2
STARDUST SAURIANS
BEING BOTH FAMOUS AND EXTINCT, dinosaurs tend to be portrayed as poster children for failure. Yet they are one of Nature’s great success stories, having persisted for about 160 million years. (In contrast, primates have been around about 55 million years, our hominid cousins 7 million years, and humans a mere 200,000 years or so.) During that time, they evolved into a wondrous DIVERSITY of forms and occupied every continent, from pole to pole, becoming the dominant large-bodied land animals of the Mesozoic Era. Plus, they aren’t completely extinct; abundant evidence confirms that birds are the direct descendants of dinosaurs, and so, in a very real sense, are dinosaurs themselves. With about ten thousand living species (and thousands of extinct forms), birds are merely the latest chapter in the lengthy dinosaurian story. And finally, extinction is not the shameful exception but the ultimate fate of all species. Over 99 percent of species that have ever existed on Earth are now extinct. Evolution depends as much on the deaths of species as it does on their births. Disappearance of the old makes way for the new.
It takes a blend of hubris and myopia to regard dinosaurs as unsuccessful. We like to view ourselves as the destiny of evolution, with the previous thousands of millions of years of evolution relegated to a warm-up act. Yet such anthropocentric hubris would not be possible (or at least palatable) without a severe case of temporal myopia. Most of our day-to-day concerns, and those of our hominid forebears, span mere seconds, minutes, hours, and days. Our lifetimes are measured in decades, whereas the age of the planet spans billions (thousands of millions) of years! Like cosmic distances, Earth history is so vast as to be virtually beyond human conception, and the tenure of our species amounts to a minute fraction of 1 percent of this duration.
When fathoming the immensity of geologic time, it helps to turn to metaphors. For example, if Earth history is a 24-hour clock, humans show up a few seconds before midnight. If depicted as the length of a football field, we appear just as time covers the distance and breaks the plane of the goal line. If compared to the old measure of an English yard—the distance from the tip of the king’s nose to the tip of his outstretched hand—a single stroke of the royal nail file on his middle finger eliminates human history. My favorite comes from Mark Twain, who noted that if the height of the Eiffel Tower were to represent Earth history, the duration of humans would amount to the thickness of paint on the ball at the tower’s pinnacle. Mocking human hubris, Twain went on to add that, of course, anybody could perceive that the tower was created for that thin layer of paint!
Along with Copernicus’s sun-centered solar system and Darwin’s common ancestry of life, recognition of DEEPTIME ranks as one of the most important discoveries in all human thought, forever altering our view of ourselves and the universe. Most profoundly, deep time reduces the duration of human existence to a veritable blink.
Of the hundreds of millions of species that have lived on Earth, we are the first to contemplate the vastness of geologic time. But imagine a world in which there was no linear time—no sense of events incessantly shifting from past to present. What if, instead, we viewed life as a continual sequence of repeating cycles, with no true sense of history? As difficult as this concept may be to grasp, it represents reality for peoples throughout most of human history and continues to be the norm for many indigenous cultures. As expressed by cultural ecologist David Abram in The Spell of the Sensuous:
To indigenous, oral cultures, the ceaseless flux that we call “time” is overwhelmingly cyclical in character. The senses of oral people are attuned to the land around them, still conversant with the expressive speech of the winds and the forest birds, still participant with the sensuous cosmos. Time, in such a world, is not separable from the circular life of the sun and the moon, from the cycling of the seasons, the death and rebirth of animals— from the eternal return of the greening earth.1
For such preindustrial peoples, even unique and extraordinary events become fully assimilated into the concept of circular time and recurrent myths, morphing as the story passes through generations. Abram argues that the concept of linear time emerged alongside alphabetic writing, perhaps with the ancient Hebrews. Only with the written word can we permanently record details of historical or mythic events, creating a nonrepeating time line.
In the same way that scientists think of light both as particles and waves, life and time can be regarded as both an arrow and a cycle—a unique sequence of historical events and a repetitive set of cyclic processes. If viewed only in arrow fashion, as a set of nonrepeating events, the history of life lacks any processes. Yet our perspective is also incomplete without time’s arrow, because evolution is a contingent phenomenon constantly buffeted by history. The scientific “discovery” of deep time greatly lengthened the arrow, bestowing on Earth and the universe an almost unfathomable prehuman history.
Deep time is a recent concept, percolating in the collective human consciousness only since the late eighteenth century. Prior to this time, experts generally agreed that Earth was about 6,000 years old. Several prominent individuals, including physicist Isaac Newton, attempted more precise calculations. Perhaps the most famous estimate came from Bishop James Ussher, who, after lengthy study of the Bible and other sources, determined that Earth was created on the eve prior to October 23, 4004 BC.
