Communication is important for organisms and involves transduction of stimulatory or inhibitory signals from other cells, organisms, or the environment. Unicellular organisms detect and respond to environmental signals. For example, they can use cell communication to locate a suitable mate, adapt metabolism rates in response to changes in nutrient availability, or make their numbers known to other members of their species (a phenomenon known as quorum sensing).
Taxis is the movement of an organism in response to a stimulus and can be positive (toward the stimulus) or negative (away from the stimulus). Taxes are innate behavioral responses, or instincts. Chemotaxis is movement in response to chemicals. For example, bacteria can control flagella rotation to direct their motion, thus avoiding repellents (such as poisons), or helping them find favorable locations with high concentrations of attractants (such as food). In our bodies, neutrophils use chemotaxis to respond to an infection and are the first responders to inflammation.
The cells of multicelled organisms must communicate with one another to coordinate the activities of the organism as a whole. Signaling can be short-range (affecting only nearby cells) or long-range (affecting cells throughout the organism). It can be done by cell junctions or signaling molecules called ligands that bind to receptors and trigger a response. Hormones and neurotransmitters are common signaling molecules, which will be covered in Chapter 9.
Signal transduction is the process by which an external signal is transmitted to the inside of a cell. It usually involves the following three steps:
a signaling molecule binding to a specific receptor
activation of a signal transduction pathway
production of a cellular response
Hydrophobic signaling molecules, like cholesterol-based steroid hormones, can diffuse across the plasma membrane. These molecules can bind receptors inside the cell and often travel to the nucleus where they regulate gene expression.
For signaling molecules that cannot enter the cell, a plasma membrane receptor is required. Plasma membrane receptors form an important class of integral membrane proteins that transmit signals from the extracellular space into the cytoplasm. Each receptor binds a particular molecule in a highly specific way. There are three classes of membrane receptors.
Ligand-gated ion channels in the plasma membrane open an ion channel upon binding a particular ligand. An example is the ligand-gated sodium channel on the surface of a skeletal muscle cell at the neuromuscular junction. This channel opens in response to acetylcholine, and a massive influx of sodium depolarizes the muscle cell and causes it to contract.
Catalytic (enzyme-linked) receptors have an enzymatic active site on the cytoplasmic side of the membrane. Enzyme activity is initiated by ligand binding at the extracellular surface. The insulin receptor is an example of an enzyme-linked receptor. After binding insulin, enzymatic activity initiates a complex signaling pathway that allows the cell to grow, synthesize lipids, and import glucose.
A G-protein-linked receptor does not act as an enzyme, but instead will bind a different version of a G-protein (often GTP or GDP) on the intracellular side when a ligand is bound extracellularly. This causes activation of secondary messengers within the cell. One important second messenger is cyclic AMP (cAMP). It is known as a “universal hunger signal” because it is the second messenger of the hormones epinephrine and glucagon, which cause energy mobilization. Secondary messengers are usually small molecules that can move quickly through the cell. They can be made and destroyed quickly and help the signal amplify throughout the cell.
Signal transduction in eukaryotic cells usually involves many steps and complex regulation. Many signal transduction pathways include protein modifications and phosphorylation cascades, in which a series of protein kinases add a phosphate group to the next protein in the cascade sequence. In contrast, bacterial cells usually use a simpler two-component regulatory system in transduction pathways.
Signal transduction cascades are helpful to amplify a signal. They are also helpful in quickly turning a response on or off, and can lead to important changes in gene expression and cell function.
