List the functions of the parts of the eye, including the cornea, pupil, iris, ciliary body, canal of Schlemm, lens, retina, and sclera
Describe parallel processing
Identify the cell types responsible for color, shape, and motion detection
Recall the structures in the visual pathway:
Vision is a highly adapted sense in human beings. With the ability to sense brightness,
color, shape, and movement, and then to integrate this information to create a cohesive
three-dimensional model of the world, the visual pathways are extremely important
to everyday life. In fact, vision is the only sense to which an entire lobe of the
brain is devoted: the occipital lobe.
Structure and Function of the Eye
The anatomy of the eye is shown in Figure 2.2.
Figure2.2.Anatomy of the Eye
The eye is a specialized organ used to detect light in the form of photons. Most of
the exposed portion of the eye is covered by a thick structural layer known as the
sclera, or the white of the eye. The sclera does not cover the frontmost portion of the
eye, the cornea. The eye is supplied with nutrients by two sets of blood vessels:
the choroidal vessels, a complex intermingling of blood vessels between the sclera and the retina, and
the retinal vessels. The innermost layer of the eye is the retina, which contains the actual photoreceptors that transduce light into electrical information
the brain can process.
When entering the eye, light passes first through the cornea, a clear, domelike window in the front of the eye, which gathers and focuses the
incoming light. The front of the eye is divided into the anterior chamber, which lies in front of the iris, and the posterior chamber between the iris and the lens. The iris, which is the colored part of the eye, is composed of two muscles: the dilator pupillae, which opens the pupil under sympathetic stimulation; and the constrictor pupillae, which constricts the pupil under parasympathetic stimulation. The iris is continuous
with the choroid, as is the ciliary body, which produces the aqueous humor that bathes the front part of the eye before draining into the canal of Schlemm. The lens lies right behind the iris and helps control the refraction of the incoming light.
Contraction of the ciliary muscle, a component of the ciliary body, is under parasympathetic control. As the muscle
contracts, it pulls on the suspensory ligaments and changes the shape of the lens, a phenomenon known as accommodation. Behind the lens lies the vitreous, a transparent gel that supports the retina.
The retina is in the back of the eye and is like a screen consisting of neural elements and
blood vessels. Its function is to convert incoming photons of light to electrical
signals. It is actually considered part of the central nervous system and develops
as an outgrowth of brain tissue. The duplexity or duplicity theory of vision states that the retina contains two kinds of photoreceptors: those specialized for
light-and-dark detection
and those specialized for color detection.
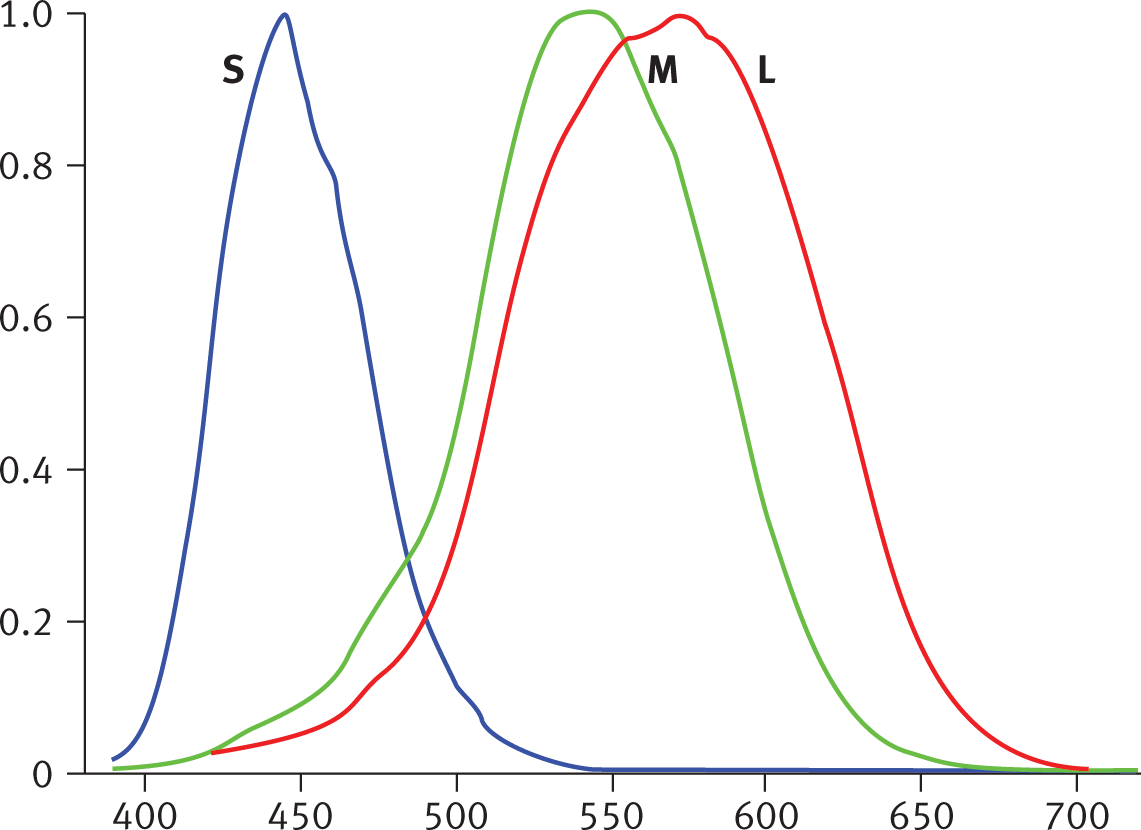
The retina is made up of approximately 6 million cones and 120 million rods. Cones are used for color vision and to sense fine details. Cones are most effective in
bright light and come in three forms, which are named for the wavelengths of light
they best absorb, as shown in Figure 2.3.
