COAST PATCH-NOSED SNAKE
Salvadora hexalepis virgultea Bogert 1935

Coast patch-nosed snake, San Diego County, California. Courtesy of John Andermann.

Status Summary
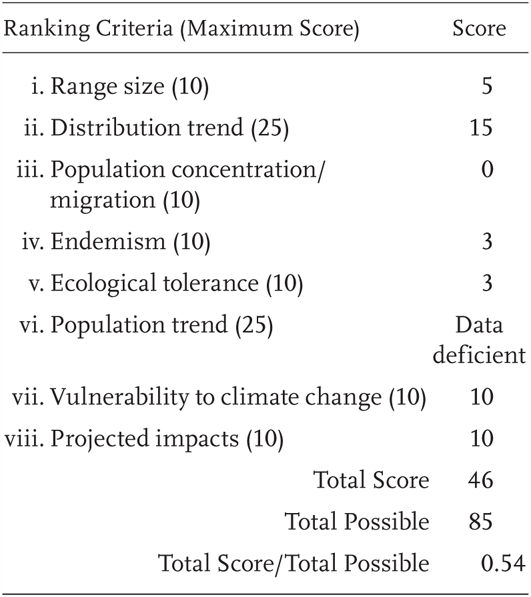
Salvadora hexalepis virgultea is a Priority 2 Species of Special Concern, receiving a Total Score/Total Possible of 54% (46/85). During the previous evaluation, it was also designated as a Species of Special Concern (Jennings and Hayes 1994a).
Coast Patch-Nosed Snake: Risk Factors
Identification
Salvadora hexalepis virgultea is a medium-sized (to 115 cm TL) snake, with an enlarged rostral scale, large eyes, and a light middorsal stripe 1.5–2 scale rows in width (Jennings and Hayes 1994a, Stebbins 2003). The dorsal stripe is yellowish to grayish and extends from the tail to the rear of the neck region (Bogert 1935, Stebbins 2003). The sides of the body are dark brown, and the head is olive or brown (Bogert 1935, Perkins 1938). Ventral coloration is generally a dull white, often with an orange wash that is more prominent toward the underside of the tail (Stebbins 2003, Lemm 2006). Usually only a single supralabial contacts the eye (Bogert 1935).
The presence of both a conspicuously enlarged rostral scale and a middorsal stripe easily differentiate this species from all other snakes within its range. Along the eastern edge of its range, S. h. virgultea is parapatric with the Mojave patch-nosed snake (S. h. mojavensis) and the desert patch-nosed snake (S. h. hexalepis), and it intergrades with both (Bogert 1945). In S. h. mojavensis, the supralabials usually do not reach the eye, and the dark lateral coloration is sometimes less pronounced and may be somewhat discontinuous. Salvadora hexalepis hexalepis usually has one supralabial reaching the eye but has a wider dorsal stripe (usually three scale rows wide) than S. h. virgultea. The spotted leaf-nosed snake (Phyllorhynchus decurtatus) occurs along the eastern edge of S. h. virgultea’s range and also has an enlarged rostral scale. This species differs in having a light-brown spotted pattern on the dorsal surface and attaining much smaller adult sizes (to 51 cm TL) (Stebbins 2003).
Taxonomic Relationships
Salvadora hexalepis virgultea was first recognized primarily on the basis of coloration, although some scalation characters also differentiate it from other subspecies of the S. hexalepis complex (Bogert 1935, Bogert 1945). Genetic analysis of differentiation among the subspecies has not been undertaken and represents a clear need for future research. The placement of Salvadora within the colubrine phylogeny is also uncertain and requires clarification.
Life History
Salvadora hexalepis virgultea is a medium-sized, active, diurnal snake, yet remains exceedingly poorly known. Here, we use information from California populations where we can, but largely rely on information from parts of the range outside of California as well as from other species and subspecies within Salvadora and assume that many aspects of the life history are similar among members of the complex. This subspecies is most active in May and June, and its normal seasonal activity period appears to extend from March until October. In Ventura County, warm (and presumably active) individuals have been found under stones between January and March (S. Sweet, pers. comm.). Individuals found in December and January in gopher burrows and woodrat nests were cold and presumably dormant (S. Sweet., pers. comm.). Specimens have been documented in the literature in all months except January and February (Klauber 1939, Grismer 2002), and more recent (1995–2011) USGS pitfall trap data have documented captures in January (three records) and February (one record; C. Rochester, unpublished data). Breeding has been observed in late April, with hatchlings appearing between mid-July and October (Klauber 1931, Grismer 2002), perhaps indicating that multiple clutches may be laid in a single year. Other members of the genus are oviparous, and we expect that S. h. virgultea is as well, although eggs have never been described (Wright and Wright 1957).
Salvadora primarily eats lizards and probably specializes on whiptails (Aspidoscelis spp.) (Cunningham 1959b, Grismer 2002, Lemm 2006). The diet of S. h. virgultea has not been described, but other members of the complex feed nearly exclusively on members of the genus Aspidoscelis, with other small lizards being taken only when they are moving rapidly, as Aspidoscelis tends to do (Bogert 1939). Several members of the complex are also known to feed on other small lizards, mammals, and snakes (Stebbins 2003). The enlarged rostral scale may be an aid for digging and possibly a specialization for feeding on lizard eggs (Grismer 2002, Lemm 2006), although this behavior has never been described in S. h. virgultea. Salvadora species have enlarged rear teeth, suggesting that they may envenomate prey. The saliva of Salvadora has not been studied, though Grismer (2002) observed an A. tigris that died soon after being bitten by a Baja California patch-nosed snake (S. h. klauberi).
