3
The Scientific Basis for Food Security
3.1 The Supply of Essential Plant Nutrients
The conversion of solar energy into biomass through photosynthesis is dependent on several factors that can limit the amount of biomass or crop yield achieved. The notable limiting factors that growers and farmers are constantly trying to optimise for greatest crop yield are light, temperature, water, and plant nutrients. While light and temperature limitations are principally dealt with by sowing periods and protecting crops from weather, plant nutrients, and water are potential limiting factors that are perhaps most under the control of the management system the grower chooses. For example, limitations to water availability can be achieved by planning storage, capture, and irrigation of water resources, but ultimate availability may well be outside the control of growers. Plant nutrients are supplied as mineral fertilisers or organic manures, and these materials supplement nutrients available from the soil. What is critical to consider at this stage is that most essential nutrients required in foods by human beings are derived from biomass derived from crops and the agronomic decisions made during production will influence dietary quality.
Plant nutrients are found in crops at distinct concentrations that correspond to the ideal range for optimal plant health and the functioning of metabolism within the plant. It is possible to determine optimal physiological and metabolic states by the analysis of specific nutrients in field trials. There are critical values of plant nutrients below which there will be a decrease in yield, and the use of tissue analysis by agronomists is proving useful in determining critical nutrient values below which yield loss may become apparent. Critical values can also be used to manage crops at a particular level of production and determine optimal application of nutrients as fertilisers. The response curve of yield, metabolism, and physiological variables is generally hyperbolic, that is an initial linear phase of increase (of biomass yield) at low concentration of a variable such as nitrate, followed by a curvilinear phase at higher concentrations (Figure 3.1). This type of response can be viewed in terms of limiting factors and control theory, where several factors are involved in limiting the rate of response associated with a system. The mass of nutrient required per hectare of land is surprisingly stable across the plant kingdom and follows the type of responses shown in Figure 3.1.
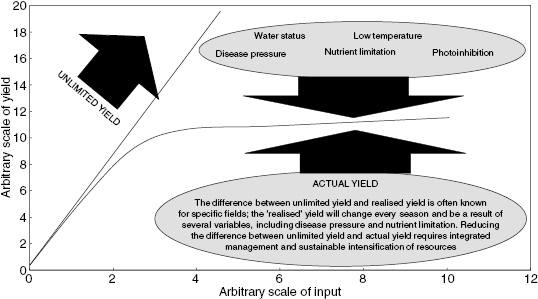
This type of curvilinear relationship has been established since Lawes and Gilbert of Rothamsted developed the fertiliser recommendation for crops in the nineteenth century, it has continually been reviewed and revised for new technologies that have an impact on crop yield in agricultural systems.1 It is currently viewed as a sum of several limiting factors that can be described in terms of control theory, that is, there are several control points in determining yield. Some control points may have more impact than others, and this value of impact will be dependent on environmental attributes, such as water availability, temperature, and light intensity.
Plant nutrients are classified on the basis of the mass required by plants to function metabolically and, in the case of crops, to provide optimal biomass quality and yield. The classification groups of the plant nutrients are described in Table 3.1. They include the major nutrients, the secondary nutrients, and the micro-nutrients or trace elements.
Table 3.1. The essential plant nutrients required for the production of all crop biomass
Source: These data were adapted from the research of W. Martindale, published for Agronomy Extension Services as Part of the Fertiliser Advisers Certification and Training Scheme Technical Information Service, and Bøckman, Kaarstad, and Richards (1990).2
| Nutrient (with common ion available to plants in brackets) | Symbol (fertiliser oxide declaration symbol in brackets) | Description and amount required by crops |
|---|---|---|
| Macronutrients | ||
| Nitrogen (NO3−) | N | Between 40 and 400 kg nutrient required by a crop per hectare for optimal crop yield. Nitrate, very mobile; ammonium, not mobile; phosphate, not mobile; potassium, mobile |
| Phosphorus (PO43−) | P (P2O5) | |
| Potassium (K+) | K (K2O) | |
| Minor nutrients | ||
| Sulfur (SO32−) | S (SO3) | Typically 10–40 kg nutrient required by a crop per hectare for optimal yield. Sulphate, mobile; calcium, mobile; magnesium, mobile |
| Calcium (Ca2+) | Ca (CaO) | |
| Magnesium (Mg2+) | Mg (MgO) | |
| Micronutrients | ||
| Iron (Fe2+) | Fe | Typically needed in quantities of 0.01–5 kg/ha Many micronutrients can be toxic at relatively low concentrations. The amount of micronutrient allowed in soils that have nutrients applied to them may be controlled by legislation. The availability of micronutrients is very much dependant on soil pH, with a general condition being greater availability of micronutrients and heavy metals at a pH below 7 in soils. This is not the condition for molybdenum, whose solubility increases at a pH above 7 in soils. Older soils >10 000 years old, very light soils are likely to have more trace element deficiencies than younger or heavier soils. Soil pH will be a critical factor in availability of trace elements. |
| Boron | B | |
| Manganese | Mn | |
| Copper (Cu2+) | Cu | |
| Zinc (Zn2+) | Zn | |
| Molybdenum | Mo | |
| Chlorine (Cl−) | Cl |
Notes: As such, these nutrients maybe considered the base of the food supply chain. They are the inputs that are ultimately contained in foods that are added to or preserved by producers, processors and manufacturers.