Science historians generally attribute the discovery of deep time to the eighteenth-century Scottish polymath James Hutton, often referred to as the father of modern GEOLOGY. In contrast to the “Neptunist” theory of the time, in which all rocks were thought to have precipitated from a single, planetwide flood, Hutton correctly inferred that heat within Earth’s interior is responsible for creating new rock. This so-called Plutonist theory, detailed in Hutton’s The Theory of the Earth (1775), also argued that Earth history is best understood as cycles of repeated events. These events, he claimed, required vast amounts of time, pushing temporal roots far deeper than previously suspected. Hutton’s ideas spawned the principle of uniformitarianism, the concept that past geologic events can be explained by present natural processes, such as volcanism, uplift of Earth’s crust, and erosion by wind and water. In short, the present is the key to the past. With sufficiently great time spans, recurring events literally recycle Earth’s surface—wind and water erase mountain ranges and geologic uplift creates new ones.
Many of Hutton’s key insights did not arise from empirical observations made in the field. Somewhat ironically, his worldview was heavily influenced by the mechanistic perspective of Newton described in the previous chapter. In an effort to forge Newtonian-type links between cosmology and geology, and thereby increase the scientific rigor of his chosen discipline, Hutton supported the idea of the cycle of time. So the man responsible for greatly extending time’s arrow was also a major proponent of time’s cycles. Only after publication of his major treatise did Hutton make several key field observations supporting this idea.
Chief among these observations was recognition of structures known today as unconformities. Within the borders of Scotland, Hutton found layers, or strata, of rocks that differed markedly in their orientations. The underlying rock layers were oriented almost vertically, whereas the overlying layers were horizontal. If a steady accumulation of sediment had occurred, the layers would all be horizontal. Hutton realized that a dynamic series of events must have generated this pattern. At a minimum, four successive steps were required: (1) an initial period in which sediments were deposited horizontally; (2) a subsequent period of deformation or tilting, followed by uplift; (3) erosion of the now-tilted sequence of rocks; and (4) a period of renewed sedimentation on top of the more ancient rocks. Here, then, was a repeating cycle requiring prodigious amounts of time—exactly what the Scotsman was looking for.
If Hutton is the father of modern geology, the nineteenth-century Englishman Charles Lyell is its first son. Lyell, in his famed three-volume Principles of Geology (1830–1833), followed Hutton in claiming that Earth’s history is properly viewed in terms of everyday, slow-acting processes summed over geologic time. By documenting numerous examples in the field, Lyell became the champion of uniformitarianism, arguing that the vastness of planetary time was ample to account for the diverse array of geologic structures seen today, from the depths of the Grand Canyon to the dizzying heights of the Himalayas. His contemporary and friend Charles Darwin embraced the uniformitarian perspective for his theory of evolution. For Darwin, evolution occurred gradually, imperceptibly from the human perspective, with the abyss of deep time more than sufficient to accumulate the diversity of life we see around us. Darwin’s adherence to gradualism and uniformitarianism is beautifully expressed in what is perhaps the best-known passage from his 1859 On the Origin of Species:
It may be said that natural selection is daily and hourly scrutinizing, throughout the world, every variation, even the slightest; rejecting that which is bad, preserving and adding up all that is good; silently and insensibly working, whenever and wherever opportunity offers, at the improvement of each organic being in relation to its organic and inorganic conditions of life. We see nothing of these slow changes in progress, until the hand of time has marked the long lapse of ages.
Yet the uniformitarian view has not been without challengers.
Humans have a curious predilection to define problems in terms of dichotomies, and scientists are no exception. The principal challenge to uniformitarianism has been its logical opposite—catastrophism. The most brilliant catastrophist was Baron Georges Cuvier, an outstanding anatomist of the early nineteenth century and the first person to document extinction in the fossil record. Cuvier noted that certain kinds of fossils were restricted to specific rock strata, and he argued for a series of cataclysmic events that wiped out one group of animals after another. Like Lyell, Cuvier acknowledged that the brevity of human life spans blinds us to the effects of deep time. Rather than invoking slow, ongoing geologic processes, however, Cuvier saw signs in the rocks of tumultuous events such as flooding and volcanism. He noted that if these events were rare, occurring once in millions of years, they would be wholly outside the experience of humans. Yet such periodic cataclysms could, given the immensity of time, conceivably result in the world we see around us.
How are we to reconcile these polar opposites? Of the two ideas, uniformitarianism has fared better since the mid–nineteenth century, becoming the guiding principle of modern geology and paleontology. Many of the geologic processes seen today can indeed be identified in the rock record. Yet, as with many dichotomies, this one is false, because the two alternatives are not mutually exclusive. That is, catastrophic, onetime events could have happened within recurring cycles to shape Earth history. In recent decades, catastrophism has made a roaring comeback, due in part to the idea that an asteroid impact killed off the dinosaurs. This hypothesis has garnered widespread support, and other mass extinctions have been plausibly linked to collisions with extraterrestrial bodies. Whatever the cause(s), these widely separated events were incontrovertibly catastrophic, vindicating Cuvier. Today, both repeated cycles and unique historical events are regarded as integral to Earth’s deep history.
