PERHAPS all scientists recall their graduate student years as an idyll. But surely some research environments are more idyllic than others, and I think there was something special about the Tinbergen group at Oxford in the early 1960s. Hans Kruuk has captured the atmosphere in his affectionate but not hagiographic biography, Niko’s Nature.42 He and I arrived too late for the heroic ‘hard core’ period described by Desmond Morris, Aubrey Manning and others, but I think our time resembled it – though we saw less of Niko himself, because his room was in the main Zoology Department while all the rest of us were housed in the annexe at 13 Bevington Road, a tall, narrow house in north Oxford, about half a mile from the main Zoology building tacked onto the University Museum in Parks Road.
The senior figure in 13 Bevington Road was Mike Cullen, probably the most important mentor in my life – and I believe most of my contemporaries in the Animal Behaviour Research Group (ABRG) would agree. To try to explain the debt that all of us owe to this remarkable man, I can do no better than quote the closing words of the eulogy that I spoke at his memorial service in Wadham College, Oxford, in 2001.
He did not publish many papers himself, yet he worked prodigiously hard, both in teaching and research. He was probably the most sought-after tutor in the entire Zoology Department. The rest of his time – he was always in a hurry and worked a hugely long day – was devoted to research. But seldom his own research. Everybody who knew him has the same story to tell. All the obituaries told it, in revealingly similar terms.
You would have a problem with your research. You knew exactly where to go for help, and there he would be for you. I see the scene as yesterday. The lunchtime conversation in the crowded little kitchen at Bevington Road, the wiry, boyish figure in the red sweater, slightly hunched like a spring wound up with intense intellectual energy, sometimes rocking back and forth with concentration. The deeply intelligent eyes, understanding what you meant even before the words came out. The back of the envelope to aid explanation, the occasionally sceptical, quizzical tilt of the eyebrows, under the untidy hair. Then he would have to rush off – he always rushed everywhere – perhaps for a tutorial, and he would seize his biscuit tin by its wire handles, and disappear. But next morning the answer to your problem would arrive, in Mike’s small, distinctive handwriting, two pages, often some algebra, diagrams, a key reference to the literature, sometimes an apt verse of his own composition, or a fragment of Latin or classical Greek. Always encouragement.
We were grateful, but not grateful enough. If we had thought about it we would have realized, he must have been working on that mathematical model of my research all evening. And it isn’t only me for whom he does this. Everybody in Bevington Road gets the same treatment. And not just his own students. I was officially Niko’s student, not Mike’s. Mike took me on, without payment and without official recognition, when my research became more mathematical than Niko could handle. When the time came for me to write my thesis, it was Mike Cullen who read it, criticized it, helped me polish every line. And all this, while he was doing the same thing for his own official students.
When (we all should have wondered) does he get time for ordinary family life? When does he get time for his own research? No wonder he so seldom published anything. No wonder he never wrote his long-awaited book on animal communication. In truth, he should have been joint author of just about every one of the hundreds of papers that came out of 13 Bevington Road during that golden period. In fact, his name appears on virtually none of them – except in the Acknowledgements section . . .
The worldly success of scientists is judged – for promotion or honours – by their published papers. Mike did not rate highly on this index. But if he had consented to add his name to his students’ publications, as readily as modern supervisors insist on putting their names on papers to which they contribute much less, Mike would have been a conventionally successful scientist, lauded with conventional honours. As it is, he was a brilliantly successful scientist in a far deeper and truer sense. And I think we know which kind of scientist we really admire.
Oxford sadly lost him to Australia. Years later, in Melbourne, at a party for me as visiting lecturer, I was standing, probably rather stiffly, with a drink in my hand. Suddenly, a familiar figure shot into the room, in a hurry as ever. The rest of us were in suits, but not this familiar figure. The years vanished away. Everything was the same – though he must have been well into his sixties, he seemed still to be in his thirties – the glow of boyish enthusiasm, even the red sweater. Next day he drove me to the coast to see his beloved penguins, stopping on the way to look at giant Australian earthworms, many feet long. We tired the sun with talking – not, I think, about old times and old friends, and certainly not about ambition, grant-getting and papers in Nature, but about new science and new ideas. It was a perfect day, the last day I saw him.
We may know other scientists as intelligent as Mike Cullen – though not many. We may know other scientists who were as generous in support – though vanishingly few. But I declare, we have known nobody who had so much to give, combined with so much generosity in giving it.
I almost wept when I spoke that eulogy in Wadham chapel, and I almost wept again just now when rereading it twelve years later.
