Every organism has a certain number of unique chromosomes. For example, fruit flies have 4 chromosomes, humans have 23 chromosomes, and dogs have 39 chromosomes. It turns out that most eukaryotic cells in fact have two full sets of chromosomes—one set from each parent. Humans, for example, have two sets of 23 chromosomes, giving us our grand total of 46.
A cell that has two sets of chromosomes is a diploid cell, and the zygotic chromosome number is given as “2n.” That means we have two copies of each chromosome.
If a cell has only one set of chromosomes, we call it a haploid cell. This kind of cell is given the symbol n. For example, we would say that the haploid number of chromosomes for humans is 23.
Remember:
Diploid refers to any cell that has two sets of chromosomes.
Haploid refers to any cell that has one set of chromosomes.
Why do we need to know the terms haploid and diploid? Because they are extremely important when it comes to sexual reproduction. As we’ve seen, 46 is the normal diploid number for human beings, but there are only 23 different chromosomes. The duplicate versions of each chromosome are called homologous chromosomes. The homologous chromosomes that make up each pair are similar in size and shape and contain the same genes in the same locations (although they might have different versions of those genes). This is the case in all sexually reproducing organisms. In fact, this is the essence of sexual reproduction: each parent donates half its chromosomes to its offspring.
Although most cells in the human body are diploid, there are special cells that are haploid. These haploid cells are called sex cells, or gametes. Why do we have haploid cells?
As we’ve said, an offspring has one set of chromosomes from each of its parents. A parent, therefore, contributes a gamete with one set that will be paired with the set from the other parent to produce a new diploid cell, or zygote. This produces offspring that are a combination of their parents.
What is genetics? In its simplest form, genetics is the study of heredity. It explains how certain characteristics are passed on from parents to children. Much of what we know about genetics was discovered by the monk Gregor Mendel in the 19th century. Since then, the field of genetics has vastly expanded.
Let’s begin then with some of the fundamental points of genetics:
Traits—or expressed characteristics—are influenced by one or more of your genes. Remember, a gene is simply a chunk of DNA that codes for a particular recipe. Different recipes affect different traits. DNA is passed from generation to generation, and from this process the genes and the traits associated with those genes are inherited. Within a chromosome, there are many genes, and each can contribute to a different trait. For example, in pea plants, there’s a gene on a chromosome that codes for seed coat characteristics. The position of a gene on a chromosome is called a locus.
Diploid organisms (organisms that have two sets of chromosomes) usually have two copies of each gene, one on each homologous chromosome. Homologous chromosomes are two copies or versions of the same chromosome in a diploid cell or organism. Humans have 23 pairs of homologous chromosomes.
The two gene copies may be different from one another—that is, they may be alternate versions, or alleles, of the same gene. For example, if we’re talking about the height of a pea plant, there’s an allele for tall and an allele for short. In other words, both alleles are alternate forms of the gene for height.
When an organism has two identical alleles for a given trait, the organism is homozygous. If an organism has two different alleles for a given trait, the organism is heterozygous.
When discussing the physical appearance of an organism, we refer to its phenotype. The phenotype tells us what the organism looks like. When talking about the genetic makeup of an organism, we refer to its genotype. The genotype tells us which alleles the organism possesses.
An allele can be dominant or recessive. This is determined by which allele “wins out” over the other in a heterozygote. The convention is to assign one of two letters for the two different alleles. The dominant allele receives a capital letter and the recessive allele receives a lowercase of the same letter. For instance, we might give the dominant allele for height in pea plants a T for tall. This means that the recessive allele would be t for short.
|
Name |
Genotype |
Phenotype |
|
homozygous dominant |
TT |
Tall |
|
homozygous recessive |
tt |
Short |
|
heterozygous |
Tt |
Tall |
One of the major ways the testing board likes to test genetics is by having you do crosses. Crosses involve mating hypothetical organisms with specific phenotypes and genotypes. We’ll look at some examples in a moment, but for now, keep these test-taking tips in mind:
Label each generation in the cross. The first generation in an experiment is always called the parent, or P generation. The offspring of the P generation are called the first filial, or F1 generation. Members of the next generation, the grandchildren, are called the F2 generation.
Always write down the symbols you’re using for alleles, along with a key to remind yourself what the symbols refer to. Use uppercase for dominant alleles and lowercase for recessive alleles.
Now let’s look at some basic genetic principles.
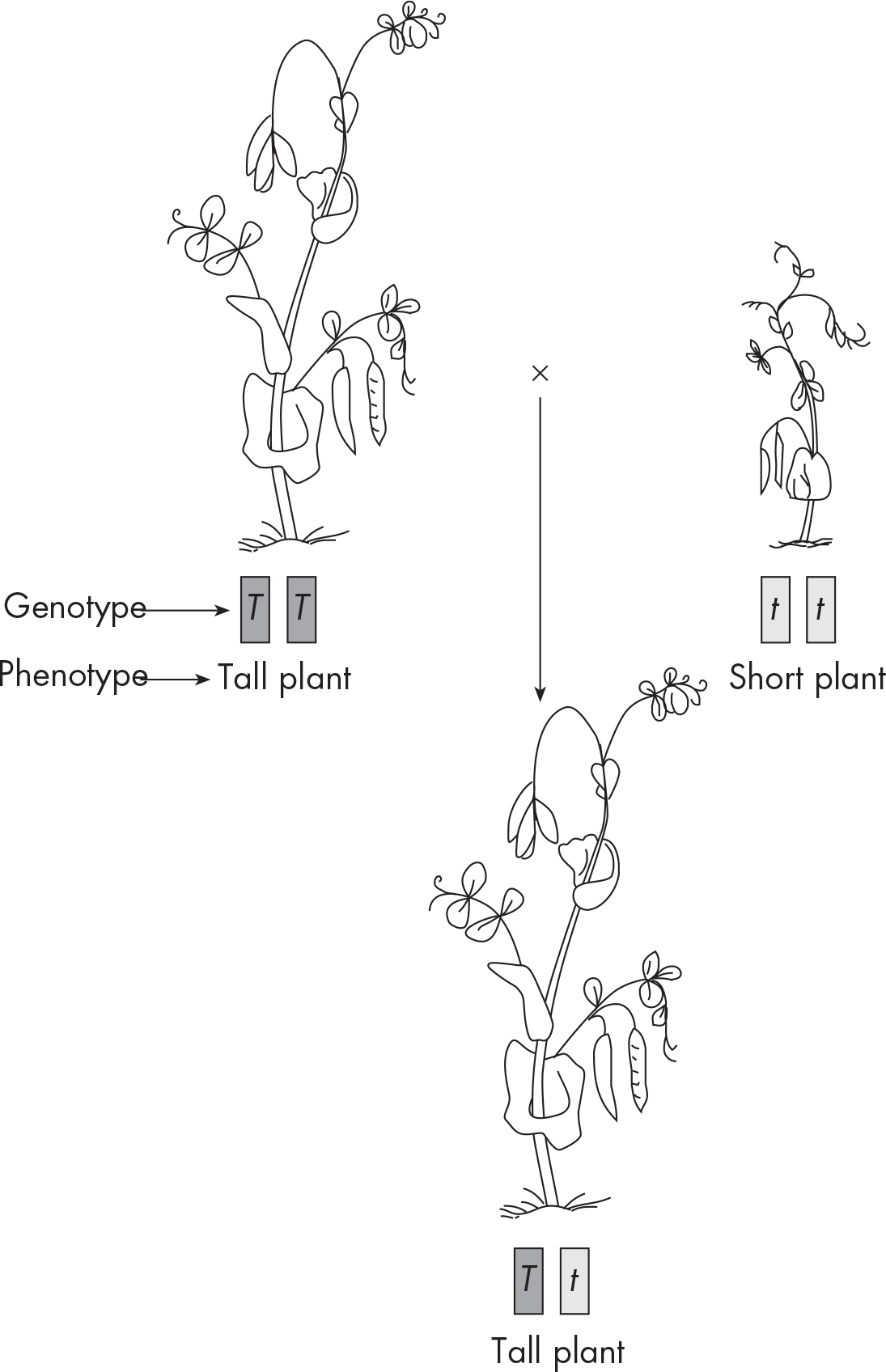
One of Mendel’s hobbies was to study the effects of cross-breeding on different strains of pea plants. Mendel worked exclusively with true-breeder pea plants. This means the plants he used were genetically pure and consistently produced the same traits. For example, tall plants always produced tall plants; short plants always produced short plants. Through his work, he came up with three principles of genetics: the Law of Dominance, the Law of Segregation, and the Law of Independent Assortment.
