CHAPTER 9
Conserving California Grasslands into an Uncertain Future

K. Blake Suttle, Erika S. Zavaleta, and Sasha Gennet
Abstract. This chapter focuses on a grassland experiment in Mendocino County, California, to examine the predictability of climate change impacts, and the lessons our work suggests for how we prepare for climate change. Grasslands provide a number of important ecosystem services to the public and are strongly shaped by climate in both composition and function. From experimental data describing multiple forms of ecological response to changing precipitation regimes through time, we show that the complexity of responses increased as the focus of attention narrowed from ecosystem- and community-level metrics down to individual species. Complexity emerged largely from changing interactions among species, which makes aggregate measures like ecosystem function and community diversity more reliable than predictions of responses by individual species. Effects of changing interactions were often lagged relative to the direct effects of physical conditions on species; this means that responses to annual variation in weather may poorly predict responses to longer-term changes in climate. The complexity apparent in species-level responses indicates that predictions for local plant and animal populations under different climate change scenarios could be more difficult than the sophistication of our modeling capabilities would otherwise suggest. In developing on-the-ground adaptation plans, it will be prudent for scientists and managers to allow for uncertainty and expect surprises, particularly where the focus is on individual species rather than broader patterns of diversity or function. Precautionary and flexible management actions should form the starting point in climate change adaptation, with more interventionist measures accompanied by careful monitoring to limit the consequences of inevitable surprises.
Key Points
• Length of the rainy season is a major factor in the production and diversity of California grasslands, much more so than the total amount of precipitation that falls. A key aspect of understanding how systems might respond is identifying what forms of climate information are most relevant, and understanding both the trends and uncertainty in those aspects of change.
• In highly dynamic systems such as arid grasslands, on-the-ground variation in species composition and diversity in response to annual variation in climate may be a poor predictor of longer-term trajectories under decadal and longer climate change. When short-term research and past observations are used to inform management, particularly at the level of individual species, this should be done with caution and an eye toward potential complexities that could derail expectations.
• Uncertainties surrounding how climate will change and how ecosystems will respond complicate efforts to adapt conservation and management practices, but do not preclude effective measures being taken. Approaches like prioritizing conservation and restoration based on abiotic factors and geographic gradients allow actions to take place without reliance on understanding species-specific response.
• Given the complexities that emerge as our geographic focus narrows to the scale of individual sites and our ecological focus narrows to the scale of individual species, the goal of maintaining a given mix of species at a given site may be less practical and less sustainable over the long term than that of maintaining native plant cover and functional and phenological group diversity
• Restoration of specific habitats should be coordinated with and complement landscape-scale conservation actions such as establishment of new protected areas, particularly in light of ongoing grassland habitat loss and conversion in California.
INTRODUCTION
To update management and conservation approaches in an era of climate change, we need to frame our expectations of what the future might look like, and acknowledge which aspects of this future are highly likely and which are most uncertain. Ecological responses to climate change are a challenge to predict, because they may combine many different kinds of direct and indirect effects (Walther 2010, Post 2013). Direct effects include changes in growth, reproduction, and survival rates based on physiological responses to climate itself (i. e., whether a change in physical conditions favors or disfavors an organism, such as by allowing its establishment or preventing its persistence). Indirect effects are those mediated by changes in the biological community around that organism (i. e., how responses to climate change by consumers, competitors, resources, and pathogens further affect its growth, reproduction, or survival). Species do not exist independently of each other, but are enmeshed in intricate webs of interrelations. As a result of the network structure of nature, any one species’ experience of climate change can influence and be influenced by the experience of many other species. We consider here how ecological interactions complicate our understanding of future climate change impacts in grasslands, and how scientists and managers can account for such complexities in developing adaptation plans.
Grasslands are widely distributed across California and host a sizeable fraction of its native biodiversity, including numerous threatened and endangered plants and animals (Stromberg et al. 2007). They are of high economic and cultural importance in the state, particularly with respect to ranching, providing critical habitat and food for wildlife and livestock, and places of recreation for the public. Grasslands vary widely in their specific composition across the state, based on geographic, climatic, and edaphic characteristics, but almost all share two important characteristics. First, grasslands are among the most heavily invaded ecosystems in California, hosting a diverse and abundant flora of European species (Mack 1989, D’Antonio et al. 2007). Annual grasses from the Mediterranean have been particularly successful, dominating millions of hectares throughout the state (Huenneke 1989). Most grasslands in California retain native plants, though these are typically scattered amidst abundant exotic plant cover (Bartolome et al. 1986, Huenneke 1989, Hamilton 1997). Second, grasslands in California are distinct from those in many other regions in that they do not follow clear patterns of succession, with composition and production fluctuating from year to year instead of shifting through a typical set of stages over time. Specifically, standing crop and species composition vary with annual variation in weather, and particularly in the timing and amount of precipitation that falls each year (Murphy 1970, Pitt and Heady 1978, Stromberg and Griffin 1996, Hobbs et al. 2007).
Grasslands, therefore, may be particularly susceptible to changes in rainfall levels that accompany long-term climate change in California. Extended drought in the late 1800s is thought to have contributed to the speed and extent of conversion of California grasslands into exotic-dominated systems (Burcham 1957, Burcham 1961, Major 1988, Corbin and D’Antonio 2003), exacerbating concomitant effects of land-use change, altered fire regimes, and repeated introductions of exotic species. Where abundant native grasses persist in northern coastal regions, their success has been attributed in part to the longer winter rainy seasons and less severe summer droughts that characterize the region (Hektner and Foin 1977, Hayes and Holl 2003). Directional changes in annual precipitation regimes could have important bearing not just for the structure and diversity of these systems but also for how we use them as a result. Conservation of threatened or valuable species, restoration of degraded habitats, and maintenance of the many services provided by California grasslands will all benefit from advance knowledge of what the future holds for these ecosystems.
In this chapter, we consider lessons for grassland management under climate change from a large-scale experiment directly manipulating climate variables over natural grassland plots in northern California. In 2001, Blake Suttle and colleagues at the University of California in Berkeley set up an experiment modifying the intensity and duration of the annual rainy season over replicate plots of grassland in a Mendocino County nature reserve. Leading climate projections at the time (National Assessment Synthesis Team 2000) called for considerable increases in annual precipitation across much of the state, with disagreement as to the specific seasonal timing of those increases. The experiment was originally set up to test how alternative scenarios of rainfall change could affect native–exotic balances among grassland plants in these already heavily invaded systems (Thomsen et al. 2006, Suttle and Thomsen 2008). Knowledge of basic life history differences among the main constituents of California grasslands suggested contrasting expectations for how major groups of native and exotic species should be affected. The most prominent group of invaders, annual species that evolved in the Mediterranean, would be more likely to benefit from additional rainfall during the winter rainy season than from increases that came during the summer drought, as these plants typically complete their life cycle in spring and persist through the summer as seed (Pitt and Heady 1978, Jackson and Roy 1986). Native perennial plants, on the other hand, could benefit from increases in rainfall during the summer drought, as this is a period of low competitive pressure from most exotic species, allowing natives to take advantage of additional precipitation to increase vegetative growth and produce more and larger seeds.
