15
THE WAY OF ALL CREATURES
WHEN IT COMES TO the death of species, the Grim Reaper assumes several guises. The most common, referred to as BACKGROUND EXTINCTION, tends to be slow paced, ongoing, and small-scale, analogous to the daily sporadic deaths of people in every major city. The causes of background extinctions are typically small-scale as well—for example, flood, drought, or the arrival a new competitor species. The term mass extinction, in contrast, is reserved for extremely rare, large-scale events in which numerous species and even entire groups vanish over a relatively brief period. Whereas each event of background extinction tends to be localized, sometimes limited to single ecosystems, mass extinctions are global, impacting the full spectrum of marine and terrestrial habitats. To wreak such widespread havoc, the causes of mass extinctions must be of that rare sort capable of generating global effects. The list of suspects implicated in these horrific “biocidal” events encompasses both Earth-based and extraterrestrial perpetrators. In between the regular ticking of background extinction and the infrequent sledgehammer of mass extinctions are a variety of midscale events with intermediate effects—for example, climate change resulting in a killer longs-term drought that causes extinctions across an entire continent.
Over the past half billion years or so, the time period for which we have a reasonable fossil record, there have been at least five significant mass extinctions. Our subdivision of Earth history into periods is based in part on the occurrence of these cataclysms, which occurred at the end of the Ordovician (450 million years ago), the Devonian (370 million years ago), the Permian (251 million years ago), the Triassic (213 million years ago), and the Cretaceous (65.5 million years ago).1 Although greater than 98 percent of all species losses are likely attributable to the inexorable ticking of background extinction, this handful of mass extinctions has had profound and long-lasting effects on the biosphere. The unbalanced influence arises from the fact that mass death events simultaneously wipe out not only species but entire clans of species, opening the door to the evolution of new replacement forms from the handful of surviving lineages. And, of course, this effect is multiplied through time; every single organism alive today is part of an unbroken lineage that survived all of these mass extinction events. In effect, then, Mother Nature periodically undertakes a severe pruning of the tree of life, hacking away entire limbs. She then watches the tree regrow, budding off entirely new branches and twigs that ultimately form the foundations of new ecosystems worldwide.
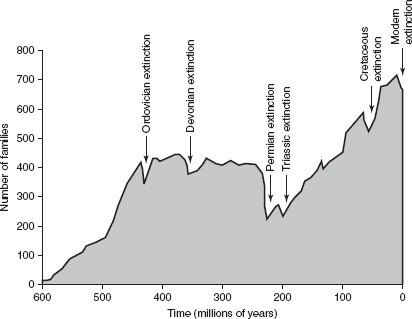
FIGURE 15.1
The five major mass extinction events on Earth during the past 500 million years. The shaded area represents the number of different families of marine invertebrates alive at any given time in the Phanerozoic Eon. Note the relative increase in families through time, with mass extinctions temporarily reducing diversity.
Of the “big five,” the Permian-Triassic (P-Tr) mass extinction—often called the “Great Dying” or sometimes the “mother of all extinctions”—was by far the largest, extinguishing about 70 percent of life on land and 90 percent in the seas, leaving in its wake a devastated biosphere of emptied ecosystems. The P-Tr extinction was perhaps the most dramatic punctuation mark in the history of life, bringing an end to the Paleozoic Era, the “Age of Ancient Life”, and ushering in the Mesozoic Era, the “Age of Middle Life,” sometimes known as the Age of Dinosaurs. Like the subsequent mass dying that wiped out the dinosaurs, the causes of the end-Permian event have long been debated. Possible culprits include asteroid impact, sea-level change, volcanism, and a sudden release of methane-rich water from the seafloor, causing a catastrophic drop in atmospheric oxygen. Of these alternatives, there appears to be growing consensus that the Great Dying was precipitated by large-scale volcanism in Siberia, which in turn led to profound atmospheric changes and ultimately the breakdown of ecosystems globally.
In a recent study, Sarda Sahney and Michael Benton (University of Bristol) suggest that the end-Permian extinction was actually the last (and biggest) of three major extinction pulses that took place in the Permian. These authors argue that Earth’s ecosystems did not fully recover from this decimating trio of events for over 30 million years, until the Late Triassic, which happens to coincide with the appearance of dinosaurs, mammals, and several other groups (see chapter 12).
Viewed in this light, the origin and subsequent radiation of dinosaurs becomes part of the recovery of Earth’s ecosystems following the mother of all extinctions. If one then considers the end-Permian and end-Cretaceous mass extinctions that bookend the Mesozoic, the dinosaur odyssey becomes inextricably interwoven with this pair of biological bottlenecks that literally transformed the face of the globe. Following the end-Permian event, vertebrate diversity on land was reduced to a smattering of amphibians, protomammals, and reptiles, paving the way for the eventual rise of dinosaurs and mammals. After the end-Cretaceous extinction—with the large dinosaurs extinguished and the amphibians, crocodiles, and turtles effectively tethered to freshwater settings— the world was primed for an explosion of mammals on land. Birds, which had begun diversifying prior to the close of the Cretaceous, also radiated following the extinction, perhaps because their major flying competitors, the leathery-winged pterosaurs, did not survive the event.
