The Stargazer lily (Lilium sp. hybrid)
A flower’s fragrance declares to all the world that it is fertile, available, and desirable, its sex organs oozing with nectar. Its smell reminds us in vestigial ways of fertility, vigor, life-force, all the optimism, expectancy, and passionate bloom of youth. We inhale its ardent aroma and, no matter what our ages, we feel young and nubile in a world aflame with desire.
—Diane Ackerman, A Natural History of the Senses, 1990
The Stargazer lily (Lilium sp. hybrid)
Here for you to identify is a living organism, much loved and admired. Having no say in the matter, millions are bought and sold, removed from their natural habitat for the pleasure of the buyers, living fast, dying young, without offspring—then discarded without a second thought. Sometimes they enjoy a better fate, free to live outdoors, reproducing prolifically, enjoy full life spans, their beauty on display for all to see.
The smell of this organism is a hauntingly sweet fragrance, once inhaled, never forgotten. The appearance is dramatic. A long, tumescent rod, topped by a broad, gray-purple tip oozing a clear, sticky liquid, juts suggestively from the center of a yellow, starlike throat. Surprisingly, this phalliclike structure is a female, not a male, organ. The broad, viscid tip evolved to catch and hold pollen grains—dustlike particles that are the male essence. Surrounding the long rod, six yellow-green arms project from the starlike mouth, the bulging, orange packages at their tips ready to split wide-open to release sperm at just the right time.
A hermaphrodite, with male and female parts united in one body, it is capable of having sex there within, though preferring to mate with others, especially nonrelatives. The largest and most spectacular part of this body consists of the two pinwheel-like whorls surrounding the sex organs, one on top of the other, outlined in white and shaded from white to pale pink to a deep almost-purple-pink hue, their surfaces spotted with raised, red dots. Below, where the whorls join the green stalk, is an ovary, containing eggs waiting to be fertilized. Lacking legs or wings, this regal-looking organism is grounded—unable to move during its entire lifetime—and thus dependent upon animal intermediates. If its concentrated appeal is able to reach out, it is the bees or perhaps other animals that transport its sperm helping it to reproduce. Earthbound and immobile, it casts a gaze upward, to the stars, as it whispers a name sending a message of fanciful dreams, optimism, and unlimited possibilities. What am I?
A flower of course, but more specifically a Stargazer lily. The Stargazer, which came to market in 1978, is the result of many years of crossbreeding efforts by the eccentric genius lily-breeder Leslie Woodriff, who wanted a beautiful, fragrant, spotted lily that would hold its blooming head proudly high, unlike any other horticultural lily of the time.
Florist-shop product it may be, but the Stargazer is nonetheless typical of all flowers—which is to say that even though a breeder tinkered with its genealogical stock to make it a crowd-pleaser in the human marketplace, its structure, appearance, and smell are ultimately all about attracting the pollinators it needs to reproduce. Outcrossing instead of selfing (self-pollination) prevails among flowering plants. Mating with nonrelatives increases genetic diversity by spreading favorable and subsequent genes through the population, thereby enhancing the fitness of the next generations of plants.
To achieve that outcrossing, flowers broadcast a powerful message to potential pollinators. Flowers are perhaps a little bit of everything, but they are, in essence, the sex organs of the plant. They need sexual favors to spread their sperm and have their eggs fertilized. Natural selection has made them into some of nature’s most successful advertisers. Their petals act as living billboards, deploying an extraordinary array of shapes, sizes, color palettes, and scents to communicate the treasures awaiting their sexual enablers. Texture matters, too. They may have matte or glossy finishes, smooth or hairy surfaces, depending on the preferences of the animals they need to attract.
Although we humans have come to love looking at and experiencing flowers in our own way, their petaloid messages were not meant for our eyes or noses. Instead, they have been beckoning for millennia to the tiny insect aerialists that became the first fliers on the planet, taking to the air during the Devonian age, which began over 400 million years ago. The two trajectories combined in a later age and have had a long and fascinating history that is by no means finished now.
Plants have not always had flowers. Certain plants, the angiosperms, “invented” flowers and never turned back. They chose wisely. (As I hope is obvious, this is anthropomorphic shorthand for a complicated set of biological processes, for the plants did not make decisions; they tried everything, and natural selection [survival of the fittest] ensured that their genes were the result of the most successful “experiments” and were reproduced.)
Flowers evolved from small leaves that bunched together at the stem tips. When they changed later and took on roles that were very different from those of their leafy progenitors, they lost their green color and began developing into the petal and bract structures we now recognize within flowers. These were their come-hither beacons and perfume dispensers, which changed over time. As traits and their gene frequencies within populations varied, they were inherited one generation to the next, creating the best available fit of flower to animal. These “matchups” are known as floral mutualisms, forms of tug-of-war, a give-and-take whereby flowers and their pollinators are mutualists in a coevolutionary endeavor that does not cross the line into antagonism. Bees prefer sweeter nectar and choose flowers with more sugar. Gradually, in their descendants, flowers with richer nectar prevailed. This is also brinkmanship. If flowers offer too little, the bees won’t come; too much and they’ve wasted precious energy. Flowers and insects are mutualists, but they are first and foremost looking out for themselves, satisfying their own needs.