The nutritional balance between the agricultural biomass producer to the food processor and manufacture in the supply chain is not only about the interface or input–output ratio of the 13 essential plant nutrients described in Table 3.1. The essential plant nutrients can optimise plant metabolism for production and influence the nutritional quality of biomass by their impact on the production of an important group of metabolites called phytonutrients that include pigments, flavours, fragrances, metabolic co-factors, and vitamins.
3.2 Plant Nutrients and Phytonutrients in the Food Supply Chain: Establishing a Nutritional Understanding Using Human Trials
Phytonutrients are a group of nutritional chemicals that are grouped collectively because they are of high nutritional value whose concentration in crop biomass is drastically changed by crop variety, agronomic management, and preservation in the supply chain.3 The phytonutrients include familiar dietary nutrients, such as vitamin A, vitamin C, vitamin B, vitamin E, carotenoids, lycopenes, and flavonoids. These are often collectively described as antioxidants in that they reduce the activity of potentially harmful free radicals in cells that result from metabolic reactions. This function of ‘mopping-up’ of oxidising capacity is linked to providing anti-carcinogenic impact, enhancing healthy metabolic attributes and improving several specific organ functions. The existence of data to support the functions of phytonutrients and antioxidants in the diets of populations is often limited by the number of people included in nutritional trials, but evidence continues to increase.4 There are excellent examples of nutritional research on micronutrient and protein consumption that can be integrated into diet and meal planning so the link between phytonutrient consumption and health can be realised by consumers.
This practice is essential if we are to enact and enable the application of nutritional research for consumers. Indeed, it may be seen as the equivalent of extension practice in agriculture for management techniques because it is delivering products with very clear nutritional aims for the supply chain. An impressive example of applying research from nutritional trials to consumer requirements is the CSIRO Total Well Being Diet, where studies have determined optimal protein intake for healthy lifestyles.5 This diet is available to consumers as a book, and it communicates CSIRO's protein consumption research, CSIRO is the Australian Government science agency and this book has sold in excess of 700 000 copies. This diet shows there is significant interest in changing diets for healthier and more sustainable lifestyles, and there are routes to make this more likely that use nutritional trials to demonstrate outcomes. A similar approach has been used by the World Wide Fund for Nature (WWF) to promote not only healthy diet planning but increased sustainability criteria associated with healthier meals.6
At a food system world view, the micronutrients and phytonutrients in food supply chains are of critical importance because of the emergence of what are known as ‘hidden hungers’. These have become important in policy arenas that aim to increase food security by enhancing the value of meal planning and health outcomes associated with different diets. This is not the food security associated with the consumption of energy and protein alone, it is concerned with the balance or quality of foods consumed. Indeed, to many commentators, overlooking the ‘hidden hungers’ has been responsible for poor dietary policies in both economically undeveloped and developed nations. This is because the primary responses of policymakers who wish to reach food security are focused on the supply of calories and protein without a considering micronutrition impacts. Indeed, global statistics and data concerned with the production and trade of primary agricultural products and foods do not report phytonutrient quality of these materials. This lack of insight can ultimately lead to deficiencies in micronutrients that have been suggested to result in the overconsumption of calories and protein.7 The micronutrients for which there are well documented cases of ‘hidden-hunger’ impacts include iron and the relationship of intake and bioaccumulation with medical conditions such as anaemia and cognitive development.8
A well-documented hidden-hunger scenario is also provided by selenium, which is essential to human and animal health and not for crop growth. It is therefore not an essential plant nutrient whose accumulation in the supply chain is critical because it is an important human nutrient. Reported research suggests that deficiency in selenium can result in over consumption of foods because more food is eaten to obtain necessary selenium and other micronutrient intake. This type of hidden-hunger scenario is thought possible for most micronutrients and phytonutrients, and it has developed innovative views of what sustainable dietary advice could be.9
Thus, attention is placed on the mineral composition of agri-produce and the hidden-hunger scenarios because zinc, selenium, copper, and perhaps most globally important, iron content of crop biomass, is influenced by soils and the plant nutrients applied to crops. These micronutrients do vary in the composition of cereal produce around the world and reflect the fertilisation policy and soil composition to a large extent.10 The optimal mix of foliage, seed, and root tissues can also influence micronutrient intake, as can how foods are preserved or cooked. These attributes of efficient micronutrient delivery are continuously overlooked in nutritional policy, and the benefits of using biofortified crop varieties, such as golden rice have yet to be realised.11
The phytonutrient content of biomass is dependent on crop species and crop variety, and specific crops are often grown for particular phytonutrients for human consumption. Indeed, the development of different-coloured vegetable and fruit varieties is associated with differing phtyonutrient content. The selection of different foods for meals based on different colour and fragrance stimuli is an important route to consuming a healthy balance of phytonutrients. This approach of developing crop variety for specific nutritional properties has led to the production of crops for pharmaceutical and industrial feedstocks. An example is the development of increased phyto-oestrogen levels in the soybean crop; these are most notably present in soya extracts and are thought important for a number of metabolic functions including maintaining bone health.12 Other compounds include the brassinosteroids found in crucifers that are suggested to offers anti-carcinogenic benefits when they are consumed.13 Perhaps the most lauded phytonutrients of a specialised nature are the poly-unsaturated fatty acids (PUFAs) and mono-unsaturated fatty acids (MUFAs) because the fatty acid composition of agri-produce has become extremely important, with linola, borage, and meadowfoam oils being grown for health supplements. The production of oils that contain increased levels of MUFAs and PUFAs has become more important for reasons relating to the health of the human circulatory system. The oilseed crops that contain the omega unsaturated fatty acids at increased levels include nuts, such as walnut and hemp, and these oils are found in oily fish and algae. The associated health benefits of increased MUFA and PUFA diets were first recognised in populations that had increased consumption of oily fish products.14,15
