3
the senses and the nervous system
Insects are bombarded by the same stimuli that we are, but their sensory and nervous systems work very differently. From the multifaceted structure of their eyes to the taste buds in their feet and their whole-body brains, insects experience and respond to the world in ways quite alien to us.
3.4 • Hearing, touch, and more
3.5 • Brains, ganglia, and nerves

The wasp’s large, prominent eyes and antennae provide it with a constant stream of sensory data about the world around it.
eyes
Insect eye anatomy is dramatically different from our own, but the size and complexity of some insects’ eyes leave us in no doubt as to how well-developed their visual abilities are.
Animals that fly in the open in daylight, as many adult insects do, need well-developed, fast-reacting vision to navigate their environment. This is at its most extreme in the dragonflies, which need not only to evade obstacles and larger predators, but also to pursue and capture other flying insects at high speed. These insects have immense, “wraparound” eyes, which occupy almost the entirety of the outsides of their heads. Flower-feeding insects such as bees and hoverflies also have very large eyes.
The primary visual organs of an adult insect are compound eyes. Under the microscope, the surface of the eye is revealed to be a tight arrangement of hexagons. Each hexagon is the surface of an individual light-sensing structure—the ommatidium. A large dragonfly has about 30,000 ommatidia per eye. Ommatidia are about ten times longer than they are wide, and narrow considerably from their surface to their base.

The eyes of a dragonfly wraparound nearly the whole of its head for almost 360-degree vision.
Each ommatidium is topped with a cornea and a crystalline pseudocone, which acts as a focusing lens, directing light into the rhabdom, a long, narrow, and transparent structure at the center of the ommatidium. It contains photosensitive pigments that respond to certain wavelengths of light. The rhabdom is formed by the combined inner parts of (usually) eight specialized nerve cells—photoreceptors. When the rhabdom’s pigments undergo chemical change in response to light, these cells send a nerve impulse to the brain.
The ommatidium also contains six pigment cells, which absorb light that strikes the cornea at an indirect angle. This ensures that the photoreceptors only receive light that passes through the cornea directly. Insects cannot move their eyes to follow a point of interest, but the pigment cells provide an alternative means of retaining focus.
What the compound eye “sees” is, as far as we can tell, a scene formed by an array of colored specks (including, in some cases, ultraviolet “color”), each speck contributed by an individual ommatidium. In dragonflies, there are enough specks to form a detailed picture, but in insects with fewer ommatidia the compound image has little detail.
ocelli
In addition to the compound eyes, many adult insects have secondary visual structures called ocelli, and these are the only visual organs in most insect larvae. They are noticeable on the heads of bees and wasps, for example, as a trio of small, shiny bumps in the space between the compound eyes. The ocelli have a cornea and a layer of photoreceptive cells, but unlike those in the compound eye, these cells only sense light levels, not color. They are thought to have an important role in maintaining stability in the air in fast-flying insects.