Mendel crossed two true-breeding plants with contrasting traits: tall pea plants and short pea plants.
To his surprise, when Mendel mated these plants, the characteristics didn’t blend to produce plants of average height. Instead, all the offspring were tall.
Mendel recognized that one trait must be masking the effect of the other trait. This is called the Law of Dominance. The dominant tall allele, T, somehow masked the presence of the recessive short allele, t. Consequently, all a plant needs is one tall allele to make it tall.
A monohybrid cross occurs when two individuals are crossed and one gene is being studied. A simple way to represent a monohybrid cross is to set up a Punnett square. Punnett squares are used to predict the results of a cross. Let’s construct a Punnett square for the cross between Mendel’s true-breeding tall and short pea plants. Let’s first designate the alleles for each plant. As we saw earlier, we can use the letter T for the tall dominant allele and t for the recessive short allele.
Since one parent was a pure, tall pea plant, we’ll give it two dominant alleles (TT homozygous dominant). The other parent was a pure, short pea plant, so we’ll give it two recessive alleles (tt homozygous recessive). To look at the types of offspring that these parents could produce, we have to figure out the gametes that each parent can possibly make (and thus contribute to the offspring).
Each parent has two copies of the height gene and each gamete gets one copy. This means each parent can make two types of gametes. For the homozygous dominant parent, the two copies are T and T. In other words, all gametes from this parent will contain the tall, or T, allele. The homozygous recessive plant can make gametes with t and with t. Notice again that because the parent plant was homozygous, only one type of allele (t, or short) will be passed on to the next generation. We will put the possible gametes for each parent on the top and left side of the square as shown below.
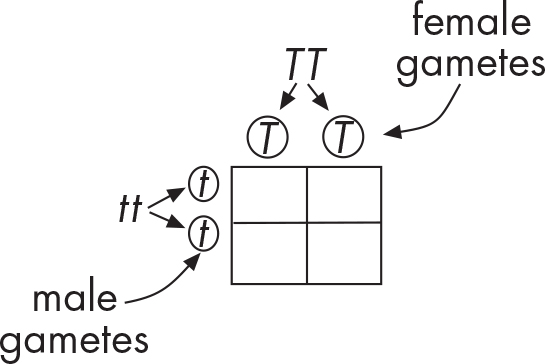
Now we can fill in the four boxes by matching the letters. What are the results for the F1 generation?
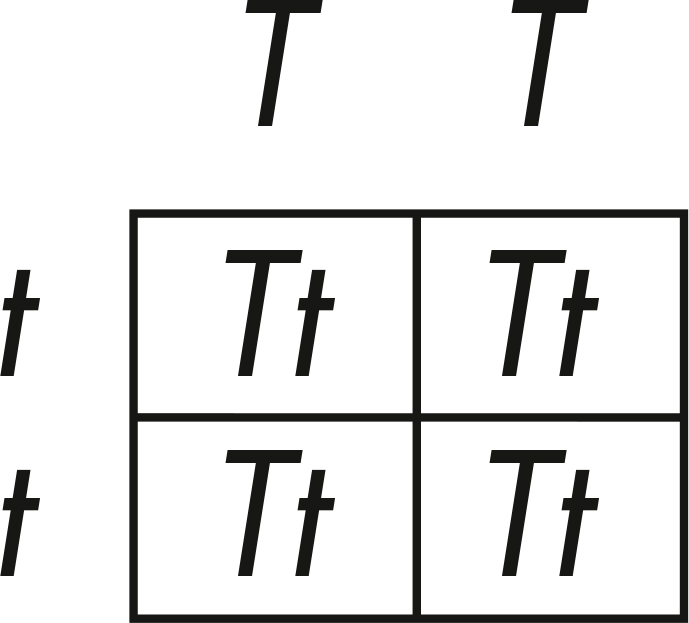
Each offspring received one allele from each parent. They all received one T and one t. They’re all Tt! So, even though the parents were homozygous, offspring are heterozygous: they possess one copy of each allele.
Next, Mendel took the offspring and self-pollinated them. Let’s use a Punnett square to spell out the results. This time we’re mating the offspring of the first generation—F1; we are crossing those heterozygotes. Take a look at the results:
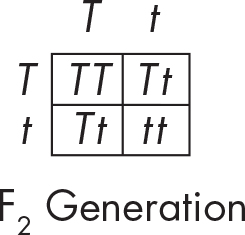
Check out the lower right box. One of the offspring could be a short pea plant!
Although all of the F1 plants were tall, they were heterozygotes that could make gametes containing the short t allele. In other words, their T and t alleles were segregated (separated) during gamete formation. This is an example of the Law of Segregation. It just means that each gamete gets only one of two copies of a gene. Gametes are haploid.
What about the genotype and phenotype for this cross? Remember, genotype refers to the genetic makeup of an organism, whereas phenotype refers to the appearance of the organism. According to the results of our Punnett square, what is the ratio of phenotypes and genotypes in the offspring?
Let’s sum up the results. We have four offspring with two different phenotypes: three of the offspring are tall, whereas one of them is short. On the other hand, we have three genotypes: 1 TT, 2 Tt, and 1 tt.
Here’s a summary of the results:
The ratio of phenotypes is 3:1 (three tall:one short).
The ratio of genotypes is 1:2:1 (one TT:two Tt:one tt).
So far, we have looked at only one trait: tall versus short. What happens when we study two traits at the same time? Each allele of the two traits will get segregated into two gametes, but how one trait gets split up into gametes has no bearing on how the other trait gets split up. In other words, the two alleles for each trait are sorted independently of the two alleles for other traits. For example, let’s look at two traits in pea plants: height and color. When it comes to height, a pea plant can be either tall (T) or short (t). As for color, the plant can be either green (G) or yellow (g), with green being dominant. Each gene comes in two alleles, so this gives us four alleles total. By the Law of Independent Assortment, these four alleles can combine to give us four different gametes.
TG Tg tG tg
In other words, each allele of the height gene can be paired with either allele of the color gene. The reason alleles segregate independently is because chromosomes segregate independently. During meiosis I, each pair of homologous chromosomes is split, and which one aligns left or right during metaphase I is different for each pair.
Now that we know that different genes assort independently into gametes, let’s look at a cross of two traits using those four gametes that we created above. A dihybrid cross is just like the monohybrid, but studies how two genes passed on to offspring
Here is the Punnett Square for a cross between two double heterozygotes (Tt Gg).

