fungi. Ectendomycorrhizae form in pot-grown nursery seedlings and with some tree seedlings, mostly pines, after forest fires. The host can be either coniferous or deciduous. A thin mantle is formed, and the hyphae of the Hartig net penetrate the cortex cells but not the plasma membrane. Eventually, these associations morph into ectomycorrhizae as the seedling matures.
SEBACINOID MYCORRHIZAE
A special group of basidiomycetes in the order Sebacinales, the sebacinoids are very diverse in terms of their mycorrhizal relationships. Some form endomycorrhizae while others are ectomycorrhizal in nature, or they form ericoid and even orchid mycorrhizae. Sebacinoid fungi can colonize the tree roots of Eucalyptus marginata, one of the most common species of native eucalyptus in Australia.
Piriformospora indica, a sebacinoid fungus discovered in desert soils in northwest India, could have huge agricultural and commercial value. When scientists grew it in the lab, it successfully colonized several crop plants, including corn, barley, wheat, parsley, and poplar. Experiments have shown that P. indica–inoculated plants are more resistant to some root pathogens and diseases.
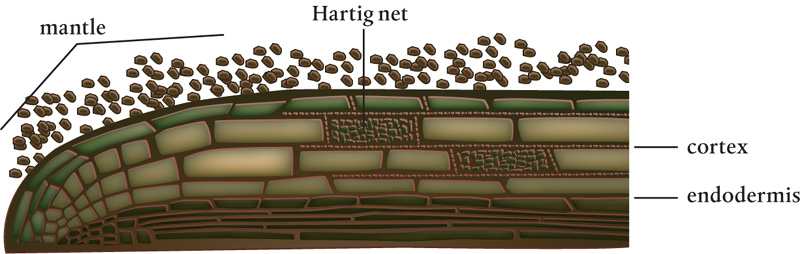
Hypha of ectomycorrhizal fungi.
PLANTS WITHOUT MYCORRHIZAL ASSOCIATIONS
Plants that do not form mycorrhizae are the exceptions. Seed and plants of the families Brassicaceae (such as mustards, kales, and cabbages), Caryophyllaceae (such as pinks and carnations), Chenopodiaceae (such as amaranths and goosefoot), Polygonaceae (such as knotweed and buckwheat), and Portulacaceae (such as purslane) germinate and remain healthy without mycorrhizal associations. These plants usually have extensive and quick-growing specialized root systems that make the most of limited nutrients in poor soils such as sand. Some nonmycorrhizal plants have very long roots covered with fine root hairs, which can explore soil pores better than most other plants, without the help of mycorrhizal fungi. Others have developed carnivorous capabilities and get nutrients from aboveground organisms such as insects. Still others, such as epiphytes (most orchids), can grab nutrients from the air, and some have evolved into parasites.
Proteas (Proteaceae) and lupines (Lupinus spp.) have cluster roots, closely spaced lateral roots that are densely covered with root hairs. These roots form near the soil surface so they can feed in the duff layer. To obtain phosphorus without mycorrhizal fungi, they support lots of phosphate-solubilizing bacteria in their rhizospheres. Their exudates are full of organic acids that decay organic matter.
Some members of the Cyperaceae (sedges) have dauciform roots—swollen, carrot-shaped lateral roots that are densely covered in root hairs. Capillaroid plants such as rushes (Restionaceae) form a mat of root clusters that are densely covered in long root hairs at the soil surface. These roots pump out lots of organic acid exudates that help them take up the nutrients they need.
Some sedges, rushes, and other monocots in Western Australia have sand-binding roots, in which sand literally sticks to the roots and covers them completely. Scientists hypothesize that the attached sand somehow enhances nutrient uptake.

The cluster roots of the red pincushion protea, Leucospermum cordifolium, are not colonized by mycorrhizal fungi.
GENERAL LOCATIONS OF MYCORRHIZAE TYPES
Determining the succession and location of mycorrhizal fungi and plants in the soil can be a chicken-or-egg situation: the type of mycorrhizal fungi present in the soil affects the distribution and diversity of the plants that grow there, but at the same time, the plants may control the local distribution of the fungi by associating or failing to associate with a particular fungal species.
If no mycorrhizal fungi are present in the soil, nonmycorrhizal plants will grow there. Eventually, facultative mycorrhizal fungi, which are parasites or saprobes, will move in. Semidependent host plants will then become established and will eventually take over. As decay increases, obligatory mycorrhizal fungi will colonize the area, along with completely dependent host plants.
Studies have shown that, in response to the presence of mycorrhizal fungi, plant diversity usually increases. Studying changes in the type of mycorrhizae in a particular area can tell scientists which way the area is headed. Nevertheless, it is clear that the presence of a particular mycorrhizal fungus influences the ability of particular plants to live in particular soils. Of course, the ability of mycorrhizal fungi to change the environment means that some plants may slow down or cease to grow there. So it goes both ways.