Initial research in the system showed that when plant groups were considered in isolation, these physiology-based mechanisms of response were largely borne out, but that interactions between different plant groups could moderate these responses in important ways (Thomsen et al. 2006, Suttle and Thomsen 2008). The contrasting phenologies of different plant groups in the system, and the differing physiological responses to seasonal rainfall they entail, therefore provide a basis for comparisons of direct and indirect effects in long-term responses; this became the focus of later work in the system (Suttle et al. 2007, Suttle et al., in review), and this work is part of our focus here in considering opportunities for prediction and preemptive management in grasslands under climate change. We compare initial and decadal trajectories in net primary production, biological diversity, and individual species abundances to highlight potential differences in predictability among different levels of ecological response, and to explore the implications of these for grassland management under climate change.
Several features of this experiment make it well suited for addressing questions of predictability. The design emphasized “naturalness”—minimizing infrastructure and observer interference so that grassland species experienced imposed changes in rainfall amid the full complexity of their biological and physical surroundings. Plots consisted of open communities in a protected grassland with no recent history of livestock grazing or anthropogenic disturbance (>75 years). Water was manipulated in a manner that did not change other environmental conditions, such as wind or shade, that would introduce other changes that could potentially confound our ability to understand responses to the treatment itself. Treatments themselves did not represent realistic approximations of climate change, but highly simplified changes in a single climatic variable, setting up straightforward expectations and hypothesis tests. By minimizing complexities introduced from the physical side, the experiment focused on biological complexities such as species interactions, response thresholds, and other nonlinearities. The work also encompassed large enough spatial scales (plots >70 m2) and long enough temporal scales (>10 years) for a rich suite of life history processes to play out in organism responses, but all within a fully controlled, randomized, and replicated framework that allowed formal hypothesis testing.
METHODS
Beginning in January 2001, 36 plots of grassland (∼70 m2 each) at the Angelo Coast Range Reserve in Mendocino County, California (39° 44′ 17.7″ N, 123° 37′ 48.4″ W) have been subjected to one of three annual precipitation treatments: A winter addition of water simulating an intensification of the rainy season (January through March), a spring addition of water simulating an extension of the rainy season (April through June), and an unmanipulated ambient control (Figure 9.1). Treatments simulate predictions for the region from climate models developed at the Hadley Centre for Climate Prediction and Research (HadCM2) and the Canadian Centre for Climate Modeling and Analysis (CCM1) (National Assessment Synthesis Team 2000). Both models projected substantial increases in precipitation throughout northern California over the next century, but the seasonality of these increases differed. The Hadley model called for increases during the current winter rainy season, while the Canadian model predicted increased rainfall extending into the summer, when rain is typically scarce and acts as a key limiting resource.
Each watered plot receives approximately 44 cm of supplementary water over ambient rainfall per year, roughly a 20% increase over mean annual precipitation (216 cm) but within the range of natural variability in both amount and timing at the study site (details in Suttle et al. 2007). Water is collected from a natural spring on a forested mountain southeast of the grassland. The spring drains to the south of the grassland, and a portion of its flow is diverted via pipe to an 1100-L irrigation tank placed on the mountainside approximately 40 vertical meters above the grassland immediately to its east. The tank is continually replenished via gravity feed from the spring, and water has been tested and found to contain nitrogen concentrations within the range present naturally in rainwater at the study site (Suttle et al. 2007).
Water is delivered evenly over the surface of each plot from a RainBird® RainCurtain™ sprinkler specially designed to simulate natural rainfall. Experimental rain additions begin approximately two hours after dawn every third day during the three-month watering period of each addition treatment, and last for one hour. The watering radius is 5 m, and all samples are collected at least 0.5 m inside of the outer edge of the watered area (Figure 9.2).

FIGURE 9.1: The experiment as seen from the air in summer 2001. The spring-addition plots, in which the rainy season is extended into summer, can be recognized from the green coloration imparted by the late flush of nitrogen-fixing forbs in these systems.
The grassland contained a well-mixed assemblage of grasses and forbs of both native and exotic origin on an abandoned terrace of the South Fork Eel River. Approximately 50 plant species are present in the system in a given year, dominated by the exotic annual grasses Bromus hordeaceus, Vulpia myuros, Aira caryophyllea, and Bromus diandrus; the exotic annual forbs Erodium cicutarium, Gallium parisiense, and Hypochaeris glabra; the native annual forbs Madia gracilis, Trichostemma lanceolatum, and Eremocarpus setigerus; and the native perennial forbs Eschscholzia californica and Sanicula bipinnatifida. Relative abundances among these species varied year to year. The three native perennial bunchgrass species Danthonia californica, Elymus glaucus, and Elymus multisetus were also present scattered throughout the system at generally low densities.
In order to understand how adding rainfall to the system would affect three different forms of biological response—productivity, biological diversity, and population abundances—the experiment was designed to meet the complementary goals of obtaining long-term observations of minimally disturbed plots and allowing follow-on experimentation in other, more heavy-use plots. Prior to any work or manipulation in the system, the 36 plots were divided randomly into “Pattern” plots and “Process” plots (18 Pattern plots and 18 Process plots, comprising six replicates each of three rainfall treatments). The Pattern plots served as the template on which the precipitation manipulations could play out over the long term and are the data presented and discussed in this chapter. These plots provide a window into the emergent result of numerous processes playing out in experimental communities in response to the altered precipitation regimes. The Process plots have been used for more intensive and potentially destructive research aimed at understanding causal mechanisms underlying the dynamics observed in the Pattern plots, providing more specific insights into the implications of altered precipitation regimes for restoration of native bunchgrasses (Suttle and Thomsen 2008), biological invasion (Thomsen et al. 2006), carbon and nutrient cycles (Hawkes et al. 2010, Berhe et al. 2012), and dynamics within the soil microbial community (Cruz-Martinez et al. 2009, Hawkes et al. 2010).

FIGURE 9.2: Manipulation and sampling of experimental plots. (a) Water is delivered evenly over the surface of each 5 m radius plot by a sprinkler. (b) Plant production was measured in two predesignated 900 cm2 subplots (small squares) on May 25, July 1, and August 25, targeting peak biomass of different phenological groups. Richness was measured across the growing season by visual inspection in two central 2500 cm2 subplots (bolded large squares). Abundances were measured by point-frame in six permanent 2500 cm2 subplots.