This is an example of the Law of Independent Assortment. Each of the traits segregated independently. Don’t worry about the different combinations in the cross—you’ll make yourself dizzy with all those letters. Simply memorize the phenotype ratio of the pea plants. For the 16 offspring there are:
9 tall and green
3 tall and yellow
3 short and green
1 short and yellow
That’s 9:3:3:1, but don’t forget the circumstances to which this applies. It is only when two heterozygotes for two genes are crossed. It is not just a magic ratio that always works.
The Punnett square method works well for monohybrid crosses and helps us visualize possible combinations. However, a better method for predicting the likelihood of certain results from a dihybrid cross is to apply the Rules of Probability. To determine the probability that two or more independent events will occur simultaneously, one can simply calculate the product of the probability that each will occur independently. This is called the Product Rule. To determine the likelihood that EITHER event occurs, but not both, use another rule called the Sum Rule. To illustrate the product rule, let’s consider again the cross between two identical dihybrid tall, green plants with the genotype TtGg.
Product Rule: If A and B are independent, then: P(A and B) = P(A) times P(B)
Sum Rule: If A and B are mutually exclusive, then: P(A or B) = P(A) + P(B)
To find the probability of having a tall, yellow plant, simply multiply the probabilities of each event. If the probability of being tall is  and the probability of being yellow is
and the probability of being yellow is  , then the probability of being tall and yellow is
, then the probability of being tall and yellow is 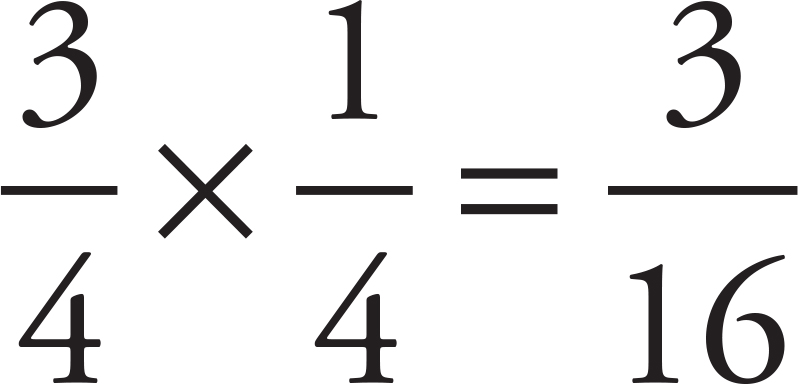 .
.
Let’s summarize Mendel’s three laws.
|
SUMMARY OF MENDEL’S LAWS |
|
|
Laws |
Definition |
|
Law of Dominance |
One trait masks the effects of another trait. |
|
Law of Segregation |
Each gamete gets only one of the copies of each gene. |
|
Law of Independent Assortment |
Each pair of homologous chromosomes splits independently, so the alleles of different genes can mix and match. |
Suppose we want to know if a tall plant is homozygous (TT) or heterozygous (Tt). Its physical appearance doesn’t necessarily tell us about its genetic makeup. The only way to determine its genotype is to cross the plant with a recessive, short plant, tt. This is known as a test cross. When the recessive plant is used, there will be only two possible outcomes in the daughter plants. Our mystery tall plant could be TT or Tt so when we mate it to a short plant, these are the possible situations: (1) TT × tt or (2) Tt × tt. Let’s take a look.
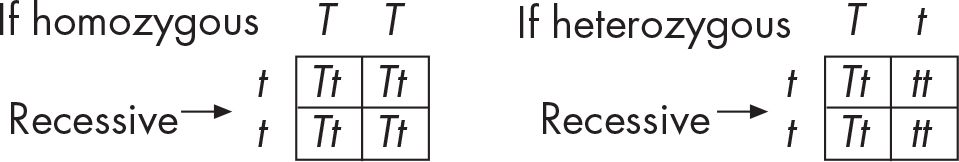
If none of the offspring is short, our original plant must have been homozygous, TT. If, however, even one short plant appears in the bunch, we know that our original pea plant was heterozygous, Tt. In other words, it wasn’t a pure-breeding plant. Overall then, a test cross uses a recessive organism to determine the genotype of an organism of unknown genotype.
Sometimes genes on the same chromosome stay together during assortment and move as a group. The group of genes is considered linked and tends to be inherited together. For example, the genes for flower color and pollen shape are linked on the same chromosomes and show up together.
Since linked genes are found on the same chromosome, they cannot segregate independently. This violates the Law of Independent Assortment.
Let’s pretend that the height and color genes we looked at in the dihybrid cross are linked. A heterozygote for both traits still has two alleles for height (T and t) and two alleles for color (G and g). However, because height and color are located on the same chromosome, one allele for height and one allele for color are physically linked. For example, maybe the heterozygote has one chromsome with Tg and one chromosome with tG. When gametes are formed, the T and g will travel together, and t and G will travel together and be packaged into a gamete together. So, in the unlinked dihybrid shown earlier, there were four possible gamete combinations (TG, Tg, tG, tg), but now there are only two (Tg and tG). The only way to physically separate linked alleles is by crossing-over. If a crossover event occurs between linked genes, then recombinant gametes can occur. However, because linked genes are physically close together, crossing-over is not likely to occur between them.
If the genes were unlinked, then four gametes (TG, Tg, tG, tg) would be equally likely (as we saw when we talked about independent assortment). However, if certain combinations of alleles are found more often in offspring than they should (based on Mendelian ratios), this is a red flag that the two genes are close together and linked.
Look at the example below from a cross of TtGg and ttgg. If you draw out the Punnett square, you should get equal numbers of each phenotype occurring if the genes are assumed to be unlinked. Yet, what if the numbers below were the actual numbers of offspring of each phenotype?
Tall and Green: 6 offspring
short and Green: 37 offspring
Tall and yellow: 45 offspring
short and yellow: 3 offspring
There are more Green/short and yellow/Tall offspring. This is a sign that these alleles are linked on a chromosome because they occur more often than they statistically should. This shows that most of the gametes of the TtGg parent had tG together and Tg together since those gamete combinations would result in Green/short and yellow/Tall offspring.
So, where did the Green/Tall and yellow/short plants come from? They are the result of gametes from the TtGg parent that are TG and tg, respectively. If the Tg and tG alleles were linked in that parent, how could they produce a few TG and tg gametes? The answer is through recombination! Even though those alleles were not found on the same chromosome, recombination must have occurred during meiosis to swap them. This allowed gametes to be formed with TG and tg, and offspring to occur that were Tall/Green and short/yellow. The offspring formed from reccombination events are called recombinants. The percentage of recombination (recombination frequency) can be determined by adding up the recombinants and dividing by the total number of offspring.
9/91 = 9.9%
This percentage can also be used as a measure of how far apart the genes are. The distance on a chromosome is measured in map units, or centimorgans.
The frequency of crossing-over between any two linked alleles is proportional to the distance between them. That is, the farther apart two linked alleles are on a chromosome, the more often the chromosome will cross over between them. This finding led to recombination mapping—mapping of linkage groups with each map unit being equal to 1 percent recombination. For example, if two linked genes, A and B, recombine with a frequency of 15 percent, and B and C recombine with a frequency of 9 percent, and A and C recombine with a frequency of 24 percent, what is the sequence and the distance between them?