Since flowers don’t get around much—unless they’re in a wedding bouquet or a high-school-prom corsage—they found ways to have sex with other plants. Not a fling or an affair, but the real thing. The animals that fly from flower to flower, along the way dispersing each flower’s sperm (in the form of pollen) to other flowers, enable plants to go on virtual dates with each other, and to reproduce. Pollinators afford a wonderful solution to their mobility handicap, and one that nature began to put into action over 100 million years ago, when the earliest flowering plants came into being.
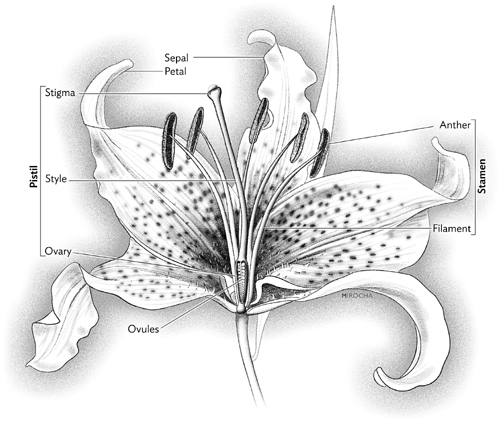
To understand the true meaning of flowers you need to be familiar with a few terms for the floral parts. The natural objects we call flowers might call on you to have a fresh-cut flower nearby as you read. Let the blossom be your guide. A lily flower would be ideal, which is why I open the chapter with the Stargazer. It’s readily available, with parts big enough to be seen without a magnifying glass, and it’s not coy about sharing the secrets of its sexuality. Although other common flowers such as roses, carnations, peonies and chrysanthemums share the same basic anatomy, they are perhaps too ruffly. Parting those floral skirts to get at what lies beneath might be a bit challenging to the amateur botanist voyeur. So let’s consider the simple lily, a big, brash beauty.
As mentioned, it is a hermaphrodite. Starting from the top of the flower, we see that long, tumescent rod, the female sex organ. It’s called the pistil (or carpel), and it has three sections. The first part you see, consisting of the style (the slender stalk) and the stigma (at the tip), rises straight up and out of the third part, the ovary, found below the petals. Floral styles can be short and squat or, as in the Stargazer, long and thin. The stigma at the tip is moist or gooey to catch and hold the pollen that come to it courtesy of a hungry pollinating animal, or perhaps a passing breeze.
Sprouting from the floral base, and surrounding the stalklike part of the pistil, are the stamens. Rising on their own long, slender stalks, called filaments, they emerge outside the swollen green ovary. Depending on the flower, there may be a few or many. The Stargazer has six stamens. Atop each are two structures that look like conjoined hot dogs. These are the anthers, containing thousands of dustlike pollen particles. As the flower matures, the anther walls peel back and release their precious cargo. Microscopic in size, and spherical or football-shaped, the pollen grains are yellow or orange, but, depending on the plant, may also be red, green, blue, purple, white, or nearly black. They’re covered with an oily coating that makes them stick together, so that they cling to pollinators—or to a probing finger. The grains are ornamented with furrows, ridges, pores, bumps, and spines arranged in patterns almost as uniquely distinctive as human fingerprints, which enable botanists to tell what family, genus, or species of plant the pollen came from just by looking at the pollen under a microscope.
Beneath the pistil and the stamens are the flower petals, and below the petals are the sepals, which are often green. In the case of the Stargazer, however, the outer whorl of what appears to be the petals is actually the set of three sepals, for in some flowers, including lilies and amaryllises and orchids, the sepals and the petals look almost identical. Sitting immediately above the sepals is the enlarged green ovary that is the bottom part of the pistil. If you cut through the ovary crosswise, you find the chambers containing the Stargazer’s ovules—its future seeds, which look like tiny peas. Depending on the plant, there can be many ovules, or just one, but regardless of how many there are, within each ovule is an embryo sac, which contains the egg and the other reproductive cells.
This weird-sounding reproductive action is so unique to angiosperms—the flowering plants—that it is one of the traits used to characterize and identify them. It’s called double fertilization because it involves not one but two sperm cells, one that fertilizes the egg, and the other to help form the endosperm, the starchy tissue that feeds the developing plant embryo. Fertilization begins when the pollen grains land upon the gooey stigma, the tip of our lily, after which things happen quickly. Each pollen grain swells up, nearly ready to burst, until it germinates, and one part of it sends a thin tube down the floral style, probing its way toward the ovary, while the other part, a generative cell, travels down the pollen tube. Finally, after several hours or a few days, as the cell approaches the embryo sac of the ovary, it divides into two sperm cells—ready to play their roles in double fertilization. The chromosomes from one sperm cell fuse with the egg cell that will develop into the tiny plant embryo found within every seed. The chromosomes from the second sperm cell fuse with different cells inside the embryo sac, developing into the endosperm.
For fertilization to occur, outside pollinators are enticed to visit flowers. But there has to be some reward for the pollinators—and some way of advertising that reward—or else they won’t bother to visit the blooms.
