Habitability as a Tool in Astrobiological Exploration
Abstract
While “follow the water” has been an extraordinarily effective strategy in the search for habitable environments in the solar system, astrobiologists will require a more refined approach to exploration during the next phase of exploration when we will begin to target specific habitable sites for deploying a new generation of in situ life detection experiments. Identifying the best sites for in situ experiments or for selecting samples for return to Earth will likely require a more refined knowledge of past and/or present aqueous environments, with an ability to detect HZs at the microscale. Success in detecting extraterrestrial life may require a spatially integrated sampling strategy that includes the ability to measure multiple microenvironmental factors at each study site. Such microscale, multidimensional approaches have been employed in the past by microbial ecologists to refine and quantify biological concepts like the niche. Such approaches may prove useful in more effectively conveying the concept of habitability.
Keywords
Habitability; Astrobiology; Extremophiles; Solar system exploration
1.1 Overview
The exploration strategy currently guiding the search for life in the solar system begins with a short list of the fundamental resources required by terrestrial life. The most important of these requirements is liquid water. Second in importance are sources of biologically essential elements, the so-called CHNOPS elements (required for all living systems), plus a dozen or so transition metals that fulfill important roles in coenzyme functions in cells. Third, life requires sources of energy, obtained through chemical redox reactions. This simple, three-pronged strategy is usually embodied in the phrase “follow the water.” This approach to habitability has served astrobiology well, with recent missions reporting evidence for past and present water on Mars and in the subsurface of tidally heated icy moons (e.g., Enceladus and Europa). While the “follow the water” strategy has proved successful in discovering potentially habitable zones (HZs) of liquid water in these and potentially other extraterrestrial environments, understanding the potential for life to actually develop and persist in these places remains unclear. On Earth, habitability depends on the coexistence of three things: liquid water, energy sources and chemical building blocks. However, it is also clear that, on Earth, life exists within a complex web of ecological interactions that, through evolution, have continually reshaped the origin, nature, and distribution of species. Stated differently, life on Earth is a powerful ecological force that itself shapes habitability.
As a testimonial to the power of eco-evolutionary forces to shape the history of the biosphere, discoveries of extremophiles have revealed that terrestrial life occupies a much broader range of environmental extremes than once thought possible. On planetary surfaces, organisms have evolved metabolic strategies that extract energy from sunlight, over an impressive range of environmental conditions (e.g., temperature, pH, and water activity). Particularly impactful, however, are life forms that do not require sunlight but rather subsist on chemical energy from their surrounding environment. These “chemotrophic” microorganisms are part of an extensive subsurface biosphere that resides in deep subsurface habitats on the Earth. Of particular interest for astrobiological exploration are “chemolithoautotrophic” microorganisms that obtain energy from the chemical by-products of aqueous weathering of mafic crustal rocks. These microbes require no connection to the surface and can exist unseen from surface exploration.
While “follow the water” has been an extraordinarily effective strategy in the search for habitable environments in the solar system, astrobiologists will require a more refined approach to exploration during the next phase of exploration when we will begin to target specific habitable sites for deploying a new generation of in situ life detection experiments. Identifying the best sites for in situ experiments or for selecting samples for return to Earth will likely require a more refined knowledge of past and/or present aqueous environments, with an ability to detect HZs at the microscale. Success in detecting extraterrestrial life may require a spatially integrated sampling strategy that includes the ability to measure multiple microenvironmental factors at each study site. Such microscale, multidimensional approaches have been employed in the past by microbial ecologists to refine and quantify biological concepts like the niche. Such approaches may prove useful in more effectively conveying the concept of habitability.
1.2 Introduction
The concept of “habitability” has emerged as a core principle in NASA's exploration strategy to search for signs of extraterrestrial life in the solar system and beyond. Goal 1 of NASA's Astrobiology Roadmap seeks to “understand the nature and distribution of habitable environments in the universe” (Des Marais et al., 2008). The general concept of habitability has been used widely to identify potential sites for future astrobiology missions. However, definitions of the term are typically very generalized and in some instances downright confusing. Looking beyond simple strategies based on the presence of liquid water, elemental building blocks, and energy sources, at a deeper level, there are environmental factors that, in combination, clearly challenge our assessments of potential habitability. The purpose of this chapter is to (1) broadly review the concept of habitability, particularly as it relates to astrobiology and exploration strategies for extraterrestrial life, and (2) illustrate how the concept of the ecological niche, originally developed by Hutchinson (1957), may provide a multivariate statistical approaches with the potential to contribute more broadly based discussions of habitability.
