Chapter 11
Incorporation and Bioconversion of Omega-3 Fatty Acids for Obtention of Enriched Fish
Oscar O. Santos, Jr.
Fabiana Carbonera
Julianna M. Vagula
Swami Arêa Maruyama
Jesuí V. Visentainer State University of Maringá, Maringá, Paraná, Brazil
Abstract
Farm-raised fish present low concentrations of polyunsaturated omega-3 fatty acids (n−3 PUFA). As the series of omega-6 (n−6) and omega-3 (n−3) fatty acids share the same enzymes, elongases, and desaturases, during bioconversion of the precursors alpha-linolenic acid (ALA, 18:3n−3) and linoleic acid (LA, 18:2n−6), the incorporation of ALA from diet favors the biosynthesis of eicosapentaenoic (EPA, 20:5n−3) and docosahexaenoic acids (DHA, 22:6n−3) in muscle tissue of freshwater fish and leads to an ideal n−6/n−3 ratio. The understanding of this metabolism has been essential to science, because while n−3 series possesses antiinflammatory functions, the excess of n−6 fatty acids affects proinflammatory processes. In this way, the incorporation of different vegetable sources of n−3 fatty acids in the diet of farm-raised fish, targeting the biosynthesis of long-chain n−3 fatty acids (n−3 LC-PUFA), becomes a viable alternative to exploit the technological and functional benefits of enriched fish meat for human consumption.
Keywords
fish
fatty acids
alpha-linolenic acid
omega-3
elongation
desaturation
1. Omega-3 Fatty Acids: Why Should They be Consumed?
The transformation of feeding habits of the population in recent decades is related to the increased consumption of foods with high sodium, simple sugars, trans fats, and cholesterol contents, along with a decrease in consumption of foods rich in vitamins and minerals. This transformation has led to an increase in the occurrence of chronic diseases (cancer, hypertension, diabetes mellitus) and their comorbidities, becoming a public health problem. Contrary to the emergence of these diseases, there is the research of foods and their compositions and/or fortification, in which the focus is the use of so-called functional foods, which act in the prevention or recovery of the human body.
The functional foods, in addition to normal nutrients, provide biologically active substances that are able to prevent the occurrence of diseases over time, if consumed according to daily recommendations. Among the various proven functional foods (oats, soy protein, lycopene, and others), this chapter will be more focused on functional foods rich in omega-3 (n−3) fatty acids, present in foods rich in lipids.
In general, fatty acids are the main components of lipids. They have an aliphatic chain and a carboxylic group, and can be classified according to carbon chain length, in short-(<8 carbons), medium-(8–18 carbons), and long(≥20 carbons) chain fatty acids. According to presence of insaturations, fatty acids can be classified as saturated fatty acids (SFA; do not have double bonds in carbon chain); monounsaturated fatty acids (MUFA; feature a double bond in carbon chain), and polyunsaturated fatty acids (PUFA; containing two or more double bonds in carbon chain) (Fig. 11.1). The insaturations in fatty acid carbon chain can lead to occurrence of cis- or trans-isomer configurations. The cis configuration, although being thermodynamically unfavorable, is predominant in the kingdoms Plantae and Animalia.
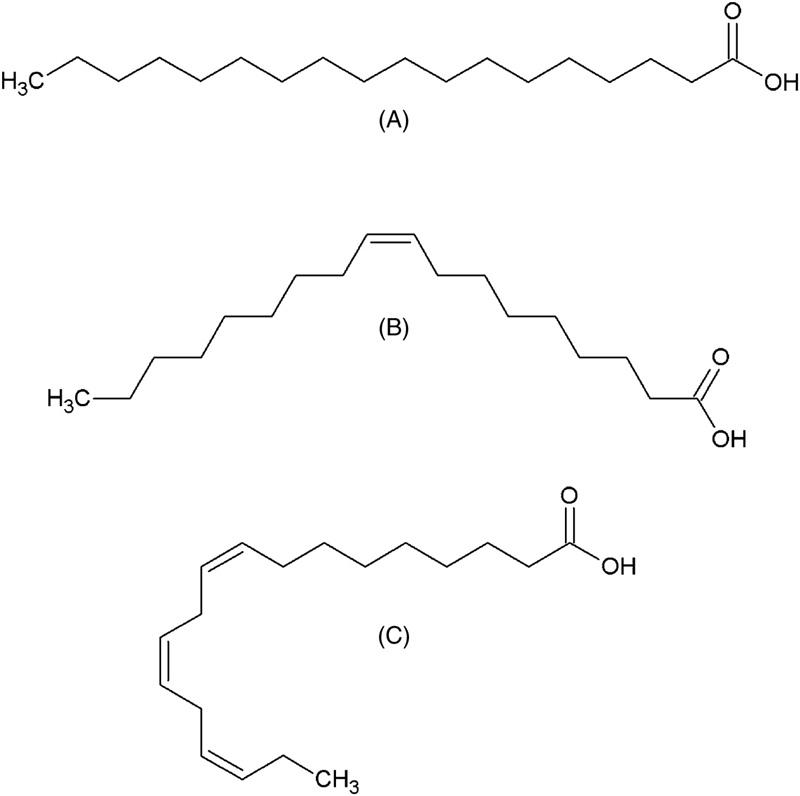
Figure 11.1 Chemical Structures of Fatty Acids.
(A) Saturated fatty acid (stearic acid, 18:0); (B) monounsaturated fatty acid (oleic acid, 18:1n−9); (C) polyunsaturated fatty acid (alpha-linolenic acid, 18:3n−3).
(A) Saturated fatty acid (stearic acid, 18:0); (B) monounsaturated fatty acid (oleic acid, 18:1n−9); (C) polyunsaturated fatty acid (alpha-linolenic acid, 18:3n−3).
Fatty acids are found in oils and fat, as free fatty acids and mainly in form of triacylglycerols (TAG). TAG molecules consist of three fatty acids (which may vary in terms of carbonic chain extension and number/orientation/position of double bonds), linked in form of esters to a molecule of glycerol (Fig. 11.2).
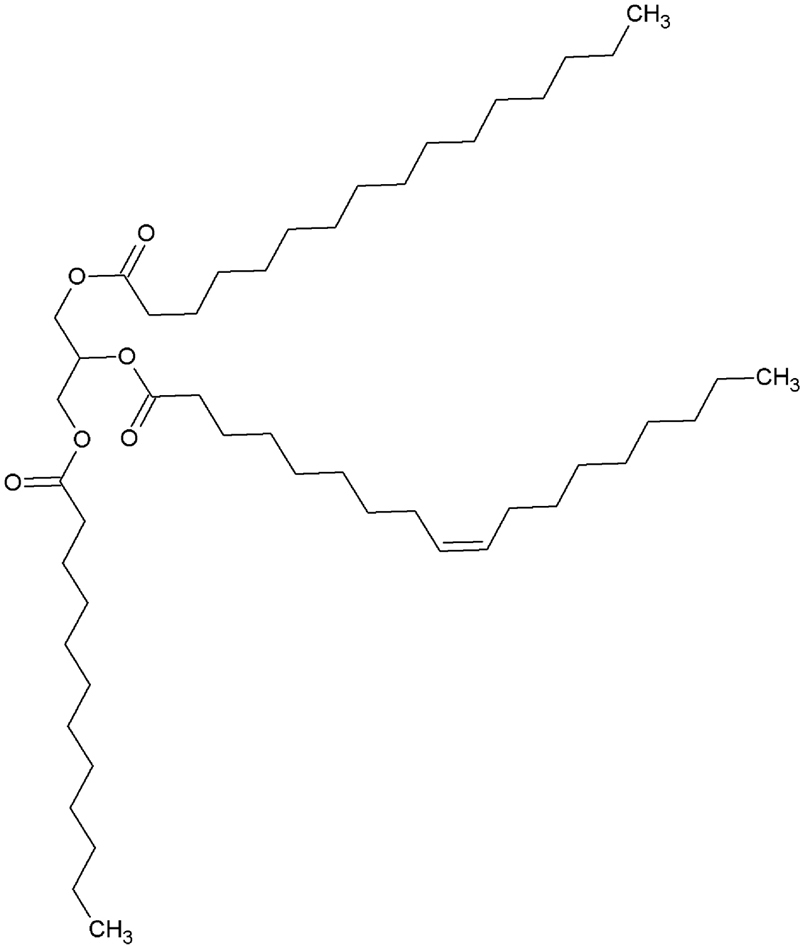
Figure 11.2 Representation of a Triacylglycerol Molecule.
Among fatty acids, two series of fatty acids, omega-3 (n−3) and omega-6 (n−6), stand out in nutritional studies for being the most important PUFA, including fatty acids with two or more double bonds and carbon chains containing 18 or more carbon atoms.