Naturally, these types of nutrients are contained within the biomass that is produced by agricultural systems, and their concentration in biomass can be enhanced by using plant nutrients applied to crops as fertiliser, which is known as agrifortification. There is also a requirement for efficient supply chains to optimize the preservation of food nutritional value or add to it by fortifying biomass and food materials in the food chain. The agrifortification and biofortifcation of foods is discussed here in the context of enhancing the delivery of phytonutrients in diet, but we do need an understanding of how nutrients are accumulated in biomass within the agricultural system. The interaction between plant genes that result in the production of phytonutrients and human health will provide many future opportunities in bringing diets to consumers that are more focused to individuals' health requirements. The role of genetically engineering crops to provide these specific nutrients offers huge potential.16
3.3 Biomass, the Base of the Supply Chain
Understanding the principles of biomass production is crucial to understanding the nutritional interface between production and food manufacture because it not only provides calories and protein, but also the materials and cellular components in which micronutrients and phytonutrients are contained. This requires a thorough understanding of how crop nutrition and crop protection are used in farming systems around the world because the use of these inputs stimulate the production of biomass and interception of solar energy. Thus, agronomic practices will influence the quality of both plant and animal biomass produced from the farming system. The principles of producing biomass in the farming system for quantity and quality of food supply chain products can be placed in the context of what are called integrated management regimes, where all aspects of crop production are considered within the food system. This includes from the molecular scale of biochemistry and physiology through to the macroscale geography and land use planning impacts that are utilised to reach optimal management decisions.
Integrated management techniques are an important suite of knowledge and thinking used by producers in the food supply chain and they should be utilised by manufacturers and processors in future. We have already introduced the value of Integrated Plant Nutrition Systems here where integrated management brings together established routes for the optimal management of producing biomass for food supply and integrates them with emergent technologies and policy frameworks. The branches of integrated management include the following:
- Integrated Crop Management (ICM): The principles of ICM are associated with determining optimal energy inputs for crop yield that provide economic and environmental outcomes that enable sustainable business practice.17
- Integrated Pest Management (IPM); The principles of IPM aim to optimise the yield of crops and livestock by reducing the costs of pest control and the impact of disease. The use of scheduled pest control actions in response to observed pest thresholds are the focus of IPM.18
- Integrated Plant Nutrient Systems (IPNS): The principles of IPNS aim to optimise crop and livestock yield in response to plant nutrient management on farms. The use of nutrient management plans, nutrient balances, and fertiliser recommendations are the focus of IPNS.19
The integrated approach provides a system-thinking view of farming operations in that no single operation used to optimise yield is taken in isolation and the goal of sustainable yield is made possible by considering all data available to growers and farmers. An important consideration already mentioned that is often outside of the direct control of the producer is that of climate, where the interception of light, impact of ambient temperature, and acquisition of water have impacts on biomass yield. While out of direct control, understanding how these resources are used can point producers to potential areas of efficiency improvement within the agricultural system.
3.4 The Interception of Light by Crop Canopies: How the Molecular Scale Impacts on Food Supply Chain Efficiency
The interception of solar radiation will be dependent on the leaf area intercepting energy from sunlight, as the leaf area index (LAI) shows; although canopy structures and effects of shading do make this statement very simplistic, its importance is often overlooked. LAI is the area of leaf per unit ground cover, and the optimisation of LAI has been crucial in developing modern wheat varieties.20 The duration a leaf is expanded and effectively photosynthesising is also important in determining the amount of energy intercepted, and this is measured as the leaf area duration (LAD). There are very good correlations between light interception, dry matter increase, and LAI and LAD. Crops will increase LAI and LAD to a point where metabolism can be maintained, and surplus energy intercepted is diverted into dry matter gain; optimising this process through integrated management or crop breeding is clearly important to increasing crop yield. While this is a relatively simple explanation that considers many different environmental variables that determine leaf expansion and duration, the largest leaves of crops are proven to be important indicators of potential yield capacity.21 What research has achieved by understanding the interaction with LAI and LAD with yields is to make what could be just anecdotal observations in the field into a specific metabolic outcomes that relates to crop performance and eventual biomass yield. Measuring the effectiveness of light capture by canopies is an important indicator for crop yield, and the activity of leaf photosystems that convert solar energy into dry matter, understanding and measuring how the activity of these photosystems changes dry matter accumulation, has important implications for projecting biomass yield.