Presumably because mammals—and, in particular, our primate ancestors—diversified in the wake of the dinosaurs’ disappearance, people are fascinated by the demise of dinosaurs. Indeed, it is one of the topics I and my paleontologist colleagues are asked about most frequently. So I would be remiss if I did not address the matter of their disappearance. Before delving into this story, however, it’s important to emphasize three points.
First, although frequently touted as the ultimate losers and exemplars of backwardness, dinosaurs were anything but failures. As argued in chapter 1, dinosaurs are more aptly regarded as a one of life’s great success stories. The misconception arises in large part from an unrealized truth: death is as inevitable for every species and group of species as it is for every organism. Although dinosaurs are the most famous members of life’s prehistoric pageant, many other groups of organisms preceded them. In every instance, these ancient clans experienced a great flowering for a time, only to be unceremoniously pruned or whacked altogether from the tree of life by a major extinction event. Far greater than 99 percent of all species that have ever lived on Earth are now past tense. To make this point another way, the millions upon millions of species alive today are but a fraction of 1 percent of the total diversity that has inhabited this planet. Thus, the only thing inevitable about the reign of dinosaurs is that it would ultimately come to an end. Nothing to be ashamed of here—just the natural progression of things. Dinosaurs were the dominant large-bodied life-forms on land for over 150 million years. Primates have been around for less than half that duration, hominids have been walking upright less than 8 million years, and humans have been present for less than a half million years. So it takes a lot of gall (or at least a severe case of temporal myopia) to claim some sort of victory over dinosaurs. You may as well snub your nose at your dead relatives because, after all, you’re alive and they’re not.
The second point is that not all dinosaurs went extinct at the same time. As described in earlier chapters, the Triassic, Jurassic, and Cretaceous witnessed an ever-changing procession of dinosaur species and families of species. On the order of seven hundred dinosaur species are currently known; and, given all the fossil-free gaps around the globe and through time, we can be confident that this number represents but a small portion of the total. Extinction and evolution worked hand in hand, eliminating and originating species in such manner as to maintain the web of life. Some-times a single dinosaur species in a single ecosystem was plucked from the web. At other times more sweeping extinction spasms devastated larger areas and many more species. So it’s somewhat misleading to claim that the end-Cretaceous extinction wiped out the dinosaurs. Assuming that the seven hundred or so currently recognized dinosaur species are but a small portion of the total number that existed in the Mesozoic, far less than 1 percent of all dinosaur species were living at the end of the Cretaceous. Like water droplets passing through a swirling whirlpool, a long succession of species sustained the flow of dinosaurs for millions of years. In one sense, then, the surprising aspect of the end-Cretaceous extinction is not that the dinosaurs disappeared, because so many kinds of dinosaurs had vanished prior to that time. No, the remarkable thing is that there were no new dinosaurs to replace those that went extinct. Something happened that was incompatible with the perpetuation of dinosaurs, or, for that matter, any large-bodied animals.
The third and final point is one that I have made several times previously in this book. Dinosaurs actually did not go extinct 65.5 million years ago, at least not all of them. Birds, the direct descendants of dinosaurs, are dinosaurs themselves in a meaningful sense. With on the order of ten thousand living representative species (far outnumbering mammals), you could even make the argument that dinosaurs remain a thriving success story to the present day. And even if your bias is that birds are fundamentally distinct from most dinosaurs, it still must be conceded that avians are a robust legacy handed down by their Mesozoic forebears.
OK, keeping these points in mind, let’s rephrase the question. Exactly what transpired 65.5 million years ago to take out Tyrannosaurus rex and most of its dinosaur contemporaries? Over the years, well over a hundred explanations have been proposed, running the gamut from the reasonable and testable to the inane and ridiculous. The list of proposed killing agents includes disease, slipped vertebral discs (because dinosaurs were so big), loss of interest in sex, poison plants (leading to diarrhea or constipation, depending on the hypothesis), fungal invasions, climatic change in the form of global cooling or warming (once again, depending on the hypothesis), cosmic radiation from a supernova, egg-eating mammals, sunspots, nasty aliens, and not enough room on Noah’s Ark.
My personal favorite is the racial senility hypothesis. According to this long-outdated idea, dinosaurs as a group experienced a period of restless youth in the Late Triassic and Early Jurassic marked by great expansion in both form and diversity. This era of youthful exuberance was followed by a kind of adulthood during the latter Jurassic and early part of the Cretaceous, a “heyday” of sorts when dinosaurs supposedly reached their evolutionary peak. Finally, there was the inevitable period of senility toward the end of the Cretaceous. Evidence put forth in support of this notion included the marvelous variety of bony excrescences on Late Cretaceous dinosaurs—the frills and horns of ceratopsians, the crests of hadrosaurs, the boneheads of pachycephalosaurs, and the tail clubs of ankylosaurs. Adherents of the racial senility hypothesis perceived these unusual features to be unnatural and useless, perhaps the result of hormones gone wild and certainly symbolic of a group on its way out.