Regarding symbology for omega fatty acids, n or ω (omega) letters are used, followed by a number indicating the number of carbons, which ranges from terminal methyl group (CH3) to the first double bond in carbon chain of a specific fatty acid (Fig. 11.3).
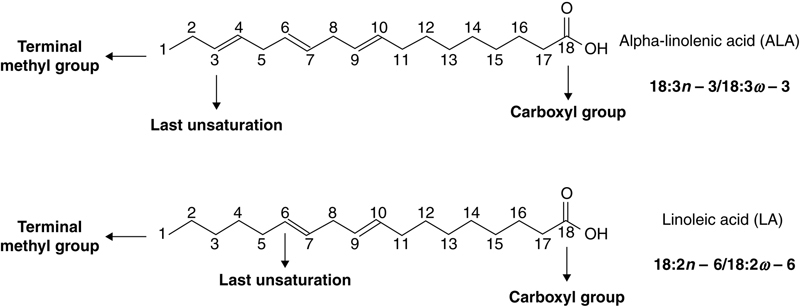
Figure 11.3 n or ω Designation of Fatty Acids.
The series of n−3 fatty acids is similar to n−6 series, because both have essential precursors for biosynthesis of long-chain polyunsaturated fatty acids (LC-PUFA). Alpha-linolenic acid (ALA, 18:3n−3) is the essential precursor of n−3 fatty acids series, while linoleic acid (LA, 18:2n−6) is the precursor in n−6 fatty acids series.
The search for functional foods, rich in n−3 fatty acids, is important due to the fact that these substances are essential to the human body. The essential nutrients are not synthesized by the human body from other nutrients, such as leucine, isoleucine, methionine, and phenylalanine amino acids, which are fundamental for protein metabolism. Therefore, the essential n−3 and n−6 fatty acids must be obtained through ingestion of adequate foods to activate the metabolic conversion of ALA and LA in other n−3 and n−6 fatty acids series.
Essential fatty acids (EFA) are part of phospholipid structures, which are important components of cell membranes. The fatty acid composition of membrane phospholipids is partly determined by the composition of ingested n−6 and n−3 fatty acids. Thus, the food lipid composition can influence various cell membrane-related functions, such as biosynthesis of hormones and activities associated with enzymes. Currently, it is known that LC-PUFA, especially from omega-3 series (n−3 LC-PUFA), are primordial in some stages of human development. In relation to biological function of n−3 LC-PUFA, their characteristics are widely known, especially when it comes to eicosapentaenoic acid (EPA, 20:5n−3) and docosahexaenoic acid (DHA, 22:6n−3) (Fig. 11.4). EPA is responsible for acting in antiinflammatory processes, while DHA acts on growth and functional brain development during the embryonic stage, and it is necessary for maintenance of normal brain functions in adults. The ingestion of DHA also leads to a learning ability improvement in children.
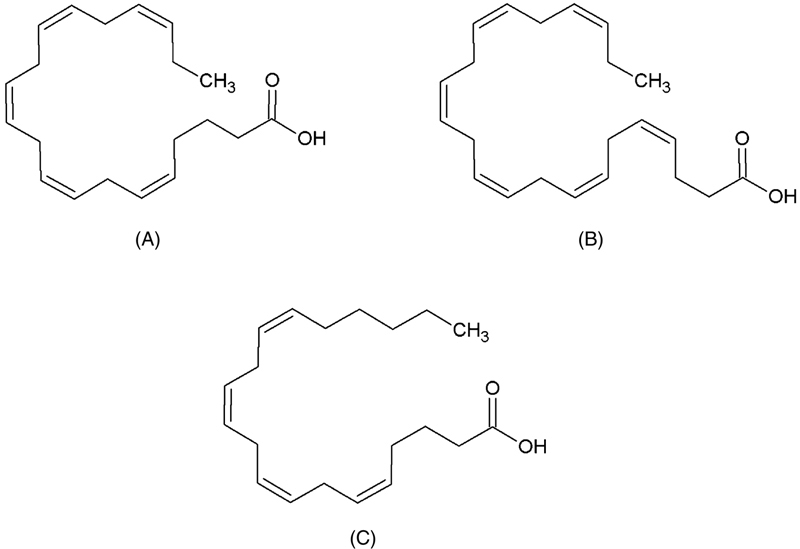
Figure 11.4 Chemical Structures of Long-Chain Polyunsaturated Fatty Acids.
(A) Eicosapentaenoic acid (EPA, 20:5n−3); (B) docosahexaenoic acid (DHA, 22:6n−3); (C) arachidonic acid (AA, 20:4n−6).
(A) Eicosapentaenoic acid (EPA, 20:5n−3); (B) docosahexaenoic acid (DHA, 22:6n−3); (C) arachidonic acid (AA, 20:4n−6).
In addition, EPA and arachidonic acid (AA) have very important roles in control and regulation of lipid metabolism, especially in relation to immune and inflammatory system, because they are precursors of eicosanoids. Eicosanoids are biomolecules produced from oxidation of its precursors. The two main enzymes involved in this process are cyclo-oxygenase, responsible for production of prostanoids derivatives (cyclic oxygenated derivatives), which include prostaglandins (PG), prostacyclins (PGI), and thromboxanes (Tx). And the second enzyme that participates effectively in oxidation of precursors is lipoxygenase, involved in production of oxygenated derivatives, such as leukotrienes (LT).
A balance in the intake of both n−3 and n−6 fatty acids is essential for a healthy diet as these two fatty acid series provide antagonistic effects. While metabolites generated by n−3 series show antiinflammatory, antihypertensive and antithrombotic functions, metabolites of n−6 series generate proinflammatory, arrhythmic, thrombotic, and hypertensive processes. In addition, eicosanoids derived from AA are biologically active in small quantities, which makes the imbalance between both series potentially alarming.
Lipids are essential nutrients to human body, an excellent source of energy (lipids provide 9 kcal/g), with extensive performed functions, such as: transportation of carotenoids and A, D, E, and K fat-soluble vitamins, and assist in hormone biosynthesis, among others. In addition, lipids present nutritional, structural, and regulatory functions, which have significant impacts on the most important physiological actions of the human body, as well as immune state and metabolism. This nutrient absorption takes place through action of gastrointestinal lipase and bile secretion, transported from intestinal lumen to microcirculation. In metabolism, when the body requires energy, triglycerides undergo lipolysis and free fatty acids are used, and carbon dioxide/water are obtained as excretion products.
Absorption of essential fatty acids (n−3) from diet occurs in the same way as other long-chain fatty acids. In metabolism, elongase and desaturase enzymes are used for conversion of ALA to EPA, and EPA is converted to DHA through β-oxidation in peroxisomes. EPA is incorporated in adipose tissue or used for biosynthesis of eicosanoids. The final metabolism products are carbon dioxide and water.
In humans, the biosynthesis of LC-PUFA is not very active, being limited by the bioconversion of precursor ALA to EPA, as well as DHA biosynthesis. In formation of n−3 LC-PUFA, the ∆6 desaturase enzyme performance is the limiting step of conversion because it is linked not only with ALA metabolism, but also with tetracosapentaenoic acid (24:5n−3) conversion. ∆6 desaturase enzyme is involved in two steps of the biosynthetic route, being used by two substrates, which leads to potential competitive inhibition of the enzyme. This caused inhibition might explain the accumulation of EPA and DPA (docosapentaenoic acid, 22:5n−3), but not of DHA when ALA dietary intake is abundant in humans.
In addition, the activity of ∆6 and ∆5 desaturase enzymes is reduced by factors, such as smoking, alcohol consumption, diabetes, stress, high intake of trans fats, and mostly by aging. Stress involves release of hormones, such as catecholamines and glucorticoides, which strongly inhibit ∆6 desaturase. In diabetes, low amounts of insulin inhibit ∆6 and ∆5 desaturases through RNA messenger modulation of these enzymes.
In animals, the 18:1n−15t, 18:1n−14t, 18:1n−11t, and 18:1n−3t trans-MUFA exert a strong inhibition on ∆6 desaturase activity, while 18:1n−15t, 18:1n−9t, 18:1n−5t, and 18:1n−3t isomers are more efficient in inhibiting ∆5 desaturase. Considering that partially hydrogenated fats present significant amounts of 18:1n−9t and 18:1n−5t isomers, a high intake of these fats can result in a significant decrease of ∆5 desaturase enzyme activity. In addition to these factors, an insufficient intake of proteins, zinc, magnesium, copper, and B3/B6/C vitamins contributes to reduce the conversion of LA and ALA into other PUFA.