Figure2.3.Relative Absorption of the Three Types of Cones at Different WavelengthsThe cones are named for the wavelengths at which they have highest light absorption:
short (S, also called blue), medium (M, green), and long (L, red).
In reduced illumination, rods are more functional and only allow sensation of light and dark because they all contain
a single pigment called rhodopsin. Rods have low sensitivity to details and are not involved in color vision, but permit
night vision.
Mnemonic
Cones are for color vision. Rods function best in “roduced” light.
While there are many more rods than cones in the human eye, the central section of
the retina, called the macula, has a high concentration of cones; in fact, its centermost
point, called the fovea, contains only cones. As one moves further away from the fovea, the concentration
of rods increases while the concentration of cones decreases. Therefore, visual acuity
is best at the fovea, and the fovea is most sensitive in normal daylight vision. There
is a blind spot where the optic nerve leaves the eye, as there are no photoreceptors
here, as shown in Figure 2.4.
Figure2.4.Specialized Regions of the Retina
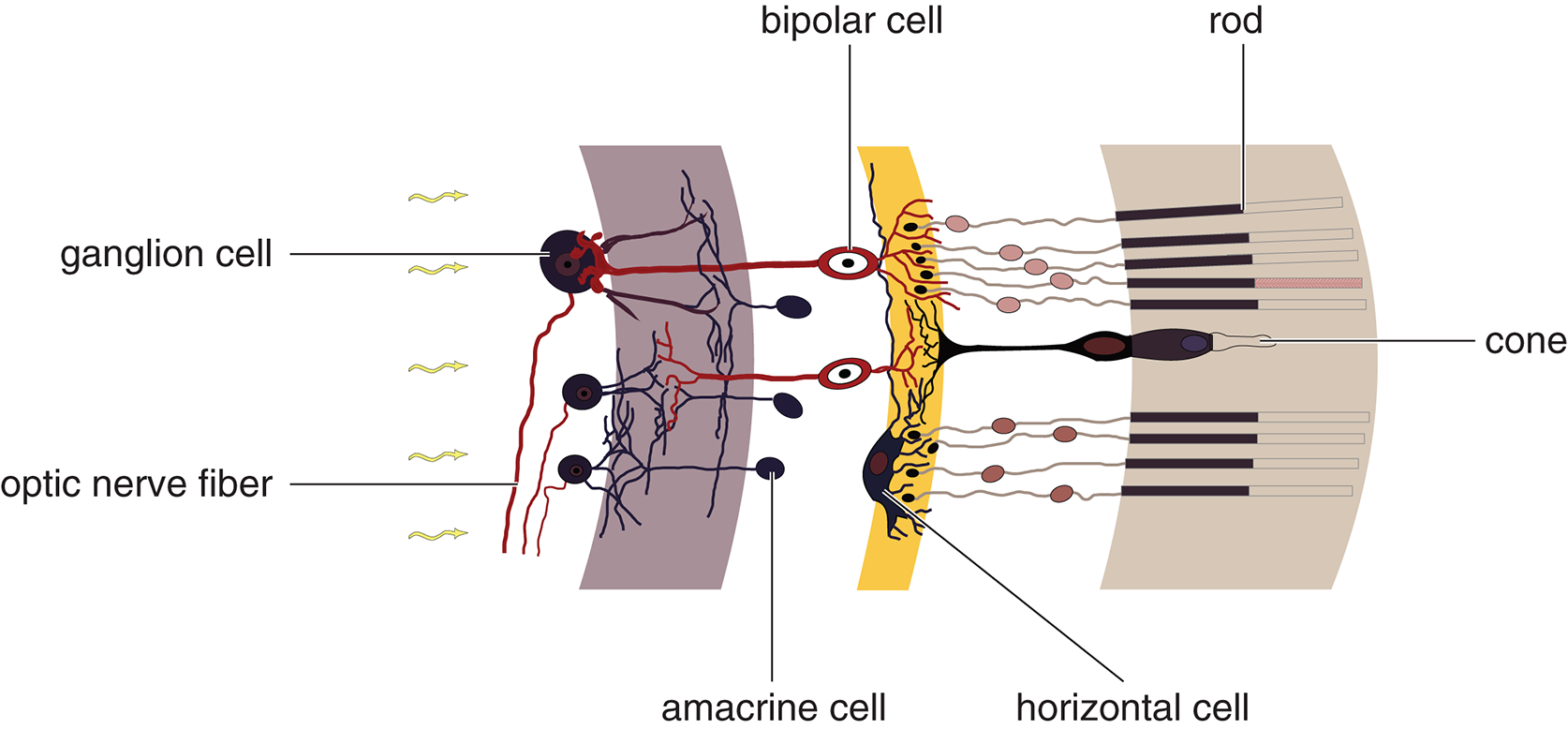
The connection between the rods and cones and the optic nerve is not direct. There
are several layers of neurons in between, as shown in Figure 2.5: bipolar cells, ganglion
cells, horizontal cells, and amacrine cells. Rods and cones connect with bipolar cells, which highlight gradients between adjacent rods or cones. Bipolar cells synapse
with ganglion cells, which group together to form the optic nerve. These bipolar, horizontal, amacrine, and ganglion cells fall “in between” the rods and cones and the optic nerve, and are located in front of the rods and cones closer to the front of the eye. This means that information is being transmitted “forward” towards the lens from the rod and cone cells, through a series of integration steps, until the signal reaches the ganglion cells. Because there are many, many more receptors than ganglion cells, each ganglion cell
has to represent the combined activity of many rods and cones. This results in a loss
of details as information from the photoreceptors is combined. As the number of receptors
that converge through the bipolar neurons onto one ganglion cell increases, the resolution
decreases. On average, the number of cones converging onto an individual ganglion
cell is smaller than for rods. Therefore, color vision has a greater sensitivity to
fine detail than black-and-white vision does.
Amacrine and horizontal cells receive input from multiple retinal cells in the same area before the information
is passed on to ganglion cells. They can thereby accentuate slight differences between
the visual information in each bipolar cell. These cells are important for edge detection,
as they increase our perception of contrasts.
Figure2.5.Cells of the Retina
Visual Pathways
Visual pathways refer to both the physical anatomical connections between the eyes and the brain
and the flow of visual information along these connections. As demonstrated in Figure
2.6, each eye’s right visual field projects onto the left half of each eye’s retina