The sequence and the distance of A-B-C is

If the recombination frequency between A and C had been 6 percent instead of 24 percent, the sequence and distance of A-B-C would instead be

We already know that humans contain 23 pairs of chromosomes. Twenty-two of the pairs of chromosomes are called autosomes. They code for many different traits. The other pair contains the sex chromosomes. This pair determines the sex of an individual. A female has two X chromosomes. A male has one X and one Y chromosome—an X from his mother and a Y from his father. Some traits, such as color blindness and hemophilia, are carried on sex chromosomes. These are called sex-linked traits. Most sex-linked traits are found on the X chromosome and are more properly referred to as “X-linked.”
Since males have one X and one Y chromosome, what happens if a male has an X-chromosome with the color blindness allele? Unfortunately, he’ll express the sex-linked trait, even if it is recessive. Why? Because his one and only X chromosome is color blind-X. He doesn’t have another X to mask the effect of the color blind-X. However, if a female has only one color blind-X chromosome, she won’t express a recessive sex-linked trait. For her to express the trait, she has to inherit two color blind-X chromosomes. A female with one color blind-X is called a carrier. Although she does not exhibit the trait, she can still pass it on to her children.
You can also use the Punnett square to figure out the results of sex-linked traits. Here’s a classic example: a male who has normal color vision and a woman who is a carrier for color blindness have children. How many of the children will be color blind? To figure out the answer, let’s set up a Punnett square.
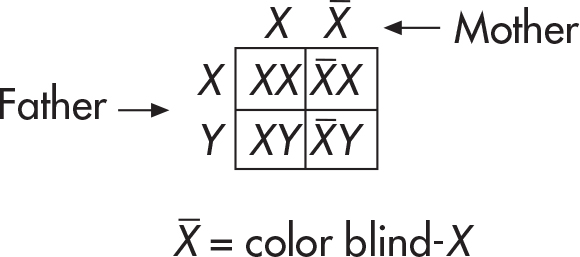
Notice that we placed a bar above any color blind-X to indicate the presence of a color blindness allele. And now for the results. The couple could have a son who is color-blind, a son who sees color normally, a daughter who is a carrier, or a daughter who sees color normally.
There are a few X-linked conditions you should be familiar with for test day. Some are caused by a dominant allele, and others are inherited by a recessive allele.
|
Examples of X-Linked Dominant Traits |
|
|
Trait |
Phenotype |
|
Rett Syndrome |
Postnatal neurological disorder of the grey matter of the brain |
|
Vitamin D Resistant Rickets |
Bone deformity including short stature and bow leggedness |
|
Fragile X Syndrome |
Developmental problems including learning disabilities and cognitive impairment |
|
Examples of X-Linked Recessive Traits |
|
|
Trait |
Phenotype |
|
Red/Green Color Blindness |
Difficulty discriminating red and green hues |
|
Hemophilia |
Impaired blood clotting, bruising easily |
|
Duchenne Muscular Dystrophy |
Muscle weakness and loss |
|
Hunter Syndrome |
Decreased ability to break down and recycle mucopolysaccharides |
|
Juvenile Gout |
Uric acid build-up, swollen joints, kidney problems |
|
Hereditary Nephritis |
Kidney disease and hearing loss |
A look at the cell nucleus of normal females will reveal a dark-staining body known as a Barr body. A Barr body is an X chromosome that is condensed and visible. In every female cell, one X chromosome is activated and the other X chromosome is deactivated during embryonic development. Surprisingly, the X chromosome destined to be inactivated is randomly chosen in each cell. Therefore, in every tissue in the adult female, one X chromosome remains condensed and inactive. However, this X chromosome is replicated and passed on to a daughter cell. X-inactivation is the reason it is okay that females have two X chromosomes and males have only one. After X-inactivation, it is like everyone has one copy.
Not all patterns of inheritance obey the principles of Mendelian genetics. In fact, many traits we observe are due to a combined expression of alleles. Here are a couple of examples of non-Mendelian forms of inheritance:
Incomplete dominance (blending inheritance): In some cases, the traits will blend. For example, if you cross a white snapdragon plant (genotype WW) with a red snapdragon plant (RR), the resulting progeny will be pink (RW). In other words, neither color is dominant over the other.
Codominance: Sometimes you’ll see an equal expression of both alleles. For example, an individual can have an AB blood type. In this case, each allele is equally expressed. That is, both the A allele and the B allele are expressed (IAIB). That’s why the person is said to have AB blood. The expression of one allele does not prevent the expression of the other.
Polygenic inheritance: In some cases, a trait results from the interaction of many genes. Each gene will have a small effect on a particular trait. Height, skin color, and weight are all examples of polygenic traits.
Non-nuclear inheritance: Apart from the genetic material held in the nucleus, there is also genetic material in the mitochondria. The mitochondria are always provided by the egg during sexual reproduction, so mitochondrial inheritance is always through the maternal line, not the male line. In plants the mitochondria are provided by the ovule and are maternally inherited. Chloroplasts also contain DNA, in the form of one circular double-stranded molecule. Both chloroplasts and mitochondria assort randomly into daughter cells and gametes.
One way to study genetic inheritance is by looking at a special family tree called a pedigree. A pedigree shows which family members have a particular trait and it can help determine if a trait is recessive or dominant and if it is sex-linked. Traits that skip generations are usually recessive. Traits that appear more in one sex than the other are usually sex-linked. In a pedigree chart, the males are squares and the females are circles. Matings are drawn with horizontal lines and the offspring are shown below the mated couple.