In newborns, ∆5 and ∆6 desaturase enzymes are active, and even premature babies are capable of producing AA and DHA. However, human milk shows higher levels of AA and DHA in the first few weeks after childbirth, diminishing at a rate that depends on the presence of these fatty acids in diet. This condition suggests that the amounts of AA and DHA produced by the newborn are still insufficient, requiring their intake. With that, the preferential accumulation of DHA in humans depends on the dietary intake of this fatty acid.
Among food sources of n−3 fatty acids, fatty fish (herring, mackerel, salmon), vegetable oils (linseed, perilla, sacha inchi, canola), and nuts (walnuts, hazelnuts, almonds) can be cited.
The reference daily intake for adults, in human diet for lipid consumption must consist of 20%–35% of this macronutrient. The consumption of omega-3 fatty acids should be 0.6%–1.2% of total lipids for all age groups. Therefore, an adult who consumes about 2.500 kcal/day, can consume up to 875 kcal/day (97.2 g) of lipids, and 30.06 kcal/day (3.34 g) of omega-3.
It is observed that the intake of EFA must be stimulated, because during human metabolism multiple cells are oxidized, and as a result over time these oxidations can lead to coronary heart disease, arrhythmias, thrombosis, and inflammatory diseases. In addition, EFA deficiency is related to immune dysfunction, thrombocytopenia, alopecia, dermatitis, and poor wound healing. In pregnancy, DHA deficiency may be associated with insufficient cognitive and visual development of fetus. The main symptoms of omega-3 fatty acid deficiency are: reduction of visual acuity, increased propensity for skin lesions, neurological symptoms, learning deficit, growth retardation, and diarrhea (especially in children).
The EFA cannot be produced by the human body; they have to be acquired through diet. A viable and efficient alternative is the consumption of fish with high concentrations of these fatty acids, what can be obtained through fortification of fish.
2. Fortification
This topic summarizes important issues about n−3 LC-PUFA as essential nutrients, in the context of aquaculture and its role in the provision of these dietary fatty acids for human consumption. In doing so, this topic aims to clarify key issues for use of marine ingredients and their replacements, highlighting the influences and limitations of fish metabolism, and critically assess options for the future, as well as sustainable supply of n−3 LC-PUFA.
The term functional food was first introduced in Japan in the mid-1980s and refers to processed foods containing ingredients that help specific body functions besides of being nutritious. These foods are defined as “foods for specific health use,” having specific effects on health due to its chemical constitution and which should not have any health risks. The usual consumption of these foods, without the need for medical supervision, promotes health benefits, confirmed by scientific studies.
A wide range of products has been characterized as functional foods, including components that can affect numerous bodily functions, which are relevant for wellness and health due to their actions in reducing risk of diseases. This class of compounds belongs to nutrition and not to pharmacology, deserving a category of its own, that does not include dietary supplements, but its role in relation to diseases will be in most cases focused more on reducing health risks than on prevention.
Functional foods have the following characteristics:
1. must be conventional foods and be consumed in the normal/usual diet;
2. must be composed of natural components in high concentrations;
3. must have positive effects besides basic nutritional value, which can increase the welfare and health and/or reduce the risk of disease occurrence, promoting health benefits in addition to increase life quality, including physical, psychological, and behavioral performances;
4. the claim of functional property must have scientific basis;
5. can be a natural food or a food in which a component has been removed;
6. can be a food in which the natures of one or more components have been modified;
7. can be a food in which the bioactivities of one or more components have been modified.
In recent years, much attention has been given to the development of meat products and meat production with functions that promote health and prevention of diseases. In this context, research has shown that it is possible to improve the functional value of meat by adding functional compounds, like some fatty acids in animal diets to improve animal production carcass composition and meat quality.
Fish accounted for 16.7% of global population consumption of animal protein and 6.5% of total protein consumed in 2010. However, in addition to protein, fish are also important dietary sources of minerals, including iodine and selenium, and vitamins, such as A, D, and E. Arguably, there is greater importance to consumers in the developed world of fish and seafood as unique and rich sources of omega-3 (n−3). LC-PUFA, particularly eicosapentaenoic (EPA, 20:5n−3) and docosahexaenoic (DHA, 22:6n−3) feature well-known beneficial effects against a variety of human diseases, including cardiovascular and inflammatory diseases, and an important role in neural development.
However, marine fisheries present themselves in a phase of stagnation, with 63% of assessed fish population units requiring resupply. So, consequently, a growing proportion of fish is being grown in captivity, being responsible for almost half of all fish for human consumption.
Food for carnivorous fish species in captivity has been traditionally based on fishmeal (FM) and fish oil (FO) and, at the same time, this practice was good for the supply of n−3 LC-PUFA. With that, the dependency on resources derived from capture fishing becomes an unsustainable practice. Therefore, the continuous growth of aquaculture was dependent on the development of more sustainable foods with alternative ingredients, usually derived from terrestrial agriculture, such as linseed (Linum usitatissimum L.), chia (Salvia hispanica L.), perilla (Perilla frutescens), sacha inchi (Plukenetia volubilis), and Japanese grape seed (Hovenia dulcis), with important consequences for the supply of n−3 LC-PUFA.
There are restrictions on the use of FO and FM in aquaculture. The main restriction is that these are essentially marine, being a finite resource with the production being necessarily limited through strict regulation of fishing and catching. The production of FM and FO features slight decline in recent years, largely due to increased regulations and reduced quotas regarding the amount of fish that can be caught in South America.
The status of FO used in 2012 showed that approximately 75% of total global production was used in aquaculture, with 22% intended for direct human consumption. More than 80% of FO was used in aquaculture feeds, being 62% consumed by salmonids, and 19% by marine fish. Despite continued growth of aquaculture, the use of FO was relatively stable over the last decade with, on average, about 0.8 million tons being used.
An increased use of seafood and aquaculture byproducts, including bycatches, to produce FM and, to a lesser extent, FO, compensated for the reduction in production from whole fish, as a result of increased fishing restrictions. The production is also subject to environmental influences and, while the potential impact of climate change is not well understood, extreme phenomena, such as El Niño, which in particular has well-known consequences and in the next significant event will have the greatest effect on production of FM and supply of FO. This is due to water heating, causing fish shoals to migrate to deeper regions, reducing the amount that are accessible to be fished.
Although the finite and limited nature of production and supply of FO is the main constraint to its use in aquaculture, there are other major factors in the sustainability issue. These are, of course, the main limiting factors of global supply, and they will have an increase in the impact with the many initiatives that have been developed with regard to national and international standards and certifications and marine ingredients.
2.1. Functions of n−3 LC-PUFA in Vertebrates
2.1.1. Metabolic pathways for LC-PUFA biosynthesis
LC-PUFA are defined as PUFA with ≥20 carbon atoms and ≥3 double bonds which, although not a perfect definition, includes all major biologically active LC-PUFA, such as EPA, DHA, and AA.
As PUFA (all fatty acids with 2 or more double bonds) cannot be synthesized by de novo biosynthesis in vertebrates, due to lack of ∆12 enzymes (or n−6) and ∆15 (or n−3) desaturase enzymes responsible for producing PUFA from oleic acid (18:1n−9), PUFA must be provided through diet. However, C18 PUFA, such as alpha-linolenic acid (ALA, 18:3n−3) and linoleic acid (LA, 18:2n−6) can be converted to biologically active LC-PUFA in vertebrates, although it varies with the species, being dependent on the presence and expression of genes of fatty acid desaturation and elongation. As a result, which PUFA can meet the need of EFA varies similarly with the species.
2.1.2. Metabolic functions of LC-PUFA
Fatty acids of LC-PUFA type have major structural roles as constituents of phospholipids, which are components of the cell membrane (biomembranes), and provide many functional properties, affecting both physical and chemical properties of the membrane (e.g., fluidity), as well as influencing membrane proteins (e.g., receptors, enzymes, and transporters) and their functions.
Similarly, as components of triacylglycerols (TAG) and other storage lipids, such as wax esters, LC-PUFA may function as an energy reserve, to be used through β-oxidation of fatty acids in the mitochondria. In addition, however, underlying the essentiality of PUFA, LC-PUFA, or their derivatives have more specific functional roles as important regulators of metabolism.
The regulation can be extracellular at tissue level, through derivatives of LC-PUFA, including eicosanoids, such as prostaglandins, leukotrienes, lipoxynes, resolvins, and protectines, or intracellular as ligands for transcription factors that control gene expression. These regulatory functions support the importance of n−6 or n−3 LC-PUFA, on the concept of ingestion balance of these two PUFA series that cannot be interconverted in vertebrates, and the impact that LC-PUFA from diet can have on health and disease prevention.