In each replicate Pattern plot (3 treatments × 6 replicates), plant production was measured every year in two predesignated 900 cm2 subplots on May 25, July 1, and August 25, in order to target the peak biomass of different phenological groups of plants present in the study system (Figure 9.2). Each subplot was harvested once and then eliminated from the sampling regime. Subplots were positioned at regular intervals along a single transect laid out in a randomly selected cardinal direction through the center of each plot and then along a second transect running perpendicular to that transect. Biological diversity was measured as the species richness presents across the growing season in two central 2500 cm2 subplots. Population abundances of individual species within experimental communities (and the overall species composition these add up to) were measured by point-frame in six permanent 2500 cm2 subplots. Point-frame measurement involved a series of eight pins dropped vertically at regular intervals into each subplot, with data recorded on the first plant species contacted by each pin as it was lowered into the vegetation, the plant species the pin landed on, and the plant species touching the pin at a marked height of 5 cm above the ground surface (3 hits per pin × 8 pins per subplot × 6 subplots, for 144 hits per plot). Sampling protocols were specifically designed to leave little potential for cross-interference among sampled variables or years (Suttle et al. 2007, Suttle et al., in review). This sampling regime was in place throughout the experiment, and was the only work undertaken in the 18 plots described in this chapter.
Plant production and species richness data were analyzed with repeated-measures ANOVA. Population abundances and species composition data are fundamentally multivariate (abundance of species a in each plot, abundance of species b in each plot, and so on across all species present), and best expressed as patterns of similarity and dissimilarity among plots though multivariate ordination. Ordinations were generated from Nonmetric Multidimensional Scaling (NMS or NMDS); these provide a quick view into compositional differences among communities by rendering variation in abundances of all species onto two axes. The ordinations cannot be read for information about any particular species, but instead show overall levels of similarity and dissimilarity in composition of the plots across all their species. These data were collected for only 15 of the 18 total plots under study, or five replicates per treatment.
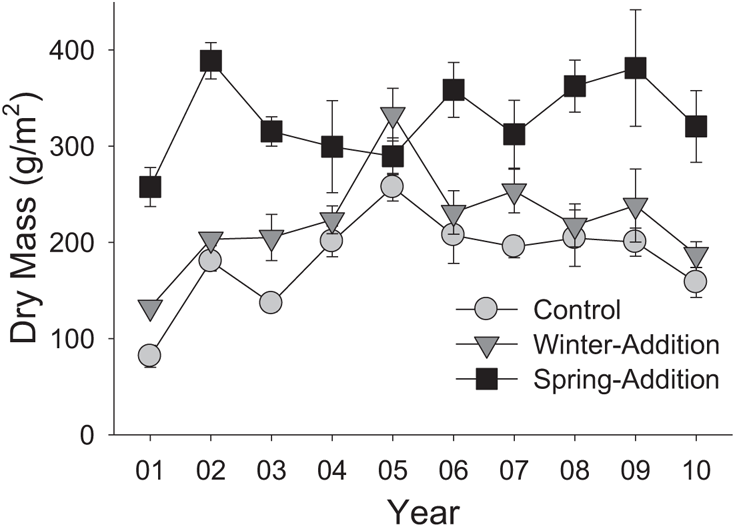
FIGURE 9.3: Effect of watering treatment on primary production. Data represent treatment means plus or minus standard errors for the cumulative biomass accrued across all plant species each year.
RESULTS
The most conspicuous feature of these data, regardless of which type of response is considered, is the difference in effect between the winter-addition treatment (an experimentally intensified rainy season) and the spring-addition treatment (an experimentally extended rainy season) (Suttle et al. 2007, Suttle et al., in review). Adding supplemental rainfall during the winter rainy season did not cause significant deviation from the control in any measured variable or year: Winter-addition plots mirrored control plots in plant production (Figures 9.3 and 9.4), plant diversity (Figure 9.5), plant composition (Figure 9.6), and species abundances (Figure 9.7) through every year of the study. Consequently, the focus of this chapter is on the spring-addition plots, which showed large differences from control plots in every measured variable, although those differences took a different form for each response variable considered.
Spring-addition plots showed large and fairly consistent increases in primary production relative to the other two treatments (Figure 9.3). In the first year of the experiment, this difference was largely due to a single group of plants: Nitrogen-fixing forbs (Figure 9.4a). Other plant species showed roughly similar levels of production across all three treatments in the first year of the experiment. Production of nitrogen-fixing forbs in spring-addition plots remained high relative to the other two treatments over most of the following decade, but was no longer the only biomass difference among treatments after the first year. Annual grass production rose sharply in spring-addition plots relative to the other two treatments in the second and third years of the manipulation, and remained at higher levels in the spring-addition treatment over most of the years that followed (Figure 9.4b). Biomass of other forb species (i. e., those not associated with nitrogen-fixing bacteria) in the spring-addition treatment, on the other hand, fell below levels seen in the other two treatments in the third year of precipitation additions and for several years afterward (Figure 9.4c).
For biological diversity, experimental extension of the rainy season caused significant increases relative to ambient levels in the grassland in the first two years of the manipulation, but this effect disappeared during the third year of rainfall addition, and had reversed by the fourth (Figure 9.5). In 2004, four years into the manipulation, species richness in spring-addition plots was starkly lower than in the other two treatments, and stayed there in 2005. In 2006, six years into the manipulation, one of the wettest months of March on record in California was followed by much higher than normal temperatures over the first few weeks of May; this was accompanied by declines in species richness in control and winter-addition plots near to levels seen in spring-addition plots. Species richness climbed again in control and winter-addition plots following that year, but stayed at significantly lower levels in the spring-addition treatment through the duration of the experiment.

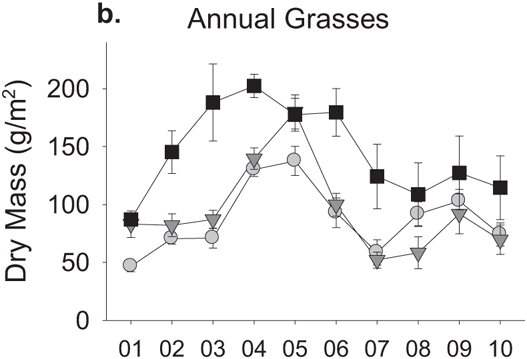
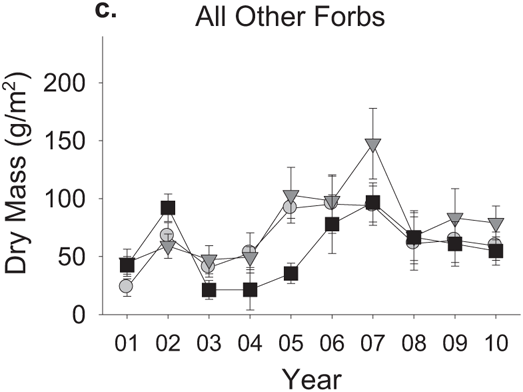
FIGURE 9.4: Effects of watering treatment on primary production by different plant groups. Data represent treatment means plus or minus standard errors for the biomass of vascular plants across each year’s growing season.