I don’t know whether the camaraderie of 13 Bevington Road was exceptional, or whether all groups of graduate students nurture a similar esprit de corps. I suspect that being housed in a separate annexe rather than in a large university building improves the social dynamics. When the ABRG (and other outliers such as David Lack’s Edward Grey Institute of Field Ornithology and Charles Elton’s Bureau of Animal Populations) eventually moved into the present concrete monster on South Parks Road, something, I believe, was lost. But it may just be that by then I was older and more weighed down by responsibilities. Whatever the reason, I retain a loyal affection for 13 Bevington Road and my comrades of those times who foregathered at the Friday evening seminars, or in the lunch room, or over the bar billiards table in the Rose and Crown: Robert Mash, whose epidemic sense of humour I later recalled in my foreword to his book How to Keep Dinosaurs;43 Dick Brown, chain smoking, hard drinking and implausibly rumoured to be religious; Juan Delius, whose deliriously eccentric brilliance never ceased to amuse; Juan’s supernormally delightful wife Uta who gave me German lessons; the tall, blond Dutchman Hans Kruuk, who later wrote Niko’s biography; the Scotsman Ian Patterson; Bryan Nelson the gannet man, known to me in my first six months only from the enigmatic notice on his door, ‘Nelson is on the Bass Rock’; bearded Cliff Henty; David McFarland, Niko’s eventual successor who, although based in the Psychology Department, was a sort of honorary member of our group because his vivacious wife Jill was Juan’s research assistant, and the couple had lunch in Bevington Road every day; Vivienne Benzie, who introduced the sunny New Zealand girls Lyn McKechie and Ann Jamieson as yet other honorary members of the lunch group; Lou Gurr, another smiling New Zealander; Robin Liley; the jovial naturalist Michael Robinson; Michael Hansell, who later shared a flat with me; Monica Impekoven, with whom I wrote a paper later; Marian Stamp, whom I was to marry; Heather McLannahan, Robert Martin, Ken Wilz; Michael Norton-Griffiths and Harvey Croze, who later formed a consulting partnership in Kenya; John Krebs, who later collaborated with me in writing three papers; the daredevil Iain Douglas-Hamilton, unwilling exile from Africa while he wrote his thesis on elephants; Jamie Smith, with whom I wrote a paper on optimal foraging in tits; Tim Halliday the newt man, Sean Neill with his lovingly restored Lagonda and gift for drawing cartoons, Lary Shaffer, master photographer, and other friends whom I apologize for omitting.
The Friday evening seminars were the highlight of the week for the Tinbergen group. They lasted two hours and frequently spilled over into the following Friday, but the time flashed by because, instead of the soporific formula of an hour spent listening to one speaker’s voice followed by questions at the end, our two hours were enlivened by argument throughout. Niko set the tone by interrupting almost before the speaker could complete his first sentence: ‘Ja, ja, but what do you mean by . . .?’ This wasn’t as irritating as it sounds, because Niko’s interventions always aimed at clarification and it was usually necessary. Mike Cullen’s questions were more penetrating, better informed and more feared. Other notable contributors – brilliant in their idiosyncratic ways – were Juan Delius and David McFarland, but the rest of us chipped in without inhibition too, almost from the first day we were there. Niko encouraged that. He insisted on absolute clarity about the question we were asking in our research. I recall how shocked I was, on visiting our sister research group at Madingley in Cambridge, to hear one of the graduate students beginning to describe his research with the words: ‘What I do is . . .’ I had to restrain myself from imitating Niko’s voice: ‘Ja, ja, but what is your question? Years later, I related this story when I gave a research seminar at Madingley. I refused to identify the culprit to a mock-scandalized Robert Hinde, the formidably intelligent and charismatic leader of the Madingley group who later became Master of St John’s College, Cambridge, and my lips are sealed to this day.
The question Niko set for me was a version of the question often la-belled with the ‘nature or nurture?’ cliché derived from The Tempest:
A devil, a born devil, on whose nature
Nurture can never stick . . .
Philosophers down the centuries have pondered the question. How much of what we know is natively built in, and to what extent is the young mind a blank slate, waiting to be written over, as John Locke believed?
Niko himself, like Konrad Lorenz (with whom he is credited with co-founding the science of ethology), was early associated with the ‘nature’ school of thought. His most famous book, The Study of Instinct,44 which he later pretty much disowned, used ‘instinct’ as a synonym for ‘innate behaviour’, defined as ‘behaviour that has not been changed by learning processes’. Ethology is the biological study of animal behaviour. Various schools of psychology also study animal behaviour, but with different emphasis. Psychologists historically tended to study animals like rats or pigeons or monkeys as substitutes for humans. Ethologists historically were interested in the animals in their own right, not as proxies for anything. Consequently they have always studied a much wider range of species, and they tend to emphasize the role of behaviour in the natural environment of the species. Ethologists also, as I have said, historically emphasized ‘innate’ behaviour, whereas psychologists were more interested in learning.
In the 1950s, a group of American psychologists started to take an interest in the works of the ethologists. Prominent among them was Daniel S. Lehrman, a big man with a deep knowledge of natural history as well as of psychology. He also spoke adequate German, which made him an effective bridge between the two approaches to animal behaviour.
In 1953 Lehrman wrote a very influential critique of the traditional ethological approach. He strongly criticized the whole notion of innate behaviour, not because he thought everything was learned (although some psychologists whom he quoted did), but because he thought it was in principle impossible to define innate behaviour: impossible to devise an experiment to demonstrate that any particular piece of behaviour is innate. Theoretically, the obvious method was the ‘deprivation experiment’. Imagine if humans were given no verbal instruction in how to copulate and no opportunity to observe other species – not even the smallest inkling. Would they know how to do it when the opportunity finally presented itself? It’s an intriguing question, and there might be telling anecdotes, perhaps about over-sheltered and naive Victorian couples. But in non-human animals we can do experiments. Deprivation experiments.
If you rear a young animal in deprived conditions without the opportunity for experience, and it still knows how to behave properly, that must mean the behaviour is innate, inborn, instinctive. Mustn’t it? But Lehrman objected that you couldn’t deprive the young animal of everything – light, food, air, etc. – and that it is never obvious how much deprivation is needed in order to satisfy the criterion of innateness.
The dispute between Lehrman and Lorenz got personal. Lehrman, whose family background was Jewish, caught Lorenz out in some suspiciously Nazi-inflected writings from the war years and did not shrink from mentioning this in his famous critique. Lorenz, on first meeting Lehrman after the critique was published, said (approximately): ‘I thought from your writings that you must be a small, mean, wizened little man. But now that I see you are a BIG man [and Lehrman was indeed a very big man] we can be friends.’ This avowal of friendship didn’t stop Lorenz trying – Desmond Morris tells the story as an eye-witness from inside the car – to intimidate Lehrman by almost mowing him down with an enormous American car that he was driving in Paris.