2.1.3. n−3 LC-PUFA in human health
Beneficial effects on human health have been related to n−3 LC-PUFA diet, generally based on two main lines of evidence, epidemiological and randomized studies. Laboratory studies that investigate biochemical and molecular mechanisms can also support these approaches. On this basis, many recommendations for intake of EPA and DHA for humans have been produced by a large number of global and national health agencies. According to the Food and Agriculture Organization of the United Nations (FAO, 2010) the recommended daily intake of DHA for 2–4 years old children is 100–150 mg, for 6–10 years old children is 200–250 mg, and for pregnant and lactating women the recommendation is 300 mg. The International Society for the Study of Fatty Acids and Lipids (ISSFAL, 2004) suggests a DHA intake of 500 mg/day for cardiovascular health of the adult population in general.
2.1.4. Neural development
There is strong evidence that the decrease of DHA concentration in the human body can lead to cognitive and visual impairments, and that DHA supplements have positive beneficial results in lactating individuals and premature babies.
In atermo children, DHA supplementation seems to be less effective, suggesting that the minimum recommendation of DHA that is effective to normalize the concentration of DHA should be rectified. There are several reports of potential beneficial effects of DHA supplementation in the diet in a series of psychological, behavioral and psychiatric disorders, including attention-deficit hyperactivity disorder (ADHD) and depression. Although there are some reports of benefits, there are insufficient studies and data to establish definitive conclusions.
However, it is generally acknowledged that n−3 LC-PUFA are potential nutrients that are essential to avoid various pathological conditions associated with normal aging process. This justifies the research on effects of n−3 LC-PUFA on dementia, Alzheimer’s disease (AD), and other age-related cognitive disabilities. In general, there is a lack of agreement on results of intervention studies to support a benefit of n−3 LC-PUFA in cognitive function, but it can also reflect intrinsic limitations in the experimental design of published studies. However, DHA supplementation in patients with some previously diagnosed cognitive disabilities indicated that this seems to slow the progression of AD. In tests with animals, the long-term administration of n−3 LC-PUFA in AD model improved cognitive function and decreased the amount of neuron loss. As chronic inflammation is observed in AD, the n−3 LC-PUFA mechanism of action is postulated as being due to negative regulation of inflammation and the promotion of inflammatory response resolution.
2.2. Fish, aquaculture, and n−3 LC-PUFA
Previously, the role of n−3 LC-PUFA in health and in prevention and/or attenuation of diseases was highlighted. The public now has a good appreciation of the problems with human diets because of high total fat, saturated fat, and cholesterol levels.
Evidences suggest that humans evolved from a diet with a ratio of n−6 and n−3 fatty acids (n−6/n−3) of about 1:1, while modern “Western style” diets called have a proportion of up to 20–25:1, indicating that modern diets are deficient in n−3 PUFA when compared with the diet in which human beings have evolved and their genetic patterns were established. Whereas the main functions of LC-PUFA are control and regulation of lipid metabolism and, especially, of inflammatory/immune responses, the imbalanced intakes of n−6 and n−3 PUFA are critical. Therefore, there is a need to rebalance our dietary intake. The n−6:n−3 ratio, regarding the provision of total fat, can be calculated from data of individual oil/fat production and fatty acid composition of the oil to be around 24:1 posthydrogenation.
Therefore, it is difficult to greatly reduce the intake of n−6 PUFA, although this is a key element of Mediterranean diets rich in olive oil. Increase the intake of n−3 PUFA is entirely feasible, but ALA has relatively little functional role besides acting as a precursor of EPA and DHA, and LC-PUFA biosynthesis is not very active in human beings. Therefore, increasing the intake of n−3 LC-PUFA is the most efficient strategy to correct the imbalance of dietary PUFA, as biological effects largely depend upon it.
Fish and seafood traditionally provide n−3 LC-PUFA to our diet, not because fish is able to produce them, but because the aquatic food chain from sea is rich in n−3 LC-PUFA. As 96% of world water comes from oceans, EPA and DHA are predominantly found in them, and fish simply accumulate these fatty acids.
Food formulations with FM and FO are also intended for aquaculture fish diets, and so fish bred in captivity are also rich in n−3 LC-PUFA. In contrast, as described earlier, FM and FO are finite resources, and the limited supply of FO in particular has affected the growth of farm-raising aquaculture and it was not increasingly substituted by other alternative ingredients, such as the ones derived from plant seeds and vegetable oils.
The need to reduce FO-use demands considerable research for the development of sustainable foods and nutrition strategies to maintain the concentration of n−3 LC-PUFA in farm-raised fish, despite the lower level of n−3 LC-PUFA obtained by diet. As a result, although modern fish diets contain much lower levels of both FM and FO, recent studies showed that farm-raised fish, especially salmon, remain one of the best sources of EPA and DHA for human consumers.
Although relative proportions of EPA and DHA are lower in salmon produced in captivity than in wild salmon (in absolute terms), farm-raised salmon presents (on average) double the amount of EPA and DHA concentrations in comparison to wild salmon. Therefore, the recommended daily EPA and DHA intake for reducing risks of cardiovascular disease (500 mg/day or 3.5 g/week), can be achieved by eating two 150 g servings of farm-raised salmon, while 4–5 servings of wild salmon would be needed.
2.2.1. Nutrition of fish and requirements of n−3 LC-PUFA
As mentioned earlier, all vertebrates, such as fish require a dietary source of PUFA. Qualitative and quantitative requirements of these fatty acids amounts for fish are reported later. Briefly, EFA requirements can be satisfied in general by C18 PUFA in freshwater and salmonid species, while saltwater species require LC-PUFA, EPA, DHA, and AA. In species where requirements can be met by C18 PUFA, LC-PUFA can satisfy at a lower level. These requirements tend to be higher in early stages of life (larvae), when relative proportions of LC-PUFA (EPA, DHA, and AA), are particularly important. EFA quantitative requirements can be described in three levels.
The first level relates to physiological requirement of EFA to avoid classical nutritional pathology (signs of EFA deficiency) and this is low, often around 1% of the diet, or eventually lower if provided by LC-PUFA. The second level relates to the maximum growth and healthy development of fish, and the third level refers to the level of final demand, which is necessary to maintain the nutritional quality based on the content of n−3 LC-PUFA from fish meat. As such, this is not a requirement of fish, but rather of human consumers. To meet this level, EPA and DHA should be supplied in excess, beyond what is already added to maintain the requirements for health and growth of fish, so that this last level is deposited and stored in fish muscle tissue.
Data for minimum requirements of EFA published for fish species represents only good estimates of this level. However, many early studies of minimum EFA requirements in the diet were carried in small fish fed diets containing relatively low lipid amounts for relatively short periods, and growth expectations were lower. In modern diets for fish, higher growth rates were observed, suggesting that EFA requirements should be reassessed.
Therefore, there may be a higher level of demand for EFA than is already known, because although this is currently defined for any species, probably it should vary according to other factors from diet and metabolism of fish.
2.2.2. Influence of endogenous metabolism in fish n−3 LC-PUFA levels
The fatty acid composition of fish is dependent on the interaction of diet (or compositions of fatty acids contained in the food chain) with endogenous metabolism.
The discussion earlier was focused on diet, and this section describes the potential influences that the metabolism can have on n−3 LC-PUFA composition in farm-raised fish.
2.2.2.1. Digestion and absorption
The efficiency of digestion and absorption of nutrients is quantified by measuring apparent digestibility coefficients (ADC), which are estimated by comparing nutrients of food and excrements with a not digestible marker. The ADC of all PUFA, including LC-PUFA, are generally high for most fish species, but they can vary depending on: lipid content from diet, composition of fatty acids, chemical form (TAG, phospholipids), and temperature. However, in general, this means that PUFA obtained from diet are efficiently digested/absorbed and, therefore, these processes do not have much influence on fatty acid compositions of diet.
2.2.2.2. Supply of n−3 LC-PUFA for aquaculture
The discussion before concluded that the composition of fatty acids in fish is fundamentally limited by biochemical and physiological mechanisms of control, in such a way that a solution to maintain high concentrations of n−3 LC-PUFA in tissues when dietary levels are low is unlikely to be found inside fish. This means that the complete cycle of food supply is the only viable option for the maintenance of high levels of n−3 LC-PUFA in farm-raised fish.
2.3. Implications of FO as a Source of n−3 LC-PUFA to Aquaculture
The approach of using FO and FM as n−3 LC-PUFA sources was perfectly sensible 30–40 years ago, during early development of intensive aquaculture. Both marine ingredients were readily accepted and digested by fish, had favorable nutrient compositions, and simply represented the natural food for fish. In addition, FM and FO are the only raw materials that are similar to other primary agricultural products that have been produced and marketed for a long time on a global scale, with aquaculture growing and becoming the biggest customer for these marine resources.