So, flowers fulfill many needs for their animal visitors. For bees as well as some other pollinators, flowers are cafés and rest stops, offering a rich abundance of food. They can also be rendezvous sites for pollinators to find each other and pair up. For predators, including crab spiders, ambush bugs, and mantids, wildflowers and garden blossoms provide ready-made perches from which to hunt their prey. Many insects seek out flowers for the warmth they find inside. This is especially true in arctic and alpine regions. The parabolic-shaped petals of these flowers not only warm the heat-seeking creatures but also speed up the development of ovules within flower ovaries. The remaining floral rewards consist mainly of nest-building materials and sex pheromone precursors in the case of some bees and butterflies. Floral oils, for example, can be eaten, but some tropical bees also turn them into protective linings for their brood cells. Fragrances produced by orchids and other sources are the pheromonelike precursors harvested by male orchid bees and used in their aerobatic courtship and mating rituals.
Of course the single most important reason insects and other animals visit flowers is to feed. Within flowers, they find nutritious food for themselves or their larvae. The adults of most flower-visiting insects seek food only for themselves. This gives them the energy they need to fly or stay warm. But other insects, including bees and certain specialist wasps, bring the pollen back to the hive or solitary nests, where the protein-rich grains feed them and also provide nourishment to their developing brood.
Flowers produce four types of rewarding food substances to draw the pollinating animals, hold their attention, and keep them coming back for more. These foods are nectar, pollen, floral oils, and the edible body tissues of some flowers.
Nectar is the sweet but watery liquid secreted from glands located near the base of the petals of most flowers. A high-energy food, nectar oozes up from hidden nectar glands into crevices deep inside most flowers, but can sometimes be produced elsewhere within flowers or from buds and leaf bases. Nectar is a high-energy, liquid food usually hidden from view. The sugars reward bees, wasps, flies, butterflies, moths, birds, and bats, but to reach those low-lying pools of nectar they have to brush up against the stamens, causing pollen to stick to them—ready to be transmitted to the next flower they visit. Floral nectar is a blend of three sugars—sucrose, glucose, and fructose—but may contain other minor sugars. When the pollinators eat and digest the sugars in the nectar, they get the energy they need to fly, or to stay warm in the cold. Certain social bees, such as honey bees, bumble bees, and stingless bees, are able to turn the nectar into the sugary “fossil fuel” we know as honey, which can be saved for the future. Honey bees visit their waxen cupboards to reclaim this stored energy during cold winters as well as periods of inclement weather.
Nectar, while an important fuel food, is not as critical for bees as the pollen that they obtain from flowers. They can’t live without it nor feed their young. Pollen contains from 5 to 60 percent protein, all of the essential amino acids, diverse lipids, and antioxidants that bees need. It’s not so easy to get at those nutrients, however. Pollen grains, tiny as they are, are actually supertough. Imagine them as hard-walled Ping-Pong balls with a nutritious liquid center. Their outer walls are a biopolymer, a natural plastic. These walls are so resistant to chemical attack that we find pollen grains fossilized in 100-million-year-old sediments. But that doesn’t stop the bees, which use a special trick to eat the otherwise indigestible pollen. They extract the pollen interiors with a combination of osmotic shock, which causes the grains to swell up and burst, and enzymes from within their digestive tracts, which break them down further allowing their nutrients to be absorbed. Once eaten, the undigested outer walls of the pollen are defecated by adults and their larvae. Looking at this waste product, we can perform a bit of forensic scatology to tell what types of pollen the bees ate, thereby knowing what plants they’d visited. I know this probably doesn’t do a lot for you, but it’s exciting for bee scientists such as myself.
Just as an aside: While pollen is the ideal balanced natural diet for bees, it may cause problems for mammalian digestive systems. Those who buy bee-collected pollen from health-food stores face a possible food-allergy risk because this pollen may come from ragweed or other allergenic plants. So, you might want to sparingly test a few pollen pellets before heaping them on your breakfast cereal. Honey is a different matter, however, since it contains only the faintest trace of pollen and may actually benefit hay-fever sufferers.
Another category of food offered to pollinators is super-high-energy floral oils contained within exotic flowers in the world’s tropical rain forests and deserts. These oils, which are used by specialist bees in the genus Centris, come mostly from flowers in the tropical family Malpighiaceae. These flowers bear unusual secretory glands on their undersides. Beneath a protective skin covering, like a blister you might get from gardening without gloves, the secretory glands contain oils with unique chemistries that are sought out by bees. Female bees land on the oil flowers and take up a four-legged stance. If you could slow down the rapid action, you’d see the bees raking their front legs across the oil glands. Their legs have broad, flattened scraper hairs that rupture the oil blisters and transfer the oils to the mats of thick hairs, almost like scrub brushes, on their back legs. When they fly home, they mix the floral oils they’ve collected with pollen, often from other kinds of flowers, creating a nutritious “bee bread” for their hungry larvae. The oil has another use, too. Some bees enzymatically convert it into a slow-hardening wall coating for the underground chambers, the brood cells, where the bee larvae grow and mature into adult bees. Other chemicals in the oil may protect the developing bees from the ravages of soil-borne bacteria and fungi. Nest-building materials may also come from the flowers that grow on the tropical Clusia tree. Its flowers are a sticky mess because they have glands that ooze tacky resins that attract pollinating bees, which harvest the resins for use in their nest-building.