Mutations in the receptor or the ligand can alter signaling. Drugs that inhibit or excite part of the pathway can also influence the gene expression within the cell.
All living things have a “steady state” that they try to maintain. This is helpful because the structures and processes within living things are sensitive to things like temperature, pH, pressure, salinity, osmotic pressure, and many other things. The set of conditions under which living things can successfully survive is called homeostasis. The body is constantly working to maintain this state by taking measurements and then responding appropriately. That might mean shivering if it is cold or releasing the hormone insulin if blood glucose is high. Depending on the condition, the body will respond accordingly.
Many of these responses are controlled by negative or positive feedback pathways. A negative feedback pathway (also called feedback inhibition) works by turning itself off using the end product of the pathway. The end product inhibits the process from beginning, thus shutting down the pathway. This is a common strategy to conserve energy. If plenty of X is made, then the pathway to make X can be turned off. Sometimes, a very distant end product will turn off a process occurring several steps back. For example, ATP turns off the beginning stages of glycolysis, although ATP is not made until much later in cellular respiration.
A positive feedback pathway also involves an end product playing a role, but instead of inhibiting the pathway, it further stimulates it. For example, once X is made, it tells the pathway to make even more X. This is less common, but it occurs during fruit ripening and labor and delivery, which ramps up and up as it proceeds.
Plants do not have a nervous system but can produce several proteins found in animal nervous systems, such as certain neurotransmitter receptors. Plants can also generate electrical signals in response to environmental stimuli, and this can affect flowering, respiration, photosynthesis, and wound healing. Light receptors are common in plants and help link environmental cues to biological processes such as seed germination, the timing of flowering, and chlorophyll production. Some plants can also use chemicals to communicate with nearby plants. For example, wounded tomatoes produce a volatile chemical as an alarm signal. This warns nearby plants and allows them to prepare a defense or immune response.
Every second, thousands of cells are dying throughout our bodies. Fortunately, the body replaces them at an amazing rate. In fact, epidermal cells, or skin cells, die off and are replaced so quickly that the average 18-year-old grows an entirely new skin every few weeks. The body keeps up this unbelievable rate thanks to the mechanisms of cell division.
The remainder of this chapter takes a closer look at how cells divide. But remember, cell division is only a small part of the life cycle of a cell. Most of the time, cells are busy carrying out their regular activities. There are also some types of cells that are nondividing. Often, these are highly specialized cells that are created from a population of less specialized cells. The body continually makes them as needed, but they do not directly replicate themselves. Red blood cells are an example of nondividing cells. Some other cells are just temporarily nondividing. They enter a phase called G0, where they hang out until they get a signal to reenter the normal cell cycle.
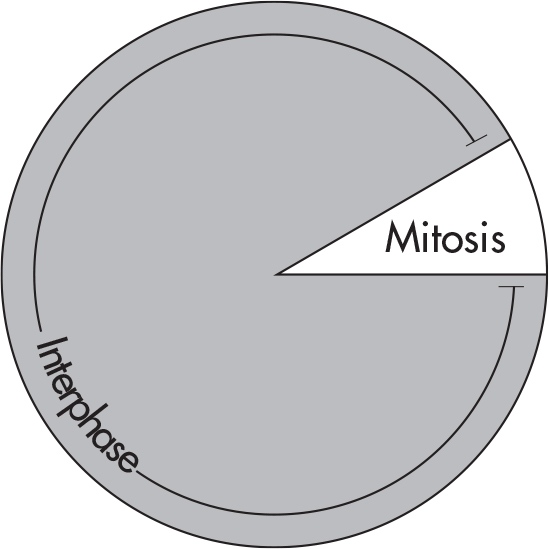
Every cell has a life cycle—the period from the beginning of one division to the beginning of the next. The cell’s life cycle is known as the cell cycle. The cell cycle is divided into two periods: interphase and mitosis. Take a look at the cell cycle of a typical cell.
Interphase is the time span from one cell division to another. We call this stage interphase (inter- means between) because the cell has not yet started to divide. Although biologists sometimes refer to interphase as the “resting stage,” the cell is definitely not inactive. During this phase, the cell carries out its regular activities. All the proteins and enzymes it needs to grow are produced during interphase.
Interphase can be divided into three stages: G1, S, G2.