However, the current danger of aquaculture stagnation, with excessive dependence on a limited range of ingredients for application as fish food, and the issues and difficulties regarding their substitution, especially for FO, led to prolonged FO use over many years.
Analyses of fish before and after receiving a “diet enriched with n−3 fatty acids” (FIFO) in aquaculture systems were developed as a potentially useful tool to evaluate the use of marine ingredients. These analyses show that FIFO for aquaculture of all fed species decreased from 1.0 in 1995 to about 0.7 in 2006, with an even more intense decline (0.2) expected for 2020. This relationship was created by groups concerned with the environment, and adopted by groups of farmers who used FIFO incorrectly, mainly for creation of farm-raised salmon, which generated a waste of precious marine resources.
The methodology for calculating FIFO and the high initial values of 4–5:1 for farm-raised salmon has been tested, and it has been suggested that FIFO can be replaced by marine nutrients diets with the dependency to which the amount of each marine nutrient used in feeding is divided by the amount of nutrients used in the product of farm-raising, producing separate indexes for protein and oil. However, FIFO proportion does not present any nutritional basis and cannot be used as a basis for production efficiency.
It is well established that, in aquaculture, there are many efficient and best performing species, such as Atlantic salmon, which has a lower ratio of feed conversion (FC) than any land animal produced for commerce with a value of 1.1 to 1.2 in relation to the value of 2 for poultry, 3 for pigs, and 6 for lamb and other meats. In addition, salmon protein promotes higher energy retention than does beef production from land animals.
Finally, the emphasis on FIFO values marginalized another important fact, which is the general improvement in the formulation of diets for fish that have led to continuous and fundamental improvements to increase the production efficiency of farm-raising that, in turn, has led to a greater efficiency regarding FO use. FIFO should be used only as a development standard for different diets in aquaculture, since with expansion of fish production and finite supplies of FO and FM, the actual trend is to reduce FIFO values. In fact, it has been shown recently that Atlantic salmon can be a liquid product of both marine proteins and n−3 LC-PUFA, although, in the latter, it must be pointed out that n−3 LC-PUFA levels in products are much lower than those in fish fed with FO.
The real problem with FM and, especially FO, as happens with many commodities, is the supply and demand. The large use of these finite resources became unsustainable, suggesting that FO and FM shall not be used in excess. The sustainability of industrial fishing is no different from any other fishery in the world. In order to be sustainable, all fish must be managed and regulated, as many species are already in several areas, including South America and Europe.
However, the supplies for production of FO and FM are always finite and limited. As FO and FM are the primary sources, this implies that the current global supply of n−3 LC-PUFA is similarly finite and limited, and the difference between supply and demand can be estimated.
Based on the recommended dose for cardiac health, the total demand of n−3 LC-PUFA is more than 1.25 million metric tons (mt), while the total supply is optimistic, estimated at just over 0.8 million mt, indicating a deficit of more than 0.4 million mt. The bulk of the supply (almost 90%) comes from fish food or through FO and FM, with relatively small additional amounts (realistically estimated) from seafood and byproducts of recycling. There is an essential problem of a worldwide lack of n−3 LC-PUFA to meet all human needs, either by direct adjustment in consumption or through aquaculture.
2.4. Application: Fortification of Fish Diet With n−3 PUFA Aimed at Incorporation of n−3 LC-PUFA in the Flesh of Fish
As mentioned earlier, fish can synthesize some LC-PUFA through their metabolism if their diet is supplied with the fatty acids LA and ALA. Therefore, we will illustrate how this happens through experimental data obtained in our analytical research group applied to lipids, sterols, and antioxidants (APLE-A).
The first step is to formulate a diet that is rich in ALA. To verify the difference, a control diet (rich in LA), which is a soybean (Glycine max) oil-based diet, must also be supplied to farm-raised fish.
Table 11.1 lists the food formulations and the results for physicochemical analyses of these diets.
Table 11.1
Food formulations and physicochemical analyses of developed diets.
| Ingredients (%) | Treatments | |
| Control | TII | |
| Corn meal | 37.39 | 37.39 |
| Soybean meal | 32.08 | 32.08 |
| Poultry viscera meal | 14.00 | 14.00 |
| Wheat bran | 8.00 | 8.00 |
| Rice grits | 5.00 | 5.00 |
| Soybean oil | 1.90 | — |
| Chia oil | — | 1.90 |
| Dicalcium phosphate | 0.22 | 0.22 |
| Common salt (NaCl) | 0.50 | 0.50 |
| Premix | 0.50 | 0.50 |
| HCl l-lysine | 0.06 | 0.06 |
| dl-Methionine | 0.06 | 0.06 |
| l-Threonine | 0.07 | 0.07 |
| Propionic acid | 0.10 | 0.10 |
| l-Tryptophan | 0.02 | 0.02 |
| BHT | 0.02 | 0.02 |
| Proximate composition* | ||
| Moisture (%) | 9.63a ± 0.04 | 8.61a ± 0.12 |
| Ash (%) | 5.40a ± 0.03 | 5.59a ± 0.04 |
| Crude protein (%) | 33.33a ± 0.48 | 32.32ª ± 0.58 |
| Total lipids (%) | 6.38a ± 0.06 | 6.08ª ± 0.16 |
| Carbohydrates (%) | 45.26a ± 0.49 | 47.40a ± 0.61 |
| Energetic value (kcal 100 g–1) | 371.78a ± 0.69 | 373.60a ± 0.86 |
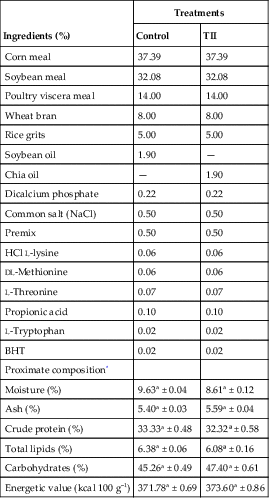
* Mean of values ± standard deviation; n = 3. Different letters in the same line indicate significant difference (P < 0.05) by t-test. Premix = vitamin complex; BHT = butylated hydroxytoluene.
As can be verified by Table 11.1, diets must be isocaloric and isoproteic to not influence the results of fattening and growth of fish.
The control diet is a common formulation, containing soybean oil in its composition, provided to farm-raised fish. Diet II is enriched with chia (Salvia hispanica L.) oil, which has high amounts of ALA, substituting soy oil.
To verify how this can influence the concentration of fatty acids in fish flesh, let’s check Table 11.2, which shows the concentration of fatty acids from diets.
Table 11.2
Fatty acids concentrations, in mg/g of total lipids (mg/g of TL), from diets.
| Fatty Acids | Treatments | |
| Control | TII | |
| 14:0 | 16.90a ± 1.05 | 16.26a ± 0.88 |
| 15:0 | 3.78ª ± 0.18 | 3.68a ± 0.24 |
| 16:0 | 957.43a ± 36.25 | 924.55a ± 30.07 |
| 16:1n−9 | 7.84ª ± 0.46 | 8.16ª ± 0.57 |
| 16:1n−7 | 51.87ª ± 1.73 | 52.67ª ± 1.73 |
| 17:0 | 11.34ª ± 0.83 | 8.83b ± 0.04 |
| 17:1n−11 | 5.31ª ± 0.43 | 5.22ª ± 0.23 |
| 17:1n−9 | 4.44b ± 0.08 | 5.73a ± 0.11 |
| 18:0 | 229.12a ± 7.63 | 232.26a ± 7.52 |
| 18:1n−9 | 1647.46ª ± 59.47 | 1397.57b ± 42.89 |
| 18:1n−7 | 76.14ª ± 4.33 | 66.47b ± 2.16 |
| 18:2n−6 | 2407.63a ± 89.77 | 1937.33b ± 64.55 |
| 18:3n−6 | 4.17b ± 0.32 | 5.13a ± 0.08 |
| 18:3n−3 | 204.80b ± 11.76 | 1141.72a ± 40.81 |
| 20:0 | 22.23a ± 0.97 | 21.47a ± 1.41 |
| 20:1n−9 | 1.64ª ± 0.04 | 1.60ª ± 0.11 |
| 20:1n−7 | 12.67ª ± 0.73 | 12.06ª ± 0.72 |
| 22:0 | 30.33a ± 1.61 | 29.42a ± 0.96 |
| 24:0 | 12.27ª ± 0.41 | 10.38b ± 0.15 |
| SFA | 1283.40ª ± 37.12 | 1246.85ª ± 31.06 |
| MUFA | 1807.36ª ± 59.66 | 1549.48b ± 42.99 |
| PUFA | 2616.60b ± 102.44 | 3084.18a ± 76.37 |
| n−3 | 204.80b ± 11.76 | 1141.72a ± 40.81 |
| n−6 | 2411.80ª ± 101.77 | 1942.46b ± 64.55 |
| n−6/n−3 | 12.27ª ± 0.07 | 1.70b ± 0.05 |

Mean of values ± standard deviation; n = 3. Different letters in the same line indicate significant difference (P < 0.05) by t-test. SFA = sum of saturated fatty acids; MUFA = sum of monounsaturated fatty acids; PUFA = sum of polyunsaturated fatty acids; n−3 = sum of omega-3 fatty acids; n−6 = sum of omega-6 fatty acids.