Perhaps the strangest floral rewards of all (at least in terms of their presentation and later use) are the essential oils—terpenes and similar chemicals—produced by specialized orchids, e.g., Stanhopea, Coryanthes, and Gongora. These orchids produce no edible rewards—not a trace of pollen or nectar for their bee pollinators, the emerald-green or cobalt-blue beauties known as euglossines. Since these orchid bees are native only to the neotropical forests—from southern Sonora, Mexico, to the rain forests of the Amazon basin—you may not have had the pleasure of seeing one of these living jewels. The male orchid bees, never the females, visit the orchids, and while they are paying their floral visit, the bees probe for nectar that isn’t there, and for pollen that does exist, but is so securely shrink-wrapped inside testicle-shaped packages (known as pollinia) that it is unavailable to them. But during the bees’ search for food, the orchid flower, whose shape has been molded by millions of years of fine-tuning, maneuvers them into exactly the correct position for the bright yellow pollinia to be glued securely to their bodies, ready to be carried to the next flower they visit. Practicing a bit of thievery of their own, the bees scrape assiduously at the scent-producing tissues on the orchid blossoms, which they stash inside hairy glands within their enlarged, hollow hind legs.
After days, weeks, or months of collecting these fragrant substances, the male orchid bees form leks. These are display areas where one or more males put on a show for choosy females. An example of this kind of courtship in birds is the prairie chicken, also known as the sage grouse. The male birds puff out their breasts, call, and strut their fancy tail feathers in front of females, who choose their eventual mate based on the best performance. We think much the same thing happens in orchid bees. Male orchid bees flash their colors, buzz loudly, and puff out little clouds of sex pheromones—the fragrances they liberated from the orchids, which they have turned into aphrobesiacs, or at least that’s what we think is happening. We don’t know quite a few things about the orchid/bee pollination arrangement, but our best guess is that the bees somehow modify, mix, or blend the purloined orchid scents for use in their own mating rituals.
Versions of this orchid/pollinator arrangement occur around the world, especially in tropical rain forests. Typically, the orchids advertise that pollen or nectar is present when, in fact, most of these offerings are a sham—just clever advertisements. Pollinators are invited in, only to end up branded with bright yellow pollen sacs, glued on, for their trouble. They must go elsewhere to satisfy their hunger or sexual lust. In the world of flowers, what pollinators see is not always what they get. The guiding principle of caveat emptor in human commerce applies equally for animals visiting flowers.
The last of the food rewards offered by some groups of flowers are food bodies on the flowers themselves. At the tips of thickened petals called tepals are protein-rich, knobby structures that sap and rove beetles like to eat. A familiar garden flower offering this reward system is the eastern and western spicebush (Calycanthus). Small beetles are attracted by the dozens to their wine-colored blossoms. Beetles can’t resist and force their way through the maze of fleshy petals and down past the stamens. Spicebush blossoms offer no nectar but feed their beetle visitors with pollen and with these small, whitish food bodies. The beetles are in no hurry to leave. They feed, mate with abandon, and defecate, before taking flight in search of the next blossom.
Scent production that sends fragrances wafting through the air is a powerful way for flowers to advertise their presence to the creatures they need for their pollination. So it won’t surprise you to discover that among their other talents, flowering plants are master chemists, scenting the air with their aromatic petals and other floral parts.
Floral scents are usually produced at the time the flowers first open, and the intensity of the fragrance will vary depending on the time of day. In general, flowers tend to release the maximal amount of scent in hot, sunny midday weather.
You can do an experiment yourself to verify this. If you come across a kind of flower that you remember as having a scent, but you find that, upon a casual first sniff, you can barely smell anything, clip some of its petals and put them in a clean, small jar with a lid and leave it in a sunny window for an hour or two. When you come back, open the jar and sniff. The flower’s scent should now be concentrated and easily detected.
Not every flower effects its perfume alchemy in daytime, however. Flowers release their scents when their pollinators are most active. As gardeners who have created so-called moonlight gardens know, certain blossoms—e.g., jasmine, moonflower, queen-of-the-night cactus, sacred datura, and other nocturnal bloomers—release their sweet essences onto the night breezes. Both the queen-of-the-night cactus and the datura’s large, white blossoms are visited by fast-flying hawk moths that follow the floral odor plumes to the flowers at sunset and into the night. Also, some temporal rhythms in scent production have to do not with the time of day but with the age of the flower. Fresh, young blossoms quickly peak in their scent production, when their stigmas are most receptive. As the flowers age and eventually wither, their fragrance production drops until it eventually stops.