Changes in genotypes can result in changes in phenotype, but environmental factors also influence many traits, both directly and indirectly. In addition, an organism’s adaptation to the local environment reflects a flexible response of its genome. For example, gene expression and protein function can be affected by temperature. Phenotypic plasticity occurs if two individuals with the same genotype have different phenotypes because they are in different environments. Here are some examples of how the environment can influence traits both directly and indirectly:
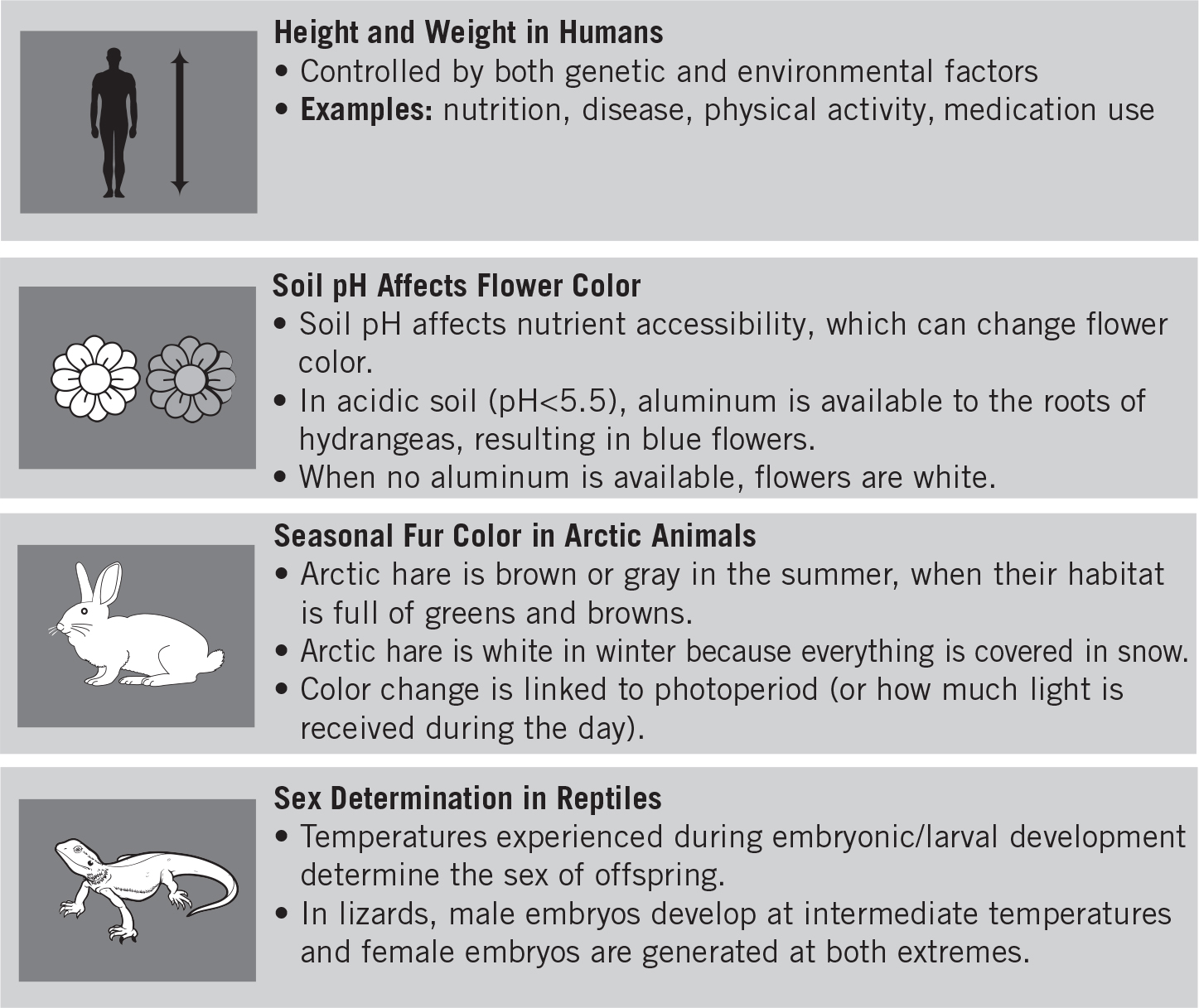
Meiosis is the production of gametes. Since sexually reproducing organisms need only haploid cells for reproduction, meiosis is limited to sex cells in special sex organs called gonads. In males, the gonads are the testes, while in females they are the ovaries. The special cells in these organs—also known as germ cells—produce haploid cells (n), which then combine to restore the diploid (2n) number during fertilization.
female gamete (n) + male gamete (n) = zygote (2n)
When it comes to genetic variation, meiosis is a big plus. Variation, in fact, is the driving force of evolution. The more variation there is in a population, the more likely it is that some members of the population will survive extreme changes in the environment. Meiosis is far more likely to produce these sorts of variations than is mitosis, which therefore confers selective advantage on sexually reproducing organisms. We’ll come back to this theme in Chapter 10.
Meiosis actually involves two rounds of cell division: meiosis I and meiosis II.
Before meiosis begins, the diploid cell goes through interphase. Just as in mitosis, double-stranded chromosomes are formed during S phase.
Meiosis I consists of four stages: prophase I, metaphase I, anaphase I, and telophase I.
Prophase I
Prophase I is a little more complicated than regular prophase. As in mitosis, the nuclear membrane disappears, the chromosomes become visible, and the centrioles move to opposite poles of the nucleus. But that’s where the similarity ends.
The major difference involves the movement of the chromosomes. In meiosis, the chromosomes line up side-by-side with their counterparts (homologs). This event is known as synapsis.
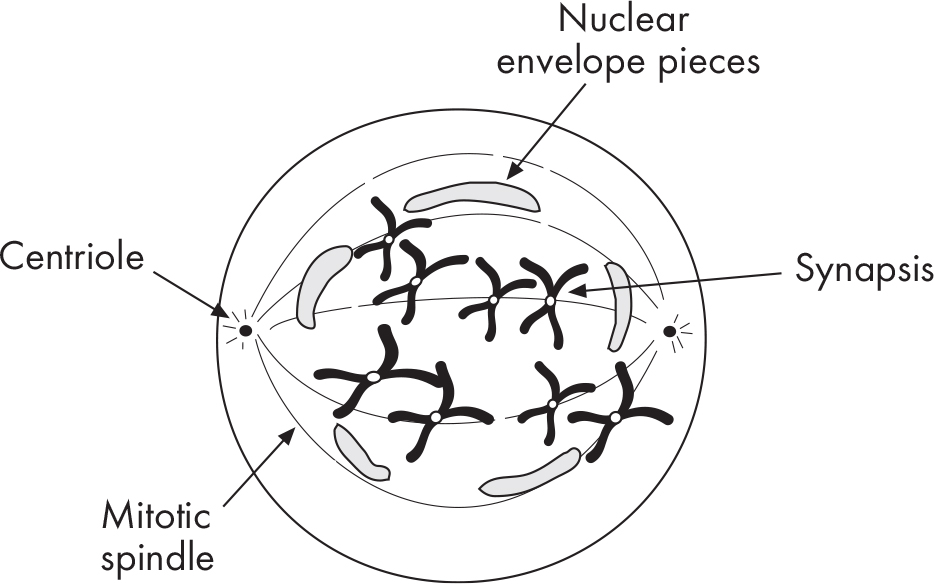
Synapsis involves two sets of chromosomes that come together to form a tetrad (a bivalent). A tetrad consists of four chromatids. Synapsis is followed by crossing-over, the exchange of segments between homologous chromosomes.
What’s unique in prophase I is that pieces of chromosomes are exchanged between homologous partners. This is one of the ways organisms produce genetic variation. By the end of prophase I, chromosomes will have exchanged regions containing several alleles, or different forms of the same gene. This means that each chromosome will have a different mix of alleles than it began with. Remember, the two homologous chromosomes are versions that came from the person’s mother and father. So, a chromosome that has experienced crossing-over will now be a mix of alleles from the mother and father versions.
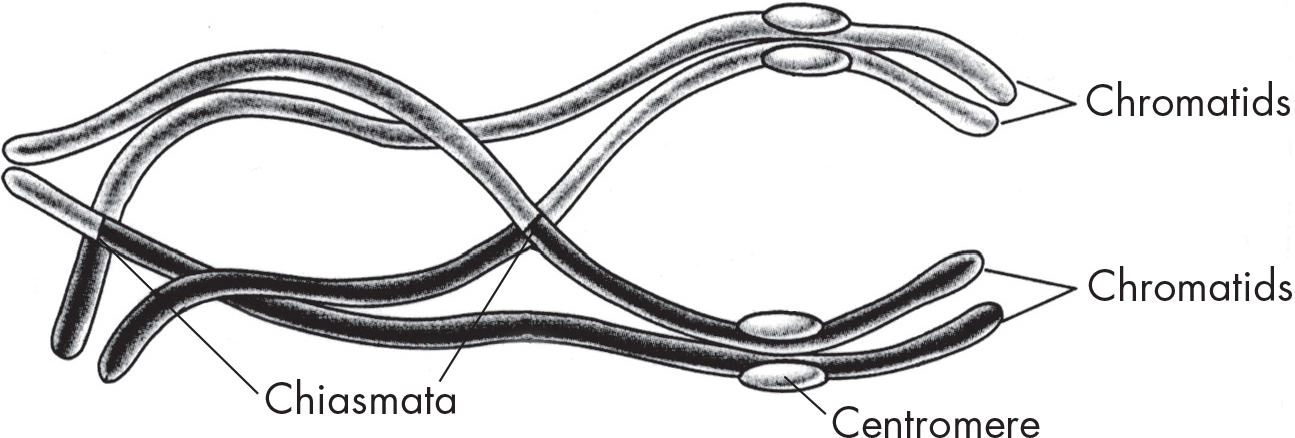
Metaphase I
As in mitosis, the chromosome pairs—now called tetrads—line up at the metaphase plate. By contrast, you’ll recall that in regular metaphase, the chromosomes line up individually. One important concept to note is that the alignment during metaphase is random, so the copy of each chromosome that ends up in a daughter cell is random. Therefore, the gamete will be created with a mixture of genetic information from the person’s father and mother. This means that the offspring created would be a combination of all four grandparents.