But back to the controversy over nurture or nature. Male sedge warblers (to take just one example) have a complex and elaborate song, and they can perform it even when reared in isolation, never having heard another sedge warbler. The Lorenz–Tinbergen school would therefore have said it must be ‘innate’. But Lehrman emphasized the complexity of developmental processes and always wondered whether learning was involved in some less obvious way. For Lehrman, it wasn’t good enough to say that the young animal had been reared under deprived conditions. For him, the question was: ‘Deprived of what?’
Since Lehrman’s critique was published, ethologists have indeed discovered that many young songbirds, including sedge warblers, even when reared in isolation, learn to sing their correct species song by listening to their own fumbling efforts, repeating the good fumbles and discarding the bad. So that looks like nurture after all. But in that case, Lorenz and Tinbergen might reply, how do the young birds know which of their fumbles are good and which bad? Surely that ‘knowledge’ – a template for what their species song ought to sound like – has to be innate? All learning does is transfer the song pattern from the sensory part of the brain (the built-in template) to the motor side (the actual skill of singing the song).
Other species, by the way, such as the American white-crowned sparrow, also teach themselves to sing in this ‘fumbling’ way, but do need to have heard the species song earlier in life. It is as though the young bird takes a ‘tape recording’ before it can sing, and uses it as a template for teaching itself how to sing. And there are intermediates between the ‘learned tape recording’ and the ‘innate tape recording’ as templates for later learning.
This was the philosophical minefield into which Niko Tinbergen released me in 1962. I think he wanted to back away from his perceived association with Lorenz and saw me as a bridge towards the Lehrman camp. My experimental subject was to be not singing birds but baby chicks pecking. I did a series of experiments of which I’ll mention only one here.
Baby chicks straight out of the egg start pecking at small objects, presumably looking for food. But how do they know what to peck at? How do they know what’s good for them? One extreme would be for nature to endow them, before they have any experience at all, with a template picture of a grain of wheat in the brain. That’s unrealistic, especially in an omnivore. Do wheat grains and mealworms and barleycorns and millet seeds and beetle larvae have anything in common, as opposed to boring and inedible marks and stains? Yes, they do. For one thing, they are solid.

How do you recognize something as solid? One way is by surface shading. Look at the photographs of moon craters. They are the same photograph, but one is rotated through 180° relative to the other. My guess is that on the left you will see hollow craters and on the right solid flat-topped hills – and the other way around if you swivel the book upside down. The illusion has been known for a long time. It depends upon a preconception about where the light is coming from: in effect, a preconception about the location of the sun. Solid objects tend to be brighter on the side nearest the sun, which will usually be approximately above. A photograph of a solid object can therefore look hollow if you turn it upside-down, and vice versa.
The sun is seldom directly overhead, but the general direction of its light is more likely to be down than up. Therefore any predator seeking solid objects as possible prey can use surface shading cues based upon that assumption. And on the other side of the predator–prey arms race, natural selection might well favour prey animals that manage to disguise their solidity by ‘countershading’. Many species of fish are darker on top, lighter below, which tends to neutralize the natural tendency for sunlight to come from above, and thereby makes the fish look flatter. One fish, the ‘upside-down catfish’, is a genuine ‘exception that proves the rule’. It habitually swims upside-down and, sure enough, it is reverse countershaded: darker on its belly than its back.
A Dutch student of Tinbergen called Leen De Ruiter did some neat experiments on reverse countershaded caterpillars, who habitually rest upside-down. The upper picture shows Cerura vinula in its normal position. It looks flat and inconspicuous. The lower picture shows what it looked like when De Ruiter turned its twig upside-down: much more conspicuous to my eyes and – more significantly – to the eyes of jays, when De Ruiter used them as experimental predators.

But none of this says anything about whether – in jays or humans – the knowledge that the sun is normally overhead is innate or learned. The solid shading illusion seemed to me to provide a good opportunity to test the question, using baby chicks in deprivation experiments.
First, did chicks see the illusion? Apparently, yes. I photographed a half ping-pong ball lit asymmetrically, and printed the image to be about the size of a tempting grain or seed. When I viewed the photograph with the illuminated side at the top, the hemisphere looked solid. When I inverted the photograph, it didn’t. When chicks were offered a choice between the two orientations, they strongly chose to peck at the apparently solid picture, the one lit from above. This suggested that chicks possess the same ‘preconception’ we do, that the sun is normally overhead.
So far, so good; but these chicks, though young, were not completely naive. They were three days old and had been feeding in normal overhead light during this time. They might have had time to learn the appearance of solid objects illuminated from above.
To test this I did a crucial experiment. I reared chicks with light coming from below and tested them under the same conditions. So, at the time of testing, they had never had any experience of overhead light. As far as they were concerned, the world into which they had hatched was a world with a sun underneath them. Every solid object they had ever seen, whether food objects or parts of other chicks, was lighter underneath than on top. I expected that, when tested with the two ping-pong ball photographs, they would prefer to peck at the one illuminated from below.
But I was delighted to be proved wrong. The chicks overwhelmingly pecked at the photograph illuminated from above. If you accept my interpretation, this means that the chicks are genetically equipped by ancestral natural selection with something equivalent to ‘advance information’: in the world in which they are to live, the sun will normally shine from above. My experiment had pinpointed a true example of innate information which is not reversed by a positive attempt to teach the contrary.