The nature of the different floral aromas broadcast by flowers is related to the types of pollinators they’re trying to entice. For example, let’s take a look at—or rather a sniff of—a potted African Stapelia. If you’ve ever owned one of these and had it bloom on your kitchen windowsill, you may remember that until you finally realized the smell was coming from your houseplant, you went looking high and low throughout the kitchen for whatever had died—a mouse? a rat? whaaaaat?! Plants such as Stapelia, and its relatives among the African milkweeds, as well as skunk cabbage and the voodoo lily, depend for their pollination on carrion flies. These flies, which normally feed on dead animals, are attracted to the nitrogen-containing aminoid compounds released by carrion during decomposition. In 1885 Berlin physician and chemist Ludwig Brieger, a man with a good nose and a keen sense of humor, gave these pungent molecules the evocative names by which we know them today—putrescine and cadaverine. These are the source of the unpleasant and unforgettable smell of death. Putrescine also plays a role in giving urine its distinctive scent.
Remarkably, natural selection enabled flowers to develop and make the same fetid-smelling compounds that carrion releases. I find it fascinating that these carrion flowers are able to tap into the smell of death, evolving in a way that they, too, could produce putrescine and cadaverine and use them to advertise their presence to the carrion flies they need to achieve their sexual unions.
Each pollinator has its own preferences for scents, and is associated with certain flowers that evolved to adapt to those preferences. Butterflies prefer the light, sweet aromas emitted by phloxes, Polemonium, and similar flowers. Working the night shift, settling moths and large, fast-flying hawk moths are attracted to blossoms such as jasmine, which have heavier, almost sickeningly sweet fragrances created by benzyl acetate and linalool (among a hundred other compounds). Flowers pollinated by bees have lively and sweet but somewhat fainter scents. Think of the scent of snapdragons, wild peas, Penstemon, or foxgloves. Often these flowers are found in the same area, where their use of similar chemical components to entice their bee pollinators makes for an economy and efficiency that suits all parties in the interaction. In addition to colors and shapes, flower-specific scents help pollinators find the same floral species. This is mutually beneficial to flowers since their pollen is moved to unrelated individuals of the same species, not wasted on the wrong blossoms. Floral fidelity, but to different individuals within a plant species, works for both partners.
Among the vertebrates and their flowers, the situation is quite different. Most of the pollinating vertebrates are birds and bats, and except for vultures and condors, most birds have an extremely limited sense of smell. So hummingbird-pollinated flowers, for example, are essentially scentless. These vivid red flowers, including Salvia, Hibiscus, and Penstemon to name a few, may look lovely in your living room, but they won’t do anything to lend it a pleasing aroma—something to keep in mind when you see beautiful bouquets of tubular, red flowers in the florist shop.
Bats, however, are among the vertebrates that enjoy floral scents. Flowers attract neotropical nectar bats for pollinating visits with batlike chemical-messenger odors, either sulfurous or fruity. Most bat odors don’t do anything for me, but I do enjoy some of the fruitier scents in some bat blossoms. A favorite scent not just of the bats but of mine is from the lofty saguaro—often growing seventy feet tall, a columnar cactus and the state flower of Arizona. Pollinated by nectar-loving bats at night and cactus bees by day, the saguaro has a floral aroma that reminds me of the lovely mild fragrance of a ripe honeydew melon. The bats seem to enjoy the smell as much as I do, greedily lapping up the saguaro’s dilute but abundant nectar as they spread cactus pollen around.
Most animals have a strong response to smell, which can play a major role in their reproductive life. But some researchers have suggested that flowers today are losing their fragrances due to climate change, and that this will disrupt links in pollination and affect our food supply. This, like so many other concerns about global warming, increasing levels of UV radiation, and other climate-change effects, should be explored more thoroughly by environmental scientists before it is too late for flowers, pollinators, and us.
Smell matters to most pollinators and so, too, do tactile sensations. The nature of surface textures has a lot to do with which plants attract which pollinators. Some floral petals are smooth even when viewed under an electron microscope. Others are noticeably hairy, with a soft matte finish. Many are thick and waxy. Others have unique microscopic layers and other features that produce a characteristic visual effect that attracts the targeted pollinators. An extreme example of a plant with reflective petals, their surfaces almost as shiny as those from a high-gloss wax applied to a prized automobile, is the buttercup, the genus Ranunculus, with about six hundred species. Its glossy petals are shinier than most others in the plant world, so shiny that if you hold a buttercup under your chin, it will reflect its yellow glow onto your skin—as you may remember from your childhood, when lore had it that if your chin turned yellow, it meant you liked butter. Bees, like children, are drawn to the specular reflections and highly directional sheen of buttercup petals.
Recently, it was discovered that bees, and likely other pollinators, are able to discriminate between flowers, or between adjacent microscopic surface textures within the same flower, solely on the basis of touch. Using the sensitive sensory cells on their legs and mouthparts, bees can differentiate between the subtle surface textures of leaves and flower petals. In one experiment, petals from different blossoms were dried and sputter-coated with gold atoms (to eliminate any possibility of color, odors, or other conflicting stimuli). Honey bees and bumble bees were trained to associate a sugar reward with one surface type, either bumpy or smooth. After a few trials, the bees clearly and easily discriminated between the petal surfaces by using only cues in their microscopic surface textures.