Global food production is dependent on the primary dry matter produced from capturing solar energy by expanded leaves. Now that we have an understanding of both the nutrient balance of agriculture and how plant nutrients can be managed, we should consider how crops produce biomass from these nutrients. The key processes to understand are what might be regarded as a suite of biochemical reactions that utilise solar energy to produce both carbohydrates and energy-transferring molecules that enable the synthesis of proteins, plant growth, and the accumulation of biomass. These energy transfers from solar to chemical energy sources are described by the metabolic reactions of photosynthesis, and the outcomes of these reactions are subject to the laws of thermodynamics. This is because photosynthesis is essentially an energy transferring activity where solar energy and heat is captured, which is not 100% efficient because there are many transfer and capture points where energy is dissipated as heat or entropy. This is an outcome of the second law of thermodynamics, which states energy can be transferred within a system but in doing so disorder or entropy is dissipated. At a macro- or physiological-scale energy transfer, inefficiencies are identified by the crop canopy interception of photosynthetically active radiation (PAR), where less than 5% PAR is typically intercepted by even the most efficient crop canopy. This is because PAR is made up of only the red and blue parts of the visible spectrum of light, and PAR must be captured by photosystems in the leaf; there are many different processes involved in getting the right type of energy to the right place—it cannot be a 100% efficient process.
At a molecular scale, the production of dry matter or biomass in plants is determined by the biochemical reactions that result in the synthesis and transport of carbohydrates that are produced from photosynthesis. The solar energy captured by crop leaf canopies is used by a number of relatively conserved chemical energy shunts that utilise the energy of electron transfer occurring in Light Harvesting Complexess (LHCs) within leaves that include pigments, such as chlorophyll. The electron transfer processes occurring in the LHCs are powered by the capture of PAR in crop canopies. This electron shunting results in the release of energy that is captured by soluble macromolecules, such as adenosine tri-phosphate (ATP) and nicotine nucleotides. While the biochemical processes described here are simplified for purposes of explanation, they have often been considered abstract when we consider the macroscale of field agronomy. However, understanding how photosynthesis is regulated by metabolic factors within leaves and environmental changes outside leaves is crucial to any agricultural management system, and it offers much future opportunity in stimulating management changes and enabling the development of improved crop varieties.
Indeed, this understanding of how molecular knowledge links to agronomic and food supply chain efficiency is the basis for the development of the bioengineering and biotechnology industries. The application of biotechnology can change both the macroscale factors, such as leaf expansion, and microscale factors, such as the transport of carbohydrate, that are influenced by the transfer of solar to chemical energy. Biotechnology has enabled the sequencing of crop genomes so that metabolic events associated with yield can be understood with respect to the regulation of specific genes. Furthermore, the establishment of genome knowledge has identified groups of genes that determine macroscale changes in crops such as leaf expansion, nutrient accumulation, and senescence or leaf duration.22 The metabolic responses to changes in the environmental variables, such as light intensity and temperature, also provide signals that can be detected by sensors, such as chlorophyll meters, that are used to determine crop health. Thus, the activity of specific genes or groups of genes that can be used to identify stress brought about by metabolic changes provides a type of sensing that can improve agronomic management.23
Photosynthesis is carried out by catalytic proteins or enzymes in plant leaves and other green tissues. The soluble enzymatic proteins that are not part of the photosystem proteins associated with membranes are functional proteins that enable the biochemical reactions that reduce carbon dioxide to carbohydrate, which in turn may accumulate to increase the dry matter in a crop. Other proteins have storage functions and accumulate in storage tissues, such as seeds; together with the accumulation of carbohydrates, the functional and storage protein content of crops often determine their nutritive value. The taste and texture attributes associated with crop ingredients are determined by many other metabolites, such as pigments and essential oils, but protein content is a good indicator of nutritional value. Many other metabolic processes will determine the eventual dry matter accumulation, including the ‘storage capacity’ of a crop to hold carbohydrate and its ability to transport carbohydrate to storage tissues. In many cases, crops have been bred by recognising these capture, transport, and storage traits, which are often observable in the field, to select for increased storage capacity for carbohydrate. Thus, the ability to observe what is known as ‘sink strength’ has been critical to the development of modern crop varieties because many of the storage tissues we harvest, such as seeds, tubers, and roots, are the result of selecting for increased sink strength and storage capacity which can be measured by field observation.
We are now able to define sink strength activities more precisely by using knowledge of the role of genes and groups of genes that control them. New biotechnological techniques in genomics can identify genes that control sink strength and provide targets for selection to improve performance in the field. An understanding of how the protein and carbohydrate content of plants are regulated in agricultural practice is critical if we are to breed more productive crops and are to understand the eventual nutritional qualities of foods and food stuffs.
3.5 The Requirement for Breeding New Crop Varieties and Selecting for Increased Sink Capacity of Crops
We know that sink strength is an extremely important component of yield determination and a complex parameter to quantify, and there are demonstrations of the ability of sink strength to influence yield and dry matter production. These are provided by greenhouse gas and global warming experiments, where plants are grown at increased CO2 concentrations. Acclimation to increased CO2 results in a decreased photosynthetic capacity over a longer term period. Acclimation results from changes in gene expression over days and not changes in metabolite concentration or enzyme activation that occur over minutes in response to environmental change. When photosynthetic tissue is harvested and allowed to regrow, such as for forage grasses, they experience a increase in sink strength even at increased CO2 concentrations and photosynthetic capacity increases. It is as if the photosynthetic system works harder at increased CO2 and the plant becomes clogged with assimilates, such as sucrose and starch; when the clogged sinks are removed, sink capacity increases again. Nitrogen can ameliorate this effect because plants grown with increased nitrogen continue to increase photosynthetic capacity at increased CO2; this is because sufficient nitrogen is available to develop new tissues and maintain sink strength. At increased CO2 and limiting nitrogen conditions, the acclimatory response of decreased photosynthetic capacity is amplified and more apparent than at high nitrogen availability. Thus, global warming and environmental change could have important implications for crop yield and nitrogen requirements of the future food system.