I can’t think of any group of humans who habitually live with underfloor lighting. If they exist, it would be interesting to test them in the same way I tested my chicks. I thought about offering an intuitive guess as to what the result would be, but I honestly prefer not to place a bet. Wouldn’t it be fascinating if we too saw the illusion innately? Having been surprised by the chicks, I’d be only slightly more surprised if humans did the same. We may never know, but there could be ways to do the experiment on very young babies. They don’t peck, but they do fixate their eyes on objects that interest them, and you can measure that. Could a developmental psychologist offer babies a version of my ping-pong ball experiment and measure the time they spend staring at each of the two photographs? Would it be considered unethical to use underfloor lighting for a baby’s room for the first few days of life? I can’t see why, but who knows what the verdict of a modern ‘ethical committee’ might be?
In the end, my work on ‘nature or nurture’ constituted only a small part of my doctoral research,45 and it was relegated to an appendix in my thesis. The main part of my thesis had little in common with it, except that it also involved pecking in chicks. And it was also an attempt to illustrate a point of philosophical interest – although taken from a different part of philosophy. It became possible through an improved technique for recording pecks.
Bevington Road, and especially its satellite research stations in the great gull colonies of the north, ran a system of ‘slaves’ – young unpaid volunteers who wanted a brief taste of the Tinbergen experience before going to university. Among them were Fritz Vollrath (who later returned to Oxford to head a flourishing group working on spider behaviour, and remains a close friend) and (also from Germany) Jan Adam. Jan and I found an immediate affinity, and we worked together. He had remarkable workshop skills – combining the very different virtues of my father and Major Campbell – and, fortunately, these were the days before health and safety regulations interfered to protect us from ourselves and sap our initiative. Jan and I had the freedom of the departmental workshops: lathes, milling machines, bandsaws and all. We (that is to say Jan, with me as willing apprentice – the younger brother syndrome again, I suppose) built an apparatus to automate the counting of chick pecks, using delicately hinged little pecking keys, elegantly made from scratch by Jan, with sensitive micro-switches. Previously, when working on the surface shading illusion, I had counted pecks by hand. Suddenly, I was in a position to collect huge quantities of data automatically. And this opened the door to a completely different kind of research, motivated by a different philosophy, Karl Popper’s philosophy of science, which I learned from Peter Medawar.
As I have already explained, I had come to know of Medawar early on through my father, who was a schoolfriend of his. As British biology’s star intellectual, Medawar came to give a visiting lecture at his old Oxford department when I was an undergraduate there, and I remember the excited buzz in the standing-room-only audience waiting for this tall, handsome, gracious figure to arrive (‘This lecturer has never been thought ungracious in his life,’ as a later critic said of him). The lecture prompted me to read Medawar’s essays, later anthologized in The Art of the Soluble and Pluto’s Republic,46 and it was from them that I learned about Karl Popper.
I became intrigued by Popper’s vision of science as a two-stage process: first the creative – almost artistic – dreaming up of a hypothesis or ‘model’, followed by attempts to falsify predictions deduced from it. I wanted to do a textbook Popperian study: dream up a hypothesis that might or might not be true, deduce precise mathematical predictions from it, and then try to falsify those predictions in the lab. It was important to me that the predictions should be mathematically precise. It was not enough to predict that a measurement X should be larger than Y. I wanted a model that would predict the exact value of X. And this kind of exact prediction demanded large quantities of data. Jan’s apparatus for counting massive numbers of pecks gave me the opportunity. Instead of pecking at photographs of ping-pong balls, my birds pecked at little coloured hemispheres mounted on Jan’s hinged windows, which triggered micro-switches. They preferred blue over red over green, but that wasn’t what interested me. I wanted to know what governed each individual pecking decision, whichever colour it was directed towards. And this, of course, was only a specimen of a more general question about how decisions are made at any time by any animal.
Medawar elsewhere made the point that scientific research doesn’t develop in the same orderly sequence as the final published ‘story’. Real life is messier than that. In my own case it was so messy that I can’t remember what gave me the idea for my ‘Popperian’ experiments. I remember only the finished story which, as Medawar would have expected, gives an implausibly tidy impression.
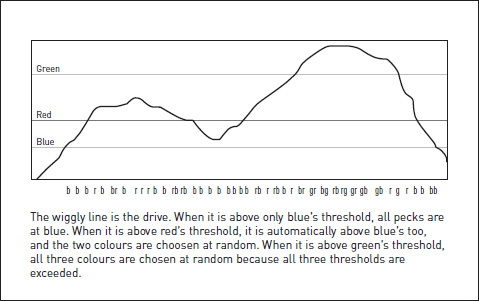
The finished story is that I dreamed up an imaginary ‘model’ of what might be going on inside a chick’s head when it makes a decision between alternative targets, did some algebra to deduce precise, quantitative predictions from the model, then tested them in the lab. The model itself was a ‘drive/threshold’ model. I postulated that there was a variable (‘drive’ to peck) in the bird’s head, whose graph was continuously wiggling up and down as the drive strengthened or weakened (perhaps at random; it didn’t matter). Every time the drive happened to rise above the threshold for a colour, the bird was capable of pecking at that colour (something else, for which I developed and tested another model which I’ll mention later, determined the timing of pecks). Blue, being a preferred colour, had a lower threshold than green. But if the drive rose above green’s threshold, it automatically had to be above blue’s threshold as well. What would the bird do then? I postulated that it would be indifferent between the two colours, since both thresholds were exceeded: it would ‘toss a coin’ to decide between them. So the model predicted that a bird’s choices over a long period would consist of periods of pecking at only the preferred colour, interspersed with periods of choosing at random between the two. There would be no periods of consistently positive choice of the less preferred colour.