The most important phase is the S phase. That’s when the cell replicates its genetic material. The first thing a cell has to do before undergoing mitosis is to duplicate all of its chromosomes, which contain the organism’s DNA “blueprint.” During interphase, every single chromosome in the nucleus is duplicated.

You’ll notice that the original chromosome and its duplicate are still linked, like conjoined twins. These identical strands of DNA are now called sister chromatids. The chromatids are held together by a structure called the centromere. You can think of each chromatid as a chromosome, but because they remain attached, they are called chromatids instead. To be called a chromosome, each needs to have its own centromere. Once the chromatids separate, they will be full-fledged chromosomes.
We’ve already said that replication occurs during the S phase of interphase, so what happens during G1 and G2? During these stages, the cell performs metabolic reactions and produces organelles, proteins, and enzymes. For example, during G1, the cell produces all of the enzymes required for DNA replication (DNA helicase, DNA polymerase, and DNA ligase, which are explained later in this chapter). By the way, G stands for “gap,” but we can also associate it with “growth.” These three phases are highly regulated by checkpoints and special proteins called cyclins and cyclin-dependent kinases (CDKs).
Cell cycle checkpoints are control mechanisms that make sure cell division is happening properly in eukaryotic cells. They monitor the cell to make sure it is ready to progress through the cell cycle and stop progression if it is not. In eukaryotes, checkpoint pathways function mainly at phase boundaries (such as the G1/S transition and the G2/M transition). Checkpoint pathways are an example of an important cell signaling pathway inside cells.
For example, if the DNA genome has been damaged in some way, the cell should not divide. If it does, damaged DNA will be passed onto daughter cells, which can be disastrous. When damaged DNA is found, checkpoints are activated and cell cycle progression stops. The cell uses the extra time to repair damage in DNA. If the DNA damage is so extensive that it cannot be repaired, the cell can undergo apoptosis, or programmed cell death. Apoptosis cannot stop once it has begun, so it is a highly regulated process. It is an important part of normal cell turnover in multicellular organisms.
Cell cycle checkpoints control cell cycle progression by regulating two families of proteins: cyclin-dependent kinases (CDKs) and cyclins. To induce cell cycle progression, an inactive CDK binds a regulatory cyclin. Once together, the complex is activated, can affect many proteins in the cell, and causes the cell cycle to continue. To inhibit cell cycle progression, CDKs and cyclins are kept separate. CDKs and cyclins were first studied in yeast, unicellular eukaryotic fungi. Budding yeast and fission yeast were good model organisms because they have only one CDK protein each and a few different cyclins. This relatively simple system allowed biologists to figure out how cell cycle progression is controlled. Their findings were then used to figure out how more complex eukaryotes (such as mammals) control the cell cycle.
A cell losing control of the cell cycle can have disastrous consequences. For example, a mutation in a protein that normally controls progression through the cell cycle can result in unregulated cell division and cancer. Cancer means “crab,” as in the zodiac sign. The name comes from the observation that malignant tumors grow into surrounding tissue, embedding themselves like clawed crabs. Cancer occurs when normal cells start behaving and growing very abnormally and spread to other parts of the body.
Mutated genes that induce cancer are called oncogenes (“onco-” is a prefix denoting cancer). Normally, these genes are required for proper growth of the cell and regulation of the cell cycle. Oncogenes, then, are genes that can convert normal cells into cancerous cells. Often these are abnormal or mutated versions of normal genes. The normal healthy version is called a proto-oncogene.
Tumor suppressor genes produce proteins that prevent the conversion of normal cells into cancer cells. They can detect damage to the cell and work with CDK/cyclin complexes to stop cell growth until the damage can be repaired. They can also trigger apoptosis if the damage is too severe to be repaired.
Cell cycle checkpoints and apoptosis have been intimately linked with cancer. In order for a normal cell to become a cancer cell, it must override cell cycle checkpoints and grow in an unregulated way. It must also avoid cell death. Tumors often accumulate DNA damage. Cancer treatments target these changes, because they are what make a cancer cell different from a normal cell.
Let’s recap:
The cell cycle consists of two things: interphase and mitosis.
During the S phase of interphase, the chromosomes replicate via DNA replication.
Growth and preparation for mitosis occur during the G1 and G2 stages of interphase.
Cell cycle checkpoints make sure the cell is ready to continue through the cell cycle.
CDK and cyclin proteins work together to promote cell cycle progression.
Oncogenes promote cell growth and tumor suppressor genes inhibit cell growth.
Once the chromosomes have replicated, the cell is ready to begin mitosis—the period during which the cell divides. Mitosis consists of a sequence of four stages: prophase, metaphase, anaphase, and telophase. It is not important to memorize the name of each phase, but it is important that you know the basic order of operations for mitosis.
One of the first signs of prophase is the disappearance of the nucleolus (ribosome-making area) and the nuclear envelope. In addition, the genome becomes visible as individual chromosomes; diffuse chromatin (which looks like a pile of scrunched up dental floss in the nucleus) condenses into densely packed chromosomes. To accomplish this, chromatin forms coils upon coils, and thickens. If you look at a human cell under the light microscope at the beginning of prophase, you would see 46 differently sized chromosomes, and each one would be the shape of an X. During interphase, individual chromosomes are not visible.