3.6 Photosynthetic Metabolism, the Biochemical Driver of Production
We now understand that photosynthesis requires proteins to catalyse reactions, and a typical protein contains 5% nitrogen and 1% sulfur by weight so they are substantial sinks for the nitrogen and sulfur nutrients in crops. These proteins, together with storage proteins, are the source of nutritional variation in many foods. The development of crops that preferentially store protein or carbohydrate in sink or storage tissue is the basis for growing many of the crops we utilise for protein or carbohydrate sources in food processing and manufacture. The protein investment made in photosynthesis is large, with a single photosynthetic protein, the Rubisco protein, making up 25% of a typical leaf protein quota. Rubisco is an acronym for ribulose-1,5-bisphosphate carboxylase-oxygenase; it is a soluble protein contained in chloroplasts within photosynthetic cells that fixes carbon dioxide gas into a reduced carbohydrate form that is synthesised into what we call sugar or sucrose, the white crystalline sugar utilised by the food system.
Sucrose is the universal carbohydrate currency of all plants, and it is a disaccharide of glucose and fructose that can be metabolised to produce transferrable energy or stored as a glucose polymer as starch. But it is the protein allocation of photosynthesis that concerns us here because the concentration of Rubisco protein in a leaf may provide a means of assessing the productivity of a crop by again acting as a sensor or indicator for production of dry matter. A further substantial protein pool within the leaf is the insoluble membrane bound or insoluble proteins within the chloroplast that are associated with light harvesting complex (LHC) or photosystem functions and pigments, such as chlorophyll and carotene. This group of proteins can make up 40% of the leaf protein total. The total photosystem insoluble protein and soluble enzyme protein in a typical leaf can account for at least 70% of the protein of a plant, and this represents a major requirement for nitrogen, sulfur, and other essential plant nutrients. Thus, the protein allocation and physiological responses of photosynthesis are important targets for methods that will help agriculturalists manage crops for the food supply chain and provide sensors for productivity.24 What is crucial in all of this is that we not only account for quantity but also for quality when we consider crop yields.
The relationship between nitrogen and leaf biochemistry demonstrates that nitrogen can substantially limit production, but above a critical leaf concentration of nitrogen, the benefits of obtaining more nitrogen are negligible. We have already discussed this with respect to increased atmospheric carbon dioxide concentration and acclimatory responses of crops to environmental stresses. The acclimatory responses of plants have implications for how nitrogen is mobilised in canopies, how much nitrogen needs to be supplied to a crop, and how much is stored in harvested biomass. The interaction between nitrogen use and distribution within the canopy cover and architecture of plants offers a means of utilising crops as sensors in optimal production systems. The basic metabolic responses of crops will be generic across most cultivated species; however, subtle but important differences exist that can provide specific targets for crop improvement. The following examples of metabolic targets demonstrate how biotechnologies can utilise natural systems to manipulate yield and productivity in agricultural systems:
- Water stress is a target for agronomic improvement because crop varieties that are able to tolerate increased levels of water stress will enable the most efficient use of land experiencing climate change25–27
- Nitrogen responders are those crops that can either provide sensors for specific nutrient requirements (in this case nitrogen) or those crops that will accumulate dry matter most effectively in response to nitrogen. They are of agronomic value because of the strong relationship between nitrogen use efficiency and crop yield.28
- Pest control using biotechnological techniques, such as inducing crops to produce pheromones that deter insect pests or develop metabolic responses that reduce the impact of diseases offer new routes to deploying crop protection strategies.29,30
- Nutrient composition of crops has been identified as an area of improvement opportunities for biofortifying crops, and with the use of biotechnologies, nutritional improvement of crops will become more innovative. This is apparent with respect to now being able to design crops with specific nutrient profiles such as those having different oil composition in their seeds.31
3.7 Environmental Stress Events and Their Impacts on Food Supply
An important area where metabolism, environmental stressors, and biotechnological opportunities exist is the understanding of how crops respond to changes in light and temperature. The combination of these two environmental variables can result in decreased temperature, which slows metabolism, and increased light intensity, which increases photoreactions of photosynthesis. These low temperature and high light conditions are not uncommon in field situations, and they result in photoinhibition in plants. In effect, plants have developed sun protection by modifying pigment concentrations in the LHCs and membranes structures to dissipate excess solar energy as heat and protect metabolic processes from photoinhibition. Photoinhibition results in the production of previously mentioned free radicals whose damaging activities are reduced by specific antioxidants that are included in the phytonutrient group of metabolites we have already discussed.