Species composition changed first in a consistent direction across spring-addition plots (as nitrogen-fixing forbs became much more abundant relative to other species) and then in different directions in each different spring-addition plot (as a different species rose to dominate in each). So from responses that were initially all in a similar direction relative to the control (Figure 9.6, 2002), spring-addition plots became steadily more dissimilar from each other as the manipulation continued (Figure 9.6, 2010––note that spring-addition plots represent virtually the entire extent of variability measured across the experiment).
Looking at changes in the abundances of individual species within these communities, we can understand how spring-addition plots have grown so dissimilar from each other over time. Certain individual species have shown wildly different responses to experimental extension of the rainy season from one plot to the next (Figure 9.7). As a result, each replicate plot within the spring-addition treatment is dominated by a single species, but it is a different species in every one. The native perennial bunchgrass D. californica exploded in one community subjected to extended rainfall each year but remained at ambient levels in each of the other four (Figure 9.7a). Likewise, the exotic annual grass B. diandrus, the annual forb Geranium dissectum, and the moss species Homalothecium pinnatifidum each rose to very high levels of abundance in a single replicate only. The pattern is less distinct for the nitrogen-fixing legume species Lotus micranthus, which has shown generally positive responses to experimentally extended rainfall across all replicate plots (Figures 9.7b–9.7e).
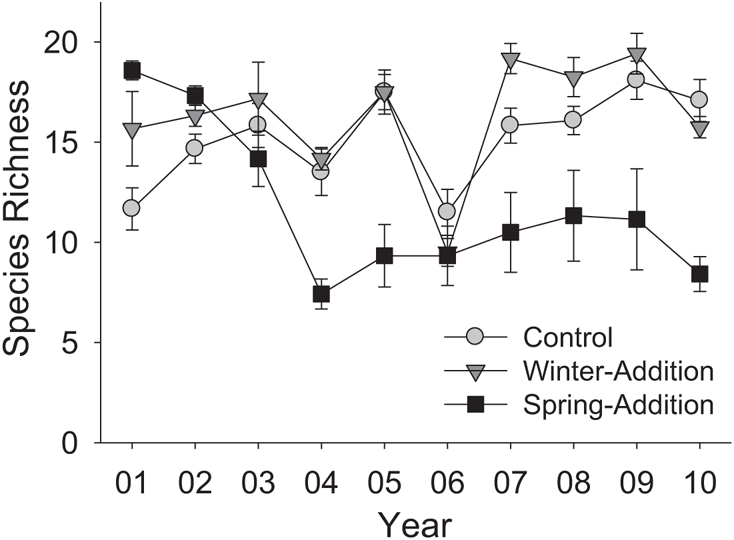
FIGURE 9.5: Effects of watering treatment on biological diversity. Data represent treatment means plus or minus standard errors for the species richness of vascular plants across each year’s growing season.
DISCUSSION
The simplicity of the rainfall additions, together with the detailed understanding of California grassland systems that has emerged out of the long history of ecological research in these systems, helped us develop a priori expectations of what simple physiology-driven responses would look like in the different response variables (e. g., Pitt and Heady 1978, Jackson and Roy 1986, Hobbs and Mooney 1995). Allowing actual responses to play out in the full complexity of a natural setting over a decade, we can identify where deviations from physiological expectations arise, and potentially identify what factors emerge to drive these deviations.
The fact that complex dynamics emerged from only one of the two watering treatments, with the other causing no meaningful changes at all, emphasizes that these ecological complexities are of secondary concern to the overlying details of climate change itself. A key take-home point here is that timing matters. Much of our dialogue on climate change focuses on changes in average annual values and extreme events, but here we see how the same annual change can have very different biological effects depending on the specific seasonal timing over which it unfolds. This illuminates a critical challenge in climate change science and adaptation—uncertainty over regional precipitation changes generally and in the annual timing of those changes specifically. Case in point, in the time since the rainfall experiment described here was established, predictions for precipitation change in California have changed dramatically, with many leading models, including more recent projections from the Hadley Centre, now forecasting decreasing annual rainfall over much of California (e. g., Wilkinson et al. 2002, Hayhoe et al. 2004, Cayan et al. 2006). Unless and until climate models achieve considerable further reductions in uncertainties over precipitation and timing, managers should consider a range of best- and worst-case scenarios and solutions with respect to how these variables will change in their systems.
In terms of understanding the processes underlying responses observed in this study system, the lack of any effect of additional winter rainfall can be understood on strictly physiological terms for all three sets of response variables: Primary production, biological diversity, and population abundances. There is relatively little overlap between periods of warm temperatures conducive to plant growth and periods of high moisture availability in northern California. Roughly 95% of annual precipitation typically falls between November and April (Major 1988), when temperatures and light levels are low and most plants are able only to use small amounts of water (Evans and Young 1989). Increases in water during this time, therefore, proved to be largely superfluous. Extending the rainy season via water addition through the spring and into summer produced many more dramatic changes in the grassland.
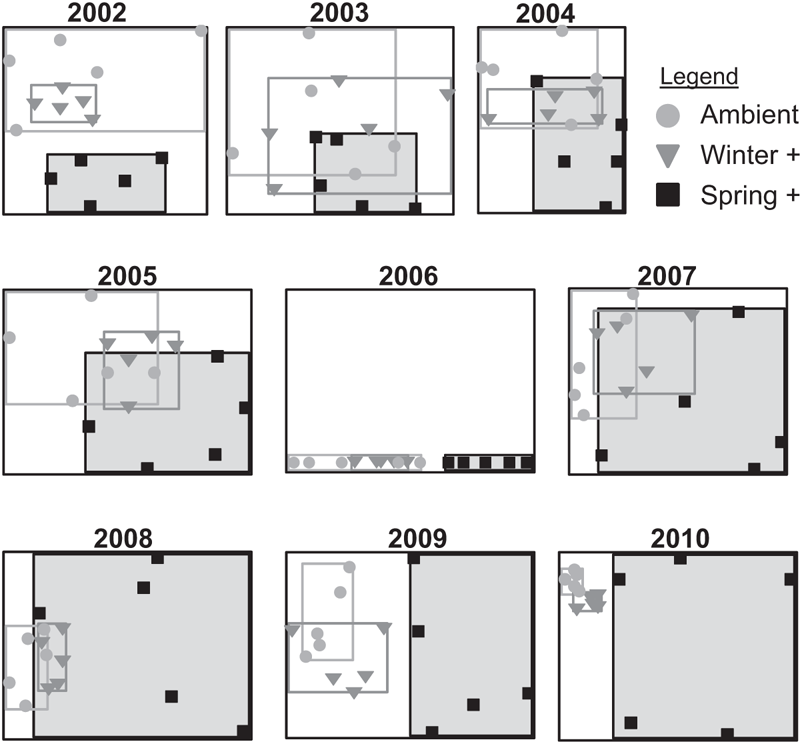
FIGURE 9.6: Treatment-level variability in the species composition of experimental plots. Data reflect patterns of similarity and dissimilarity in relative abundances across all species, as expressed by Nonmetric Multidimensional Scaling (NMS). The compositional similarity and dissimilarity are expressed as distances along two axes, so that the closer two points on a given figure, the more similar those communities. Rectangles are drawn as a visual guide to the total amount of variation along each axis accounted for by the plots of a single treatment.