As can be checked through the data shown in Table 11.2, all the fatty acids of the two diets do not present significant differences, with the exception of LA and ALA, since the control diet is rich in LA while treatment II contains chia oil (which has a high ALA content).
This reflects directly on the n−6/n−3 ratio, as can be verified in the same table. The control diet has a ratio of 12.27 and the treatment II n−6/n−3 ratio is 1.70, while the maximum n−6/n−3 ratio value recommended by the National Ministry of Health is 4.
After 15, 30, and 45 days of feeding with these diets, fish were slaughtered and the results of concentration of fatty acids found in fish fillets are shown in Table 11.3.
Table 11.3
Fatty acid composition (mg/g of TL) and total lipid content (g 100 g−1 of sample) of tilapia filets from the treatments.
| Control | 15 days | 30 days | 45 days | |
| Total Lipids (g 100 g−1 of sample) | 2.82 ± 0.23 | 3.79 ± 0.15 | 3.61 ± 0.34 | 4.18 ± 0.01 |
| Fatty acids (mg/g of TL) | Control | 15 days | 30 days | 45 days |
| 14:0 | 17.17a ± 0.62 | 16.84a ± 0.54 | 14.78b ± 0.69 | 15.23b ± 0.22 |
| 16:0 | 196.22a ± 7.46 | 199.82a ± 10.67 | 187.03a ± 1.70 | 158.15b ± 2.38 |
| 16:1n−9 | 5.07a ± 0.18 | 5.01a ± 0.29 | 4.92a ± 0.39 | 4.52ª ± 0.07 |
| 16:1n−7 | 31.08ª ± 1.00 | 32.86a ± 1.92 | 34.00a ± 2.69 | 31.02a ± 0.27 |
| 17:0 | 3.09a ± 0.12 | 3.20a ± 0.24 | 2.57b ± 0.14 | 2.41b ± 0.13 |
| 18:0 | 69.97a ± 3.55 | 69.80a ± 2.66 | 59.85b ± 4.94 | 51.05c ± 1.38 |
| 18:1n−9 | 265.86a ± 10.58 | 286.56a ± 15.14 | 280.63a ± 3.61 | 285.16a ± 2.99 |
| 18:1n−7 | 24.05a ± 0.91 | 25.00a ± 0.78 | 25.68a ± 1.73 | 24.49a ± 0.03 |
| 18:1n−5 | 2.18ab ± 0.09 | 1.96bc ± 0.16 | 2.34a ± 0.07 | 1.83c ± 0.01 |
| 18:2n−6 | 157.78c ± 3.21 | 166.69bc ± 8.67 | 171.27b ± 3.95 | 186.56a ± 1.74 |
| 18:3n−6 | 7.13ab ± 0.10 | 6.67b ± 0.33 | 7.48a ± 0.49 | 6.52b ± 0.07 |
| 18:3n−3 | 9.65d ± 0.13 | 28.30c ± 1.40 | 47.18b ± 2.07 | 59.21a ± 1.39 |
| 20:0 | 2.47a ± 0.06 | 2.14a ± 0.09 | 1.50b ± 0.43 | 1.94ab ± 0.15 |
| 20:1n−9 | 12.55a ± 0.63 | 12.20a ± 0.15 | 12.85a ± 0.76 | 13.64ª ± 0.70 |
| 20:2n−6 | 6.17c ± 0.28 | 8.59ab ± 0.27 | 7.96b ± 0.53 | 9.14a ± 0.34 |
| 20:3n−6 | 9.32a ± 0.36 | 9.55a ± 0.27 | 8.65ab ± 0.77 | 7.88b ± 0.75 |
| 20:4n−6 (AA) | 13.46a ± 0.34 | 13.74a ± 0.50 | 13.17a ± 1.74 | 10.45b ± 0.25 |
| 20:4n−3 | 2.32d ± 0.07 | 4.76c ± 0.19 | 7.70b ± 0.67 | 9.86a ± 0.12 |
| 22:0 | 0.28c ± 0.02 | 0.75b ± 0.03 | 1.40a ± 0.09 | 1.39a ± 0.06 |
| 22:1n−9 | 0.44c ± 0.05 | 0.74b ± 0.02 | 1.14a ± 0.08 | 1.24a ± 0.09 |
| 20:5n−3 (EPA) | 0.57c ± 0.03 | 0.67bc ± 0.03 | 0.91a ± 0.05 | 0.76b ± 0.08 |
| 22:4n−3 | 7.41ab ± 0.49 | 6.95bc ± 0.19 | 7.80a ± 0.15 | 6.53c ± 0.18 |
| 24:0 | 0.21d ± 0.01 | 0.50c ± 0.03 | 1.17b ± 0.04 | 1.92a ± 0.09 |
| 24:1n−9 | 3.65d ± 0.16 | 4.23c ± 0.12 | 5.70a ± 0.24 | 5.12b ± 0.15 |
| 22:6n−3 (DHA) | 11.96c ± 0.37 | 13.06b ± 0.26 | 15.95a ± 0.23 | 13.07b ± 0.28 |
| SFA | 289.41a ± 8.28 | 293.05a ± 11.01 | 268.30a ± 11.01 | 232.08b ± 11.01 |
| MUFA | 344.88a ± 10.69 | 368.55a ± 15.28 | 367.26a ± 15.28 | 367.01ª ± 15.28 |
| PUFA | 223.44d ± 3.32 | 254.22c ± 8.81 | 280.36b ± 8.82 | 300.12a ± 8.82 |
| n−3 | 29.01d ± 0.62 | 48.31c ± 1.43 | 70.92b ± 1.44 | 78.81a ± 1.44 |
| n−6 | 173.27c ± 3.26 | 184.83bc ± 8.69 | 187.88b ± 8.70 | 203.58a ± 8.70 |
| n−6/n−3 | 5.97a ± 0.03 | 3.83b ± 0.05 | 2.65c ± 0.05 | 2.58c ± 0.05 |

Means of values ± standard deviation; n = 3. Different letters in the same line indicate significant difference (P < 0.05) by Tukey’s test. SFA = sum of saturated fatty acids; MUFA = sum of monounsaturated fatty acids; PUFA = sum of polyunsaturated fatty acids; n−3 = sum of omega-3 fatty acids; n−6 = sum of omega-6 fatty acids. AA = arachidonic acid; EPA = eicosapentaenoic acid; DHA = docosahexaenoic acid; TL= total lipids.
As it can be seen in Table 11.3, as long as the diet enriched with chia oil is supplied, the fish gradually incorporates the n−3 PUFA in their flesh and metabolize the excess of these fatty acids in LC-PUFA, especially EPA, DHA, and AA. This reflects directly on the composition of fish fillet, where it can be observed that the fish that received a diet enriched with chia oil during 45 days presented the best n−6/n−3 ratio (2.58). With this, it is clear that it is not enough to simply ingest fish meat, because its nutritional quality is directly related to the food that was provided to the fish. In this way, only those who receive a diet enriched with n−3 fatty acids will show fillets with better lipid quality and nutritional value.
2.4.1. Identification and quantification of fatty acids by gas chromatography with flame ionization detector (GC–FID)
In the determination of fatty acid composition, total lipids of the samples are normally extracted from diets and fillets according to the methodology described by Bligh and Dyer (1959), as it uses a mixture of cold solvents for extraction, not compromising the lipid structure. In addition, this method extracts polar, neutral, and apolar lipids.
Gas chromatography has occupied a prominent position among the methods of analysis due to the ease and efficiency for performing the separation of different lipid components from a mixture, being coupled to detectors that aid in identification and absolute quantification of constituents. However, it is a method that requires stable substances at the temperature that is employed during the chromatographic analysis, and the analyte must be volatile or it must have a volatile derivative.
Thus, prior to chromatographic analysis, a methylation step of the total lipids is required (conversion of free fatty acids and esters linked to triacylglycerol in methyl esters). This step is necessary to enable the volatilization of the compounds of interest and allow the determination by gas chromatography.