The mechanisms of nutrient partitioning have been investigated since the 1960s with regard to leaf and canopy structures, and the biochemical elucidation of photosynthesis has enabled an understanding of carbohydrate synthesis in low temperature and high light environments to be understood in the 1980s. Biotechnological techniques enabled manipulation of pigment genes that conferred elements of low temperature resistance and increased acclimation in the 1990s. The development of low temperature and frost resistance in crops is of clear agronomic application since susceptibility to low temperature during germination and grain filling can have detrimental impact on yield. Indeed, the following case study shows how biotechnological techniques have enabled, firstly, understanding low temperature responses of crops, and then to target specific metabolic pathways that enable crops to acclimate or protect cells from low and freezing temperatures.32
Low temperature decreases photosynthetic capacity, and this can result in the acclimation of crops to lower temperatures. Acclimation will result in an increase in the ability of leaves to carry out gas exchange, that is, carbon dioxide entering and water vapour being removed from the leaves via the stomata. The response to decreased temperature will be associated with a number of variables, including the acclimation of stomata and many metabolic changes. These responses can be limited by protein synthesis, and this can be demonstrated using the photosynthetic response curves that show a limiting factor response we have seen previously. These show that acclimation to low temperature and high light is associated with an increase in the rate of carbon assimilation and the capacity to utilise photosynthetic metabolites in cells. An increase in the capacity of gas exchange of the leaf during photosynthesis is related to increased amounts of photosynthetic enzymes within the leaf. An important enzyme is Rubisco, which is a sensor for crop health and production capacity; it makes up between 20% and 30% of the leaf soluble protein and therefore is easy to detect and its activity is easily measured. Placing these diagnostic methods from laboratories into field situations is challenging, but the detection of the amount of Rubisco in leaves or greenness of leaves may provide a means of determining the health and acclimation potential of crops. An important implication of acclimation to low temperature is that increases in enzyme activities will require a greater amount of protein synthesis, and therefore nitrogen availability must be increased or nitrogen use efficiency improved.33 This is similarly the case for phosphate use in photosynthetic cells, since the amount of sugar phosphate metabolites in cold-acclimated leaves is known to increase. An increase in the capacity to synthesis soluble sugars, principally sucrose in many plants, is closely related to the ability for crops to acclimate to low temperature where sugars such as sucrose can act as cryoprotectants that maintain the integrity of cell membranes.34 Thus, a consequence of low temperature acclimation is the production of free sugars such as sucrose in cells that confer cryoprotective effects on membranes within cells. High concentrations of sucrose increase the viscosity of fluid around membranes, thus protecting them against the disruptive formation of ice crystals. This principle is used in food preservation and freezing, where the integrity of foods can be maintained during cold treatment by maintaining free sugar concentrations. Yet again, this type of process can be influenced by agronomic management of biomass in that foliage can be low temperature sweetened prior to freezing so that the concentration of free sugars is increased.
This has implications for plant storage organs that are cold stored, such as potatoes, sugar beet, and many fruit and vegetable crops. The storage and low temperature sweetening of potatoes, fruits, and vegetables for the retail market is a recognised characteristic associated with changes in taste. There has been substantial interest in finding out what determines cold sweetening in potatoes that undergo storage periods at low temperature. Sweetening has implications for the processing industry because free reducing sugars in potato tissue can cause discolouration during frying of potato chips. Increased sugar synthesis in plants at low temperature may also provide insights into the improved storage of phytonutrients since specific sugars, such as trehalose, synthesised in plants and microbes have extremely good cryoprotective properties.
The light environment of leaves can stimulate acclimatory responses of the light capture systems and other photosynthetic components of a leaf if the light environment is decreased or increased in intensity within a canopy. Under such conditions, photoacclimatory responses occur to maintain photosynthetic capacity, and if acclimation does not occur, photosystems in leaves become super energised with no sink for absorbed PAR and energy, resulting in increased photoinhibiton processes within a leaf, which can decrease productivity. Light is rarely thought of limiting photosynthesis when it is at high intensities, although photoinhibition has been shown to occur in all plant species to some degree at all times if photosystems are operating at maximum rates. There are metabolic mechanisms that result from photoinhibition of the light capturing systems in leaves that cause the degradation of proteins involved in electron transport, thus decreasing energy capture and photosynthetic production. Furthermore, there is a metabolic mechanism known as photorespiration that results in the consumption of oxygen and results in the release of CO2 and an ammonia molecule. This process of photorespiration is thought to be important in dissipating light energy at high light intensities. Many temperate crops will experience photoinhibitory conditions of high light and low temperature, particularly if they are autumn sown. Hence, understanding the relationship between photosynthesis, photoinhibition, and photorespiration with environmental variables can help us understand how crop metabolism responds to these particular stresses. These types of investigation may also identify metabolic indicators of stress that can be utilised by agronomists.
3.8 The Principles of Integrated Management across the Food Chain: A Food Supply Chain Perspective
Integrated management of pests, nutrients, and crops depends on a number of different approaches to optimising yield being utilised to develop profitable and sustainable farms. They can include the following headline actions.
- The use of Geographic Information Systems (GIS's) has started to replace traditional field maps and have the potential to revolutionise farming practices. GIS have provided more fit-for-purpose means of using maps within an Information Technology (IT) framework; the map remains the same whether it is digital or on paper or transparency formats. Indeed, this is the principle of precision agriculture and the techniques associated with it that target agricultural inputs and outputs to specific areas. The use of soil and plant sensors to target inputs is becoming important in farming systems, and metabolic sensors have been discussed. The use of precision agricultural techniques for outputs has focused on harvesting efficiency and transport of products. All farm maps will include general reference points, including streams, residences, well heads, number of hectares, and soil types. This is the basis for the rest of the plan. The use of GIS systems to map farms is becoming more common as computer power increases and their price decreases. Maps of sufficient resolution for farms are available free or at minimal cost on the World Wide Web and ‘off-the-shelf’ software providing GIS applications. GIS software has become compatible with other standard software packages that handle spreadsheets and databases so that a GIS system can be interfaced with other expert systems. These systems are able to store long-term records for assurance and traceability purposes. Furthermore, GIS systems can be utilised to map activities across food supply chains from producers to consumer functions.