The fact that each response variable behaved differently under experimentally extended spring rainfall points to a different set of processes driving changes at each ecological level considered. Ecosystem functions typically involve a large number of species, making them relatively robust to changes in the number and identity of particular species. Thus primary production—the function studied most closely in experimental plots—changed according to processes we understand well (i. e., physiological tolerances and preferences, which can be tested directly or gleaned from time series data or distribution records). Plant production more than tripled in the first year and doubled in the second year compared to the ambient levels in the grassland (Figure 9.2). Summer is typically a period of high water stress in northern California grasslands; plants responded to alleviation of this stress with increased growth and, in certain species, extended longevity (Suttle et al. 2007). These effects were apparent immediately and remained relatively consistent throughout the duration of the experiment.

FIGURE 9.7: Effects of watering treatment on individual species abundances. Data represent plot-level population abundances for each of five individual species in five replicate plots of each treatment. Spring-addition plots are represented by black lines; for visual simplicity and to highlight the divergent trajectories evident in spring-addition plots, winter-addition and control plots are represented by lines in the same shade of gray.
Diversity measures are a little more narrowly focused than overall production, in that the number of species becomes important, if not their identity. Accordingly, diversity changes in experimental plots have been more complex to model, requiring that we account for a set of species interactions in addition to basic physiological responses to climate to understand the reversal in the effect of extended rainfall through time (Suttle et al. 2007). Dividing overall plant production into three broadly defined functional groups (i. e., nitrogen-fixing forbs, annual grasses, and non-fixing forbs) sheds light on the nature of these interactions. The strongest initial response to the extended rainfall season came from nitrogen-fixing forbs (Figure 9.4a). Exotic annual grasses showed no significant response to the first year of the spring-addition treatment, but after the proliferation of nitrogen-fixing forbs that year, annual grass production rose dramatically (Figure 9.4b). Exotic grasses are generally the first plants to germinate each year and among the earliest to complete their life cycle and senesce. This early phenology limits direct responses to extended spring rainfall (Pitt and Heady 1978, Jackson and Roy 1986), but allows these plants to benefit in the subsequent growing season as abundant N-fixer litter decomposes and provides increased nitrogen (Bentley and Green 1954, Bentley et al. 1958, Jones et al. 1990). As this process was repeated over several years, the accumulated annual grass litter apparently suppressed germination and regrowth of leafy forbs (Figure 9.4c), driving sharp declines in plant species richness (Figure 9.5).
Forb species after forb species disappeared from spring-addition plots until these plots were made up of only a few species of annual grasses and the nitrogen-fixing forb L. micranthus (which, with its spindly growth form and narrow leaves, was able to penetrate the grass litter and remain a productive constituent of spring-addition plots). These dynamics, too, however, turned out to be short-lived, for within all but one of these annual grass-dominated communities, another species took hold and ultimately rose to dominance. The most surprising finding from this research, however, is that the “winner” was a different species in every plot. Ten years into the altered precipitation regimes, a different species now dominates each replicate spring-addition plot (Figure 9.7) despite the fact that each of these species is present in every replicate community (Suttle et al., in review).
From initially well-mixed communities of roughly similar composition, spring-addition plots have grown increasingly dissimilar from each other over a decade of extended seasonal rainfall (Figure 9.6). Thus a simple manipulation of one environmental variable, imposed identically over discrete plots of well-mixed grassland in a single meadow, has given rise to communities that are conspicuously different in their makeup from replicate to replicate. Population trajectories for individual species, very much at the center of aspirations in predictive ecology, here show a degree of context dependency that prevents our being able to explain or model them based on experimental data. Many species in the experiment have failed to show any kind of trajectory at all, responding in different directions from replicate to replicate under a single experimental treatment. The extent of idiosyncrasy seen in how populations of individual species changed in response to rainfall addition suggests a high degree of uncertainty around predictions of local population changes, as these appear to be governed by factors that vary over fine spatial scales.
The scale of study plots (70 m2) and therefore of the context dependency seen in population trajectories make it important to consider what changes might look like if rainfall were added at the scale of whole grasslands. The single-species dominance we see at the scale of experimental plots would likely manifest as increased spatial heterogeneity in vegetation composition at the scale of the meadow as a whole over the decadal scale considered here, but may well lead to dominance by some smaller subset of these species as the new climate regime took hold over longer time periods. Extrapolating from plot-scale to whole-treatment differences, species richness remains lower under the extended-rainfall treatment; even with a different species dominating each replicate plot, there are so few other species able to persist amid those levels of dominance that collective richness across plots is still lower than that measured in winter-addition and control plots. And richness differences notwithstanding, whether we view results at the scale of experimental plots or at the scale of the whole experiment, complexity increases as we narrow from our focus down toward individual species.
SOLUTIONS, ADAPTATION, AND LESSONS
Uncertainty and scale deserve serious consideration in plans for preemptive management around climate change (Wiens and Bachelet 2009). We cannot simply delay action while we wait for more precise forecasts, because in some cases the problems will grow worse and solutions more difficult and in some cases that hoped for precision will remain elusive. As climate scientist Stephen Schneider often said, “This is not a world where you can let the perfect crowd out the good.” Where uncertainty in how some species or ecosystem will respond to future climate change arises not from a lack of appropriate empirical data, but from the fundamental nature of the underlying processes, we can act now, even if it means reframing certain priorities. There are many courses of remedial action with effectiveness not bound to any specific vision of the future (e. g., reduce emissions, restore degraded landscapes, expand protected areas, and improve connectedness among them) (Heller and Zavaleta 2008). These are likely to be most effective if undertaken in complement with each other. The step-wise changes in response complexity from ecosystem to community to population levels in the experiment suggest that, at local scales, changes in individual species will show greater variation and uncertainty than changes in overall diversity or ecosystem function. When management targets are more narrowly ecologically focused, the most robust management actions will be broader and more precautionary, allowing for greater uncertainty in target responses.