While Lyell, a barrister by training, was successfully convincing the scientific world of the ubiquity of uniformitarianism, geologists in Europe were piecing together the geologic time scale. The largest and most encompassing units of time were referred to as EONS. All plants and most animals are restricted to the Phanerozoic Eon, which began about 540 million years ago. Eons are composed of ERAS, such as the Mesozoic, which in turn comprise PERIODS like the Jurassic. Smaller still are EPOCHS—for example, the recent Pleistocene or the present Holocene. The segeologic units of varying magnitude are distinguished by the composition of their rocks and fossils, as well as by the presence of unconformities. Many subdivisions within the Phanerozoic coincide with the simultaneous disappearance of multiple groups of animals, now recognized as mass extinction events. Geologists also discovered that the same general sequence of rocks occurs in other parts of the world, allowing investigators to correlate geologic units in, say, North America, with those in Europe.
Although cognizant of deep time, nineteenth-century geologists were unable to determine the actual age of the rocks they studied. For example, they could not state with confidence whether dinosaurs lived tens of thousands of years ago or millions of years ago. Put simply, they lacked an appropriate clock. It was only after the discovery of radioactivity at the end of the nineteenth century that scientists devised an effective tool for this purpose.
Variants of a given chemical element possessing the same number of PROTONS in the nucleus but differing numbers of orbiting ELECTRONS are called ISOTOPES. Geologists make use of the steady decay of UNSTABLE ISOTOPES into more stable forms as clocks to measure the ticking of deep time. Specifically, they compare the abundance of naturally occurring radioactive isotopes (parent isotopes) with their decay products (daughter isotopes). Once the ratio of parent-to-daughter isotopes is assessed, the rate of decay is then used to calculate the time elapsed since decay began—that is, the age of the organic matter (in the case of radiocarbon dating) or rocks (most other methods). Fortunately, radioactive isotopes decay at different rates. For example, radiocarbon dating takes advantage of the relatively rapid decay of carbon-14 into carbon-12, estimating the age of organic remains less than about 60,000 years old. In contrast, potassium-40 decays to argon-40 more than two hundred thousand times slower, allowing age estimates in the realm of tens to hundreds of millions of years. Potassium-argon dating is a commonly used method for determining the age of dinosaur fossils and other Mesozoic remains. Based on radioactive isotope dating, Earth’s age is now estimated at 4.6 billion years. Astronomers have used an entirely different set of techniques to assess the age of the universe at 14,000,000,000 (14 billion) years, so Earth has been around just over one-third of that duration.
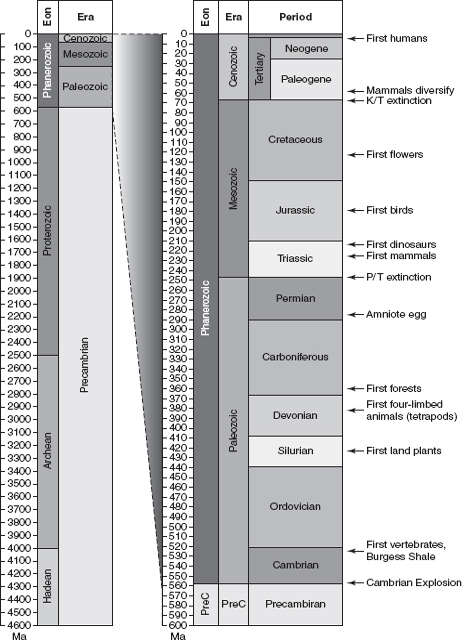
FIGURE 2.1
The geologic time scale. The left column depicts the entire 4.6 billion years of Earth history, noting eons and eras discussed in the text. The right column zooms in on the Phanerozoic Eon (the last 560 million years), showing eras and periods along with key evolutionary events. Both columns include a time scale (denoted in millions of years ago, abbreviated as “Ma”) on the left side.
This temporal immensity offers a new perspective on the web of life. The web of life is vast, at any given moment encompassing the entire BIOSPHERE. Yet it has a historical aspect as well, encompassing most of Earth’s duration. So life’s web is both vast and deep. Viewed from a deep time outlook, the web is never fixed or rigid. Rather, its innumerable, interlinked parts are constantly morphing, as ecological threads spun from the sun’s energy continually replace one another. Meanwhile, evolutionary strands, spun from genetic information, morph at a slower pace, transforming together with the life-forms that temporarily house them.