- Soil and agricultural product testing is critical because this will determine how much nutrient is being utilised as N, P, K, trace nutrients and provide assessment of pH and SOM so that nutrient balances can be developed. Again, the use of analytical data provides a means to use sensor-related and threshold values for developing management actions, such as the use of fertiliser, pesticides, and processing operations.
- Cropping plans should be prepared to represent market trends and the environmental sustainability of farming enterprises. This will include assessment of estimated yield; factors affecting yield are numerous and complex. Using historic series of yield records is important in developing yield estimates, and the more accurate the yield estimate, the easier it is to potentially optimise input use efficiency. Furthermore, the use of market information regarding the price trends and volatility agricultural products and food ingredients will be utilised in determining crop plans.
3.9 The Modern Agricultural System, the Dietary Interface, and Food Supply
Modern agriculture exists in a rapidly changing commercial, regulatory, and economic environment where new technologies, genetically modified crops, food traceability and safety, environmental quality, and financial survival in global markets must be balanced within businesses. The only way to achieve this is by considering the farm enterprise as a system where technologies enable the production of biomass, improve crop and livestock production, and result in improved environmental management. The food production systems of the world depend upon primary production of dry matter or biomass by plants to provide ingredients for the manufacturing and processing functions of the food supply chain. Biomass and crop products are consumed in our food chain directly by us, or via livestock as meat and dairy produce or recycled within the biosphere as an energy source. The integration of biological knowledge into understanding plant production systems is an important goal for growers of biomass to strive for. The historical development of farming systems shows us that extension and agri-education are an important part of disseminating management knowledge, particularly when they are concerned with new technologies and methods of utilising food ingredients. Extension of biological principles to the food supply chain requires a framework of analysis, such as LCA or energy balance to incorporate economic, environmental, and agronomic decisions together so that they can be measured and assessed in terms of their performance.
The basis for any decision made in the food supply chain made by the farmer, manufacturer, retailer, or consumer must integrate economic, social, and environmental considerations. Here is our first and foremost difficulty in understanding the balance of both quantitative and qualitative attributes that result in management decisions. Ultimately, the decisions made will be an integration of several sources and disciplines. What a formalised integrative approach, such as LCA and energy balance, does is to provide terms of reference that performance can be measured against. This provides a platform to explain decisions and evaluate them in increasingly important arenas, such as the global trade of products, where supply chains are held to account by regulators for actions around the world that need to be reported to achieve optimal profitability, communicate responsibility, and ensure food safety.
Interfacing the food supply chain with the initial points of field trials and agricultural operations is critical because we have seen that how crops are grown drastically influences food quantity and quality. It is possible to detect metabolic changes in crops at field and landscape scales so that these foods-based quality attributes can be managed. This means precision techniques that map inputs and outputs from cultivation to manufacturing and processing are valuable tools that will tackle threats to yield loss and enhance the ability to develop management systems that can meet the demands of the food supply chain.
References
- 1 Scarseth, G. D. (1962). Man and his earth. Ames: Iowa State University Press.
- 2 Bøckman, O. C., Kaarstad, O., Lie, O. H., & Richards, I. (1990). Agriculture and fertilizers. CAB Direct Record Number 19901948903. Oslo: Norsk Hydro AS.
- 3 Martin, C., Butelli, E., Petroni, K., & Tonelli, C. (2011). How can research on plants contribute to promoting human health? The Plant Cell Online, 23(5), 1685–1699.
- 4 Ames, B. N. (2004). A role for supplements in optimizing health: the metabolic tune-up. Archives of Biochemistry and Biophysics, 423(1), 227–234.
- 5 Noakes, M., & Clifton, P. M. (2006). The CSIRO total wellbeing diet. Camberwell, Australia: Penguin.
- 6 Macdiarmid, J., Kyle, J., Horgan, G., Loe, J., Fyfe, C., Johnstone, A., & McNeill, G. (2011). Livewell: a balance of healthy and sustainable food choices. Livewell report 2011. London: WWF UK.
- 7 Ames, B. N. (2006). Low micronutrient intake may accelerate the degenerative diseases of aging through allocation of scarce micronutrients by triage. Proceedings of the National Academy of Sciences, 103(47), 17589–17594.
- 8 McCann, J. C., & Ames, B. N. (2007). An overview of evidence for a causal relation between iron deficiency during development and deficits in cognitive or behavioral function. The American Journal of Clinical Nutrition, 85(4), 931–945.
- 9 Ames, B. N. (2006). Low micronutrient intake may accelerate the degenerative diseases of aging through allocation of scarce micronutrients by triage. Proceedings of the National Academy of Sciences, 103(47), 17589–17594.
- 10 Welch, R. M., & Graham, R. D. (2004). Breeding for micronutrients in staple food crops from a human nutrition perspective. Journal of Experimental Botany, 55(396), 353–364.
- 11 Martin, C., Butelli, E., Petroni, K., & Tonelli, C. (2011). How can research on plants contribute to promoting human health? The Plant Cell Online, 23(5), 1685–1699.