Imagine a highly charged bumble bee landing on a flower, then sensing the flower’s electrical condition—and possibly being able to draw useful information from it. This isn’t science fiction. In spring 2013, researchers at the University of Bristol’s School of Biological Sciences published a paper describing their discovery of a previously unknown flower-to-bee signaling modality: electrical signals.
We’ve known since 1929 that plants have electrostatic fields. Plants and their flowers are both literally and electrically grounded, which means that flower petals and pollen grains have weak, typically negative, electrical charges. We also know that every object flying through the air, whether it is a Boeing 787 Dreamliner jumbo jet, a baseball, or a bumble bee, acquires a strong positive electrostatic charge from friction with the air. A bee might be carrying a charge as high as 450 volts. The waxy surface cuticle of the bee’s body helps hold on to the charge, which discharges and dissipates slowly. So what happens when our positively charged bee lands on a negatively charged blossom? What kinds of interactions might occur between these electrical opposites? Can the bee sense patterns and changes in the flower’s negative charge, somehow using that information to its advantage? Conversely, could flowers be using their electrical charges to advertise their food supplies to the bees?
Working with Professor Daniel Robert at Bristol, Dominic Clarke and Heather Whitney detected patterns in the electric fields on the sexual parts and petals of certain flowers and speculated that the bees might be able to detect those patterns and use them as guides to the flowers’ stores of pollen and nectar—much in the way that bees use ultraviolet-light patterns on flower petals to lead them to food (as you’ll read about below).
By placing electrodes on the stems of petunias, the Bristol researchers discovered that when a bee lands on it, the flower’s charge changes for several minutes. From my own research on electrostatic charges on flowers and bees, especially buzz-pollinated ones, it seems plausible to me that by leaving this “electric footprint” when they land on a flower and feed, bumble bees and other bees might passively be “labeling their empties”—so that the bee, and its hive mates, would not make the mistake of returning to the same flower too soon. After all, why bother with a flower you just visited, likely one already emptied of its nectar and pollen, when you can quickly move on to virgin territory? I agree with the Bristol researchers that it makes sense that bees might be using the electrical signals naturally present on flowers or left by the bees themselves.
I’m reserving judgment, however, about whether these electrical differences can be actively produced by flowers, perhaps fine-tuned for bees, until other behavioral studies are completed. But nature has unending surprises, so almost anything seems possible.
Flowers blush. It may seem remarkable to say this, but it’s true. Many flowers change colors after they are pollinated. This physiological shift happens because of a pH change from basic to acidic within their petals, possibly triggered by pollination or subsequent fertilization. In other flowers, colors often change with age, transitioning from a young, rewarding bloom (rewarding to a pollinator, that is) to an older, senescent one. In lupines, a white spot at the base of the banner petal blushes pink as they age. In the Mexican paloverde, a small legume tree that grows in the Sonoran Desert around my home in Tucson, Arizona, the uppermost petal, the banner petal, turns from yellow to a bright red. In red-hot pokers (Kniphofia), the flowers start out a brilliant red then become yellow close to the end of their lives, when they are no longer secreting nectar. Many garden or wildflowers show similar color changes. Unlike demure, young human female blushing beauties, flowers tend to do their blushing toward the later stages of their lives. These floral color changes are their way of shouting out their new status as they move from floral adolescence into maturity, senility, and finally death. The yellow to red shift is the most common signal to pollinators that the flowers are no longer rewarding, but others may use different colors as their signal. Whatever the colors, the flowers might as well be holding up a sign that reads NO FOOD HERE, PLEASE MOVE ALONG.
This is an efficient arrangement for both partners. The color-shifted flowers have already been visited and likely fertilized. Further bee visits would be wasted on them. For bees it means that they can learn quickly whether the next flower is likely to contain nectar or pollen, which allows them to become more efficient foragers, visiting flowers that are rewarding while skipping those that aren’t. As you’ll read below, most bees don’t register the color red, so although the shift to red make these flowers more visible to humans, it may lessen or somehow change their visibility to passing bees.
Color is one of the main signaling strategies flowers use to beckon their pollinators. Floral colors also serve as filters, selectively attracting specialist pollinators to certain blooms while eliminating others. For many of those pollinators, the brighter the better, which must be why floral colors, especially those created by the petals, are among the most highly saturated colors found in nature. Technically speaking, saturation is the strength of any given color in relationship to its own brightness. The purest, most highly saturated colors are of one or a few close wavelengths and contain almost no white light (the combination of all wavelengths), while unsaturated colors appear faded or washed-out because white light has diluted the colors. Saturated colors can be equated with their perceived intensity.
The colors of a flower are determined by the interaction between light (from either the sun or the moon) and the floral pigments inside its petals. Air spaces and fluid-filled vacuoles within the petals also play a role. The pigments are of at least three types—carotenoids, betalains, and flavonoids (which include the red- and sometimes blue-reflecting pigments, the anthocyanins). When these pigments interact with light, they absorb certain wavelengths and reflect others. Those that they reflect determine the colors that we see. For example a red rose is red because its various colorful anthocyanins selectively absorb blue and green light from the sun, while reemitting the red light that hits its petals. A deep blue larkspur (Delphinium) is blue because delphinidin, an anthocyanin within its tissues absorbs red and green wavelengths but reflects blue. While the anthocyanins tend to produce red and blue colors, carotenoids are responsible for the oranges and yellows (as well as some of the reds) we see in various flowers, and also in carrots and tomatoes. Betalain pigments produce some of the most highly saturated and beloved reds of all, as in certain cactus flowers. You also know the betalains from common garden beets, Portulaca blossoms, amaranth stems, and the prickly pear cactus.