I didn’t at first look directly at sequences of pecks. That was to come later, after I moved to California. I think the reason I didn’t test sequences at first was as unambitious as the fact that Jan’s apparatus could count pecks but not record the exact order in which they happened; and Jan himself had by now gone back to Germany, so wasn’t there to modify his apparatus. I think, too, that I was simply seduced by the Popperian elegance of deducing a mathematical formula which would predict some measured quantity from some other measured quantities.
The chicks happened to prefer blue over red over green. I imagined an experiment in which I would present Blue versus Green, Blue versus Red, and Red versus Green, counting the proportion P of pecks to the preferred colour in each case. This would give me three numbers (PBestWorst, PBestMedium, PMediumWorst). It’s only to be expected that PBestWorst would be larger than either of the other two. But could the model predict precisely how much bigger? Could I deduce from the model a formula to predict exactly what PBestWorst should be, if I fed in PBestMedium and PMediumWorst? Yes, that is exactly what I succeeded in doing. I defined algebraic symbols to stand for the time spent by the drive between various thresholds, did some school algebra (simultaneous equations as taught by Ernie Dow) to eliminate the unknown variables, and was pretty pleased when, at the end of pages of algebra, a simple, precise, quantitative prediction dropped out. The drive/threshold model predicts that
PBestWorst = 2(PBestMedium + PMediumWorst – PBestMedium • PMediumWorst) – 1.
I called this Prediction 1. The thing that interested me about Prediction 1 was that it is quantitatively precise.
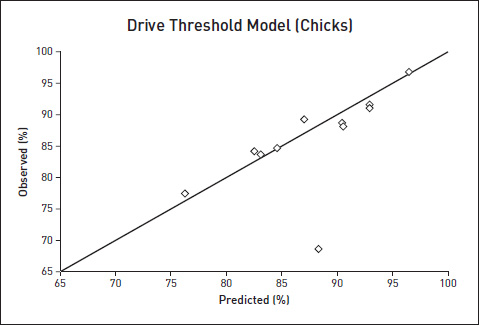
So now to test it. Would the chicks obey the prediction? Yes: to my delight and amazement, in seven out of eight repeats of the experiment they did, very closely. The eighth experiment was way off, so much so that, to my acute embarrassment, when one of my papers was published in the journal Animal Behaviour,47 the printer removed the relevant point from the graph, thinking it must be a blemish on the block! Fortunately the offending datum was clearly present in the accompanying table, otherwise I might have been accused of dishonesty. I did another set of experiments on chicks, involving not pecking but walking into chambers illuminated by light of different colours. The graph shown here combines the two sets of experiments and plots the observed against predicted percentages for all 11 chick experiments.
If the model’s predictions were perfect, the points should all lie exactly along the diagonal line. With the exception of Experiment 8, as already mentioned, the Drive Threshold Model does a far better job than we ever dare to expect in animal behaviour experiments (physicists expect higher precision because there is usually less statistical error in their measurements).
I also used all the same data to test the predictions of an alternative model, one which simply assumed that each colour has a ‘value’ for the animal, and that the animal allocates its choices in proportion to the colour’s value. The two models gave similar predictions, so that if one is right the other one can’t help being nearly right. But the Drive Threshold Model was consistently more accurate in predicting the observed result. The ‘colour value’ model consistently overestimated PBestWorst. The ‘colour value’ model was falsified. The Drive Threshold Model triumphantly survived the attempt to falsify it, and indeed its predictions were (with the exception of the one experiment) remarkably accurate.
Does this good performance of the model really mean that there is something equivalent to a fluctuating ‘drive’ in the chick’s head, crossing ‘thresholds’, and that something equivalent to tossing a coin happens when the drive is above more than one threshold? Well, Popper would say that the model survived a strong attempt to disprove it; but that says nothing about what the ‘drive’ and the ‘thresholds’ actually correspond to in the language of nerves and synapses. It is at least an interesting thought that you can make inferences about what is going on inside the head without cutting it open.
The same method of imagining a model and testing its predictions has proved enormously productive in many branches of science. In genetics, for example, you can infer the existence of chromosomes as one-dimensional linear sequences of genetic code without ever looking down a microscope, using only the data from breeding experiments. You can even work out the order in which the genes are arrayed along the chromosomes, and how far apart genes are from each other, entirely by imagining what might be the case and testing predictions in breeding experiments. As with my experiments on solidity and shading, I think of my Drive Threshold Model as an illustrative example of the kind of thing that can be done with a model, rather than as a conclusive discovery of what is really going on inside a chick’s head.
I elaborated the Drive Threshold Model in various directions (that’s also something that is supposed to happen according to Popperian philosophy) and tested nine predictions in all, with good success. One of these elaborations of the model, as I mentioned above, was an attempt to explain the exact timings of pecks (‘samplings’ of the position of the ‘drive’ relative to the ‘thresholds’). The predictions of this model stood up well against data on black-headed gull chicks from my colleague and close friend Dr Monica Impekoven, a visitor to Bevington Road from Switzerland. We published a joint paper on this work.48
Another elaboration of the model, which I published as the ‘Attention Threshold Model’,49 was an attempt to probe more deeply the ‘penny tossing’ of the original Drive Threshold Model: the indiscriminate choice of target when more than one threshold is exceeded. Briefly, I suggested that chicks attend to one dimension at a time – colour, shape, size, texture etc. – and in a definite order. Each of these attention systems has its own version of the Drive Threshold Model. The chick attends to the first dimension – say, colour. If the colour system’s drive/threshold delivers a definite choice, the chick goes for the preferred colour, say Blue. But if the colour system’s verdict is a ‘penny toss’ the chick switches its attention to something else, say shape, and ignores colour. From the colour system’s point of view, choosing by shape is equivalent to choosing at random. But of course it is not random from the shape system’s point of view. This trickle-down process continues through all the attention systems. If all else fails, the equivalent of a ‘penny toss’ is something like ‘choose the nearest one’. The Attention Threshold Model yielded a series of further predictions (making nine predictions in all), which I tested with success.