1.3 Defining Habitability
Habitability has been defined as the capability of an environment to support life. Often, the term is used to target specific locations (“habitable zones”) within the solar system, on other planets, or moons where extraterrestrial life may be present. The potential for habitability is presently based on comparisons with the known requirements for life on Earth.
1.3.1 Follow the Water
Arguments for the habitability of other planetary bodies in the solar system have focused on the inferred presence of liquid water at the surface, or within the subsurface of a planet or moon. Because all living organisms require liquid water for survival and growth, it is often referred to as a universal requirement of life. Given the close relationship between liquid water and life, it is not surprising that the search for water, in all its forms (liquid, ice, or vapor), has provided a consistent strategic focus in NASA's search for past or present HZs in the solar system. This is reflected in the widely embraced mantra “follow the water.” Indeed, the effectiveness of this strategy is evident in recent discoveries of water (past and present) by NASA's missions to Mars (e.g., Squyres et al., 2004; Ruff et al., 2011; Grotzinger et al., 2013) and the outer solar system icy moons, Europa (e.g., Roth et al., 2013; Lowell and DuBosse, 2005) and Enceladus (Meyer and Wisdom, 2007; McKay et al., 2014; Glein et al., 2015).
But why water? Water is often identified as a “universal solvent.” In large part, this traces to the basic dipolar structure of the water molecule and its ability to form hydrogen bonds with other water molecules and/or cations/anions in solution. In addition to water's extraordinary solvent properties, it easily outcompetes other potential biological solvents in its combination of biologically favorable physical and chemical properties (Table 1.1; Plaxco and Gross, 2011). Water remains liquid over a broad temperature range and has a high molar density, heat capacity, and dielectric constant, all of which can favor habitability. Given its properties, it is perhaps not surprising that liquid water is the required medium for carrying out all of the basic cellular functions of organisms (e.g., energy transduction, reproduction, and locomotion).
Table 1.1
| Physical properties of potential biological solvents | |||||
|---|---|---|---|---|---|
| Solvent | Formula | Liquid range (°C at 1 atm) | Molar density (mol/L) | Heat capacity (cal/g °C) | Dielectric constant |
| Water | H2O | From 0 to + 100 | 55.5 | 1.0 | 80 |
| Hydrogen fluoride | HF | From − 83 to + 19 | 48.0 | 0.8 | 84 |
| Ammonia | NH3 | From − 78 to − 34 | 40.0 | 1.1 | 25 |
| Hydrogen sulfide | H2S | From − 85 to − 6 | 26.8 | 0.5 | 9 |
| Methane | CH3 | From − 182 to − 161 | 26.4 | 0.7 | 25 |
| Hydrogen | H2 | From − 259 to − 253 | 35.0 | 0.002 | 1 |
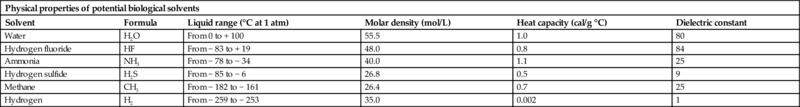
From Plaxco, K.W., Michael, G., 2011. Astrobiology: A Brief Introduction, second ed. Johns Hopkins University Press, Baltimore. ISBN 978-1-4214-0096-9.
However, the recent discovery of hydrocarbon lakes on Titan (Mitri et al., 2007) highlights the importance of considering the potential for alternative solvents for life. A comparison of some of the basic physical and chemical properties of water and other solvents (see Table 1.1) places this in perspective. While a strong case has been made for water as the solvent for life, it is not the only possibility. Some forms of life may have followed different evolutionary pathways involving other hydrocarbon-based solvent systems (e.g., HF or NH3). It seems prudent to keep this in mind as we think about habitability.