Whereas some of these extinction hypotheses are just plain wacky, most suffer from a common problem. They consider only the charismatic megafauna, dinosaurs, and ignore the many other groups that suffered major losses in the end-Cretaceous extinction. This event is usually referred to as the Cretaceous-Tertiary, or K-T MASS EXTINCTION. (Here K is used instead of C because the latter designates a much earlier period—the Carboniferous.) In the marine realm, the entire spectrum of top vertebrate predators, including the plesiosaurs and mosasaurs, disappeared together with several groups of fishes. The ichthyosaurs, those dolphin-look-a-like reptiles, disappeared about 30 million years before the end of the Cretaceous, so their demise must have had a different cause. Among nonvertebrates, several important, long-lasting groups were also wiped out in the K-T event; these include a variety of hard-shelled marine organisms, such as the nautilus-like ammonites and reef-making rudist clams. Also victimized were the chalk-making algae, famous for generating massive limestone deposits like the White Cliffs of Dover. Although some forms survived, decimation of the chalk producers likely had deep and cascading effects, because these microscopic organisms formed much of the base of the marine food web. Overall, it appears that the hardest hit in the marine realm were free-swimming or surface forms such as plankton and ammonites (although many varieties of fishes made out just fine). Among bottom-dwellers, several filter-feeding groups were decimated, including the colony-forming bryozoans, “sea-lily” crinoids, and corals, whereas scavengers took only minor hits. All of these losses interrupted the flow of energy and cycling of nutrients through the biosphere, in some cases shutting down nutrient cycles altogether.
Meanwhile, up on land, the pattern of extinction and survival has been more difficult to assess. The biggest problem is that few well-studied rock and fossil sequences span the time period immediately before and after the K-T extinction. Based on this very limited sample, it appears that lizards and mammals were severely decimated, and pterosaurs were extinguished altogether. Freshwater sharks, common predators in Mesozoic streams and rivers, also disappeared in the end-Cretaceous event. On the flip side, several groups of vertebrates weathered the storm with few to no species losses, including fresh-water fishes, amphibians, turtles, crocodiles, and a strange group of narrow-snouted, crocodile-like reptiles called champsosaurs. At least two major conclusions emerge from this pattern of losers and winners. First, with few exceptions, all animals larger than about 10 kilograms (22 pounds) were killed off, so being big turned out to be a lethal liability. Second, freshwater vertebrates—including crocodiles, turtles, amphibians, champsosaurs, and fishes (but not sharks)—did much better than land dwellers. In the near future, it may be possible to compare these findings with similar inventories from other locales around the world. Nevertheless, the bottom line is that any reasonable hypothesis of the K-T extinction must explain much more than the dinosaurian demise, accounting for patterns of extinction and survival across the full spectrum of marine and terrestrial life.
We are left with only two plausible hypotheses for explaining the K-T extinctions. The first of these I refer to as the SILVER BULLET HYPOTHESIS. This now-familiar idea posits that the extinction was brought about by a singular, devastating event—an asteroid impact. According to this view, first proposed in 1980 by the father-son team of Luis and Walter Alvarez and their colleagues, an asteroid about 10 kilometers (6 miles) in diameter collided with Earth one fateful day about 65.5 million years ago. Slamming into the planet at a velocity of about 100,000 kilometers per hour (62,000 miles per hour), the resulting explosive force of this extraterrestrial bullet exceeded that of all of the present stockpiles of nuclear weapons combined. The event triggered a massive earthquake (perhaps greater than 10 on the Richter scale), well beyond anything in recorded history, causing tsunamis to radiate away from ground zero–like monstrous ripples on a planet-sized pond.
Upon impact, the asteroid disintegrated, vaporizing a chunk of Earth’s crust with it. The huge volume of ash and pulverized rock ejected high into the atmosphere enveloped the world, blocked the sun, and transformed day to night. What followed was a prolonged period of cold and dark that lasted about 4 months. Lacking a solar energy source, photosynthesis effectively ceased, shutting off the energy supply to most marine and terrestrial ecosystems worldwide. As if a giant asteroid plunging the biosphere into darkness were not sufficiently hellacious, additional lethal side effects may have included rampant wildfires and acid rain. Another possible killing agent was a massive pulse of infrared heat in the few hours following impact, as millions of bits of molten rock ejected into the atmosphere from the impact site rained down around the globe. In short, the Silver Bullet model claims that dinosaurs and their contemporaries were thriving during the latest Cretaceous until a giant rock collided with Earth, bringing their charmed lives to an abrupt halt.
The major competitor of the Silver Bullet scenario is an idea that I term here the BLITZKRIEG HYPOTHESIS. Blitzkrieg, a German compound word that means “lightning war,” refers to an offensive military strategy developed in World War II by the German Wehrmacht. It entailed use of multiple weapons, including air bombardments and deployment of ground forces, designed to outmaneuver and overwhelm the enemy. With regard to the K-T event, the name fits well with the view that this mass extinction was precipitated not by a single cause but by multiple factors working in unison from the ground and air. Specifically, advocates of the Blitzkrieg view argue for three disruptive agents working in concert: receding sea levels, erupting volcanoes, and an asteroid impact. In other words, instead of a single strike from outer space, a barrage of attacks on the latest Cretaceous world reshuffled the biological deck and discarded the dinosaurs.