Given that all organisms are descended from single-celled ancestors, the web of life forms an uninterrupted network extending back to the first BACTERIA, with inestimable numbers of transformations in the intervening eons. All lineages can be thought of as individual genetic threads, with the origin of new forms marked by the branching of distinct strands, some of which evolve into substantial threads themselves. Over time, the web became increasingly complex, with greater and greater numbers of interwoven threads. It’s a fascinating thought experiment to select one particular strand and attempt to trace its convoluted evolutionary history through the abyss of time. With a solid deep time framework now in hand, I will attempt to tease out the thread leading to dinosaurs and take you on a whirlwind, “dinocentric” tour of the epic of evolution, from the big bang to the biggest animals ever to walk on Earth.
About 14 billion years ago, the universe flared forth in the big bang, unfurling space, time, light, and matter in a singular, immense explosion. Less than one minute after this “Great Radiance,” protons and NEUTRONS emerged and combined to form nuclei of the simplest elements, mostly hydrogen and helium. About 300,000 years later, electrons combined with these nuclei for the first time, transforming matter into hydrogen and helium atoms. An infinitesimally small amount of these atoms would one day form the bodies of all dinosaurs. About 11 billion years ago, matter condensed into billions of swirling spiral galaxies. One of these would eventually become known as the Milky Way. Within each immense galactic pinwheel, billions of stars condensed from gas and dust, igniting nuclear fusion that converted vast amounts of hydrogen into helium. After consuming most of their hydrogen fuel, a large portion of these stars—those at least eight times the size of our sun—generated even greater extremes of heat and temperature, forging heavier elements such as carbon, nitrogen, oxygen, silver, magnesium, copper, and iron within their interior furnaces. Ultimately, the giant stars exploded, disgorging their heavy elements into surrounding galactic space. All life on Earth would one day be formed from this wandering stardust. The shock waves from these supernovae also triggered the formation of second-generation stars, resulting in additional cycles of stellar births and deaths.
About 4.6 billion years ago, our sun, an average-sized second-generation star, formed in an outlying spiral arm of the Milky Way galaxy, about 26,000 light-years from the nucleus. The remaining debris disk of gas and dust orbiting the primordial sun condensed into eight planets, together with a bunch of smaller planetoids and moons. The heavier elements—including the atoms that would one day walk the Earth as Stegosaurus and Velociraptor—were concentrated closest to the central star, forming the four rocky worlds we know as Mercury, Earth, Venus, and Mars. Abundant debris remained in the early solar system, however, and impacts with the planets were commonplace. Earth, the third rock from the sun, experienced a particularly violent impact from a Mars-sized world about 4.5 billion years ago. This fortuitous collision carved off a huge chunk of our globe, ejecting countless molten moonlets into Earth’s orbit. In less than a century, these moonlets coalesced to form our moon. The moon stabilized Earth’s axis of rotation and helped set up conditions for life, including the cyclical rise and fall of oceanic tides.
The extraterrestrial bombardment persisted through most of Earth’s first eon, the Hadean, finally coming to an end about 3.9 billion years ago. The effects of this meteoric barrage remain visible on the heavily cratered surface of the moon, which has undergone minimal change during the succeeding billions of years. Earth, in contrast, has always been a dynamic world, receiving regular face lifts from above and below. The Hadean Eon—named after Hades, the mythological hell of the Greeks—is an apt name for Earth’s primordial phase. As the planet was being pummeled from space, geologic turmoil boiled within, generating great bouts of volcanism on the heaving, molten surface. About 4 billion years ago, Earth’s crust formed. Soon after (in the deep time sense), torrential rains began to fall, eventually forming expansive oceans.
Perhaps 3.8 billion years ago (the fossil evidence is equivocal), the dinosaurian thread came to life, literally, animated by single-celled bacteria. Referred to as PROKARYOTES, these microscopic bits of stardust—Earth’s first life-forms—appeared shortly after the planet was cool enough to sustain water. The origin of life remains a mystery, but its rapid emergence suggests to some that life may not have been a lottery-like stroke of luck but a virtual inevitability, given the right conditions. Despite their diminutive sizes, and lacking even a nucleus, these early bacteria were complex. Like us, they had active metabolisms, consuming energy and carrying out hundreds of chemical transformations involving vitamins, proteins, sugars, nucleic acids, carbohydrates, and fats. The environment in which life first evolved remains in question; candidates include the planetary crust miles below the surface and hydrothermal fissures on the ocean floor.
Over the course of the Archean Eon, from 3.8 to 2.5 billion years ago, bacteria blossomed into an array of colors (green, purple, and red), shapes (oval, eel, and rod shaped), and metabolisms (fermenting, photosynthetic, sulfide producing, and oxygen producing). Remarkably, these millions of bacterial forms were not distinct species, at least not in the same way we differentiate lions and tigers, or tyrannosaurs and allosaurs. Like all life, they were able to reproduce, with the vital information of inheritance stored in specialized macromolecules of deoxyribonucleic acid (DNA). Lacking immune systems and rigid barriers, bacteria possess the capacity to share their DNA, enabling them to evolve quickly in response to changing environments. In a sense, then, the bacteria alive at any single moment in time, from the Archean to the present, can be thought of as a single, astoundingly variable species.