The fatty acid methyl esters (FAME) are separated into a gas chromatograph equipped with flame ionization detector and a CP-7420 (Select FAME, 100 m length, 0.25 mm internal diameter, and 0.25 μm of cyanopropyl thickness) fused silica capillary column. The flow rates of gases are 1.2 mL/min for carrier gas (H2); 30 mL/min for the auxiliary gas (N2); and 35 and 350 mL/min for H2 and synthetic air, respectively, for the detector flame. The injected volume is 2.0 μL, using sample split of 1:80. The injector and detector temperatures are 200 and 240°C, respectively. The column temperature is set to 165°C during 7.00 min, followed by a heating ramp of 4°C/min until it reaches 185°C. The column is kept at this temperature during 4.67 min, followed by another heating ramp of 6°C/min until the column reaches 235°C, kept by 5.00 min, therefore totaling 30.00 min. The retention times and peak areas of FAME are determined through a computer software.
The fatty acids are identified through the comparison of retention times with standards of known composition and the absolute quantification of FAME is accomplished through the internal standardization, using the methyl ester of tricosanoic acid (23:0) as standard. Theoretical correction factors (Visentainer et al., 2014) are employed for the determination of concentrations values. The amount of fatty acids in the samples is calculated in mg/g of total lipids (mg/g of LT) using Eq. (11.1):
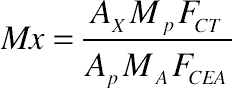 (11.1)
(11.1)Where: Mx is the concentration of fatty acid “X” in mg/g of total lipids, Ax is the fatty acid “X” peak area, Ap is the internal standard (23:0) peak area, Mp is the internal standard mass added to the sample in mg, MA is the sample mass in grams, FCT is the theoretical correction factor of the flame ionization detector (FID) and FCEA is the conversion factor from fatty acid to methyl ester.
The flame ionization detector (FID) is widely used in the analysis of fatty acid methyl esters due to its low cost, high efficiency, quick and almost universal response, great linearity range and high sensitivity, approximately 10–12 g. However, the identification of compounds using FID cannot be conclusive. To increase the efficiency of this identification, the following procedures are used: addition of “spiking” standard, use of secondary standards, graphics methods and systematic retention indexes.
Normally the projects carried out in the FAME food analysis use the method of area normalization, where the results are expressed in percentage of relative area. In this method, all the components of the sample must be detected and, in the case of omission or estimation of a component, the other components are affected. Besides, the results presented by this method present difficulties of interpretation.
Recent works express the results in absolute amounts, that is, mass concentrations of FAME by mass of raw material or sample. It is one of the best methods for quantification of fatty acids, as it provides reliable results that are easily interpreted, providing results with greater accuracy while allowing comparisons with works that use modern techniques of quantification. Given that, the use of internal standards has been increasing, as it allows researchers to quantify the analytes of interest, as well as minimize experimental errors.
The use of internal standard methodology calls for some requirements, as the substance cannot be part of the sample, it must present a high degree of purity, be accessible and cheap, must not coelute with components of the sample, yet at the same time, it must elute close to them. As the fatty acid methyl esters respond differentially in FID, it is necessary to use correction factors for the methyl esters in relation to the internal standard, which is also an ester. The used factors are the experimental or empirical correction factor (FCEA), whose value is determined experimentally, and the theoretical correction factor (FCT), theoretically determined from the number of active carbons (C*). Both factors are based on comparisons between studied methyl esters and the internal standard.
3. Biosynthesis of Fatty Acids in Fish
3.1. Omega-3 and Omega-6 Fatty Acids
The freshwater and saltwater fish have different needs of n−3 and n−6 fatty acids. While freshwater fish need to acquire minimum levels of essential dietary PUFA (ALA and LA), especially in the early stages of development, to prevent nutritional deficiency, the fish of marine origin have a greater demand for LC-PUFA, especially EPA, DHA, and AA. This distinction occurs mainly because freshwater species have, in general, all enzymes required in the process of elongation and desaturation that compose the biosynthesis of LC-PUFA from essential precursors. Elongase enzymes act by adding two carbon atoms in the initial part of the fatty acid carbon chain, and the desaturase enzymes oxidize two carbons in the chain, resulting in a double bond with the cis configuration.
The saltwater species, on the other hand, have very large variations regarding enzyme activity, as certain species have low or zero ability for bioconversion of fatty acids. Low activity of elongase enzyme (C18 to C20) was found in Turbot (Scophthalmus maximus), as well as low ∆5 desaturase enzyme activity was shown by carnivore saltwater fish, such as gilt-head sea bream (Sparus aurata). Thus, a proper diet for saltwater fish must be constituted of LC-PUFA (EPA, DHA, and AA). This problem is overcome, because the diet of saltwater fish, consisting of phytoplankton and macroalgae, presents significant amounts of LC-PUFA, especially from n−3 series, which can be absorbed and accumulated in fish.
However, the diet of freshwater fish, especially of those bred in captivity, is recognized for being rich in n−6 fatty acids, due to the use of vegetable oil sources, mainly soybean (G. max), which is composed of high levels of these fatty acids. Although the enzymatic preference is for substrates with higher levels of insaturation (n−3), it should be emphasized that there is competition between the two fatty acid series by the same enzymes for biosynthesis of LC-PUFA, and this competition is predominant when there is an excess of one of the precursor substrates. Thus, the diet should be alternately enriched with n−3 fatty acids to meet the minimum requirements of the fish and, to maintain the nutritional quality of the fillet for human consumption, the supply must be high so the fish can carry out the biosynthesis of other fatty acids and deposit them in the muscle lipid.
3.2. Metabolic Pathways for Biosynthesis of Fatty Acids in Fish
All known organisms are able to synthesize SFA through the de novo biosynthesis, or to get them through diet. The de novo biosynthesis involves the formation of endogenous fatty acids from nonlipid precursors through the action of the multienzymatic complex fatty acid synthetase (FAS), via lipogenesis. The main products of FAS are the 16:0 (palmitic acid) and 18:0 (stearic acid) fatty acids (Fig. 11.5). However, the de novo biosynthesis of saturated fatty acids is inversely proportional to the levels of SFA obtained through diet, indicating that higher amounts of SFA ingested by the body lead to lower amounts of synthesized SFA, and vice versa.
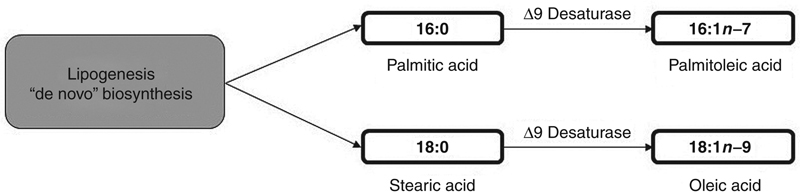
Figure 11.5 Biosynthesis of Saturated and Monounsaturated Fatty Acids in Living Organisms.
Additionally, the MUFA may have their origins through ingestion of lipids, or MUFA can be synthesized by all organisms through the action of the ∆9 desaturase enzyme in their equivalent saturated substrates, 16:0 and 18:0. In this way, it is possible to obtain, respectively, the 16:1n−7 (palmitoleic acid) and 18:1n−9 (oleic acid) MUFA (Fig. 11.5).
However, the vertebrate organisms, including fish, are not able to synthesize PUFA by de novo biosynthesis, as they do not have the ∆12 and ∆15 desaturase enzymes that are needed for the biosynthesis of LA and ALA precursors, respectively (Fig. 11.6).
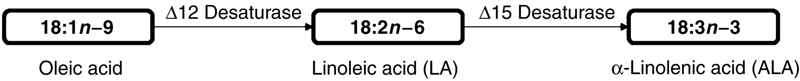
Figure 11.6 Biosynthesis of Essential Fatty Acids Precursors of the n−6 and n−3 Series Observed in the Kingdom Plantae.
Vertebrate organisms do not have the ∆12 and ∆15 desaturase enzymes that are required to synthesize these fatty acids.
Vertebrate organisms do not have the ∆12 and ∆15 desaturase enzymes that are required to synthesize these fatty acids.
Therefore, these precursor fatty acids must be obtained through diet to endogenously enable the biosynthesis of the remaining fatty acids from both series, in particular from n−3 LC-PUFA series, through the action of elongase and desaturase enzymes, as shown in Fig. 11.7.