This Ashy Mining Bee’s ocelli, on the upper part of the head in between the two large compound eyes, resemble small glossy beads.
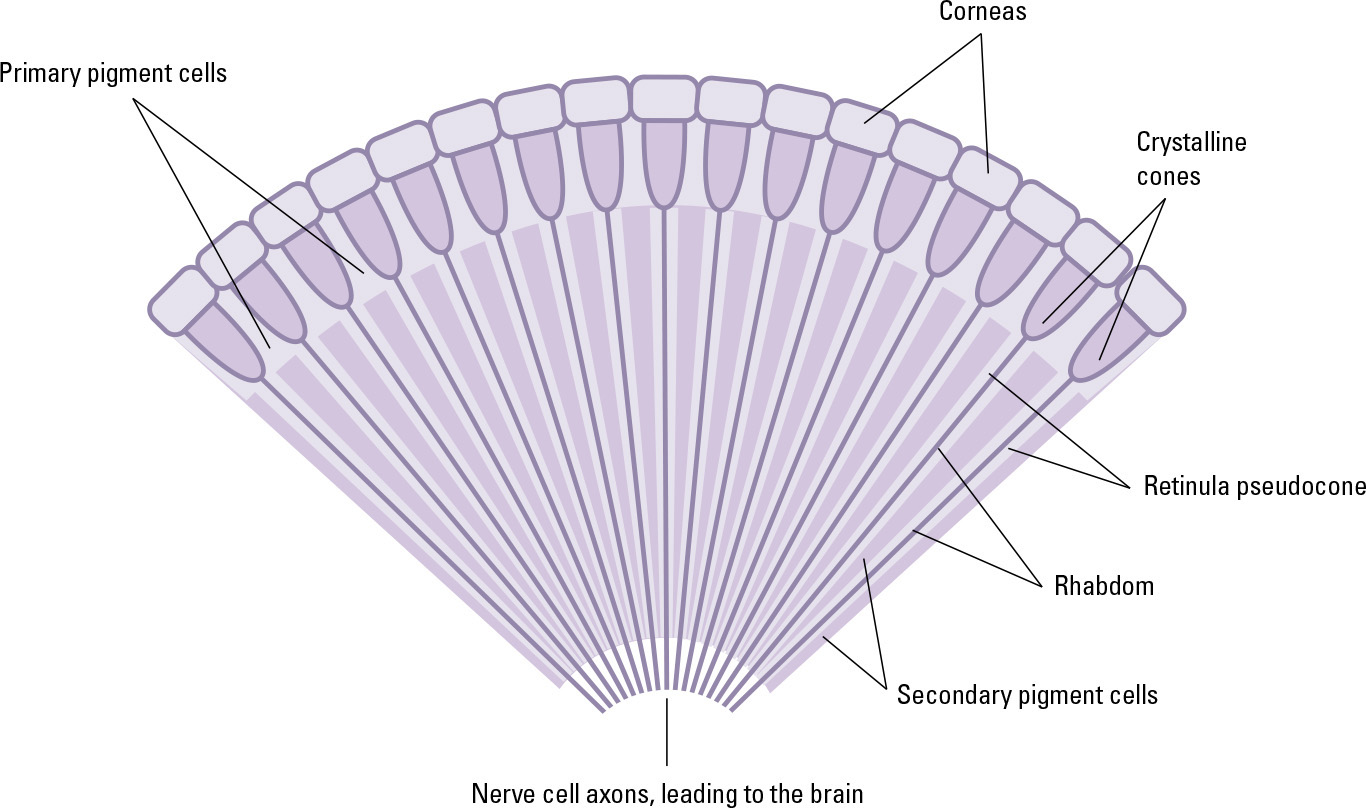
A cross-section through part of an insect’s compound eye.
antennae
The paired antennae, or feelers, are, after the compound eyes, the most noticeable sensory structures in insects. They are highly diverse in form and also in function.
The antennae are the appendages that develop from the second segment of the insect head. Like the legs, they have a segmented structure, which can give them great mobility. The constant antenna movement of a longhorn beetle as it walks across a flower-head shows how actively it uses these appendages to collect information about its environment.
Each antenna has three main sections. The base segment is the scape, which is small and sits within a sclerotized ring projecting from the head capsule. This connection is springy and flexible, allowing free movement of the antenna as a single unit. The second segment is the pedicel—this is usually relatively long and attached by a hinge joint to the outer section, the flagellum, which is often formed by many small segments or flagellomeres. The flagellomeres contain odor-detecting cells, and the joints between them permit this outer part of the antenna to move with considerable flexibility, allowing the insect to pinpoint the source of a scent. The pedicel contains a bundle of sensory cells (the Johnston’s organ) that responds to involuntary movement of the flagellum. It can function as an organ of hearing, sensing air vibrations produced by sound, and its input can also help the insect maintain control of stability during aerial twists and turns.

Some impressive antennae: Black Dancer Caddisfly

Gypsy Moth

Alpine Longhorn Beetle.
Antenna shape varies from simple and straight to bulbous-ended, feather-like (plumose), serrated, or tipped with a fan of blades. The plumose antennae of certain male moths are very large, enabling them to sense molecules of the female’s airborne pheromones at extremely low concentrations (and to react to them with remarkable speed).