Unsurprisingly, the media immediately jumped on the Silver Bullet scenario. Here in one tidy package was an unimaginably violent solution to one of nature’s greatest outstanding mysteries—the death of the dinosaurs. The idea had the added bonus of a possible repeat performance, with the potential to wipe out humankind next time round. News outlets have to wait a long time for science stories of this caliber. The impact hypothesis also sparked a worldwide research effort in a variety of scientific fields. Not long after the idea took hold, a pair of paleontologists reviewed the fossil record over the past 500 million years and reported that mass extinctions appeared to be cyclical, occurring about once every 26 million years. This announcement prompted astronomers, in turn, to search for an extraterrestrial cause to explain such metronome-like spasms of extinction. One innovative solution with a terrific name was the Deathstar hypothesis. Just outside the outer limits of our solar system orbits a thin band of comets known as the Oort cloud, named after its discoverer. Proponents argued that an unseen dwarf star—known as “Nemesis” or, simply, the Deathstar—might also exist not far beyond our solar system. According to this idea, the dwarf star perturbs the Oort cloud every 26 million years or so, unleashing a plague of comets on the solar system. Several sensationalist books were written on this topic before paleontologists determined that the original hypothesis of a 26-million-year extinction cycle was flawed, in part because of poor resolution of the timing of extinctions. Although debate about extinction cycles continues, the Deathstar hypothesis resides with cold fusion and other discarded ideas in the gutters of science.
Nevertheless, the Silver Bullet hypothesis is alive and well, with the asteroid impact scenario supported by multiple lines of evidence. The first (and in many ways still most pivotal) of these lines was identified by the Alvarezes in their 1980 article. These geoscientists found an anomalous abundance of the element iridium in a thin stratum of rock within the K-T boundary layer. Iridium, which comes from the same chemical group as platinum, is rare in Earth’s crust but common in extraterrestrial bodies like asteroids. The “iridium spike,” as it’s called, was first identified in a clay layer near Gubbio, Italy, but has since been confirmed in K-T boundary sediments at many other locations around the world. Additional geologic evidence includes the presence of shocked quartz and glass spherules in the boundary layer. Shocked quartz is quartz modified by intense heat and pressure, leaving behind characteristic lines—exactly the kind of thing that might be produced by an asteroid impact. Similarly, glass spherules form during impact events as rock is melted, blasted into the air as a spray of droplets, frozen almost immediately, and then dropped back to Earth.
On the biological side, paleobotanists have documented a radical decimation of plant life at the end-Cretaceous boundary. Just below the iridium layer, fossil pollen and spores from many locations around the world indicate a diverse flora. Immediately above this layer, the pollen and spore evidence is heavily dominated by a single plant type—ferns. Ferns are one of the first plants to recover after major environmental catastrophes, and this “fern spike,” as it’s been called, is cited as evidence of a sudden cataclysm—that is, death by Silver Bullet.
The scientific world was stunned by the Alvarez impact hypothesis, with the iridium layer providing the geologic equivalent of a smoking gun. Almost equally surprising was discovery of the actual wound in Earth’s surface—a crater about 200 kilometers (125 miles) across in the Gulf of Mexico, near the Yucatán Peninsula. The Chicxulub crater appears to be the right age and size to account for the impact of a 10-kilometer (6-mile) diameter asteroid. It’s also associated with a number of physical effects consistent with a major impact, including shocked quartz, glassy spherules, and tsunamis or tidal waves. The sediments are even arranged in a layer-cake sequence consistent with the Silver Bullet model. First to be deposited were large particles like the glassy spherules, which would have rained down from the sky in the minutes following the impact. Immediately above this stratum are sediments indicative of tsunamis, or tidal waves, which in turn are topped by iridium and other fine sediments that would have remained airborne longer before falling back to the surface.
In short, although considerable debate persists as to the relative roles played by such factors as acid rain, wildfires, and the cessation of photosynthesis, there is strong consensus that an asteroid hit Earth about 65.5 million years ago and that its effects on the biosphere were catastrophic. But was the asteroid the overriding factor in the K-T extinctions (the Silver Bullet model), or were additional killing agents involved (the Blitzkrieg model)?
It turns out that strong support exists for protracted environmental changes in the latest Cretaceous, well prior to the asteriodal collision. As noted in the previous chapter, Earth’s atmosphere experienced a gradual cooling between about 90 and 65 million years ago, culminating in an estimated 2°–3°C drop in temperature by the K-T boundary. Many investigators see evidence of a particularly intense pulse of global climate change in the last few million years of the Cretaceous, marked by a distinct warming trend. And there is now abundant evidence that much of the latest Cretaceous environmental shift resulted from a combination of volcanism and sea-level change, both cited as key factors of the Blitzkrieg model.
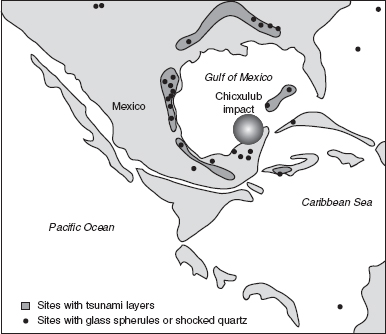
FIGURE 15.2
Map of Central America during the Late Cretaceous, showing the Chicxulub impact site and localities that have yielded additional evidence (glassy spherules and tsunami deposits) of an asteroid strike that may have caused the K-T extinction.