Those earliest Archean bacteria grabbed energy directly from the environment through chemical reactions, synthesizing organic compounds with the help of carbon dioxide. Then, sometime between 3.0 and 2.7 billion years ago, a subset of Earth’s bacterial world “learned” to convert light energy from the sun into chemical energy, tapping into a virtually limitless energy source. These blue-green CYANOBACTERIA had discovered PHOTOSYNTHESIS, a process that uses carbon dioxide and water as raw materials and the sun as an energy supply, with oxygen generated as a by-product. This evolutionary event was critical on the road to dinosaurs, because virtually all subsequent life-forms on Earth have depended on the energy of sunlight captured by photosynthesis.
Initially, oxygen generated by the activities of water-living, photosynthetic bacteria was transformed into dissolved iron in the oceans and deposited as iron ore. Eventually, after saturating the seas and painting much of the land red, increasing oxygen levels in the Archean atmosphere triggered an environmental crisis. The biosphere turned poisonous for most bacteria, unable to metabolize this rogue element. Evolution responded to the so-called Oxygen Catastrophe of 2.4 billion years ago by evolving bacterial forms capable of respiring oxygen. Suddenly, using a form of controlled combustion, the dinosaur thread could breathe oxygen. The same substance that had been killing life now enabled it to flourish and diversify in new directions. Oxygen levels in the atmosphere stabilized at a concentration of about 21 percent.
The world had now entered the Proterozoic Eon, which lasted from about 2.5 billion years ago until 545 million years ago. The next major event in the history of the dinosaurian thread occurred about 2 billion years ago, when life became nested in cells with nuclei—the EUKARYOTES. Our present understanding is that certain bacterial cells invaded others as parasites. For some reason, the host did not digest the parasite, and the parasite did not kill its host. Over time, the intruders became permanent, cooperative residents, providing energy to the host cell in return for protection. The newly incorporated bacteria-turned-ORGANELLES were MITOCHONDRIA, destined to become the miniature powerhouses within the cells of dinosaurs (and all other complex life). Another key merger involved photosynthesizing bacteria, relatives of those ancient cyanobacteria, entering some cells containing mitochondria. The merging photosynthesizers were transformed into CHLOROPLASTS, energy-capturing organelles later passed on to plants and algae. The dinosaurian thread, however, had already veered off on a different evolutionary course, tracking with life-forms called PROTISTS.
All of the early eukaryotes were protists—slimy, nucleated organisms whose ranks would eventually include algae, amoebas, water molds, and slime molds.2 After almost 2 billion years in bacterial guise, the dinosaur thread entered a lengthy protist phase that would last another 1.5 billion years. Throughout most of Earth history, all life, including the thread that would one day lead to dinosaurs, remained microscopic. Only in the last 700 million years has the living world exploded into the macro realm, with those slimy protists giving rise to all other complex organisms—plants, animals, and FUNGI. Life’s early history of bacterial mergings also means that every cell of every complex organism that has ever lived on Earth has housed multiple collections of DNA acquired from single-celled ancestors. Dinosaurs were products of ancient evolutionary mergers, as we humans are today. So, contrary to the standard Darwinian outlook of “nature red in tooth and claw,” there is much more to evolution than ruthless, dog-eat-dog-style competition. Cooperation, too, is a prevalent theme in the history of life, one that has been given short shrift by evolutionary biologists.
The first multicellular life-forms appeared about 1.2 billion years ago, consisting of simple cell colonies. Sexual reproduction originated around the same time, accelerating the rate of evolution. A bout of global glaciation embraced the world from 850 to 630 million years ago. At the end of this chilly interval, multicellular life experienced its first major blooming. This was the Ediacara Period (635–542 million years ago), the final days of the Proterozoic Eon. The “Garden of Ediacara,” as it’s been called, included an amazing diversity of animals, both predators and prey, ranging in size from millimeters to meters. Most had soft bodies, but a small portion developed calcified hard parts. Some resembled palm fronds, others were jellyfish-like disks, and still others were reminiscent of mud-filled bags. Most of the strange creatures would soon be gone, but the dinosaur thread was there, having adopted a wormlike guise that would persist for millions of years.
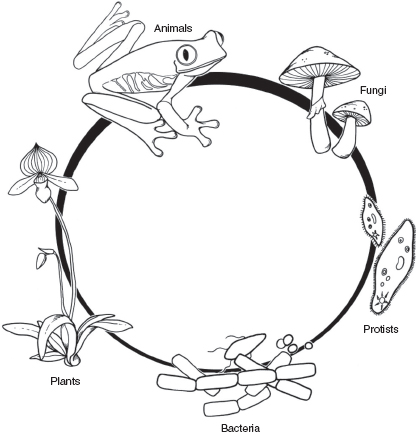
FIGURE 2.2
The five kingdoms of life: bacteria, protists, fungi, plants, and animals. This is one of several schemes used in recent years to organize the diversity of life on Earth.