Male fairy longhorn moths gather on sunlit vegetation and perform short display flights to attract females—their long white-tipped antennae wafting about and catching the light.

Direct contact between the antennae is a precursor to mating in many butterfly species.
social signaling
Some insects use their antennae to convey messages to other insects. The extraordinarily long, white-tipped antennae of fairy longhorn moths (family Adelidae), for example, dance eye-catchingly in the air as the males perform their communal courtship display dances. The presence of certain chemical compounds on ants’ antennae enables them to recognize whether other individuals they encounter are from their own nest or not.
chemoreception
The senses we experience as smell and taste are both classed as chemoreception, as they involve sensing particular chemical compounds through direct contact of that compound’s molecules with our sensory cells.
Although they do not have nostrils or tongues laden with taste buds, most insects are still highly sensitive to smells and tastes. Without these senses, they would have difficulty finding the right food, detecting mating partners, and locating suitable places to take shelter either for hibernation or to lay their eggs.
The organs of chemoreception are called sensilla. These small structures are located on the outside of the cuticle, and consist of one or a few receptor cells plus a few accessory cells, which moisten and protect the receptor cells, as well as the endings of the nerve cells that communicate with the receptor cells. When a molecule of a particular compound makes contact with a receptor cell, it binds to a certain part of that cell’s membrane, causing a chemical change within the cell. This in turn stimulates the nerve cell that is in contact with the receptor cell, sending a nerve impulse toward the brain.
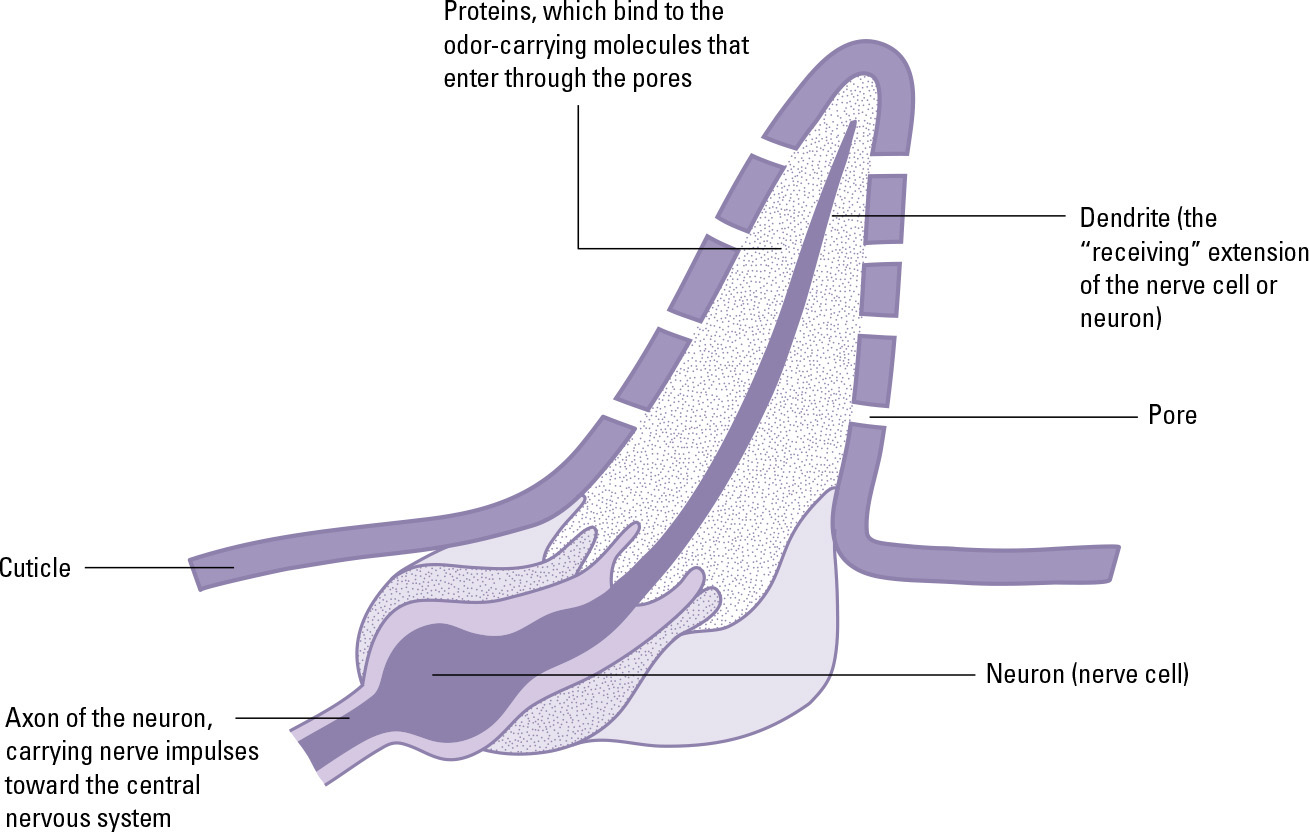
A chemoreceptive sensillum in an insect’s outer covering: Molecules of a chemoactive substance enter through pores in the sensillum’s cuticle, and bond to receptors on the dendrite of the nerve cell that extends into the sensillum.
The antennae are the key organs of chemoreception, their sensilla being sensitive to smell (picking up airborne molecules of volatile compounds) and in some cases also taste (picking up the same kinds of molecules through direct contact with their source). However, most insects primarily taste through their mouthparts. Moths, butterflies, and true flies can also taste through sensilla in their tarsi (the outermost part of their legs), and some insects have taste receptors on their ovipositors (egg-laying tubes). Females with eggs to lay use the taste sensors in their feet or ovipositors to locate suitable leaves or other food sources for their future larvae. Insect larvae also have a well-developed sense of taste—finding food is a much more dominant driver in their lives than it is for adults, and in herbivorous insect larvae in particular, the range of foods that their bodies can process may be very narrow.