Now the cell has plenty of room to “sort out” the chromosomes. Remember centrioles? During prophase, these cylindrical bodies found within microtubule organizing centers (MTOCs) start to move away from each other, toward opposite ends of the cell. The centrioles will spin out a system of microtubules known as the spindle fibers. These spindle fibers will attach to a kinetochore, a structure on each chromatid. The kinetochores are part of the centromere.
The next stage is called metaphase. The chromosomes now begin to line up along the equatorial plane, or the metaphase plate, of the cell. This happens because spindle fibers are attached to the kinetochore of each chromatid.

During anaphase, sister chromatids of each chromosome separate at the centromere and migrate to opposite poles. Chromatids are pulled apart by the microtubules, which begin to shorten. Each half of a pair of sister chromatids now moves to opposite poles of the cell. Non-kinetochore microtubules elongate the cell.

The final phase of mitosis is telophase. A nuclear membrane forms around each set of chromosomes and the nucleoli reappear.

The nuclear membrane is ready to divide. Now it’s time to split the cytoplasm in a process known as cytokinesis. Look at the figure below and you’ll notice that the cell has begun to split along a cleavage furrow (which is produced by actin microfilaments).

A cell membrane forms around each cell, and the cell splits into two distinct daughter cells. The division of the cytoplasm yields two daughter cells.
Here’s one thing to remember: cytokinesis occurs differently in plant cells. The cell doesn’t form a cleavage furrow. Instead, a partition called a cell plate forms down the middle region.
Once daughter cells are produced, they reenter the initial phase—interphase—and the whole process starts over. The cell goes back to its original state. Once again, the chromosomes decondense and become invisible, and the genetic material is called chromatin again.
For mitosis, you may already have your own mnemonic. If not, here’s a table with a mnemonic we created for you.
|
IPMAT |
|
|
Interphase |
I is for Interlude |
|
Prophase |
P is for Prepare |
|
Metaphase |
M is for Middle |
|
Anaphase |
A is for Apart |
|
Telophase |
T is for Two |
Mitosis achieves two things:
the production of daughter cells that are identical copies of the parent cell
maintaining the proper number of chromosomes from generation to generation
The impetus to divide occurs because an organism needs to grow, a tissue needs repair, or asexual reproduction must take place. For our purposes, we can say that mitosis occurs in just about every cell except sex cells. When you think of mitosis, remember: “Like begets like.” Hair cells “beget” other hair cells, skin cells “beget” other skin cells, and so on. Mitosis is involved in growth, repair, and asexual reproduction.
quorum sensing
taxis
chemotaxis
ligands
receptors
signal transduction
ligand-gated ion channel
catalytic (or enzyme-linked) receptor
G-protein-linked receptor
homeostasis
negative feedback pathway (feedback inhibition)
positive feedback pathway
cell division
G0 phase
cell cycle
interphase
mitosis
G1 phase
S phase
G2 phase
sister chromatids
centromere
cyclins
cyclin-dependent kinases (CDKs)
cell cycle checkpoints
apoptosis
cancer
oncogene
tumor suppressor gene
prophase
metaphase
anaphase
telophase
chromatin
spindle fibers
kinetochores
metaphase plate
cytokinesis
cleavage furrow
cell plate
Sometimes cells do not need to pass any molecules through the membrane, but instead they just pass a message. This is signal transduction. A ligand binds to a receptor on the outside of the cell and causes changes to the inside of the cell. Ligand-gated ion channels, catalytic receptors, and G-protein-linked receptors are common examples.
The cell cycle is divided into interphase and mitosis, or cellular division.
The three stages of interphase are G1, G2, and S phase.
S phase is the “synthesis” phase, when chromosomes replicate.
Growth and preparation for mitosis occur in G1 and G2.
Cell cycle progression is controlled by checkpoint pathways and CDK/cyclin complexes.
Cancer occurs when cells grow abnormally and spread to other parts of the body. Tumor-suppressor genes are genes that prevent the cell from dividing when it shouldn’t. Proto-oncogenes are genes that help the cell divide. If either type is mutated (a mutated proto-oncogene is called an oncogene), it can lead to cell growth that is out of control.
Mitosis, or cellular division, occurs in four stages: prophase, metaphase, anaphase, and telophase.
During prophase, the nuclear envelope disappears and chromosomes condense.
Next is metaphase, when chromosomes align at the metaphase plate and mitotic spindles attach to kinetochores.
In anaphase, chromosomes are pulled away from the center.
Telophase terminates mitosis, and the two new nuclei form.
The process of cytokinesis, which occurs during telophase, ends mitosis, as the cytoplasm and plasma membranes pinch to form two distinct, identical daughter cells.
Answers and explanations can be found in Chapter 15.
1. A scientist is testing new chemicals designed to stop the cell cycle at various stages of mitosis. Upon applying one of the chemicals, she notices that all of the cells appear as shown below. Which of the following best explains how the chemical is likely acting on the cells?