Think of the last rainbow you watched as it winked briefly into view before evanescing into the vapor. Its brilliant colors ranged from red to orange, yellow, green, blue, indigo, and, finally, violet. The reds are at the longer-wavelength side of the color spectrum, with violet at the opposite, short end. However, that part of the rainbow you are able to see occupies only a small middle portion of the wider electromagnetic spectrum, with light continuing beyond red into the infrared at the far end of that spectrum, and ultraviolet at the opposite end. Human beings cannot see colors in the infrared or, as we’ll discuss below, in the ultraviolet.
Like us, birds and insects and other animals are capable of sensing only part of the electromagnetic spectrum. That much—the fact of being limited—we have in common. Yes, bees and people both have trichromatic vision, which means we each detect and respond to three main color regions due to three types of sensory cells, but which three color bands is where we differ, and that in turn means that those regions of the visual spectrum we see, and how we process them, differ. The result is that what bees view of their world differs greatly from what we see. For example, bees are not capable of seeing either red or infrared light. Therefore bees (and most other insects, too; except perhaps for the Asian honey bee Apis dorsata, a few butterflies, and buprestid beetles) are red-blind; for them, the brilliant red flower colors we have been talking about might as well be black, the absence of colors. But their lack is a signal, too, as we discussed, because it allows them to focus on the more visible—and more rewarding—flowers.
While bees, including the familiar honey bees and bumble bees, don’t see the infrared part of the spectrum, they as well as ants, wasps, and other insects do see ultraviolet colors. The sensory regions of their eyes allow them to sense ultraviolet light at wavelengths measuring between three hundred and four hundred nanometers. (For us, these UVA longwave and UVB shortwave wavelengths fall just below our visual detection limit—though they do have an effect on us, since these are the ranges that can give us a sunburn, tan us, or lead to skin cancers.) That wasps, ants, and bees can sense the presence of ultraviolet light was first noticed by the great naturalist Sir John Lubbock in 1877, during his observations on British ants; and that flowers reflect ultraviolet light was proven photographically and discussed in a 1922 article by Americans Frank Lutz and F. Richtmyer.
Humans—except for infants less than three months old, whose crystalline lenses within their eyes have not yet stabilized—are generally unable to see the ultraviolet patterns visible to bees and other flower-visiting insects. However, we can get around this handicap by a bit of scientific sleight of hand, commandeering the technology inside special cameras, sensors, and filters to give us at least a sense of what the bee’s eyes can see.
If we took one of these devices, say an ultraviolet-sensitive camcorder, and went on a field trip to a colorful meadow or local garden, how would our favorite flowers look as we aimed the camera at them? Would we even recognize them as the same flowers?
Panning around the garden with our newfound bee vision, we discover that bright yellow flowers are excellent places to look for the ultraviolet floral patterns that we can’t ordinarily see. Of these, members of the large and common sunflower family (Asteraceae) give us particularly vivid displays of the UV signposts we’re looking for. So let’s consider one of them—the familiar black-eyed Susan (Rudbeckia hirta), which is abundant throughout the eastern United States.
A sunflower relative (Bidens ferulifolia) in visible light (human view, left) and ultraviolet light, as seen by insects, on the right
Without bee vision what we would see are long, narrow yellow petals surrounding the dark brown center of the sunflower “face.” Bees, however, see something quite different. Though they do see the dark brown center, they don’t see the petals as a homogeneous yellow color. Instead, the bottom portion of the petals, the part closest to the center, looks “dark” to a bee because it absorbs UV light while also reflecting yellow. In contrast, the uppermost portion is reflecting yellow plus a little or a lot of UV and looks bright. What this combination of yellow and UV wavelengths actually looks like to a bee we don’t know, but insect neurobiologists call the resulting color bees’ purple. The dividing line between bees’ purple and plain yellow appears about halfway along the length of the petals. Viewed in its entirety, the sunflower face has two rings of different colors surrounding the dark brown center, making for a dramatic bull’s-eye effect, and turning the flower into a gigantic signaling unit, all the better to catch the attention of bees, butterflies, flies, and beetles. Many yellow flowers have ultraviolet-enhanced bull’s-eye patterns similar to that found in the black-eyed Susan or the Bidens above.
But other flowers have non-bull’s-eye patterns created by ultraviolet light that vary in ways that also speak eloquently to their pollinators. When we turn our techno-gaze upon the lowly marsh marigold (Caltha palustris), an herbaceous plant of temperate, wet woodlands, we see thin, wavy, dark UV-absorbing lines radiating outward on the petals. These UV-absorbing markers, set against their reflective leafy backgrounds, help direct bees to nectar produced but concealed from direct view in crevices at the petal bases.