A female Monarch Butterfly uses chemoreceptors on her feet to assess the suitability of a leaf before she lays her eggs on it.
filtration
Because there are so many volatile chemical molecules around in the air and on all kinds of substrates, the insect needs to be able to ignore all of those that are not relevant to its interests. Its chemoreceptors are therefore highly specific to particular molecule types. However, potentially dangerous substances (such as ammonia or strong acids) seem to trigger a strong avoidance response across all species of insects, even if all their known chemoreceptors have been temporarily blocked. The pathways of this common chemical response are not yet known. Research into insects’ behavioral responses to different chemicals is important when attempting to control insect species that carry deadly diseases (for example, mosquitoes in areas where malaria is common).

Wasps have an unerring ability to sense and home in on sweet-tasting substances.

Mantises are able to learn to avoid attacking prey that has a bitter taste.
hearing, touch, and more
Vision and chemoreception are vitally important to most insects, but they have other sensory inputs that are valuable, too, including some that seem quite alien to us.

Worker ants are in constant communication with each other, through chemical and touch receptors on the antennae and other body parts.
Because insects usually seem entirely unresponsive to the sounds we make, it is easy to assume that they have no appreciable sense of hearing. However, noisy insects such as grasshoppers, crickets, and cicadas clearly do communicate through sound. Other ways that sound is important to insects are often lost on us, as they involve frequencies outside our range of hearing—for example, some moths can hear the echolocation calls of hunting bats.
The anatomical structure of hearing is the tympanal organ. It comprises a thin membrane stretched across an air-filled space, with sensory cells below. Sounds create pressure waves in the air, which vibrate the membrane. The air within is moved by this vibration, and this movement triggers the sensory cells. Tympanal organs may be found on almost any part of the body.
Insects sense touch through sensilla similar to those involved in chemoreception. However, the receptor cells in touch sensilla respond to contact, movement, and pressure. They can be found on all body parts. Touch sensilla often take the form of long, fine hairs, but other types include stretch receptors (which sense, for example, when it is time to stop feeding) and water-pressure receptors (which keep aquatic insects from diving too deep).
other senses
Some insects are also sensitive to Earth’s magnetic field, and to electrical fields. Honeybees use geomagnetic information to help them navigate directly back to their hive after a long and convoluted foraging flight. A foraging bumblebee generates a small electrical field with its wingbeats, which briefly alters the electrical field of each flower that it visits. When another bumblebee comes along, it can sense this altered state, and will avoid that flower as it “knows” that the nectar has already been taken. How insects can sense magnetic and electrical fields is still the subject of debate and research.