Another property that is important to consider with regard to habitability is the chemical activity of water (Aw). This provides a measure of water's availability to carry out chemical reactions in aqueous solutions. Aw provides a thermodynamic measure of salinity, another variable of common interest in planetary exploration. Aw values range from 0.0 to 1.0, where 1.0 (the value for pure water) indicates that water is 100% available to host reactions. An Aw value of 0.0 indicates that water is unavailable. With minor exceptions, Aw values for life on Earth fall between 1.0 and 0.6 (Beuchat, 1983). Aw has the potential to provide a more sensitive indicator of habitability than just “follow the water” by addressing the specific composition of the water (e.g., hypersaline brine vs fresh water). This was shown in geochemical modeling of Martian water activities by Tosca et al. (2008). This study revealed how the salinity of the water at Meridiani Planum may have exceeded the limit for terrestrial life based on the range of Aw values obtained using models that were constrained by independent in situ measurements of Martian mineralogy.
Astrobiologists are also interested in the search for liquid water in far-flung places beyond the solar system. Astrophysicist Su-Shu Huang (1960) was the first to suggest the presence of an orbital zone around sun-like, main-sequence stars where planetary surface environments are likely to meet two fundamental requirements for the long-term habitability of a planet. These requirements include a sustained energy source (the central star), coexisting with liquid water on the surface of the planet. This orbital region where liquid water is stable at the surface of a planet is referred to as the HZ (Kasting et al., 1993). It is notable that the HZ has also been applied in searching for habitable extrasolar planets in orbit around other stars in the nearby galaxy (Lammer et al., 2016). Near the end of 2013, the Kepler mission team announced that there could be as many as 40 billion Earth-sized planets orbiting within HZs of sun-like stars and red dwarfs within the Milky Way Galaxy.
The discovery of subsurface zones of liquid water within the interiors of some outer solar system icy moons, maintained by internal frictional heating from tidal flexing (Peale, 2003), has further broadened our view of the HZ to include icy moons of gas giants in the outer solar system (Williams et al., 1997). And with the discovery of a deep, hot biosphere on Earth (Gold, 1999; Farmer, 2000), populated by subsurface thermophilic chemotrophs, an even broader shift in thinking has been required by astrobiologists. This has opened up potentially large subsurface zones of liquid water within the interiors of terrestrial planets and moons. More recently, the interior HZ concept has been expanded further to include subsurface hydrothermal habitats populated by subsurface chemolithoautotrophic organisms that obtain energy by fixing carbon dioxide (CO2) released by the aqueous weathering of mafic crustal rocks. These organisms convert the CO2 derived from aqueous weathering to energy-storing glucose (Kuenen, 2009). The impact of these discoveries on potential habitability is noteworthy (Chapelle et al., 2002) and indicates that such ecosystems may survive independently of surface energy sources, with the potential to thrive within extensive, stable subsurface environments that are basically undetected by orbiting spacecraft.
1.3.2 Follow the Bioessential Elements
We now know of many potential destinations in our solar system where zones of liquid water likely exist to support life. And while liquid water is usually regarded as the most crucial requirement of living systems, as noted above, life also requires sources of elements for the synthesis of complex macromolecules that comprise the basic building blocks of living systems. Almost 99% of the human body consists of six elements: oxygen, carbon, hydrogen, nitrogen, calcium, and phosphorus (i.e., the so-called “CHNOPS” elements). In the human body, < 1% is accounted for by just five elements: potassium, sulfur, sodium, chlorine, and magnesium. The remaining elements are present in trace amounts. These trace elements are mainly transition metals that include V, Cr, Mo, Mn, Fe, Co, Ni, Cu, Zn, and Se. Despite their low abundance, these trace elements are considered to be micronutrients that fill essential cellular functions, mainly as components of coenzymes.
While they are essential for life, the CHNOPS elements also fulfill many nonbiological roles. Thus, they may provide fewer constraints for inferring habitability. It can be argued that the bioessential trace elements may actually be more sensitive indicators of habitability because they are micronutrients that may be limiting and may require biological concentration mechanisms for their enrichment in organisms. In other words, the trace metals that fulfill vital roles in living systems, while far less abundant, may be more useful as biological indicators than the CHNOPS elements.