The remaining 545 million years of Earth history comprise the present eon of “visible life,” the PHANEROZOIC, which includes three periods: the PALEOZOIC (“ancient life”; 545–225 million years ago), MESOZOIC (“middle life”; 248–65.5 million years ago), and CENOZOIC (“new life”; 65.5 million years ago to the present). The Paleozoic, in turn, is subdivided into five periods: Cambrian, Ordovician, Silurian, Devonian, Carboniferous, and Permian.3
The Paleozoic opened with a bang. Often referred to as the CAMBRIAN EXPLOSION, this blast of evolutionary change about 530 million years ago resulted in an astonishing menagerie, including ancestors of most major animal groups alive today. Hard-shelled trilobites were here, as were clams, sponges, snail-like mollusks, and echinoderms—the radially structured precursors of starfish, urchins, and sand dollars. Interactions between predators and prey intensified, with teeth, drills, and claws evolving in response to protective shells. Of course, the thread that would one day connect to dinosaurs and other back-boned animals (VERTEBRATES) was present as well. The closest known representative, Pikaia, still looked very wormy but now possessed multiple body segments. Segmented bodies were an important innovation, enabling complex interactions among parts. The notochord, a flexible rod running the length of Pikaia’s body, would one day become the multipart backbone that gives vertebrates their name. As in trilobites and some other Cambrian INVERTEBRATES, eyes and other sensory organs were now concentrated at one end of the body. The dinosaur thread had added sight to its sensory repertoire.
Committed to the vertebrate path, our thread of interest morphed multiple times and spawned numerous strands along the way. First came small jawless fish, circa 450 million years ago, which later developed elaborate bony armor within the skin. Gill arches were modified to form jaws, resulting in a succession of jawed fishes. By the Ordovician Period (about 460 million years ago), an interior skeleton had developed in fish, initially made of cartilage (as is still the case in sharks and rays) and later of bone. A mass extinction—the first of six in the Phanerozoic and the second-most devastating on record—hammered the biosphere at the close of the Ordovician (about 440 million years ago). Many families of trilobites and clamlike brachiopods vanished, along with entire reef-building communities. The dinosaur thread, however, managed to persist in fish form.
In the wake of the Ordovician extinction, bony fishes evolved alongside the highly successful nautilus-like ammonoids. Sometime during the Devonian, no later than 375 million years ago, one line of bony fishes carried the dinosaur thread from water to land. A long series of transitional fish-amphibians, propelled by stout, muscular fins, now endured the effects of gravity. Vertebrates were latecomers to the terrestrial realm, which had already been occupied for millions of years by all the other kingdoms (bacteria, protists, fungi, and plants) and even other animals (insects). Fungi and land plants had made the jump during the Ordovician, about 100 million years earlier, forming symbiotic partnerships that have persisted to the present day.
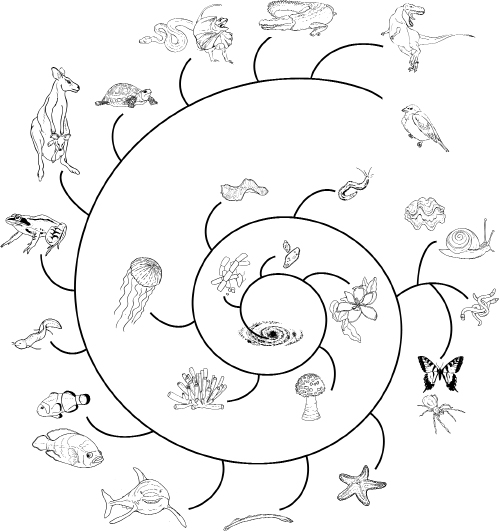
FIGURE 2.3
The dinosaur thread, showing the evolutionary pathway from stardust to dinosaurs, with major branches of life noted. From the center of the spiral to the outside, the branches off the main thread of dinosaur evolution are as follows: bacteria, protists, plants, fungi, sponges, jellyfish, flatworms, round-worms, mollusks, annelid worms, arthropods (the previous three forming a distinct group), echinoderms, lampreys, sharks, bony fishes, lungfish, amphibians, mammals, turtles, snakes and lizards, crocodiles, dinosaurs, and birds. The spiral galaxy at the core of the image symbolizes the fact that all life-forms are made of stardust. Life-forms are not to scale.