The tectonic pulse in the Late Cretaceous generated volcanism in various areas around the globe, especially in and around the Pacific Ocean. By far the greatest episode of volcanism, however, the largest since the end-Permian extinction, produced the DECCAN TRAPS, an incredible flood of lava that inundated much of present-day India and Pakistan. In all, the Deccan Traps released on the order of 1 million cubic kilometers (240 cubic miles) of basalt, equivalent to covering the entire continent of North America with lava about 20 kilometers (12 miles) deep! The duration of this event is still hotly debated. Whereas older estimates pointed to a lengthy duration, on the order of 10 million years, more recent estimates are consistent with a much briefer volcanic burst, with peak eruption levels occurring in the 1–2 million years immediately surrounding the K-T boundary. A bout of volcanism of such great magnitude undoubtedly had dramatic and devastating environmental effects, yet the exact nature of those effects remains unclear. If the Deccan Traps pumped vast amounts of carbon dioxide into the atmosphere, this event could have resulted in global warming. Conversely, ejection of tons upon tons of particulate matter into the atmosphere may have reduced available sunlight at the surface, resulting in a long-term cooling trend. What we do know is that the eruptions tended to be viscous rather than explosive—more akin to volcanoes on the Hawaiian Islands than to Mount St. Helens.
Meanwhile, the sea-level highs that resulted in shallow seas atop many of the world’s continents finally subsided in the latest Cretaceous, leading to a global marine regression. Maximal retreat of the seaway occurred just prior to the K-T boundary, uncovering vast swaths of continental crust that had been underwater for millions of years. This transformative event greatly increased the amount of area available for land-dwelling and freshwater animals, while vastly reducing coastal and marine habitats. Additional effects included lengthening of stream systems (which had to flow much farther to reach the ocean) and emplacement of land bridges between previously separate continents. The land bridges resulted in exchanges of plants and animals between landmasses. One example mentioned in the previous chapter stretched from present-day Alaska to eastern Asia at least twice during the Late Cretaceous, forming a corridor for the movement of dinosaurs and other life-forms between North America and Asia. Viewed as a cause of the K-T event, however, regression is problematic, because even larger regressions occurred at other times in the Mesozoic without the consequent extinctions.
Nevertheless, with a wealth of evidence supporting the latest Cretaceous occurrence of an asteroid impact alongside dramatic episodes of volcanism and marine regression, it might seem that the Blitzkrieg hypothesis would emerge victorious over its Silver Bullet rival. Yet, before any such pronouncement can be made, we need more information about species extinctions during the K-T event. At the core of this debate is the pace of extinction. A rapid extinction—say, as brief as a week or a year, and no more than thousands of years—would support a single, devastating cause (Silver Bullet), whereas a more protracted period of dying—on the order of millions of years—might indicate multiple causes (Blitzkrieg). Many paleontologists claim that the fossil record shows numerous groups—for example, ammonites, plesiosaurs, pterosaurs—on the wane for millions of years prior to the K-T asteroid impact, with much lower diversities than earlier in the Cretaceous. Others argue that ecosystems remained relatively diverse until 65.5 million years ago, when a major event (presumably the asteroid impact) resulted in a virtually instantaneous killing strike. Unfortunately the rock record rarely provides the degree of resolution necessary to distinguish between such alternatives. An inch (2.5 centimeters) of sedimentary rock may record part of a day or many thousands of years. A study by Michael Benton found that the K-T extinction wiped out 64 out of 210 vertebrate families, with the greatest losses among the land-dwelling vertebrates, or tetrapods (43 percent of families lost). Yet these figures cover a period of about 5 million years. So we need more data on whether these losses were concentrated into a relatively brief instant of deep time or spread out over millions of years.
I think it’s safe to say that, over the past couple of decades, most dinosaur paleontologists have tended to reside in the Blitzkrieg camp, arguing for a gradual extinction precipitated by multiple factors. However, despite all the fuss about their extinction, the fossil record of dinosaurs is remarkably sparse for the final stage of the Mesozoic. Indeed, to date, only one place on Earth—the Hell Creek Formation exposed most abundantly in eastern Montana and the Dakotas—has been investigated in detail. Several authors have argued that the Hell Creek fauna possessed far fewer species than earlier dinosaur faunas. In particular, comparisons have been made with the approximately 75-million-year-old Dinosaur Park Formation of southern Alberta, with its bewildering array of herbivorous and carnivorous dinosaurs (see the previous chapter).
For a long time, I regarded myself as a card-carrying member of the gradualist-Blitzkrieg camp. However, three lines of evidence have caused me to change my mind and join the ranks of Silver Bullet enthusiasts.2 First is the documentation of many additional K-T boundary sites preserving that anomalous iridium layer; the presence of this asteroidal signature at far-flung locations around the globe confirms that the Chicxulub impact was truly a devastating, global event. Second is a growing fossil database indicating that the terminal Cretaceous world was not stressed to the breaking point, awaiting arrival of an extraterrestrial coup de grâce. With regard to dinosaurs in particular, recent work in the Hell Creek Formation (see chapter 5) and nearby Lance Formation have documented a much more diverse fauna than previously realized. Certainly some dinosaur lineages (e.g., short-frilled ceratopsids and crested duck-bills) went extinct prior to the end of the Cretaceous, but overall dinosaur diversity remained relatively high, with many groups represented by truly gigantic exemplars such as Tyrannosaurus, Ankylosaurus, and Pachycephalosaurus.