and each eye’s left visual field projects onto the right half of each eye’s retina.
As the signal travels through the optic nerves toward the brain, the first significant
event occurs at the optic chiasm. Here, the fibers from the nasal half (closer to the nose) of each retina cross paths.
These fibers carry the temporal visual field (further toward the side of the head)
from each eye. Because the temporal fibers (which carry the nasal visual field) do
not cross in the chiasm, this reorganization means that all fibers corresponding to the
left visual field from both eyes project into the right side of the brain, and all
fibers corresponding to the right visual field from both eyes project into the left
side of the brain. These reorganized pathways are called optic tracts once they leave the optic chiasm.
Figure2.6.Visual Pathways
From the optic chiasm, the information goes to several different places in the brain:
the lateral geniculate nucleus (LGN) of the thalamus, through radiations in the temporal and parietal lobes to the visual cortex in the occipital lobe. There are also inputs into the superior colliculus, which controls some responses to visual stimuli and reflexive eye movements.
Real World
When a there is a loud, sudden sound, the superior colliculus aligns the eyes with
the likely stimulus. In other words, it’s the superior colliculus (as well as the
sympathetic nervous system) that gives us the “deer in the headlights” appearance
during the startle response.
Processing
While being able to sense all of the light information around us is useful in its
own right, we must be able to make sense of visual stimuli to be able to interact
with the environment. One process that helps create a cohesive image of the world
is parallel processing. Parallel processing is the ability to simultaneously analyze and combine information regarding color,
shape, and motion. Then, these features can be compared to our memories to determine
what is being viewed. For example, most people can recognize a moving car very easily
from a distance because they are familiar with the usual motions and shapes of cars.
Parallel processing is not only a psychological model, but also has a correlate in
neuroscience called feature detection. Our visual pathways contain cells specialized in detection of color, shape, or motion.
Color
As described above, cones are responsible for color vision.
Shape
Shape refers not only to the three-dimensional geometry of an object, but also our
ability to discriminate an object of interest from the background by detecting its
boundaries. Shape is detected by parvocellular cells, which have very high color spatial resolution; that is, they permit us to see very fine detail when thoroughly examining an object.
However, parvocellular cells can only work with stationary or slow-moving objects
because they have very low temporal resolution.
Motion
Motion is detected by magnocellular cells because they have very high temporal resolution. However, magnocellular cells have
low spatial resolution, so much of the rich detail of an object can no longer be seen
once it is motion. Magnocellular cells therefore provide a blurry but moving image
of an object.
Mnemonic
Magnocellular cells specialize in motion detection.