The second major mass extinction took place at the end of the Devonian, about 370 million years ago. Once again, the reef builders were decimated and, along with them, brachiopod clams, trilobites, and armored fishes (placoderms). Seemingly undaunted, the dinosaur thread tracked along with the amphibian niche for a substantial chunk of deep time, now borne by sprawled limbs and adapted to breathing air. These early amphibians also developed ears capable of detecting airborne sound waves, radically improving the lineage’s ability to hear (previously limited to sensing low frequency sounds moving through the ground). Initially, members of this line were unable to venture far from their aquatic heritage. But that all changed during the Carboniferous (about 320 million years ago) with the appearance of the amniote egg. Featuring a semi-permeable shell that allowed gas exchange without losing water, the egg acted like a portable sea, enabling AMNIOTES to move permanently onto dry land, no longer tethered by the need to reproduce in or around water. Assuming the guise of reptiles, the thread spread across Earth’s landmasses, unaware of the profound changes that lay ahead.
Later in the Carboniferous, a new strand emerged that would ultimately generate countless generations of mammals and mammal relatives. It is here, in the latter half of the Paleozoic, that the evolutionary paths of humans and dinosaurs diverged. Our last common ancestor with the dinosaur thread was a lightly built lizardlike critter perhaps 20 centimeters (8 inches) long, with splay-toed feet and sprawled limbs transporting its long body and tail. This animal, akin to Carboniferous creatures like Hylonomus, offered few hints of the two grand lineages it was about to spawn. During the following Permian Period, another important parting of the ways occurred when the dinosaur thread split from that of lizards and snakes. Not long afterward, animals within the dinosaur thread first began to look distinctly dinosaurian. These so-called ARCHOSAURS and their near relatives—which would give rise to crocodiles and pterosaurs, as well as to dinosaurs— grew to larger sizes while retaining a predilection for carnivory. As the Permian drew to a close, all of Earth’s continents docked together to form the supercontinent Pangaea, with profound implications for the subsequent course of evolution.
The boundary between the Permian and Triassic periods (and thus between the Paleozoic and the Mesozoic eras) was punctuated by another global paroxysm, the third Phanerozoic mass extinction and the worst overall. This time greater than 90 percent of Earth’s species were exterminated. Somehow, for reasons that remain obscure, the dinosaur thread survived. And, with the ecological slate all but wiped clean, there was now plenty of space for evolution to explore new avenues. The archosaurdinosaur line took full advantage, blossoming into a great variety of mostly midsized carnivorous reptiles that became interwoven into the Triassic web of life. (The evolutionary story of archosaurs will be explored in more detail in chapter 12.) Finally, late in the Triassic, the evolutionary thread that began in stardust more than 10 billion years earlier produced the first true dinosaurs.
We recognize three periods in the Mesozoic: the TRIASSIC (248–206 million years ago), JURASSIC(206–144 million years ago), and CRETACEOUS (144–65.5 million years ago). Dinosaurs underwent an initial evolutionary flourish during the Triassic. Among the earliest forms were meat eaters like Eoraptor and Coelophysis. Although these carnivores perpetuated the bloodthirsty legacy of their ancestors, they were forced to compete with many other, often larger archosaurs. The two major lineages of plant-eating dinosaurs, sauropodomorphs and ornithischians, also appeared at this time. Plateosaurus and other long-necked sauropodomorphs became the largest animals to have existed on Earth, as well as the most common big herbivores on many parts of Pangaea.
Following another mass extinction at the end of the Triassic, Jurassic dinosaurs seized most of the big-bodied niches, herbivore and carnivore alike. Sauropods pushed body size to record-breaking extremes that have never been topped. Also appearing in this period were ornithischian plant munchers like the four-footed stegosaurs and ankylosaurs, as well as the bipedal ornithopods. This was also a period of expansion among the meateating THEROPODS, with predators like Allosaurus and Torvosaurus exceeding 1,000 kilograms (2,200 pounds) for the first time in the long history of carnivory. Beginning in the latter part of the Jurassic, evolution was accelerated by the breakup of Pangaea, leaving dinosaur assemblages isolated on smaller landmasses to pursue independent evolutionary paths.
Dinosaurs reached their acme during the Cretaceous, after the dinosaur thread had spun off several new strands. Dome-headed pachycephalosaurs, horned ceratopsians, and crested hadrosaurs were added to the mix, at least in much of the Northern Hemisphere. Down south, a group of long-necks called titanosaurs reigned supreme. Abelisaur carnivores became top predators on many southern landmasses, while tyrannosaurs terrorized the north. At the very end of this period, Tyrannosaurus shared its evolutionary moment with Triceratops and Ankylosaurus.
The fifth and most famous of the major mass extinctions occurred about 65.5 million years ago, wiping out most of the remaining dinosaurs and virtually all other large animals alive at the time. Yet the dinosaur thread was not completely severed by this cataclysm. Sometime during the Jurassic, one group of small carnivorous dinosaurs found a way to become airborne, recruiting scales modified into filamentous, air-trapping structures. Together with the leathery-winged pterosaurs, these feathered wonders we know as birds graced the skies above Cretaceous dinosaurs. Unlike their larger, earth-bound brethren, birds managed to eke through the extinction bottleneck that brought an end to the Mesozoic. For the past 65 million years, they have diversified into a bewildering array of forms, from menacing ground-dwelling predators to tiny, hovering nectar sippers. The sheer tenacity of the dinosaur thread is underscored by the thousands of bird species alive today.