Bumblebees use their sensitivity to electrical fields to make decisions about which flowers to visit.

A touch-sensitive sensillum in an insect’s cuticle: When it is moved by an external force, the resultant stretching of its socket stimulates the nerve cell that extends into it.
brains, ganglia, and nerves
The input received from the insect’s sense organs needs to be processed before the insect can “decide” an appropriate behavioral response. This is the job of its central nervous system.
The insect brain, like ours, is essentially a concentration of neurons, or nerve cells, which communicate with each other and with nerves that reach out into the rest of the body. It receives input along afferent nerves (those whose impulses head toward the central nervous system) from the sensory organs, and responds by sending out signals via efferent nerves (those whose impulses head out from the central nervous system) to the muscles, to trigger the appropriate physical movements. The efferent nerves also communicate with glands, causing them to release hormones in response to certain stimuli.
The brain is located in the back of the head, and has three distinct lobes, each region associated with the structures that arise from different head segments: the protocerebrum at the front (which is linked to the visual system), the deutocerebrum in the middle (which is linked with the antennae), and the tritocerebrum at the back (which communicates with the mouthparts).
These three regions are each formed from a pair of ganglia—a bundle of nerve endings. Though in the brain the ganglia are fused, the insect’s body contains many more, separate ganglia, one pair in each body-segment as a general rule (though in some cases they are fused with neighboring ganglia). In a sense, therefore, each body-segment has its own miniature “brain,” handling its own affairs—for example, each of the three segments of the thorax contains a pair of ganglia that control movement of that segment’s pair of legs.
neurons
A neuron is a cell with one or more long, filamentous offshoots (axons), which can conduct electrical impulses. An axon ends with a cluster of very fine, branching projections (dendrites), which are separated from the dendrites of nearby neurons by tiny gaps (synapses). When the electrical impulse reaches the dendrites, it causes them to release a special chemical (neurotransmitter), which crosses the synapse and binds to the cell membranes on the next neuron’s dendrites, sparking a new electrical impulse. In this way, an impulse can travel between the brain or a ganglion to the farthest extremities of the body.
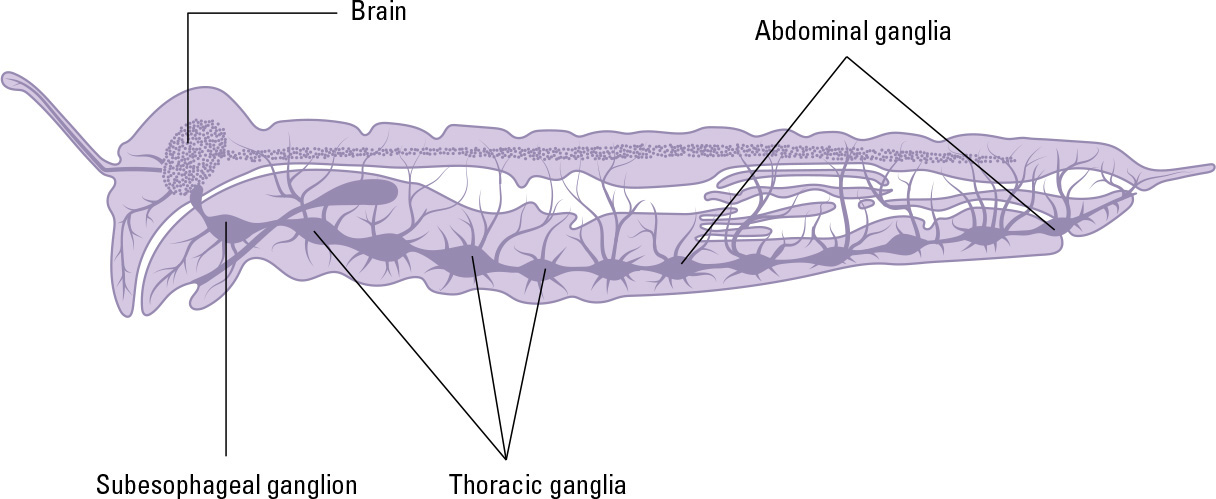
Each body-segment of an insect has its own governing ganglion, while the brain is formed by fused ganglia in the head segments.
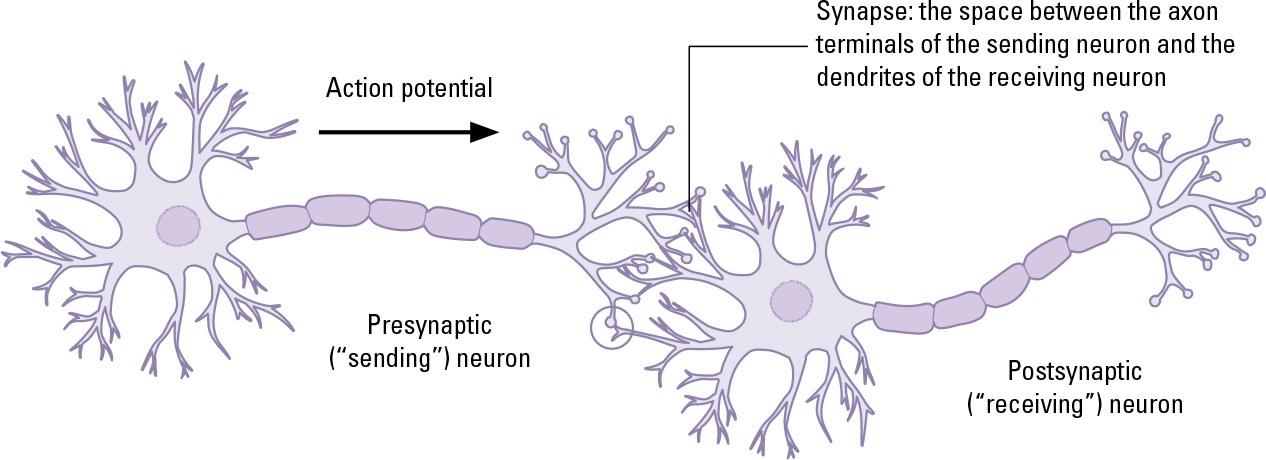
A neuron is adapted to carry an electrical signal through the body and pass it on to adjoining neurons.
Nerves that leave (or arrive at) the brain and ganglia are thicker, formed from bundles of many neurons. In more peripheral body parts they become thinner, containing fewer neurons. Each sensillum may be innervated by just two or three individual neurons.