Many studies and management efforts across grassland types in California have demonstrated that reversing biological invasions once established and restoring native species once lost is difficult, whether in mesic coastal prairie or arid interior grassland (Corbin et al. 2004). Although the restoration component of resource management and conservation is critical, site-scale efforts (e. g., 1–1000 ha) will be insufficient to achieve protection of the full array of grassland types and constituent communities. Restoration must be coordinated with and complement landscape-scale conservation actions such as establishment of new protected areas, particularly in light of ongoing grassland habitat loss and conversion in the state. Put another way, protecting large areas of grasslands that can be flexibly managed for conservation and compatible uses is a prerequisite for adaptive management or restoration in an uncertain climate future. Protected areas must be large enough and adequately linked so that species with a variety of life histories, from wide-ranging mammals and breeding birds to host-specific pollinating insects and edaphic-specialist plants, can maintain viable populations and potentially migrate to new areas when portions of current ranges become unsuitable.
Because grasslands and savanna cover millions of hectares in California, a complete inventory and biological assessment of sites is impractical. However, landscape-scale ecological research can help to identify and prioritize areas for protection where diverse communities are more likely to persist into the future (Klausmeyer and Shaw 2009, Klausmeyer et al. 2011). For example, recent research along the Central Coast of California suggests that grassland plant species richness in that ecoregion is correlated with elevation, steep north-facing slopes, and coarse soils, with higher cover of native species on less fertile soils (high C:N ratio, low phosphorous) (Gea-Izquierdo et al. 2007, Gennet, unpublished data). Grassland areas that are current and, to the best of our knowledge, likely future hotspots of species diversity can be identified and prioritized for protection using readily available digital GIS datasets (in this case, digital elevation models, geology, and soil layers).
Surprises are in store, however, so another practical step may be to broaden acceptable ecological outcomes and the scales by which they are defined. Scaling outward from a species-level focus to functional groups, phenological groups, or ecosystem processes, we can expect less idiosyncratic responses to climatic change (Zavaleta et al. 2003, Bai et al. 2004, Hobbs et al. 2007). Scaling outward geographically from individual sites to regional and broader scales, we can expect less idiosyncrasy in responses by individual species (Wiens 1989, Pearson et al. 2004). The goal of maintaining a given mix of species at a given site may be less practical and less sustainable over the long term than that of maintaining native plant cover and functional and phenological group diversity [e. g., managing to retain deeply rooted perennial plants to help control star thistle (Centaurea solstitialis)]. The goal of protecting a given species or mix of species, on the other hand, will be more practically accomplished through a regional perspective that views individual sites as part of a broader network. For any particular species or site, however, an expectation of surprises, a monitoring effort sufficient for early detection of the same, and a responsiveness and adaptability built into conservation and management plans are all commensurate with the challenge we face.
Changes in California’s climate could have profound consequences for grassland structure and species composition, and some of these will unfold in highly complex ways. With impacts varying by species and life stage, tied strongly to seasonal timing, and propagating widely along interaction networks, species- and site-level predictions of ecological response may carry high levels of uncertainty, and management strategies built upon them should allow for this. The most practical path forward requires acknowledging uncertainties and asking “what then?” These are discussions in which all stakeholders should be engaging. We will not be able to preempt every harmful impact of climate change, but acting along these lines, to borrow a phrase from USGS scientist Nate Stephenson, we should be able to usher in some intended consequences among all the unintended ones.
Manager Comments
K. Blake Suttle
in conversation with
Andrea Craig
Suttle: Have you observed changes in the systems that you manage that seem linked to changes in climate? As climate continues to change, what ecological changes do you expect to see?
Craig: Ranchers and other land managers in our area are observing a long string of abnormal weather years, with the timing of precipitation being particularly unusual and unpredictable. Cattle-production operations and wildlife are forced to follow the timing of precipitation. As climate continues to change, I would expect to see a shift in forage and drinking water availability, which would shift the rate and timing of stocking, and migratory patterns and life cycles of wildlife. I would expect to see change in vegetation composition and availability (timing and abundance), leading to a cascade of changes related to soil properties and the organisms most suited to persist and evolve in this landscape.
Suttle: How do you see this information fitting in with the work that you do to conserve grassland systems in California? What kinds of decisions could this type of research be used to inform?
Craig: This information fits in with my rangeland work by informing the decisions involved in determining lease terms, and by informing strategic planning decisions for conservation of the targets that brought the Nature Conservancy to this region. Abnormal weather patterns force us to adapt, and lease and easement terms need to be written to accommodate uncertainty. Climate change research informs my preparation of functional agreements with ranchers and other land managers.
Suttle: Is understanding climate change responses an area where stronger or continued partnership with academic researchers is highly valued? Can you give an example of an ecological study that would be of particular interest to you, and that would have the potential to influence management actions?
Craig: Continued partnership with academic researchers is valued, especially when studies support anecdotal evidence seen on the ground. Examples include: (1) Are precipitation events occurring later in the growing season? (2) Will weather patterns support some summer grazing, or should we expect deer to stay longer at high-elevation habitats? (3) Do we have more grass and less forb forage? (4) Have black bears stopped hibernating, thus pressuring their food supply and other big game? Land managers can certainly make more informed decisions when informed by scientific research. I feel this especially holds true for those managing expansive and complex landscapes that may cover multiple watersheds and have a variation of overlapping ecological systems. The complexity and interconnectivity of these systems is too diverse to manage without a strong research partnership.
Suttle: When you think about your ability to change management approaches or shift conservation priorities in this system, what do you see as the main constraints? Are there particular areas of study (ecological science, climate science, policy, data management, etc.) that could help overcome this constraint?
Craig: The main constraints for me to change a management approach are the leases and conservation easements that are not currently accommodating extreme intra-annual variations seen on the ground. Policies within public agencies, land trusts, and other leasing entities should be modernized to ensure a certain level of flexibility in the terms applied to all new leases and conservation easements, and how these fit into a greater management plan. The degree to which lease and easement terms could accommodate unpredictable change can only be determined with the support of ecological and climate science studies examining weather trends, forage productivity, and changes in biodiversity across the landscape.
Suttle: What do you see as the main challenge to linking information on potential future climatic conditions to your work protecting grassland systems into the future?
Craig: The main challenge in linking potential future climate change information to my conservation work is regular and open communication among involved parties: Landowners, ranchers, lessees, field-based staff, senior managers, and others. Monitoring and decision-making frameworks will need to be defined. By sharing information effectively, and being in tune with what is happening on the land, greater flexibility can be built into the whole system.
Suttle: To what extent is monitoring an important management objective at the places where you work? Are monitoring plans set up in a way that captures responses to variation in the seasonality of rainfall? Has this chapter led you to consider any changes in objectives or methods for monitoring (e. g., less species-specific)?