Mycorrhizal fungi can also be grouped based on the geographic location in which they grow, which in turn is based on soil type, the availability of nutrients, and the prevailing climatic conditions. For example, some particular ectomycorrhizal tree hosts grow only in particular areas, and that is where you will find their associated mycorrhizal fungi.
Soils in subarctic regions have extremely low pH levels. Peat dominates there, as do ericoid mycorrhizae. Ascomycete fungal partners penetrate the root’s cortex cells, but they don’t send much mass into the soil. They do, however, mineralize nutrients near the roots that the host plant could never access alone, producing the inorganic ions needed by the host. Some tundra mycorrhizal fungi provide organic nitrogen directly to their host, a feat that harkens back to the days of the Humus Theory (which held that plants ate organic matter) and bucks the norm of taking up inorganic forms of nitrogen. (If this process could be translated into agricultural plants, it could change the way the world is fed.)
In conifer forests and hardwood forests, with warmer temperatures, higher pH, and more available nutrients, ectomycorrhizal fungi and their associations predominate. These fungal hyphae send out lots of hyphal mass, which not only brings back lots of nutrients to the hosts but also mineralizes a tremendous amount of organic material, making it available to other plants and fungi.
In temperate grasslands with warmer temperatures, where the soil pH is even higher and nitrates are the predominant form of nitrogen, arbuscular mycorrhizae prevail. The huge increase in surface area available to the system as a result of mycorrhizae enables the host plants to meet their nutrient needs.
MYCORRHIZAL BEHAVIOR
Scientists have demonstrated that nutrients are shared throughout mycelial networks, both between different and among the same plant species. In fact, several different mycelial networks can fuse together and act as one. A mother tree can thus support other trees throughout the forest, preferentially distributing more nutrients to the younger ones. Cooperation exists between different plants’ mycelial networks. This is amazing when you consider that each plant may also contain substances that can inhibit the growth of another.
It is clear from genome sequencing that mycorrhizal fungi contain special genetic expressions that cause them to function as they do. Some genes affect nutrient uptake, others manage the production of signaling proteins used in controlling the delicate dance between mycorrhizal symbionts, and others help the fungi reach the plant without triggering its defense mechanisms. These subtle genetic differences, however, may result from DNA exchange with other mycorrhizal fungi or with other organisms, such as bacteria, in the mycorrhizosphere or the fungal spore. Perhaps this occurrence depends on the conditions to which the spore is exposed and in which the fungus will need to live.
Knowing how mycelial networks function and figuring out how we can duplicate their operation could be a major advance, especially with regard to food crop production. Scientists have discovered that microorganisms exchange genetic material. They can create maps of the genetics of particular mycorrhizal fungi and study these in relation to a whole range of biological influences. This new science, transcriptomics, enables scientists to identify mycorrhizal strains that carry specific traits that can benefit a specific crop or plant. We still have a lot to learn, and perhaps we will someday use all this knowledge to benefit humankind.
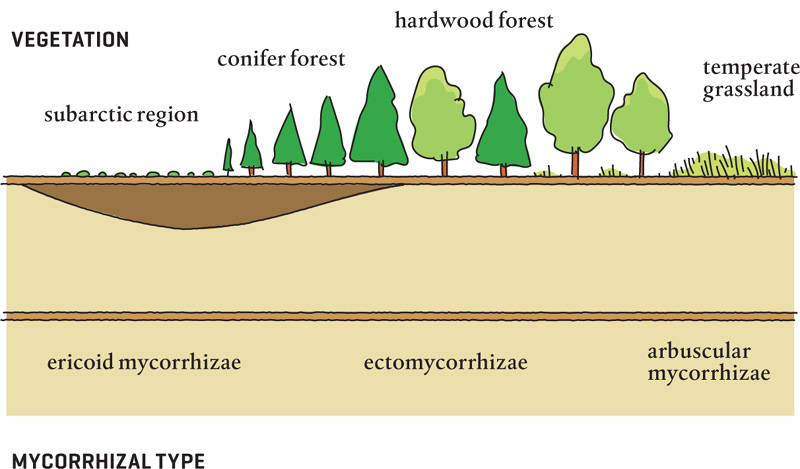
Locations of vegetation and corresponding mycorrhizal types.
 Mycorrhizal is the adjective used to describe the fungus or fungi, as in mycorrhizal fungi. The term has nothing to do with roots or the relationship of fungus to root.
Mycorrhizal is the adjective used to describe the fungus or fungi, as in mycorrhizal fungi. The term has nothing to do with roots or the relationship of fungus to root.