(A) The chemical has arrested the cells in prophase and has prevented attachment of the spindle fibers to the kinetochore.
(B) The chemical has arrested the cells in metaphase and has prevented dissociation of the spindle fibers from the centromere.
(C) The chemical has arrested the cells in metaphase and is preventing the shortening of the spindle fibers.
(D) The chemical has arrested the cells in anaphase and is preventing the formation of a cleavage furrow.
Questions 2 and 3 refer to the following graph and paragraph.

An experiment is performed to evaluate the amount of DNA present during a complete cell cycle. All of the cells were synced prior to the start of the experiment. During the experiment, a fluorescent chemical was applied to cells, which would fluoresce only when bound to DNA. The results of the experiment are shown above. Differences in cell appearance by microscopy or changes in detected DNA were determined to be phases of the cell cycle and are labeled with the letters A–D.
2. Approximately how long does S phase take to occur in these cells?
(A) 15 min
(B) 20 min
(C) 30 min
(D) 40 min
3. During which of the labeled phases of the experiment would the cell undergo anaphase?
(A) Phase A
(B) Phase B
(C) Phase C
(D) Phase D
4. Trisomy 21, which results in Down syndrome, results from nondisjunction of chromosome 21 in humans. Nondisjunction occurs when two homologous chromosomes, or two sister chromatids, do not separate. Which of the following describes the mechanism of this defect?
(A) During DNA replication in S phase of the cell cycle, the two new strands do not separate.
(B) During mitosis, at the metaphase plate, non-sister chromatids do not separate.
(C) The mitotic spindle attaches to chiasmata rather than kinetochores.
(D) The same microtubule in the spindle attaches to both sister chromatids during meiosis II.
5. A researcher isolates DNA from different types of cells and determines the amount of DNA for each type of cell. The samples may contain cells in various stages of the cell cycle. In order of increasing DNA content, which of the following would have the least amount of DNA to the greatest amount of DNA?
(A) Sperm, neuron, liver
(B) Ovum, muscle, taste bud
(C) Liver, heart, intestine
(D) Skin, neuron, ovum
6. Which of the following mutations would be least likely to have any discernable phenotype on the individual?
(A) Translocation of the last 1,000 base pairs of chromosome 1 onto chromosome 2
(B) Nondisjunction of chromosome 7 to produce trisomy-7
(C) Nondisjunction of chromosome 8 to produce monosomy-8
(D) Deletion of 400 base pairs, resulting in the loss of an enhancer
Respond to the following questions:
Which topics from this chapter do you feel you have mastered?
Which content topics from this chapter do you feel you need to study more before you can answer multiple-choice questions correctly?
Which content topics from this chapter do you feel you need to study more before you can effectively compose a free response?
Was there any content that you need to ask your teacher or another person about?