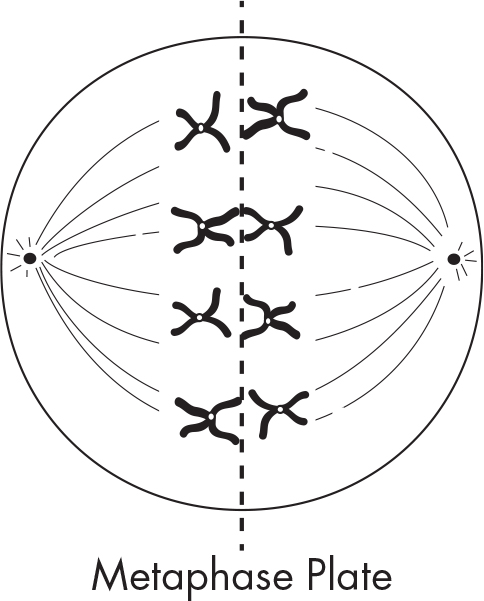
Anaphase I
During anaphase I, each pair of chromatids within a tetrad moves to opposite poles. Notice that the chromatids do not separate at the centromere. The homologs separate with their centromeres intact.
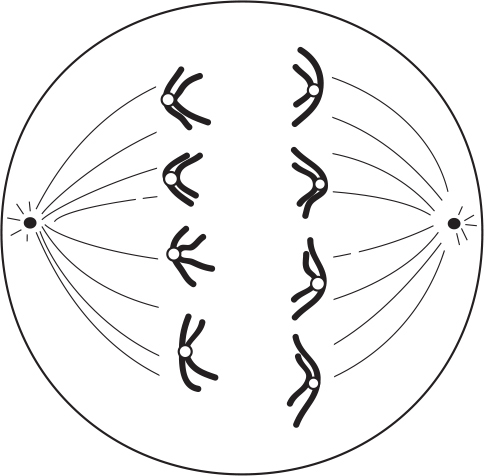
The chromosomes now move to their respective poles.
Telophase I
During telophase I, the nuclear membrane forms around each set of chromosomes.
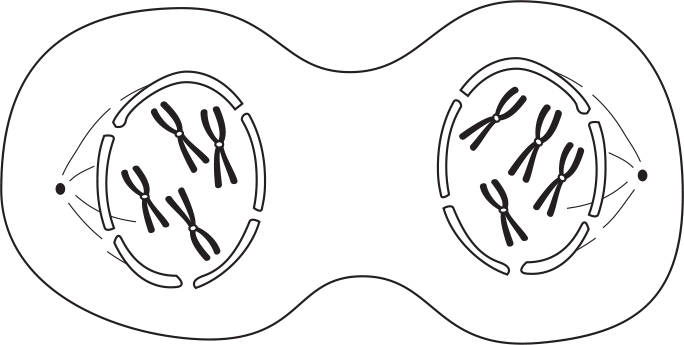
Finally, the cells undergo cytokinesis, leaving us with two daughter cells. Notice that at this point the nucleus contains the haploid number of chromosomes, but each chromosome is a duplicated chromosome, which consists of two chromatids.
The purpose of the second meiotic division is to separate sister chromatids, and the process is virtually identical to mitosis. Let’s run through the steps of meiosis II.
During prophase II, chromosomes once again condense and become visible. In metaphase II, chromosomes move toward the metaphase plate. This time they line up single file, not as pairs. During anaphase II, chromatids of each chromosome split at the centromere, and each chromatid is pulled to opposite ends of the cell. At telophase II, a nuclear membrane forms around each set of chromosomes and a total of four haploid cells are produced.
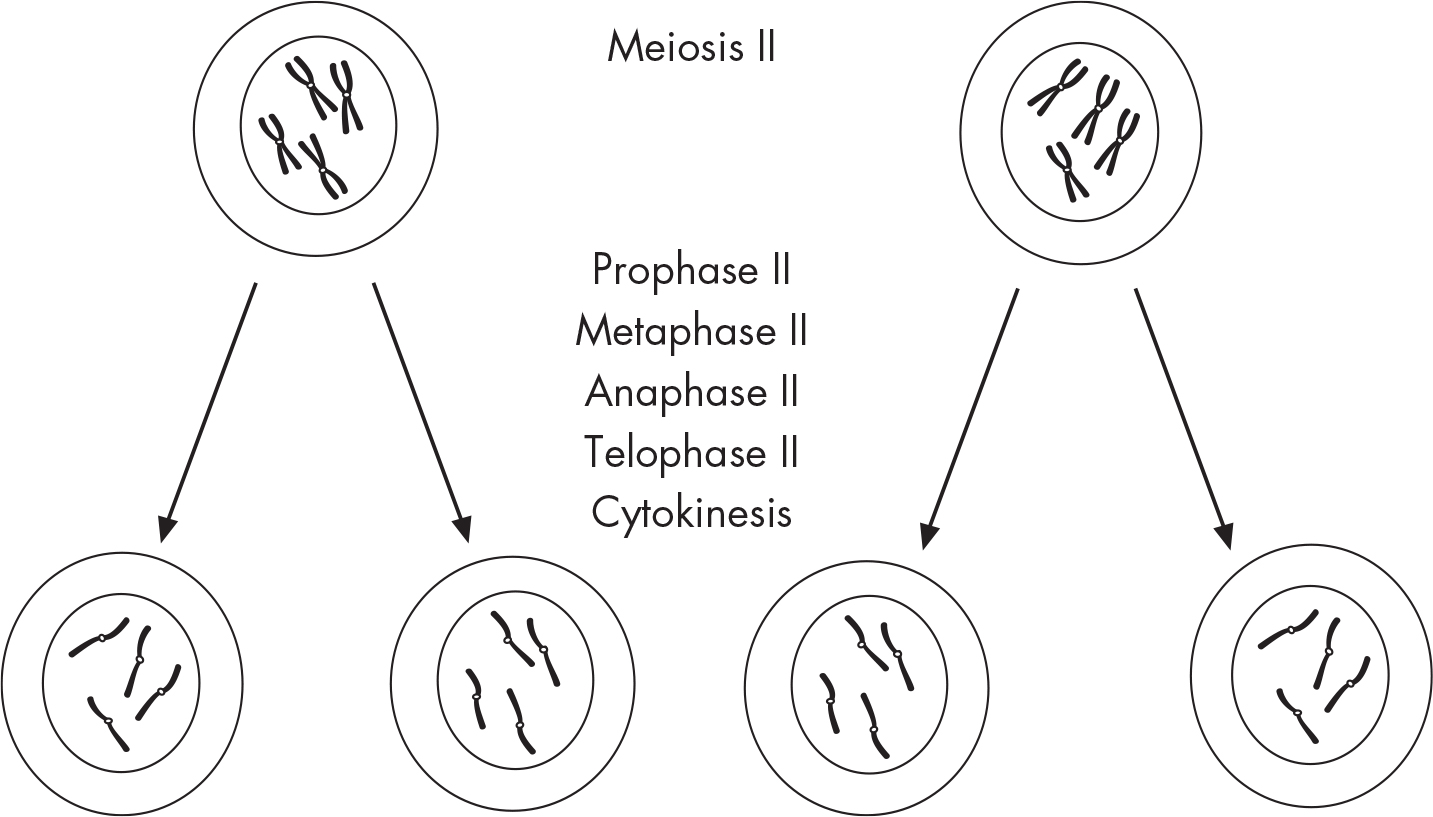
Meiosis is also known as gametogenesis. If sperm cells are produced, then meiosis is called spermatogenesis. During spermatogenesis, four sperm cells are produced for each diploid cell. If an egg cell or an ovum is produced, this process is called oogenesis.