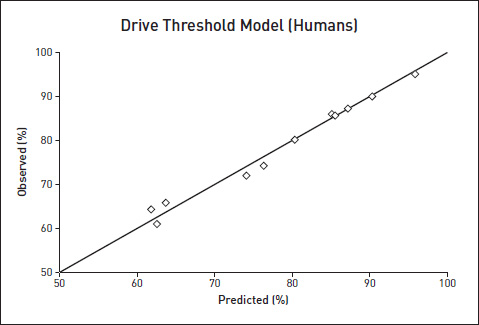
Again, as with the solid shading experiments: is it possible that a version of the Drive Threshold Model might apply to humans? I searched the scientific literature and found that several psychologists had done pairwise preference tests on humans. Their motivation was different from mine, but I could use their published results. There are various reasons why a psychologist might present a range of choices in all possible pair combinations: to test an idea in Voting Theory, for instance. Instead of offering a three-way choice between Conservative, Liberal and Socialist, with either winner-take-all or rank-order voting, a pollster might investigate the benefits of pairwise testing: ‘How would you vote between Conservative versus Liberal (if there were no other choice), between Liberal versus Socialist (given no other choice), and finally between Conservative versus Socialist?’ Anyway, for whatever reason, psychologists have presented humans with choices in all possible pairwise combinations. I was therefore able to feed their measurements of Best versus Medium, and Medium versus Worst, into my formula, and test my model’s prediction of Best versus Worst. The data came from a diverse set of studies: American students choosing handwriting samples, American students choosing vegetables, American students choosing bitter/sweet tastes, and Chinese students choosing colours. In addition, I was especially delighted to be able to use a big study of preferences for composers shown by the members of the Boston Symphony Orchestra, the Philadelphia Orchestra, the Minneapolis Symphony Orchestra and the New York Philharmonic. Following is a graph pooling all the results from humans. Once again, if the Drive Threshold Model’s predictions were perfect, the points would fall along the diagonal line. I must say I was pretty excited when I saw how closely the prediction was fulfilled. Predictions in behavioural biology just aren’t usually fulfilled that precisely!
The orchestra study was large, and processing the data laborious. I discussed the problem with my Uncle Colyear, who was by then in the Oxford Forestry Department, lecturing and advising on statistical methods. He suggested that I should learn to program the university computer. He and his wife Barbara got me started and helped me to write a program for the composer preferences. And so began my forty-year time-wasting and soul-consuming love affair with computer programming, a love affair that is now happily over: I am still an intensive computer user, but I now leave the programming to professionals.
Back then in the mid-1960s there was only one computer in Oxford University: a brand new English Electric KDF9, less powerful than an iPad today, but state-of-the-art in its time and filling a large room. The programming language favoured by my uncle and aunt was K-Autocode, a British alternative to Fortran with a similar structure and grammar, and a similar tendency to foster bad programming practice (absolute jumps, for instance). At that time American computers used great stacks of punched cards (vulnerable to being dropped and irretrievably shuffled) and British ones used punched paper tape (spewed out into great spaghetti mounds on the floor, which then had to be rolled up and were vulnerable to tearing). Thank goodness those days are over. Thank goodness, too, that computers now communicate with us via a screen or loudspeaker, not on reams and reams of paper – and immediately, instead of after a 24-hour delay.
But in those days we knew no better, and I was captivated. I was entranced by the idea of pre programming a sequence of operations and chugging through them step by step with a pencil to check them, then unleashing them in the computer to be rerun at very high speed and thousands upon thousands of times. I had a terrible night when I dreamed that I was a computer running my program and I spent – as it seemed – the whole night going through repeat loop after repeat loop in my fevered brain. To be fair, conditions that night were not ideal for sleeping anyway. Along with several of the Bevington Road crew, I had been persuaded by my friend Robert Mash to spend the weekend hunting the Surrey Puma.
Ever since 1959, sightings of a mysterious large carnivore had been reported in the woods of Surrey in southern England. Dubbed the Surrey Puma, it had achieved the status of a minor Yeti-style myth, and a group of us joined together to spend a weekend in May 1966 trying to find it. The newspapers heard about the scheme and, with news material getting thinner as the summer Silly Season approached, the Observer printed a photograph of me in a British Empire pith helmet (solar topee) of the kind that I had worn as a child. I have forgotten where my companions pitched their tents, but my assigned role was to spend the night in a sleeping bag, out in the open under the stars, surrounded by great haunches of raw meat. I was allocated a camera with a flash, and my marching orders were to photograph the puma if it should come to take the meat – or me, I suppose. I didn’t sleep peacefully, to say the least, so perhaps it was not surprising that my computer nightmare should have come on that particular night. Dawn came as a great relief to me and my companions, and a dreamily misty dawn it was (as you can see from the photograph reproduced in the picture section). We never did find the Surrey Puma, and it may be significant that sightings went on being reported as late as 2005, which would seem to suggest that the Surrey Puma lived more than twice as long as the maximum recorded lifespan of its species, even in captivity.