Craig: Monitoring is very important and structured, performed on an annual basis on all properties managed with a conservation easement in this project area. Monitoring plans are not currently set up in a way to capture variation in rainfall seasonality. This study has led me to consider adding to existing monitoring objectives, such as measuring vegetation change outside of the effect of grazing (current quantitative comparisons are for residual dry matter in grazed areas only). This however raises a concern for added staff time and data management responsibilities.
Suttle: Have changes in climate led to changes in your approach to management? Have your tools for addressing the challenges of exotic plants changed or become easier or harder to use (i. e., prescribed fire, use of grazing or herbicides)? Have costs of management changed?
Craig: Weighing in climate change certainly has changed my approach to management and responding to the non-static nature of our environment. During the relatively short time frame in which I’ve been managing invasive species it has become more challenging to prioritize goals while using limited resources. Prescribed fire is increasingly restricted, and herbicide use, though seemingly cost-effective, also has its limitations. The need for additional monitoring and research has increased, and with an eye on the importance of adequate stewardship, management costs have without a doubt risen.
The Angelo Reserve study results confirm that the biggest rangeland-management challenge the California landowners and cattle operators currently face—uncertainty—is already being exacerbated by climate change. Many ranchers and land managers are already looking for solutions. One landowner grazing under easement in the Lassen Foothills of northeastern California recently asked me, “When is everyone going to stop talking about how weird the weather is and just start planning for it?” Managers at all levels know the question is not whether the future will be increasingly unpredictable; it’s what they and policy makers can and should do about it.
Freshwater springs, vernal pools, and swales dot the Lassen Foothills, enabling wildlife and cattle to find water throughout much of the year. However, as in much of California, thin soils limit plant cover to annual grasses and forbs with abbreviated and highly opportunistic life cycles. This vegetation may be adaptable to climate change, and is in turn a highly limiting factor for ranching operations. The most successful managers are those that can be as adaptable and responsive to conditions within a given year as is the vegetation on which the livestock rely.
Here is one solution that would improve intra-annual flexibility for managers at the individual property or landscape scale: Legally binding land ownership and management agreements such as leases and conservation easements could be written to better accommodate the increasingly unpredictable nature of California’s grazing season. Specifically, season dates defined in leases for turning out and removing cattle should be based on field observations of resource conditions in the field, including weather patterns, water availability, and productivity, rather than predetermined dates. This would enable better utilization of forage, supporting the cattle industry, and would not unduly impact grasslands. Policies within public agencies, land trusts, and other leasing entities should be modernized to ensure these types of flexible terms are applied for all new leases and conservation easements.
Making these decisions in real time will require ranchers and field-based staff to communicate regularly and openly with lessees and agency staff, and for those managers to trust the knowledge and good intentions of those with their eyes on the ground. In some cases this will be a shift. Monitoring and decision-making frameworks for these quick-turnaround decisions will need to be defined. By sharing information and being in tune with what is happening on the land, greater flexibility can be built into the whole system.
Some cattle operators are currently working enough land to juggle abnormal weather patterns, without undue hardship. Operators with less acreage do not have as much flexibility. On a recent mid-April day, at least one local rancher was branding in a steady spring rain, working with a team of cowboys to keep irons hot despite mud- and moisture-soaked coats. We have been catching up with an average year’s precipitation the last few weeks which has resulted in, for the second year in a row, ranchers needing to gather cattle out of the valley while conditions are finally favorable and the high country is blanketed in the season’s deepest snowpack yet. Consistently abnormal weather patterns associated with climate change may put smaller operators out of business sooner than later. Conservation of this landscape is dependent on keeping the patchwork of ranchers ranching, large scale and small scale alike. Scientific study of climate change will continue to provide valuable information that land managers can integrate into decision-making frameworks. But solutions must also be welcome from those who know each patch of ground best, and must be found quickly. The success of managing for uncertainty will hinge upon the close communication we are able to maintain with our neighbors and other stakeholders in this rural community.
LITERATURE CITED
Bai, Y., X. Han, J. Wu, Z. Chen, and L. Li. 2004. Ecosystem stability and compensatory effects in the Inner Mongolia grassland. Nature 431:181–183.
Bartolome, J. W., S. E. Klukkert, and W. J. Barry. 1986. Opal phytolyths as evidence for displacement of native California grassland. Madroño 33:217–222.
Bentley, J. R. and L. R. Green. 1954. Stimulation of native clovers through application of sulfur on California foothill range. Journal of Range Management 7:25–30.
Bentley, J. R., L. R. Green, and K. A. Wagnon. 1958. Herbage production and grazing capacity on annual-plant range pastures fertilized with sulfur. Journal of Range Management 11:133–140.
Berhe, A. A., K. B. Suttle, S. D. Burton, and J. F. Banfield. 2012. Contingency in the direction and mechanics of soil organic matter responses to increased rainfall. Plant and Soil 358:371–383. doi: 10.1007 / s11104-012-1156-0.
Burcham, L. T. 1957. California Range Land: An Historico-Ecological Study of the Range Resource of California. Division of Forestry, Department of Natural Resources, State of California, Sacramento, CA.
Burcham, L. T. 1961. Cattle and range forage in California 1770–1880. Agricultural History 35:140–149.
Cayan, D., A. L. Luers, M. Hanemann, G. Franco, and B. Croes. 2006. Scenarios of Climate Change in California: An Overview. California Climate Change Center, Sacramento, CA.
Corbin, J. D. and C. M. D’Antonio. 2003. Competition between native perennial and exotic annual grasses: Implications for a historical species invasion. Ecology 85:1273–1283.
Corbin, J. D., C. M. D’Antonio, and S. J. Bainbridge. 2004. Tipping the balance in the restoration of native plants: Experimental approaches to changing the exotic: Native ratio in California grassland. In M. Gordon and L. Bartol (eds), Experimental Approaches to Conservation Biology. University of California Press-Cooperative Extension, Berkeley, CA. 154–179.
Cruz-Martinez, K., K. B. Suttle, E. L. Brodie, M. E. Power, G. L. Andersen, and J. F. Banfield. 2009. Despite strong seasonal responses, soil microbial consortia are more resilient to long-term changes in rainfall than overlying grassland. The ISME Journal 1:1–9.
D’Antonio, C. M., C. Malmstrom, S. A. Reynolds, and J. Gerlach. 2007. Ecology of invasive non-native species in California grassland. In M. R. Stromberg, J. D. Corbin, and C. M. D’Antonio (eds), California Grasslands: Ecology and Management. University of California Press, Berkeley, CA. 67–85.