Third, new and improved estimates of the stratigraphic age and duration of dinosaur species from earlier in the Late Cretaceous suggest that these somewhat older ecosystems (dating to about 75 million years ago) were not nearly as diverse as previously supposed. For a long time, the Dinosaur Park Formation in Alberta, Canada, has been hailed as the exemplar of dinosaur diversity. Yet recall from the previous chapter that many dinosaur species recovered from these Alberta badlands do not co-occur in time. Instead, recent study of the rock record reveals that these dinosaurs had much lower ecological diversity than previously thought, a finding more consistent with rapid replacement of species (high rates of origins and extinctions) rather than numerous coexisting species at any one time. Considered in unison, the second and third lines of evidence suggest that the difference in co-occurring species between the two formations is not nearly as large as previously believed. So, even within the well-sampled Western Interior of North America (let alone the rest of the world, for which we currently have no hard data), I see no grounds for arguing that dinosaurs (or other groups of animals or plants) were undergoing a slow, attritional demise in the latest Cretaceous. We must keep in mind, however, that our knowledge of dinosaur diversity at the very end of the Cretaceous is effectively limited to one place in western North America. Fossil evidence from elsewhere in the world may one day tell a very different story.
Finally, it seems questionable to argue for a lethal cascade of agents when compelling evidence exists for a single agent capable of doing the job on its own. Would the dinosaur-laden biosphere have persisted long after 65.5 million years ago if an asteroid had not slammed into the Gulf of Mexico, bringing an abrupt close to the Mesozoic? We will never know for sure, but my hunch is that it would have, with evolution pumping out new and wondrous species. Why not, given that dinosaurs had already been around for 160 million years? Although the scythe of extinction was ultimately inevitable, it might have been millions of years before another such devastating event. Some authors, such as paleontologist Niles Eldredge of the American Museum of Natural History, now argue that not much happens in evolution until ecosystems are upturned by external forces like those that produce mass extinctions. Certainly without that asteroidal collision, the grand radiation of mammals would have been thwarted or at least significantly delayed, and there would be no humans today to contemplate this dramatic series of events.
I offer just one caveat. Research is growing that the Deccan Traps formed over a relatively brief period (less than a million years). One team of researchers recently argued that a 600-meter- (2,000-foot-) thick portion of the massive lava flows may have piled up in only 30,000 years. If further research supports these findings and points to a pulsed episode of volcanism in association with the K-T boundary, then we may have a problem. It would seem to be a gargantuan coincidence that a lethal asteroid strike would co-occur with one of the biggest bouts of volcanism known. Gerta Keller (Princeton University) and her colleagues have argued on geologic and biologic grounds that the Chicxulub impact evidence does not support a global killing event from space. Instead, they (and others) point to the Deccan Traps as the likely “Silver Bullet” that killed off the dinosaurs and other life-forms at the close of the Mesozoic. So the final chapter on this story may not yet be written.
Sometime during the mid-1980s, dinosaur paleontologist Jack Horner got fed up with being asked about the dinosaur extinction. One day, after facing that same question for the umpteenth time, Horner famously replied, “I don’t give a damn how they died. I want to know how they lived.” I confess to being with Horner on this matter. Indeed, this book serves as a testament to my passion for understanding dinosaur lives. To me, the most interesting aspect of mass extinctions is not the presumed causes of all the death and mayhem (OK, so volcanoes release lots of lava and Earth occasionally collides with big rocks). I am much more fascinated by life’s response in the aftermath of such extinctions. How do the survivors get back on track? And what has prevented the biosphere from collapsing altogether when decimated by mass extinctions?
To provide at least a partial answer, I conclude this chapter with a brief discussion of a radical idea. Life is no mere innocent bystander constantly changing in response to environmental perturbations; instead, organisms actively generate and maintain the biosphere, in part by regulating the temperature and composition of the atmosphere. This dramatic, encompassing, and surprising idea, known as the GAIA HYPOTHESIS (named after the Earth goddess of Greek mythology), pertains as much to the present day as to the world of dinosaurs. Over the past few decades, this hypothesis has acquired increasing numbers of adherents, though it is still a long way from becoming mainstream, broadly accepted science. I position myself firmly within the ranks of its supporters.
Over the course of deep time, global climates on Earth have witnessed dramatic variations. This book has explored the hothouse conditions that characterized the Mesozoic, including degrees of global warming and sea-level rise well beyond the direst predictions of the present day. And there have been other, equally extreme hothouse intervals during the history of life. Yet Earth has been an icehouse world as well. You may be surprised to learn that the tenure of humanity has occurred largely within an icehouse interval (the so-called Ice Age), albeit one that may be coming to a rapid close thanks to human inputs into the atmosphere. Far more severe icehouse conditions than the Pleistocene ice ages occurred in the remote past. Between about 730 and 630 million years ago, during the Proterozoic eon, the planet may have been virtually enveloped in ice, a phenomenon aptly called “Snowball Earth.”3
Underlying such dramatic extremes, however, is a story of astonishing climatic stability. Astronomers tell us that our star, the sun, has become markedly brighter over the past 4 billion years, increasing its radiation output by about 25 percent. Yet, rather than a progressive rise in temperature, as might be expected, the surface of Earth has tended to become cooler. Despite notable icehouse and hothouse fluctuations, Earth’s surface temperature has remained relatively constant for most of its history, within the relatively narrow temperature range suited for life. What is the source of such enduring atmospheric stability and biological perseverance in the face of changing external conditions?