Oogenesis is a little different from spermatogenesis. Oogenesis produces only one ovum, not four. The other three cells, called polar bodies, get only a tiny amount of cytoplasm and eventually degenerate. Why does oogenesis produce only one ovum? Because the female wants to conserve as much cytoplasm as possible for the surviving gamete, the ovum.
Here’s a summary of the major differences between mitosis and meiosis.
|
MITOSIS |
MEIOSIS |
|
|
Sometimes, a set of chromosomes has an extra or a missing chromosome. This occurs because of nondisjunction—chromosomes failed to separate properly during meiosis. This error, which produces the wrong number of chromosomes in a cell, usually results in miscarriage or significant genetic defects. For example, humans typically have two copies of each chromosome, but individuals with Down syndrome have three—instead of two—copies of the 21st chromosome.
Nondisjunction can occur in anaphase I (meaning chromosomes don’t separate when they should), or in anaphase II (meaning chromatids don’t separate). Either one can lead to aneuploidy, or the presence of an abnormal number of chromosomes in a cell.
Chromosomal abnormalities also occur if one or more segments of a chromosome break and are either lost or reattach to another chromosome. The most common example is translocation, which occurs when a segment of a chromosome moves to another nonhomologous chromosome.
Here’s an example of a translocation.
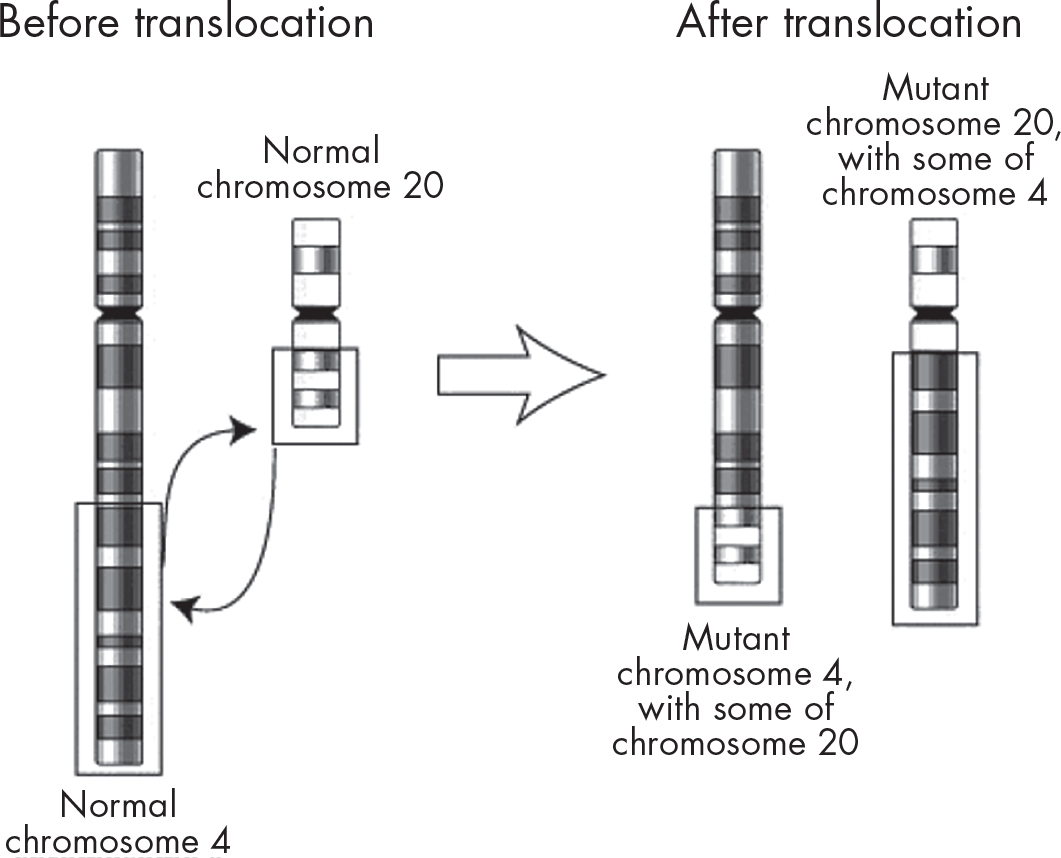
Translocations can occur when recombination (crossing-over) occurs incorrectly. Fortunately, in most cases, damaged DNA can usually be repaired by special repair enzymes.
Many ethical, social, and medical issues surround human genetic disorders. Individuals who are carriers for genetic diseases are at increased risk of having a child with a genetic disease. A prenatal diagnosis can identify some genetic diseases before birth, and parents can then prepare to have a child with special needs or have the option of terminating the pregnancy. In addition, individuals who carry genetic diseases sometimes use an egg or sperm donor instead of using their own gametes.
Civic issues are also important to consider. Genetic discrimination occurs when people are treated unfairly because of actual or perceived differences in their genetic information that may cause or increase the risk of developing a disorder or disease. For example, a health insurer might refuse to give coverage to an individual who has a genetic difference that raises their odds of getting cancer. Many G7 countries protect genetic test information to help eliminate genetic discrimination and safeguard individuals’ genetic privacy.
diploid cell
haploid cell
homologous chromosomes
sex cells (or gametes)
Gregor Mendel
trait
genes
locus
alleles
homozygous
heterozygous
phenotype
genotype
dominant
recessive
parent, or P generation
filial, or F1 generation
F2 generation
Law of Dominance
Law of Segregation
Law of Independent Assortment
monohybrid cross
Punnett square
dihybrid cross
Product Rule
Sum Rule
test cross
linked genes
recombinants
percentage of recombination (recombination frequency)
map units (centimorgans)
autosomes
sex chromosomes
color blindness
hemophilia
sex-linked traits
carrier
Barr body
incomplete dominance (blending inheritance)
codominance
polygenetic inheritance
non-nuclear inheritance
pedigree
phenotypic plasticity
meiosis
gonads
testes
ovaries
germ cells
meiosis I
meiosis II
synapsis
tetrad (bivalent)
crossing-over (recombination)
alleles
gametogenesis
spermatogenesis
oogenesis
polar bodies
ovum
nondisjunction
Down syndrome
aneuploidy
translocation
Genetics is the study of heredity. The foundation of genetics is that traits are produced by genes, which are found on chromosomes.
Diploid organisms carry two copies of every gene, one on each of two homologous chromosomes, one that came from their mother and one that came from their father.
When two alleles are the same, the organism is homozygous for that trait. When the two alleles are different, the organism is heterozygous for the trait.
Genes are described as the genotype of the organism. The physical expression of the traits, or what you see in the organism, is the phenotype.