My computer habit moved on from the KDF9 to a smaller but more accessible computer. Oxford Zoology had acquired a dynamic new professor (head of department in the Oxford-speak of the time) in succession to the genial and slightly batty-looking Sir Alister Hardy. The stiff new broom from Cambridge was ‘Laughing John’ Pringle (one of those ironic nicknames, like ‘Lofty’ for a very short man), and the department was thrown into a whirlwind of modernization. In one way after another, dear old Alister Hardy’s dear old department was ‘upgepringled’, no doubt for the better. Among the more exciting instances of upgepringleheit was the arrival from London of an equally dynamic group of X-ray crystallographers (think Watson/Crick, but with protein molecules instead of DNA). And, most exciting for me, they brought with them their own computer, which I was allowed, by its friendly custodian Dr Tony North, to use at night when it wasn’t needed for number-crunching the patterns of X-ray scattering by crystals. An Elliott 803, it was even more primitive by modern standards than the KDF9, but it had the enormous advantage that I was allowed to get my hands on it.
This was the time when I became fully aware of the addictive lure of computers. I really did literally – and frequently – spend all night in the warm, glowing computer room, entangled in a spaghetti of punched paper tape, which must have resembled my insomnia-tousled hair. The Elliott had the charming habit of beeping an acoustic rendering of its inner processing. You could listen to the progress of your computation through a small loudspeaker which hummed and hooted a rhythmic serenade, doubtless meaningful to Dr North’s expert ear but merely companionable to my nocturnal solitude. Nocturnal dalliance with computers in youth is characteristic of those – now called geeks – whose love affair with computers lasted longer (and more profitably) than mine: Bill Gates, to name one. With hindsight I cannot say that my affair with the Elliott was productive. No doubt I obtained some valuable practice in the art of programming. But Elliott Autocode was not a language that could be used on any other computers, and my nocturnal geekery, though diligent and very hard work, bore the same relation to serious programming as my tootling in the Oundle music school bore to real music.
I gave a talk on my Drive Threshold Model at the 1965 International Ethological Conference in Zurich. For the talk, I built a physical model of my theory, incorporating a rubber tube filled with mercury which I jiggled up and down to represent fluctuating ‘drive’. The rubber tube was attached to the bottom of a vertical glass tube, into which were let three electrical contacts at different depths, representing ‘thresholds’. Mercury is an electrical conductor, so when the jiggling column hit any of these contacts (the ‘drive’ exceeded the ‘threshold’) a circuit was completed. Obviously, if mercury was in contact with any electrode, it was automatically in contact with all lower electrodes too, which captured the major assumption of the model. I implemented the rules of the model by means of a noisy system of clattering electromechanical relays switching on coloured lights to represent pecks at different colours. The whole Heath Robinson50 affair was calculated to bring the house down, just as, at an earlier Ethological Conference in Oxford, a spoof hydraulic simulation devised by Desmond Morris, Aubrey Manning and friends reputedly had. How I managed to transport it from Oxford to Zurich evades my memory, and indeed my comprehension. There’s not a chance that today’s airport security would allow anything remotely like it through, bristling as it was with amateurishly soldered wires, relays, batteries and mercury.
Alas, just as I was about to go on the big stage for my first ever conference performance, something went wrong and my contraption wouldn’t work. In a sweat of panic and unable to think straight, I was kneeling on the floor outside the theatre, frantically tinkering, when I suddenly became aware of an amused Austrian accent barking out peremptory orders at great speed behind me. The rapid-fire, stuttering voice told me exactly what to do. As in a dream I obeyed – and it worked. I turned to look at my saviour, and beheld Wolfgang Schleidt, whom I hadn’t previously met although I knew who he was. Without any prior knowledge of what my infernal machine was supposed to do, this rising star of continental ethology had come upon my panic, instantly sized up the problem and dictated the solution to me. I have been grateful ever since to Dr Schleidt, who had, as I later learned without surprise, a reputation for technical ingenuity. I bore my strange device up into the theatre and at the end of my talk its spluttering coloured lights and air of Heath Robinson amateurishness received a reception little short of an ovation. Thank you, Wolfgang Schleidt: and for more than saving my blushes. For in the audience that day was the handsome figure of George Barlow, rising star of American ethology, and he was sufficiently impressed by my talk to get me invited to become an assistant professor at the University of California at Berkeley, without an interview or submission of a curriculum vitae: my first proper job.
But that was to come later. Meanwhile, back in Oxford, Niko Tinbergen had a sabbatical leave in 1966 and he invited me to take over the undergraduate lectures on animal behaviour for that year. He offered me his lecture notes, but I decided instead to develop my own lectures from scratch. Because this was the first course of lectures I ever gave, I wrote them out pretty fully. I thought I had lost them long ago but, to my surprise, they turned up in a cardboard box in the basement of my house while I was writing this memoir, and it was quite interesting for me to read them forty-six years on – especially the lecture on social behaviour, because it so clearly demonstrates both the central message and the style of The Selfish Gene, despite having been written a full ten years before that book.
In 1964, the Journal of Theoretical Biology had published two long and rather difficult mathematical papers by W. D. Hamilton, a young graduate student at the University of London whom none of us knew at the time, although he was later to become a close colleague. Mike Cullen characteristically recognized the importance of Hamilton’s papers before almost anyone else in the world except John Maynard Smith, and he expounded them one evening to the Bevington Road group. Mike’s enthusiasm was infectious, and I was immediately fired up, to the extent that I wanted to explain Hamilton’s ideas to the undergraduates in my stand-in lectures on animal behaviour.