The apparent solution to this mystery came from a British atmospheric scientist by the name of James Lovelock. NASA hired Lovelock in the 1960s to design experiments for the Viking space probes going to Mars. His job was to determine how to use the probes to test for the presence or absence of life on the Red Planet. After contemplating the matter, Lovelock decided that we should be able to determine whether a planet has life simply by examining its atmospheric makeup. After all, he reasoned, life on the surface of any planet would have to use its atmosphere to cycle the elements necessary for existence. Nutrients must be supplied and wastes removed. Earth’s atmosphere is composed of approximately two-thirds nitrogen and one-fifth oxygen, with several other elements such as carbon dioxide and methane occurring in trace amounts. Oxygen is highly reactive, particularly in the presence of methane. Therefore, in order to maintain constant levels, oxygen must be constantly replenished by the metabolic activities of organisms such as plants and certain bacteria, which continually produce oxygen as a waste product of photosynthesis. An increase in the amount of oxygen to a level of 25 or 30 percent could cause the atmosphere to burst into flames. A decrease to below 10 percent would be lethal to the vast majority of life-forms.
So it seems that something has kept the composition of the atmosphere, as well as its temperature, within narrow limits conducive to life for billions of years. In contrast, the atmosphere of Mars is effectively “dead”—or, more precisely, at equilibrium—composed almost entirely of carbon dioxide, with no free oxygen. Based on this observation, Love-lock concluded that Mars could not presently harbor any life. A similar claim could be made for the other nearby rocky world, Venus. The difference between the two is that carbon dioxide has turned the surface of Venus into an oven, whereas the atmosphere on Mars is so thin that it cannot retain heat. In contrast, here on our blue planet the oceans have not boiled away, and the atmosphere has maintained a temperature and composition suitable for life. Lovelock, sparked by a radical insight, argued that the planetary signature of life is an atmosphere far from equilibrium that includes highly reactive elements such as oxygen. Remove all traces of life on Earth, and our atmosphere would soon migrate to an equilibrium state dominated by carbon dioxide.
By all accounts, NASA was not overly enthusiastic about Lovelock’s findings, because his “remote-sensing” argument could be construed as negating any need to travel to Mars in search of life. Lovelock’s employment with the space agency was terminated soon thereafter. Of course, the presumed absence of life on Mars today does not mean that it has never existed there. On the contrary, the ancient presence of water, a key ingredient for life, was recently confirmed by geologic evidence recovered by Martian rovers. Remarkably, then, our best shot at discovering life elsewhere in the universe may rest with paleontologists exploring the rocks of Mars!
Unperturbed by his exit from NASA, Lovelock realized that he was on to something big. In contrast to the traditional view of environment as a purely physical phenomenon, his hypothesis implicated life in the generation of its own environment. Just as a thermostat keeps a room within a given temperature range, the gas-exchanging metabolic processes of life must interact with the physical processes of a planet in order to regulate the temperature and composition of the atmosphere. Some have misconstrued the Gaia hypothesis as claiming that Earth is “alive” or, worse, a single organism, pronouncements embraced by New Age spiritualists and scorned by hard-nosed scientists. However, at its core, this idea makes no such assertion. Rather, Lovelock and other Gaia supporters argue that the sum of planetary life has an integrated physiology that enables the biosphere to regulate its environment. One of Lovelock’s closest colleagues, microbiologist Lynn Margulis, puts it this way: “The planet’s surface is not just physical, geological, and chemical, or even just geochemical. Rather, it is geophysiological: it displays the attributes of a living body composed of the aggregate of Earth’s incessantly interactive life.”4
Yet how could such a regulatory process occur on a planetary scale? How does Gaia “know” when to raise or lower oxygen levels in the atmosphere, and what has prevented a catastrophic buildup of greenhouse gases like carbon dioxide? Whatever the mechanism involved, it must be driven by an unconscious, automatic process, akin to the way your body regulates its internal temperature by shivering or sweating, heals itself following an injury, and allows you to maintain balance while walking.
To answer this perplexing question, Lovelock and Margulis compiled a compelling argument. In short, they postulate that, merely by growing and reproducing, consuming nutrients and generating wastes, bacteria, fungi, plants, and animals participate in self-regulating feedback loops. Feedback loops, briefly introduced in chapter 5, are every-where in nature. Instead of conscious monitoring, these loops employ circular control, gathering information about past performance and using this input to self-correct as necessary. Whereas negative feedback loops tend to bring a system back to a previous stable state, positive feedback loops drive a system to change until a new stable state in achieved. An example of negative feedback is the way in which the animals maintain relatively stable internal conditions (homeostasis) through adjustments to such regulatory factors as body temperature, blood pressure, and blood glucose levels. A biological example of positive feedback, or amplification, is the runaway evolution of bizarre features like peacock tails (chapter 10).