Evans, R.A. and J.A. Young. 1989. Characterization and analysis of abiotic factors and their influences on vegetation. In L.F. Huennecke and H. Mooney (eds.), Grassland Structure and Function: California Annual Grassland. Kluwer Academic Publishers, Dordrecht. 13–28.
Gea-Izquierdo, G., S. Gennet, and J. W. Bartolome. 2007. Assessing plant-nutrient relationships in highly invaded Californian grasslands using non-normal probability distributions. Applied Vegetation Science 10:343–350.
Grebmeier, J. M., J. E. Overland, S. E. Moore, E. V. Farley, E. C. Carmak, L. W. Cooper, K. E. Frey, J. H. Helle, F. A. McLaughlin, and S. L. McNutt. 2006. A major ecosystem shift in the northern Bering Sea. Science 311: 1461–1464.
Hamilton, J. G. 1997. Changing perceptions of pre-European grasslands in California. Madroño 44:311–333.
Hawkes, C. V., S. K. Kivlin, J. Rocca, V. Huguet, M. Thomsen, and K. B. Suttle. 2010. Fungal community responses to precipitation. Global Change Biology 17:1637–1645.
Hayes, G. and K. D. Holl. 2003. Cattle grazing impacts on annual forbs and vegetation composition of mesic grasslands in California. Conservation Biology 17:1694–1702.
Hayhoe, K., D. Cayan, C. B. Field, P. C. Frumhoff, E. P. Maurer, N. L. Millers, S. C. Moser, S. H. Schneider, K. C. Cahill, E. E. Cleland et al. 2004. Emissions pathways, climate change, and impacts on California. Proceedings of the National Academy of Science of the United States of America 101:12422–12427.
Hektner, M. M. and T. C. Foin. 1977. Vegetation analysis of a northern California Coastal prairie: Sea Ranch, Sonoma County, California. Madroño 21:83–103.
Heller, N. E. and E. S. Zavaleta. 2008. Biodiversity management in the face of climate change: A review of 22 years of recommendations. Biological Conservation 142:14–32.
Hobbs, R. J. and H. A. Mooney. 1995. Spatial and temporal variability in California annual grassland – results from a long-term study. Journal of Vegetation Science 6:43–56.
Hobbs, R. J., S. Yates, and H. A. Mooney. 2007. Long-term data reveal complex dynamics in grassland in relation to climate and disturbance. Ecological Monographs 77:545–568.
Huenneke, L. F. 1989. Distribution and regional patterns of California grasslands. In Huenneke, L. F. and Mooney, H. A. (eds), Grassland Structure and Function: California Annual Grassland. Kluwer Academic Publishers, Dordrecht. 1–12.
Jackson, L. E. and J. Roy. 1986. Growth patterns of mediterranean annual and perennial grasses under simulated rainfall regimes of southern France and California. Acta Ecologica / Ecologica Plantarum 7:191–212.
Jones, M. B., M. W. Demment, C. E. Vaughn, G. P. Deo, M. R. Dally, and D. M. Center. 1990. Effects of phosphorus and sulfur fertilization on subclover-grass pasture production as measured by lamb grain. Journal of Production Agriculture 3:534–539.
Klausmeyer, K. R. and M. R. Shaw. 2009. Climate change, habitat loss, protected areas and the climate adaptation potential of species in Mediterranean ecosystems worldwide. PLOS ONE 4:e6392.
Klausmeyer, K. R., M. R. Shaw, J. B. MacKenzie, and D. R. Cameron. 2011. Landscape-scale indicators of biodiversity’s vulnerability to climate change. Ecosphere 2:art88.
Mack, R. N. 1989. Temperate grasslands vulnerable to plant invasion: Characteristics and consequences. In J. A. Drake, H. A. Mooney, F. di Castri, R. H. Groves, F. J. Kruger, M. Rejmanek, and M. Williamson (eds), Biological Invasions: A Global Perspective. John Wiley and Sons, New York, NY. 155–179.
Major, J. 1988. California climate in relation to vegetation. In M. Barbour and J. Major (eds), Terrestrial Vegetation of California. California Native Plant Society, Sacramento, CA. 11–74.
Murphy, A. H. 1970. Predicted forage yield based on fall precipitation in California annual grasslands. Journal of Range Management 23:363–365.
National Assessment Synthesis Team. 2000. Climate Change Impacts on the United States: The Potential Consequences of Climate Variability and Change. US Global Change Research Program, Washington, DC.
Pearson, R. G., T. P. Dawson, and C. Liu. 2004. Modelling species distributions in Britain: A hierarchical integration of climate and land-cover data. Ecography 27:285–298.
Pitt, M. D. and H. F. Heady. 1978. Responses of annual vegetation to temperature and rainfall patterns in northern California. Ecology 59:336–350.
Post, E. 2013. Ecology of Climate Change: The Importance of Biotic Interactions. Princeton University Press, Princeton, NJ, USA.
Stromberg, M. R. and J. R. Griffin. 1996. Long-term patterns in coastal California grasslands in relation to cultivation, gophers, and grazing. Ecological Applications 6:1189–1211.
Stromberg, M. R., J. D. Corbin, and C. M. D’Antonio. 2007. California Grasslands: Ecology and Management. University of California Press, Berkeley, CA.
Suttle, K. B. and M. A. Thomsen. 2008. Climate change and grassland restoration: Lessons from a rainfall manipulation in California. Madroño 54:225–233.
Suttle, K. B., M. A. Thomsen, and M. E. Power. 2007. Species interactions reverse grassland responses to changing climate. Science 315:640–642.
Suttle, K. B., M. A. Thomsen, M. Sullivan, D. T. Gerber, and M. E. Power. In review. Limits on predictability emerge in grassland responses to decade-scale climate forcing.
Thomsen, M., C. D’Antonio, K. B. Suttle, and W. P. Sousa. 2006. Ecological resistance seed density, and their interactions determine patterns of invasion in a California coastal grassland. Ecology Letters 9:160–170.
Walther, G. R. 2010. Community and ecosystem responses to recent climate change. Philosophical Transactions of the Royal Society, B 365:2019–2024.
Wiens, J. A. 1989. Spatial scaling in ecology. Functional Ecology 3:385–397.
Wiens, J. A. and D. Bachelet. 2009. Matching the multiple scales of conservation with the multiple scales of climate change. Conservation Biology 24:51–62.
Wilkinson, R., K. Clarke, M. Goodchild, J. Reichman, and J. Dozier. 2002. The Potential Consequences of Climate Variability and Change for California: The California Regional Assessment. U.S. Global Change Research Program, Washington, DC.
Zavaleta, E. S., M. R. Shaw, N. R. Chiariello, B. D. Thomas, E. E. Cleland, C. B. Field, and H. A. Mooney. 2003. Grassland responses to three years of elevated temperature, CO2, precipitation, and N deposition. Ecological Monographs 73:585–604.