Hamilton’s theory, now often labelled ‘kin selection’ (Maynard Smith’s name, not Hamilton’s own), follows directly from the neo-Darwinian ‘Modern Synthesis’ – directly, in the sense that kin selection is not an extra, not an addition bolted on to the neo-Darwinian synthesis: it is a necessary part of the synthesis. You cannot divorce kin selection from neo-Darwinism, any more than you can divorce the Pythagorean theorem from Euclidean geometry. A field biologist attempting to ‘test’ kin selection is in the same position as Pythagoras setting forth with a ruler seeking triangles to measure.
The neo-Darwinian synthesis, as opposed to Darwin’s own version of his theory, is centred on the gene as the unit of natural selection. Genes are discrete entities that can be counted in a population, more or less ignoring the fact that they are in fact sitting in the cells of organisms. Each gene has a frequency in the ‘gene pool’, which is approximated by the number of reproductive individuals possessing it. Successful genes are those whose frequency increases at the expense of unsuccessful alternatives, which decrease in frequency. Genes that cause an animal to be good at caring for its offspring tend to increase because they are carried in the bodies of the offspring cared for. Hamilton realized (as Fisher and Haldane had too, sort of, but they didn’t make much of it) that offspring are not the only category of relatives who share genes, and who therefore might be beneficiaries of evolved caring.
Hamilton derived a simple rule (now called Hamilton’s Rule): any gene ‘for’ altruism towards kin will tend to spread through the population, if the cost to the altruist C is less than the benefit B to the recipient devalued by the degree of relatedness r between the two. The degree of relatedness r is a proportion (i.e. a number between 0 and 1) which Hamilton showed how to calculate (its exact meaning is hard, though not impossible, to explain intuitively).51 Between full siblings, r is 0.5. Between uncles and nephews it is 0.25, between first cousins 0.125. Hamilton had a special interest in the social insects, and he made brilliant use of his theory of kin selection in explaining how ants, bees, wasps and (in a rather different way) termites evolved their remarkable habits of social altruism.
A typical underground nest of ants is a factory for propagating genes and spreading them around the countryside. The genes are churned out from the factory packaged up inside the winged bodies of young queens and males. These flying ants (which you might not realize are ants because of their unfamiliar wings) erupt from holes in the ground and fly up to mate on the wing. During her mating flight, each female (young queen) collects a lifetime’s supply of sperm, which she will store inside her body and eke out over the course of a long life. Laden with sperm, the mated female flies off and settles down to dig a hole and found a new nest. In some species, she bites or breaks off her wings, as she will need them no more in her role as subterranean queen.
Most of her offspring will be sterile workers, but the important children from the point of view of gene propagation are the young (winged) queens and males. Workers (all female in the case of ants, bees and wasps; male and female in the case of termites) normally have no prospect of passing on their genes through offspring, and devote their efforts to feeding and caring for their fertile collateral relatives, young queens and males, their siblings or nieces, for instance. A gene that makes a sterile worker care for a sister who is destined to become a queen can pass into future gene pools, carried there in the body of the young queen. The caring behaviour may never be expressed by the young queen herself, but the gene for that behaviour will be passed on to her worker daughters, who will consequently care for young queens and males who can pass it on.
The social insects are just a special case. Hamilton’s Rule applies to all animals and plants, whether or not they in practice care for relatives. If they don’t care for relatives, the reason will be that the economic costs and benefits in Hamilton’s Rule (the Bs and Cs) don’t pan out in such a way as to favour it, in spite of the fact that the coefficient of relatedness r may be high. And – a point that is often misunderstood, even by professional biologists – individuals care for their offspring for the same reason that elder siblings care for younger siblings (when they do): in both cases they share the genes for caring.

As I said, I was fired with enthusiasm when Mike Cullen introduced us to Hamilton’s brilliant ideas, and I very much wanted to try my hand at explaining them in my own way, in the lectures that I was to give as understudy for Niko Tinbergen. I was diffident about departing so far from Niko’s message and substituting my own rhetoric – about ‘selfish genes’ inhabiting a succession of mortal bodies, to be thrown away in the genes’ relentless march into the future. Seeking reassurance, I showed my typed-out lecture to Mike Cullen, and seeing again his handwritten marginalis today reminds me that I was hugely encouraged by it at the time (see the facsimile). It was Mike’s ‘lovely stuff’ that emboldened me to persist with my plan to lecture on this topic and in this style. And I suppose this could be said to be the moment of conception of The Selfish Gene, born ten years later. My lecture notes even contained the phrase ‘genes will be selfish’. I’ll return to this later when I come to the book itself.
In the summer of 1967, in the tiny Protestant church of Annestown on the south coast of Ireland, where her parents had a holiday cottage, I married Marian Stamp, a member of Niko Tinbergen’s group of graduate students, who later became his successor but one as Oxford’s Professor of Animal Behaviour and the world’s leading authority on the experimental science of animal welfare. I was by now committed to taking up the offer of an assistant professorship at the University of California at Berkeley. Niko was confident of Marian’s ability to continue her doctoral research there, with a minimum of long-distance supervision from him, and indeed his confidence was well justified. We had a brief honeymoon driving around Ireland in a hired car. Marian had to drive because I had forgotten my licence, and we had an awkward moment when the car hire clerk discovered she was a ‘graduate’ (it seems that graduates had a poor track record). Almost immediately after the honeymoon we set off for San Francisco, where we were met at the airport by the ever-kind George Barlow. A new life in the New World had begun.