Within the last million years, barely a sneeze in deep time, the mammal thread attained self-awareness within a strand of gangly, two-legged primates, the most recent of which is Homo sapiens. Gifted with this newfound level of consciousness, humans became the first of Earth’s organisms to contemplate the vast scope of the universe and, ultimately, the place of dinosaurs within the epic of evolution.
Most of us subdivide the living world into three major groups: plants, animals, and microbes or germs. Germs, of course, are regarded as something to be avoided or, better yet, vanquished. Biologists take a broader view based on formal rules of classification. Traditionally, their primary subdivisions of life have been referred to as KINGDOMS. The number and composition of kingdoms has changed over time, largely in response to changing conceptions about evolutionary relationships. One recent scheme, the one followed here, recognizes five kingdoms: bacteria, protists, fungi, plants, and animals. During the epic of evolution, the dinosaur thread was interwoven with three of these kingdoms: in temporal sequence, bacteria, protists, and animals.
Stepping back, several patterns emerge from this stunning, multibillion-year saga. One of these is an overall increase in complexity. The concept of increasing complexity, or progress, has a contentious history among biologists. Yet when viewed in a limited sense—as “production through time of increasingly complex and controlling organisms and societies, in at least some lines of descent”4 —there can be little doubt this pattern applies to the history of life. Complexity clearly increases within the evolutionary sequence that includes hydrogen atoms, molecules, bacteria, protists, fishes, and elephants. The most complex structure known in the universe is also one of the most recent—the human brain. Of course, as noted in the previously cited quotation, not all evolutionary lineages increased in complexity. Indeed, most major groups, from bacteria through mammals, include descendants that differ only in minor ways from primitive forms. Nevertheless, a directional trend toward advancing complexity is apparent.
The increase in complexity was achieved through integration, yielding another pattern: smaller wholes becoming parts of larger wholes. Over and over we see the progressive development of multiple-part individuals from simpler forms. Thus, atoms become integrated into molecules, molecules into cells, and cells into organisms. At each new emergent stage, older forms are enveloped and incorporated into newer forms, with the end result being a nested, multilevel hierarchy.
So the epic of evolution has apparently been guided by two counterbalancing trends that we can refer to as complexification and unification. Complexity increased through unifying simpler forms into ever-more complex wholes. In this way, the wholes at each level fed back on one another, changing as necessary, such that all levels persisted. This journey was not an inevitable, orchestrated march but a creative, quixotic unfolding in which future biospheres could not be predicted. Perhaps most remarkable of all is the sheer scale of the transformations. A short version of our dinocentric story might go something like this: take a bunch of hydrogen atoms, wait several billion years, and you get T. rex.
So there you have it—a glimpse of the entire dinosaur odyssey in a single chapter. The rest, as they say, is details. But, oh, what discoveries await those who are patient enough to rummage through the details. The rest of the book explores various aspects of the world of dinosaurs, progressively building an ever-more detailed picture before attempting a step-by-step walk through the Mesozoic. First things first, however. With the dramatic preamble established, let’s narrow our focus and concentrate on the diversity of dinosaur players that participated in the ever-changing Mesozoic biosphere.
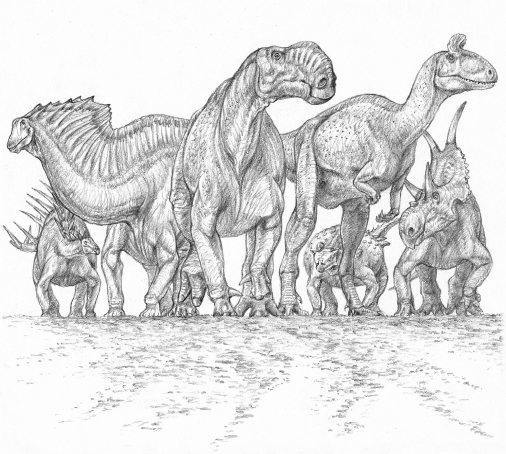
A representative assemblage of dinosaurs from around the world. From left to right: Kentrosaurus(a stegosaur from the Late Jurassic of Africa), Amargasaurus(a sauropod from the Early Cretaceous of South America.), Altirhinus(an ornithopod from the Early Cretaceous of Asia), Cryolophosaurus(a theropod from the Early Jurassic of Antarctica), Minmi(an ankylosaur from the Early Cretaceous of Australia), and Diabloceratops (a ceratopsian from the Late Cretaceous of North America).