An ant’s brain, though obviously tiny, can make up as much as 15 percent of its total body-weight.
insect intelligence
With a brain the size of a sesame seed, a bee seems an unlikely candidate for exceptional intelligence. Yet bees and other insects can perform complex behaviors, suggesting brainpower beyond their size.

The degree of organization and cooperation required for these ants to form a living bridge for their comrades suggests real intelligence.
Modern research on various animals, particularly birds, has overturned our traditional view that brain size is the only important factor that determines intelligence. A raven and a chimpanzee show comparable intelligence, because the raven’s ½-ounce (15g) brain has a much higher density of neurons than the chimp’s, and a more efficient structure generally. What about insects, though? Their brains are vastly different from those of vertebrates, and are relatively little-studied.
Defining intelligence is scarcely straightforward. However, most biologists agree that its elements include memory, innovative behavior, ability to learn, planning, problem solving, and social interaction (including teamwork). All of these can be observed in certain insects, but probably most of all in the social species, such as bees, wasps, and ants.
Honeybees have to learn how to reach the nectaries of various flower types, but they can also be taught skills that clearly are not essential to their survival, such as recognizing different human faces. The famous figure-eight “waggle dance,” performed in the colony to tell other bees where to go for new nectar sources, conveys information about direction, distance, and even quality of the nectar available. This use of symbolism to express concepts is a sort of language, the like of which is not known in many apparently “higher” organisms.
Ants’ social structure can show great complexity. Some species are “slave-makers,” stealing ant pupae from other species’ nests to replenish their own workforce. They are capable of cooperative behavior, such as forming living bridges to allow others to traverse gaps, and working together to carry large prey items back to the nest.

Intelligent behavior is most apparent in social insects like bees, and predators like dragonflies.
predatory minds
Carnivores are often smarter than herbivores, because it takes more brainpower to capture an animal than it does to find and eat a plant. Dragonflies have been shown to be capable of predicting the flight path of the insect they are chasing, and are able to ignore all distractions (including other insects, when their target is in a swarm) as they home in, making a successful strike more than 90 percent of the time. Curiosity, another aspect of intelligence, is also apparent when a dragonfly hunts—it will even pause midair to examine you before moving on.

To chase down living, fast-moving prey takes a finely adapted nervous system.
INSECT BUILDERS
From the exquisitely molded little mud nest of a potter wasp to the towering monolith that houses a colony of termites, the insect world includes many master builders.
Most of the insects that construct shelters, either for themselves or for their young, are members of the order Hymenoptera (bees, wasps, sawflies, and ants). Wasps of the genus Eumenes are among those that craft particularly pleasing structures from mud, which, when complete, resemble dainty vases with a large bowl narrowing to a small entrance passage. The female wasp collects mud, which she sticks to a substrate (often a high vertical surface such as a wall or tree trunk, for protection). She sculpts and manipulates her building material with her mouthparts and forelegs, using her saliva for moisture where required. When it is complete, she provisions it with some paralyzed prey, lays an egg inside, and may then seal up the entrance. The egg hatches and the larva lives on its food store, pupating inside and then breaking out after it emerges from its pupa.
Termite mounds may be tall and narrow, or wide with multiple humps. They include shafts or “chimneys” for ventilation, and a maze of tunnels and chambers inside, leading to the main nest chamber in the ground beneath, where the king and queen termite live and breed. They are built and maintained by worker-caste termites, while the larger soldier caste defends the nest from invaders. As the mounds are built from a considerable amount of termite droppings, they encourage a different and often richer plant community than the surrounding terrain. Clusters of mounds in close proximity effectively create different biomes, and greatly increase biodiversity.

Social insects make their nests from materials such as paper from chewed wood (top: wasp’s nest) and the earth (bottom: termite mound).

Some moth caterpillars live socially in large “tents” made from silk that they produce from facial glands.

Depending on the materials they use, caddisfly larval cases can be quite ornate in appearance.
The little-known tropical insect order Embioptera—the webspinners—live permanently in aggregations within webs spun from glands on their feet. Only the adult males develop wings and disperse from the colonies.
Some insects build miniature shelters just for themselves. The most famous of these are the larvae of caddisflies. These delicate-bodied, slow-moving larvae are aquatic and feed on detritus, so protection is more important than speed. They form their cases from tiny fragments of gravel, twigs, sand, and other materials they find on the lake bed, spun together with silk.
Webs
It is not unusual in some areas to come across a shrub or hedgerow that is completely draped in what look like spider webs. In fact, these large silk webs are the work of young moth caterpillars, such as the ermine moths (genus Yponomeuta). The young larvae spin their web out of silk secreted from their mouthparts, and under its shelter they can feed on the plant’s foliage in relative safety. Some other insect larvae use silk to make individual shelters. Moths of the family Tortricidae are sometimes known as leaf rollers, as they roll up and secure a section of a leaf of their food plant and feed within this makeshift shelter.