All this may sound a little mystical, so let’s look at a concrete example—carbon. Over many millennia, volcanoes have spewed millions upon millions of tons of carbon dioxide into Earth’s atmosphere. Yet, in contrast to Venus and Mars, it has remained a trace component of the atmospheric makeup. Therefore, there must be mechanisms that pump carbon dioxide out of Earth’s atmosphere. One such mechanism comes in the form of plants, which consume huge amounts of carbon dioxide through photosynthesis, converting the carbon into a solid form that is then stored within their bodies. Animals balance this carbon sequestering at least in part by exhaling carbon dioxide with every breath. Another method of trapping carbon is rock weathering. As rocks break down in response to erosion, they combine with rainwater and carbon dioxide to form various carbon-based compounds called carbonates. The gaseous carbon dioxide is thus taken from the atmosphere and bound up into liquid form. Yet calculations show that the purely physical processes of rock weathering are insufficient to remove the necessary carbon dioxide from the atmosphere.
It turns out that many soil bacteria, through partnerships with fungi and plants, act as superb catalysts for rock weathering, increasing the rate of this process by many times. To give just one example, some bacteria secrete compounds that swell inside cracks and fissures within the rock, accelerating the pace of weathering about a thousand-fold. Most of this water-borne carbon ends up in the oceans, where a variety of life-forms tap into the resulting chemical compound, calcium carbonate, to make their shells. Shelled organisms include large-bodied animals such as clams and snails. Far more abundant are microscopic organisms called coccolithophores, the same life-forms responsible for the formation of vast reservoirs of Cretaceous chalk such as the White Cliffs of Dover. So the carbon dioxide that started out as a gas and was transformed to a liquid ultimately becomes locked up in a solid. When these animals die, their shells descend to the ocean bottom, piling up in massive layers that turn to limestone. After a lengthy residence on the sea floor, this limestone-entombed carbon is transported into Earth’s interior via the process of subduction (see chapter 4). Ultimately, a portion of this carbon dioxide is released again as a gas through volcanic activity. The entire cycle, from volcanoes to rock weathering to soil bacteria to oceanic algae to limestone and back to volcanoes, forms a feedback loop that contributes to regulating Earth’s temperature. Remarkably, it’s estimated that this cyclical and ongoing process of rock weathering cools the planet by some 15°–45 °C (60°–110°F)!
As the theory goes, an increase in atmospheric carbon dioxide initiates a corresponding increase in photosynthetic and rock-weathering organisms, which in turn accelerate the rate of carbon capture from the atmosphere. Once carbon dioxide levels in the atmosphere drop sufficiently, the biomass of photosynthesizing plants and bacteria decreases, reducing the rate of carbon capture and allowing the system to reenter a state of equilibrium.5 Other feedback loops involving biological and geologic processes have been identified, and many more likely await discovery. It is mind-boggling to think of life playing a pivotal role in its own fate, keeping the planet habitable for eon upon eon.
The Gaia hypothesis gives us a new perspective on the world of dinosaurs. The Mesozoic web of life, including the dinosaurs, was maintained by a complex, robust interweaving of life-forms, most of them microscopic. So fully integrated were these with Earth’s physical processes that they formed a system of feedback loops regulating the Mesozoic atmosphere and perhaps the biosphere more generally. The end-Permian and end-Cretaceous extinctions—which precipitated the origin and demise of the dinosaurs, respectively—undoubtedly had major impacts on this regulatory process, as well as the composition of the web. Nevertheless, the biosphere, Gaia, has proved remarkably resilient, rebounding from such cataclysmic events multiple times over billions of years.
The Mesozoic odyssey of dinosaurs is a tale of fantastic creatures in distant lands. As with any such epic, the narrative includes lengthy periods of quietude punctuated by brief intervals of upheaval. We have seen that Mesozoic Earth bears a strong resemblance to the planet we live on today. Then, as now, ecology and evolution worked in tandem to sculpt resilient ecosystems bursting with splendor and diverse life-forms, many of which would look familiar to us. Yet this ancient Earth would seem alien as well, and not only because of the plethora of dinosaurian monsters. Dinosaurs inhabited a hot-house world lacking rain forests, icy polar caps, and grasslands. Pterosaurs graced the skies, while mosasaurs, ichthyosaurs, and plesiosaurs plied the oceans. Today, we still grope to understand even the basics of Mesozoic biomes—the large-scale communities of plants, animals, and soil organisms. Paleontology and its allied sciences will continue to refine our conceptions of these ancient worlds. An equivalent review of dinosaurs written 20 years from now will be very different indeed, achieving deep insights we cannot yet imagine. And future versions 40, 60, and 100 years hence will be equally distinctive. It’s an exciting time to be a paleontologist, and the excitement shows no signs of abating anytime soon. For the few who elect to pursue paleontology, as either a professional or an amateur, rest assured that many, many lifetimes of groundbreaking work remain to be done. And for the vast majority who prefer a front-row seat on the sidelines, all I can say is, enjoy the show!

A rock wren alights on the horn of a Triceratops skull eroding out of the Hell Creek badlands, Montana, momentarily underscoring an evolutionary link between the present day and the deep past.