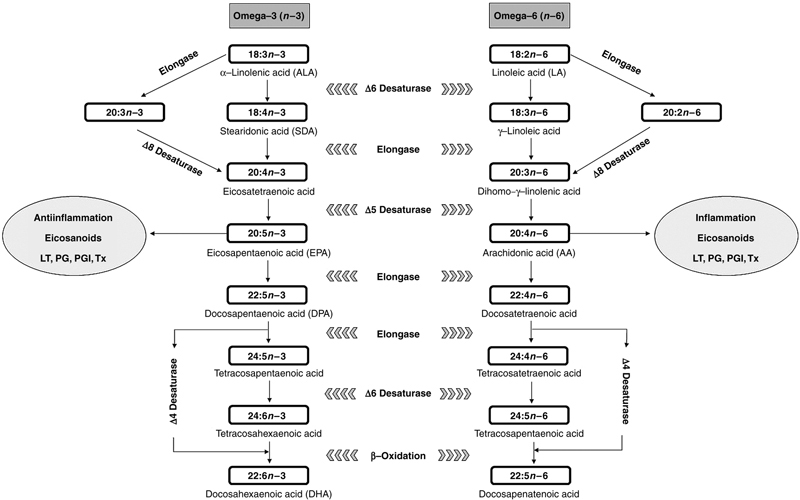
Figure 11.7 Schematic Representation of the Biosynthesis of Long-Chain Polyunsaturated Fatty Acids From Essential Precursors.
LT, Leukotrienes; PG, prostaglandins; PGI, prostacyclins; Tx, thromboxanes.
LT, Leukotrienes; PG, prostaglandins; PGI, prostacyclins; Tx, thromboxanes.
The production of eicosapentaenoic acid (EPA, 20:5n−3) from ALA precursor requires inclusion of two insaturations via activities of ∆6 or ∆8 desaturase enzymes (depending on whether the ALA was elongated before desaturation or not) and ∆5 desaturase. Sequentially, the biosynthesis of docosahexaenoic acid (DHA, 22:6n−3) from EPA may proceed directly through elongation and subsequent inclusion of one insaturation through ∆4 desaturase enzyme (alternative pathway), or via “Sprecher shunt,” which involves the elongation of EPA to tetracosapentaenoic acid (24:5n−3), inclusion of a sequential insaturation through ∆6 desaturase enzyme and a shortening of the chain via β-oxidation.
Analogously, the biosynthesis of arachidonic acid (AA, 20:4n−6) from the LA precursor involves the inclusion of two double bonds and chain elongation, through the action of the same previously discussed enzymes (∆6/∆8 and ∆5 desaturase enzymes, depending on the route, and elongase enzymes).
The LC-PUFA intermediaries of the n−3 and n−6 pathways (EPA and AA, respectively) have very important roles in the control and regulation of the lipid metabolism in fish, because they are precursors of eicosanoids (Fig. 11.7). The understanding of metabolism is necessary, because during the process of elongation and desaturation the same enzymes are shared by n−3 and n−6 series for biosynthesis of LC-PUFA, such as AA and EPA. Thus, there is a competition between n−3 and n−6 fatty acids for these enzymes, and the excess of a particular fatty acid directly affects the metabolism of another. Consequently, the production of eicosanoids is also changed.
Another important characteristic of metabolism that indicates that the balanced intake of ALA and LA precursors should be taken into consideration, is the fact that the n−3 and n−6 fatty acids series can be considered as special series, because the n−3 fatty acids cannot be transformed into n−6 series members, and vice versa, as the inclusion of a double bond by ∆-desaturase enzymes and the inclusion of two carbon atoms by elongase enzymes occurs between carboxyl and the first double bond of the fatty acid carbon chain. Therefore, the position of the double bond relative to the terminal methyl group of the carbonic chain, which determines whether a fatty acid belongs to n−3 or n−6 series, is not changed (Fig. 11.8).
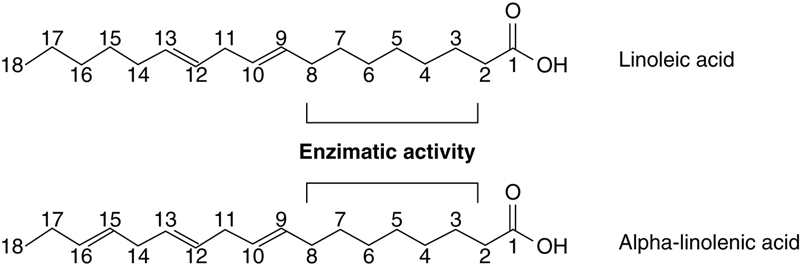
Figure 11.8 Enzimatic Activity Regions of Elongase and Desaturase Enzymes in the Biosynthesis of Fatty Acids, Following Delta Designation.
The original positions of characteristic double bonds of n−6 and n−3 series remain unchanged.
The original positions of characteristic double bonds of n−6 and n−3 series remain unchanged.
The name delta (∆), which is based on the numbering of the fatty acid chain from the carboxyl group, is used to indicate the position for enzymatic activity in chemical reactions involving fatty acids.
3.3. Factors That Affect the Biosynthesis of Fatty Acids in Fish
Although the vertebrate organisms, including fish, are not capable of producing the fatty acids precursors of n−3 and n−6 series, they can metabolically convert them in biologically active LC-PUFA, as seen earlier. However, the biosynthesis is affected by several factors that can lead to inefficient and/or imbalanced production of the fatty acids from both series, and the final composition of fatty acids found in fish is directly dependent on two crucial factors: endogenous metabolism and the diet.
Within the metabolic context, besides the differences of enzyme activity in fish, which determine the course of the fatty acid biosynthesis in n−3 and n−6, another factor must be taken into consideration: the oxidation of fatty acids.
The priority of oxidation obeys the following order: monounsaturated/saturated fatty acids > polyunsaturated fatty acids > long-chain polyunsaturated fatty acids, with the n−6 series being oxidized before n−3 series. However, with regard to the deposition of fatty acids, higher amounts of ingested fatty acids lead to a lower observed deposition, resulting in an increase of oxidation with subsequent indication that the overall oxidation rate of fatty acids depends on the balance between the enzymes involved in the metabolic process and substrate concentration (ingested fatty acids, in question), and the latter appears to be predominant.
This trend is valid for most PUFA, such as EPA. DHA is an exception, being preferentially deposited in fish in larger concentrations to those obtained through diet, rather than being oxidized. This occurs because of the greater difficulty of DHA oxidation through β-oxidation process.
Taking into consideration the great contribution that the fish have on food as a primary source of n−3 LC-PUFA, and the increasing production need of farm-raised fish, the understanding of the biosynthesis mechanisms for these fatty acids becomes of extreme importance for specific nutritional determination of fatty acid precursors for the guarantee of a good growth and development of the fish, and especially to maintain high levels of n−3 LC-PUFA for human consumption.
4. Bioavailability and Conclusions
As seen previously, there is a need for production of farm-raised fish, to meet the necessities for human feeding, both in availability of a fish with excellent quality attributes (such as free from physical, chemical, and biological contaminants; an appearance without stains, holes, or cuts on the surface; the skin must be moist, tight, and adhered to the flesh; the eyes must be brilliant, protruding while occupying the entire eye cavity) and regarding fish centesimal composition quality, with the improvement in the production chain for breeding of fish fortified with omega-3.
Therefore, investments in the production chain are necessary, because according to the Food and Agriculture Organization of the United Nations, in 2030 fish deficit will reach approximately 100 million tons. The recommendation of the World Health Organization is 12 kg of fish per inhabitant/year, equivalent to the consumption of 250 g of fish per week.
For human feeding, the consumption of fish must be encouraged through divulgation of its specific health benefits, as the flesh of fish shows excellent bioavailability.
The bioavailability of an ingested nutrient can be defined as its accessibility for normal physiological and metabolic processes, that is, the efficiency in which a component of the diet is used systematically through normal metabolic pathways. The bioavailability is a response of the interaction between diet, nutrient, and the individual.
Another relevant factor is the protein composition of fish, being made up of proteins with high biological value, related to the human body’s ability to absorb the amino acids contained in foods. Fish presents a composition of essential amino acids (valine, leucine, isoleucine, threonine, methionine, lysine, phenylalanine, and tryptophan) in appropriate proportions. With respect to the amount of fish protein, the content is always high, ranging from 15% to 25%.
The digestibility (portion of the food which is not excreted through feces) is high, over 95%, depending on the species, and higher than common meat and milk types, due to the minimal amount of connective tissue. The biological value is higher than other sources, such as eggs, milk, and beef. With processing, fish protein quality is maintained
The advantages of fish, in relation to beef, are mainly the content and quality of the lipids that compose fish meat. The fish, especially from cold seawater or enriched farm-raised species, are rich in monounsaturated and polyunsaturated fats and contain very low levels of cholesterol (substance belonging to the class of lipids which is present predominantly in animals), unlike other meats that have high concentrations of cholesterol in their composition.
In conclusion, consumption of these fats is of extreme relevance for prevention of cardiovascular and neurodegenerative diseases, and for the treatment of respiratory diseases, because it contains large amounts of the omega-3 fatty acids. In general, the lipid composition can vary according to: (1) the species of fish, (2) analyzed corporal muscle type, (3) sex, (4) type of food provided or eating habits, (5) age, (6) time of the year, (7) habitat, and (8) degree of gonadal maturation.