- 12 Branca, F. (2003, November). Dietary phyto-oestrogens and bone health. Proceedings: Nutrition Society of London, 62(4), 877–887). CABI Publishing. 1999.
- 13 Moreau, R. A., Whitaker, B. D., & Hicks, K. B. (2002). Phytosterols, phytostanols, and their conjugates in foods: structural diversity, quantitative analysis, and health-promoting uses. Progress in Lipid Research, 41(6), 457–500.
- 14 McCann, J. C., & Ames, B. N. (2005). Is docosahexaenoic acid, an n−3 long-chain polyunsaturated fatty acid, required for development of normal brain function? An overview of evidence from cognitive and behavioral tests in humans and animals. The American Journal of Clinical Nutrition, 82(2), 281–295.
- 15 Kris-Etherton, P. M., Harris, W. S., & Appel, L. J. (2003). Fish consumption, fish oil, omega-3 fatty acids, and cardiovascular disease. Arteriosclerosis, Thrombosis, and Vascular Biology, 23(2), e20–e30.
- 16 Martin, C. (2012). The interface between plant metabolic engineering and human health. Current Opinion in Biotechnology, 24(2), 344–353.
- 17 Leake, A. (2000). The development of integrated crop management in agricultural crops: comparisons with conventional methods. Pest Management Science, 56(11), 950–953.
- 18 Barzman, M., & Dachbrodt-Saaydeh, S. (2011). Comparative analysis of pesticide action plans in five European countries. Pest Management Science, 67(12), 1481–1485.
- 19 Dicks, L. V., Bardgett, R. D., Bell, J., Benton, T. G., Booth, A., Bouwman, J., Brown, C., Bruce, A., Burgess, P. J., Butler, S. J., Crute, I., Dixon, F., Drummond, C., Freckleton, R. P., Gill, M., Graham, A., Hails, R. S., Hallett, J., Hart, B., Hillier, J. G., Holland, J. M., Huxley, J. N., Ingram, J. S. I., King, V., MacMillan, T., McGonigle, D.F., McQuaid, C., Nevard, T., Norman, S., Norris, K., Pazderka, C., Poonaji, I., Quinn, C. H., Ramsden, S. J., Sinclair, D., Siriwardena, G. M., Vickery, J.A., Whitmore, A. P., Wolmer, W. & Sutherland, W. J. (2013). What do we need to know to enhance the environmental sustainability of agricultural production? A prioritisation of knowledge needs for the UK food system. Sustainability, 5(7), 3095–3115.
- 20 Austin, R. B. (1999). Yield of wheat in the United Kingdom: recent advances and prospects. Crop Science, 39(6), 1604–1610.
- 21 Evans, L. T., & Fischer, R. A. (1999). Yield potential: its definition, measurement, and significance. Crop Science, 39(6), 1544–1551.
- 22 Varshney, R. K., Hoisington, D. A., & Tyagi, A. K. (2006). Advances in cereal genomics and applications in crop breeding. Trends in Biotechnology, 24(11), 490–499.
- 23 Varshney, R. K., Graner, A., & Sorrells, M. E. (2005). Genomics-assisted breeding for crop improvement. Trends in Plant Science, 10(12), 621–630.
- 24 Pirie, N. W. (ed.) (2012). Food protein sources (Vol. 4). Cambridge, UK: Cambridge University Press.
- 25 Roy, S. J., Tucker, E. J., & Tester, M. (2011). Genetic analysis of abiotic stress tolerance in crops. Current Opinion in Plant Biology, 14(3), 232–239.
- 26 Eisenstein, M. (2013). Plant breeding: discovery in a dry spell. Nature, 501(7468), S7–S9.
- 27 Peleg, Z., Apse, M. P., & Blumwald, E. (2011). Engineering salinity and water-stress tolerance in crop plants: getting closer to the field. Advances in Botanical Research, 57, 405–443.
- 28 Schachtman, D. P. (2012). Recent advances in nutrient sensing and signaling. Molecular Plant, 5(6), 1170–1172.
- 29 Gross, M. (2011). New directions in crop protection. Current Biology, 21(17), R641–R643.
- 30 Aliferis, K. A., & Jabaji, S. (2011). Metabolomics—a robust bioanalytical approach for the discovery of the modes-of-action of pesticides: a review. Pesticide Biochemistry and Physiology, 100(2), 105–117.
- 31 Haslam, R. P., Ruiz-Lopez, N., Eastmond, P., Moloney, M., Sayanova, O., & Napier, J. A. (2013). The modification of plant oil composition via metabolic engineering: better nutrition by design. Plant Biotechnology Journal, 11(2), 157–168.
- 32 Guy, C. L. (1990). Cold accelimation and freezing stress tolerance: role of protein metabolism. Annual Review of Plant Biology, 41(1), 187–223.
- 33 Holaday, A. S., Martindale, W., Alred, R., Brooks, A. L., & Leegood, R. C. (1992). Changes in activities of enzymes of carbon metabolism in leaves during exposure of plants to low temperature. Plant Physiology, 98(3), 1105–1114.
- 34 Martindale, W., & Leegood, R. C. (1997). Acclimation of photosynthesis to low temperature in Spinacia oleracea L. I. Effects of acclimation on CO2 assimilation and carbon partitioning. Journal of Experimental Botany, 48(10), 1865–1872.