Homozygous individuals have only one allele that will determine the phenotype. However, heterozygous individuals have two different alleles that could contribute to the phenotype. The allele that “wins out” and directs the phenotype is the dominant allele. The one that doesn’t show up in the phenotype is the recessive allele.
Simple genetics with dominants and recessives was illuminated by Gregor Mendel. In Mendelian genetics, there are three important laws: the Law of Dominance, the Law of Segregation, and the Law of Independent Assortment.
If two genes are on the same chromosome and close together, they are linked. Completely linked alleles on the same chromosome are locked together and travel together during gamete formation. Recombination frequency can be used to measure how far apart two genes are on a chromosome.
Humans have 23 pairs of homologous chromosomes for a total of 46. Of these, 22 pairs are autosomes and 2 are designated as sex chromosomes:
Females have two X chromosomes; males have one X and one Y chromosome.
Because females have two X chromosomes, one is inactivated in each cell and condenses into a Barr body.
Non-Mendelian genetics refers to a situation in which traits do not follow the Mendelian laws. Examples include:
incomplete dominance
codominance
polygenic inheritance
Meiosis produces four genetically distinct haploid gametes (sperm or egg cells). Meiosis involves two rounds of cell division: meiosis I and meiosis II.
During meiosis I, the homologous chromosome pairs separate, but before they can, they undergo crossing-over and swap some DNA.
During meiosis II, sister chromatids separate.
Mutations, such as nondisjunction events or whole chromosome translocations, can lead to genetic diseases.
Answers and explanations can be found in Chapter 15.
1. Two expecting parents wish to determine all possible blood types of their unborn child. If both parents have an AB blood type, which of the following blood types will their child NOT possess?
(A) A
(B) B
(C) AB
(D) O
2. A new species of tulip was recently discovered. A population of pure red tulips was crossed with a population of pure blue tulips. The resulting F1 generation was all purple. This result is an example of which of the following?
(A) Complete dominance
(B) Incomplete dominance
(C) Codominance
(D) Linkage
3. In pea plants, tall (T) is dominant over short (t) and green (G) is dominant over yellow (g). If a pea plant that is heterozygous for both traits is crossed with a plant that is recessive for both traits, approximately what percentage of the progeny plants will be tall and yellow?
(A) 0%
(B) 25%
(C) 66%
(D) 75%
Questions 4–6 refer to the following pedigree tree and paragraph.
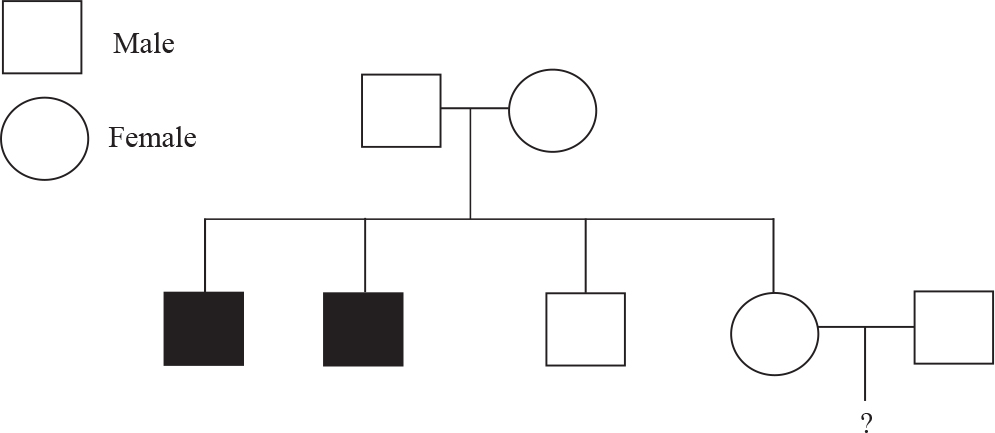
Hemophilia is an X-linked disease associated with the inability to produce specific proteins in the blood-clotting pathway. Shown above is a family pedigree tree in which family members afflicted with the disease are shown with filled-in squares (male) or circles (females). A couple is trying to determine the likelihood of passing on the disease to their future children (represented by the ? symbol above) because the hemophilia runs in the woman’s family.
4. Assuming that the woman in the couple is a carrier, what is the probability that the couple’s first son will have hemophilia?
(A) 0%
(B) 25%
(C) 50%
(D) 100%
5. Why were both women in the family tree above free of the disease?
(A) They were lucky because they didn’t receive an X chromosome with the diseased allele.
(B) They cannot get hemophilia because it is associated only with a diseased Y chromosome.
(C) They are carriers and will get the disease only if they have children.
(D) They have at least one normal (unaffected) copy of the X chromosome.
6. Turner syndrome is a disease in which an individual is born with only a single X chromosome. Suppose the woman in the couple is a carrier for hemophilia and has a child with Turner syndrome. Would this child have the disease?
(A) Yes, because the child would have only one copy of the X chromosome and it would be affected.
(B) No, because women cannot be affected by hemophilia.
(C) No, because the child would have to receive a normal copy of the X chromosome from its mother.
(D) Maybe, it depends on which X chromosome she receives from her mother.
7. Neuronal ceroid lipofuscinosis (NCL) is a group of autosomal recessive diseases characterized by blindness, loss of cognitive and motor function, and early death. One of the genes that is mutated in this disease is CLN3. When functional CLN3 protein is absent, neurons die because of increased storage material in the cells, presumably because the lysosomes aren’t working properly. What is a valid explanation as to why parents of children with this devastating disease can be completely normal?
(A) Parents of the autosomal recessive disease must both be carriers of the mutation on the CLN3 gene.
(B) Carriers of the CLN3 mutation genotype do not show the phenotype because one normal CLN3 allele is present to provide a functioning CLN3 protein.
(C) Redundant proteins take over the function of the mutant CLN3 protein in the parents.
(D) X inactivation prevents expression of the CLN3 mutated protein in the parents.
8. A breeder of black Labrador puppies notices that in his line a gene predisposing his dogs for white spots has arisen. He believes the white spot allele is autosomal recessive, and he wants to prevent it from continuing in his dogs. He sees that one of his female dogs, Speckle, has white spots, and he therefore assumes she is homozygous for the gene. But he must determine which males in his pack are heterozygous to avoid breeding them in the future. What is a reasonable plan?
(A) Stop breeding Speckle.
(B) Pair Speckle with a male with white spots.
(C) Perform a testcross of Speckle with a black Labrador male. If any pups are born with white spots, stop breeding that male because he is heterozygous.
(D) Breed Speckle with a chocolate Labrador.
9. Which of the following is a reason why certain traits do not follow Mendel’s Law of Independent Assortment?
(A) The genes are linked on the same chromosome.
(B) It applies only to eukaryotes.
(C) Certain traits are not completely dominant.
(D) Heterozygotes have both alleles.
10. The gene for coat color in some breeds of cats is found on the X chromosome. Calico cats are mottled with orange and black colorings. What is a possible explanation for the fact that true calico cats are only female?
(A) The allele for coat color is randomly chosen by X-inactivation.
(B) The coat color is linked with genes that are lethal to male cats, so male calico cats never live past birth.
(C) Coat orange and black color is expressed only in female cats; males have only one color.
(D) Male cats have Y chromosomes.
Respond to the following questions:
Which topics from this chapter do you feel you have mastered?
Which content topics from this chapter do you feel you need to study more before you can answer multiple-choice questions correctly?
Which content topics from this chapter do you feel you need to study more before you can effectively compose a free response?
Was there any content that you need to ask your teacher or another person about?