1.3.3 Follow the Energy Sources
Life also requires sources of energy to support basic cellular functions. Energy for metabolism is produced through electron transfers during oxidation-reduction reactions. Redox-based energy systems have been discussed by Nealson and Conrad (1999) who concluded that the extreme metabolic plasticity observed among prokaryotes allows them to exploit essentially every redox couple available on Earth.
Metabolic plasticity in energy production is well illustrated by the bacterium Shewanella putrefaciens (Nealson and Saffarini, 1994). In the absence of oxygen, this species is able to use a number of alternative electron sources for respiration. That terrestrial organisms obtain energy using transduction systems that share many similarities in common suggests that these pathways have been widely conserved in biological evolution (Hoehler et al., 2007; Thauer et al., 1977).
As noted, metabolic flexibility is a general feature of prokaryotes (see Anderson et al., 2014) and is particularly widespread in all low-oxygen environments. Nealson and Conrad (1999) noted the importance of this “metabolic extremophile” in shaping our approaches to the exploration for extraterrestrial life. Certainly, such examples impact our view of potential habitability by opening up alternative environments (e.g., low oxygen) that, until recently, were not believed possible. This illustrates how the discovery of alternative biochemistries on Earth can provide an important reality check when considering the potential of an extraterrestrial environment to support extreme forms of “weird life” (National Research Council, 2007).
1.4 Exploring the Extremes of Life
On Earth, life occupies a broad range of environmental extremes (Table 1.2; see also National Research Council, 2009; Rothschild and Mancinelli, 2001; Plaxco and Gross, 2011). The upper temperature limit for microbial growth is presently reported as ~ 121°C (Kashefi and Lovley, 2003), with a survival temperature of perhaps 130°C. At > 150°C, complex biomolecules are degraded to their basic components. In between, specialized survival mechanisms, such as heat shock proteins, work to stabilize molecular structures (e.g., Trent et al., 1994).
Table 1.2
| Parameter | Classification | Definition | Examples |
|---|---|---|---|
| Temperature | Hyperthermophile Thermophile Psychrophile | > 80°C Growth from 60°C to 80°C Growth < 15°C Active at − 18°C | Archaeal strain 121; 121°C Pyrolobus fumarii; ~ 116°C Synechococcus lividis; ~ 73°C Psychrobacter Himalayan midge |
| pH | Acidophile Alkaliphile | Low pH (< 5) High pH (> 9) | Ferroplasma acidarmanus pH 0 Alkaliphilus transvaalensis, pH 12.5 Natronobacterium; pH 10.5 |
| Salinity | Halophile | 2–5 Molar NaCl | Halobacteriaceae |
| Oxygen tension | Aerobe Microaerophile Anaerobe | Requires O2 Tolerates some O2 Not tolerant of O2 | Bacteria, archaea Neutral pH Fe2 +-oxidizing bacteria Methanogens, SO42 − reducers |
| Dessication | Xerophile | Anhydrobiotic | Lichens, cyanobacteria; arid deserts |
| Radiation | Radiophile | Ionizing radiation to 15 kGy | Deinococcus radiodurans |
| Pressure | Piezophile | Pressure-loving | Obligate strain MT41 |
| Chemical extremes | Gases | Metalotolerant | Cyanidium caldarium |
| Metals | Ferroplasma |
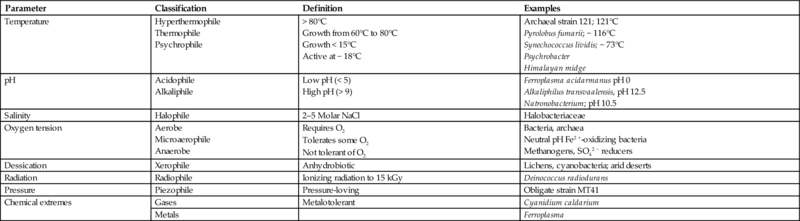
Modified from NAS, 2009.