I have studied an Arizona plant, the Mexican yellow-show wildflower (Amoreuxia palmatifida), which may be the world’s UV champion. Reflecting 90 percent of ultraviolet wavelengths from their large, curved orange petals, these flowers are highly saturated color beacons of amazing intensity, living signposts beckoning to its bees. How could any bee resist dropping in for a closer look? Amoreuxia flowers are visited and buzz-pollinated by carpenter bees, bumble bees, and a few other smaller native bees.
A visit to an orchid fancier’s greenhouse reveals many other examples of floral patterns created by ultraviolet light. Orchid flowers, especially those of the neotropical Oncidium, have broad lower lips that reflect large amounts of UV light, signaling fast-flying Centris bee pollinators in their neotropical forests.
Imagine a bumble bee flying low over an alpine meadow in riotous bloom somewhere in the Colorado Rockies. Looking down, it sees hues of green and brown vegetation punctuated by highly attractive points of saturated light. Those lights popping in and out of view across its visual field indicate single flowers or clumps of plants in bloom. The bee takes notice. Bees in flight can detect objects much better than we can when we are in rapid motion. In fact, researchers Peter Skorupski and Lars Chittka have recently demonstrated that bumble bees have the fastest color vision of all animals. These bumble bees see the world almost five times faster than we do and can therefore make extremely fast decisions about how to respond even to moving objects while they themselves are in rapid motion, flying at 15–20 mph. They, and likely other bees, can easily navigate through shady bushes to find food, escape predators, or apprehend potential mates midair.
They can do all this even though they see almost no details from far away. Unlike us, bees have to be extremely close to objects to make them out with any clarity, because from even a medium distance they see only a fuzzy image. In fact, bumble bees and honey bees can’t recognize that a colorful object is a flower until they are almost upon it, perhaps four inches away. Smaller bees (and other pollinators, too) possess even more limited vision and have to be closer yet to see and differentiate objects, including flowers. So forget those wondrous “mosaic” or kaleidoscopic views of the world that all insects supposedly have. This fantasy idea about insect vision, which has been perpetrated by movies, television, and magazines, is incorrect.
That said, the speed of bees’ vision is remarkable because a huge amount of processing is going on every second inside a bee’s tiny but powerful brain. The salt-grain-size neuropile (brain) of the familiar western honey bee, Apis mellifera, houses 960,000 neurons and a staggering number—109—of neural synapses. We, the brainy, naked apes, have 85 billion neurons to our credit, with synapses totaling 1015. Sounds like a lot, but remember that they are housed within a brain that is 1.2 million times larger than that of our flower-visiting honey bee.
To get an idea how the bee’s vision benefits from all that neural power within the bee’s brain, imagine a honey bee flying into a movie theater, then pausing to watch a Hollywood blockbuster. Watching a movie is nothing more than fooling the brain into thinking that a series of individual static images (frames, as they’re called in the language of cinema) projected rapid-fire onto the screen is in seamless motion—which is similar to what happens when you riffle through the pages of a flip book. Behavioral scientists call this phenomenon flicker fusion, and the frequency at which these flashing images become smooth, fusing into a steady image for the observer, is the flicker-fusion rate, which is a function of how rapidly visual signals are processed by the brain and seen in our mind’s eye. For people, the flicker-fusion frequency is considered to be sixteen hertz—an image flashing on and off sixteen times every second. For bees and flies, their flicker-fusion rate is higher, by almost twenty times, an amazing three hundred times per second. A bee watching a fast-moving, action-packed thriller would probably wonder what all the fuss was about and be bored out of its mind by the slowly changing still images flashed on the big screen. (That is, if it could see as far away as the movie screen—which it can’t.)
The reason for this digression into the physics and physiology of bee vision is that what bees see has had a powerful impact on why flowers look the way they do. From the distant past to the present, bees and other insects, along with a few birds and bats, have been the primary selective (natural-selection) forces causing wildflowers to have the diverse palette of colors that they do. Flowers signal to insect and other visitors—via food rewards, color, shape, size, texture, scent, etc.—and the animals respond. If everything goes right, bees and other insects carry away the precious pollen grains, acting as go-betweens for the plants they visit. The stronger the floral attraction and the more frequent the visitation, the higher the survival and reproductive rate of that flower, and its descendants.
Today’s wildflowers and flowering garden plants have colorful blooms to attract, guide, and hold the attention of bees and other pollinators. But they haven’t always looked the same. The world of big, bold, colorful, and lavish flowering plants or cut flowers from the florist’s shop that we experience today took a long time to develop, with painstaking breeding trials. So in the next chapter we will go back into geological time, long before the arrival of modern humans, and of many of the insects currently on the planet, to the origin of the earliest flowering plants and their blooms. We will discover when angiosperms first experimented with showy flowers, and how that evolutionary experimentation was encouraged when diverse pollinating creatures began noticing and using them for food, rest stops, and more. We will explore an ancient world of flowers trapped and preserved within golden amber, and others that were compressed into stone, leaving faint carbonized residues of their former glory. This is the story of the first flowers on the planet—known to us today only as the rarest of fossils.