In moderate-temperature surface environments, where water is lost through high rates of evaporation (e.g., warm playa lakes), the main challenge for life is high salinity and alkalinity. But salinity can also be an important constraint on the habitability of low-temperature environments where interstitial brines form by freeze exclusion. Some groups of psychrophilic halophiles have been shown to grow and reproduce down to − 18°C, well below the freezing point of fresh water. They do this by being halophilic and by living within brine films around grains of rock and soil found within permafrost, or in brine-filled microfractures or fluid inclusions in glacial ice (Mazur, 1984). Psychrophilic organisms have been shown to stabilize bimolecular structures with heat shock proteins that are similar to those employed by hyperthermophiles (Trent et al., 1994). The main problem faced by microorganisms living near the freezing point of such brines is the potential for cell lysis due to the formation of ice crystals (Mazur, 1984).
Even where the minimum requirements for sustaining metabolism, growth, and reproduction are absent, many organisms survive combined extremes of temperature, pH, radiation, and desiccation (including freezing) as endospores, or other resistant structures. In some cases, survival has been documented over extremely long periods of time. Navarro-González et al. (2003) confirmed a lower limit for habitability in the Atacama Desert of Chile where water activities fell below 0.6. Indeed, below this value, living organisms were absent, as well as endospores, suggesting the environmental limits for both cells and endospores had been exceeded.
Life has also been shown to occupy the full range of pH, with the fungus, Ferroplasma acidarmanus, growing at a pH of 0.77, in waters that originate from an acid mine drainage (Bond et al., 2000). At the other end of the pH scale, hypersaline alkaline lakes support microbial alkaliphiles at pH values > 10. Adaptations for extremes in radiation are seen in terrestrial microbes that have evolved extremely rapid DNA repair systems, such as Deinococcus radiodurans (Battista, 1997). This organism lives in association with natural radioactive mineral deposits and on fuel rods in nuclear power plants (Rothschild and Mancinelli, 2001). Rapid repair of DNA basically allows Deinococcus to maintain a working genome.
The observations presented support the following conclusions: terrestrial life is extremely robust and has evolved to occupy a broad range of (as yet unknown) environmental and metabolic limits, seemingly constrained by only the presence of liquid water, sources of biogenic elements, and metabolic energy. This perspective has had a profound impact on our perception of habitability, greatly expanding the scope of environmental possibilities for life on Mars and other planets or moons in our solar system. However, as researchers debate the habitability of the ocean beneath the icy crust of Europa, or in the deep subsurface of Mars where liquid, likely hypersaline water, circulates through sediments and cracks in permeable igneous rocks, the first-order question is whether life is permitted to be present at all. To answer, this question requires more specific studies of habitability, in particular, those that can lead to quantification.
1.5 Niche-Based Multivariate Approach to Habitability
The concept of the ecological niche is rooted in a complex and interesting history (see MacArthur, 1968), with early definitions ranging from the niche as the role played by a species in a community, to the type of environment a species occupies. In 1957, immanent ecologist G.E. Hutchinson formulated a new definition using set theory to quantify the concept of the niche (Hutchinson, 1978). In this new definition, the niche was represented as an n-dimensional hypervolume in multidimensional biospace, occupied by the species being investigated, including the complete range of environmental factors (dimensions) required for successful reproduction of the population (Green, 1971). In this niche model, the n-dimensional hypervolume occupied by the species, including all of its required resources, was deemed the fundamental niche of the species. This approach has achieved popularity among ecologists, primarily because there are so many biologically relevant environmental factors in nature that can be easily measured and quantified (Chase and Leibold, 2003; Austin, 2006).
To further illustrate Hutchinson's niche concept, Fig. 1.1 shows three environmental variables (conditions) displayed in one-, two-, and three-dimensional “biospace.” Each dimension of the fundamental niche represents an independent environmental variable that is required for the growth and survival of the species. The realized niche is the portion of the fundamental niche that is actually occupied by the species in question, whereas the unoccupied part of the hypervolume represents the unrealized portion of the niche (essentially the potential biospace not yet occupied).
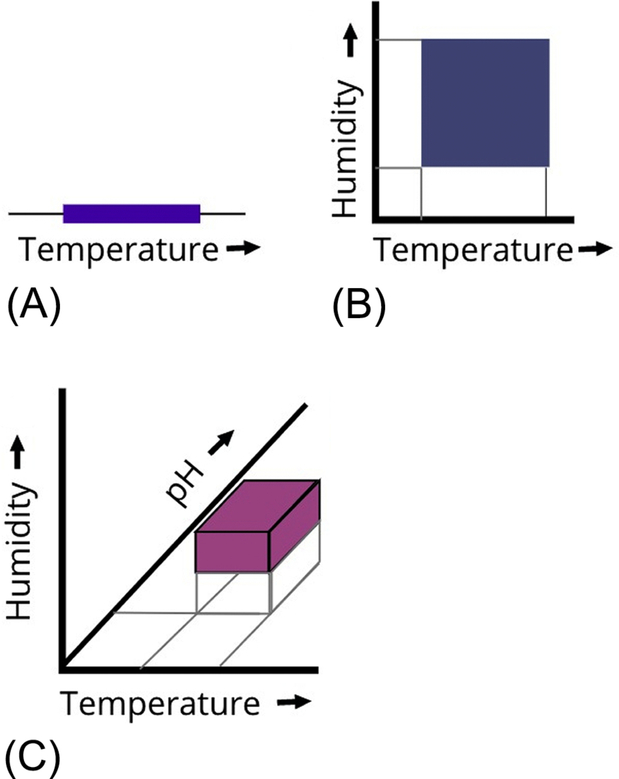
As discussed by Guisan et al. (2006), there have been a number of modifications to the Hutchinsonian niche concept, including the development of multivariate statistical approaches (e.g., multiple discriminate analysis; see Green, 1971). Another example uses habitat suitability models (HSMs; Hirzel and Le Lay, 2008) that relate a set of environmental variables to the likelihood of occurrence of a particular species. Results from HSMs are usually displayed as 2D maps in environmental space that can be quickly compared with the 2D spatial distribution of a species in geographic space. Such comparisons of spatial (geographic) distributions and correlated environmental data can lead to testable hypotheses regarding the nature of species-environment interactions.
Returning to the simplified, unscaled 3D representation of Hutchinson's niche (Fig. 1.1C), we can see how the hypervolume/biospace approach might be repurposed to represent “habitability” space. For example, we could redefine the three axes (conditions) to represent environmental variables, such as temperature, salinity and water activity, and/or spatial dimensions. To expand the analysis to n-dimensional habitability space, we could add the environmental limits for each of the extremophilic species shown in Table 1.1. By including the full range of environmental data for all known microbial species, we could begin to define (qualitatively) the “shape” of n-dimensional habitat space for terrestrial extremophiles. We could then add additional dimensions to the distribution (e.g., radiation tolerance, redox, and hydrostatic pressure). The uncolored part of the boxed region in Fig. 1.1A–C could be redefined as “prospective” habitability space. A multidimensional data set for extremophiles may provide new insights into the nature of habitability and how certain groups (e.g., polyextremophiles) have evolved to occupy certain region's habitability space and how the competition for resources might have occurred, driving evolution. Factors selected for analysis could focus more specifically on key environmental dimensions for extremophiles that hold high scientific interest for understanding the limits of habitability on Earth. This information could be used in strategic mission planning.
1.6 Conclusions
While we acknowledge the recent successes of solar system exploration and extrasolar planet research based on the state and distribution of water, we still lack a system-level understanding of how other factors required by life influence habitability. In articulating a more complete conceptual framework for habitability and one that extends beyond simply “follow the water, elements, and energy,” we need to also consider the importance of biological factors that directly contribute to habitability. It seems clear that astrobiological exploration of the solar system has benefitted greatly from basic discoveries in extremophile research, which have significantly expanded the environmental limits of life on Earth. This work has been foundational for astrobiology in expanding our understanding of where extraterrestrial life could exist and how we might be able to detect it. But next steps are needed to further refine our understanding of habitability. On Earth, species and populations are organized into ecosystems made up of interactive networks of individuals that manage energy flow through various ecological interactions. The success of future life detection experiments could ultimately depend on recognizing spatially integrated ecological interactions between species at the microscale, with the capability to quantify associated processes in situ. Multidimensional ecological models like those developed by Hutchison for the niche could provide useful direction to lead us to more focused strategies for astrobiological exploration, including science-driven technology developments that will allow us to identify and measure dimensions of the environment most critical for life.