The movement ecology of S. h. virgultea is unknown. Like other members of the genus, it is a fast and active species that is probably active at higher temperatures than most other diurnal colubrids (Jacobson and Whitford 1971). The species readily climbs and has been found foraging in brush, off the ground (Grinnell and Grinnell 1907, Lemm 2006; S. Sweet, pers. comm.). The species probably ranges widely, as do other large diurnal colubrids.
Habitat Requirements
Salvadora hexalepis virgultea shows an apparent preference for brushy chaparral habitat (Bogert 1935, Grismer 2002). Klauber (1939) located 35 specimens in “Heavy brush, Chaparral” habitat, which was more than the number found in all other habitat types combined. In Baja California, they are also often seen in riparian areas in the vicinity of Tecate (Grismer 2002). In California unpublished data suggest that this species has a preference for chamise and red shank and often basks at or near the tops of bushes (S. Sweet, pers. comm.). Schoenherr (1976) noted that this taxon was widespread but uncommon on the Pacific slope of the San Gabriel Mountains and that it may prefer coastal sage scrub to chaparral, at least in this area. He recorded a sighting at 830 m elevation, the highest site for coastal sage scrub habitat in the San Gabriels. At two burn sites in southern California, brush was reduced, Aspidoscelis numbers increased, and S. h. virgultea numbers remained stable or increased (C. Rochester, pers. comm.), suggesting that prey abundance may be at least as important as vegetation per se. If the species is a wide-ranging predator, it may be susceptible to habitat fragmentation. Because a large component of its diet probably consists of Aspidoscelis species, S. h. virgultea may be susceptible to decline in areas where Aspidoscelis are declining. The two species found within its range, the orange-throated whiptail (A. hyperythra) and the coastal whiptail (Aspidoscelis tigris stejnegeri), are both under threat. Aspidoscelis hyperythra is a Watch List species, while A. t. stejnegeri is a Priority 3 Species of Special Concern (see accounts, this volume).
Distribution (Past and Present)
The historical distribution ranges from Ventura and Los Angeles Counties south to the United States–Mexico border and south to the vicinity of El Rosario, Baja California, Mexico. It ranges from sea level along the coast up to 2130 m (Jennings and Hayes 1994a, Lemm 2006). The eastern edge of the range extends to the vicinity of Campo, San Diego County; Banning, Riverside County; and San Bernardino, San Bernardino County (Bogert 1935).
Today, the species is declining or absent from large areas of the Los Angeles basin and along the coast to San Diego (Jennings and Hayes 1994a; R. Fisher, pers. comm.). Survey data are available from Torrey Pines State Park where this species was formerly present. It has not been re-documented there despite intensive trapping efforts (Wells 1998).
Trends in Abundance
No historical or current abundance data exist for this taxon, and anecdotal reports are rare and inconclusive. Records in southern California are infrequent (Lemm 2006), although this taxon may be more commonly encountered in riparian areas south of the United States– Mexico border (Grismer 2002). The species may have historically been rare in California. Klauber (1924) reported only two specimens despite intensive collecting efforts spanning 2 years and stated that “it seems to be uncommon.” Bogert (1930) also noted that it was “uncommon in the chaparral.” USGS pitfall data for San Diego, Orange, and Los Angeles Counties collected between 1995 and 2011 indicate that 2.6% of snake captures were Salvadora hexalepis virgultea (123 captures out of 4680 total snake captures), compared to 3.8% (61/1601) of captures for daytime road driving records for snakes recorded by Klauber (1939). These data confirm that this taxon is relatively uncommon and may suggest a moderate decline over the last 60 years.
Nature and Degree of Threat
The declines in Salvadora hexalepis virgultea are most likely due to the conversion of the preferred brushy habitat to other vegetation types. Development of rangeland, combined with increasingly frequent and intense wildfires, has converted large blocks of chaparral habitat to grassland (R. Fisher, pers. comm.), which appears to be unsuitable for this species (Jennings and Hayes 1994a; S. Sweet, pers. comm.). Climate change may exacerbate the intensity of wildfires in southern California (Cayan et al. 2008b), although current models range from a 29% decrease to a 28% increase in wildfires in the region (PRBO 2011). However, climate models for 2070 project an estimated 38–44% decrease in the chaparral/coastal scrub habitat preferred by this species. Ongoing urbanization in the populated areas within this taxon’s range is also destroying, degrading, and fragmenting large areas of remaining habitat. Salvadora hexalepis virgultea’s probable preferred prey, Aspidoscelis lizards, are also in decline, which could cause cascading declines in snake populations. Finally, diurnally active widely foraging snakes are particularly affected by road mortality, and the volume of vehicular traffic in much of its range is large and increasing.
Status Determination
Salvadora hexalepis virgultea has a relatively small range in California and has disappeared from significant areas centered in the southern portion of its range where it was formerly known. It continues to lose habitat, which is also causing declines in a significant component of its prey-base. Habitat loss, due to direct anthropogenic changes, climate-change-driven habitat loss, and wildfire, is unlikely to stop in the near future, which we expect will cause further declines. For all of these reasons, a Priority 2 designation is justified.
Management Recommendations
The primary management goal for Salvadore hexalepis virgultea should be to protect large, intact patches of brushy chaparral and/or coastal sage scrub habitat that support this snake. Ideally, these patches should be those that are least likely to be directly affected by future climate change. Pending further study of the species’ movement ecology, habitat protection efforts should focus on remaining large blocks of intact habitat. Habitat fragmentation from roads is a key issue in these efforts. Road overcrossings, if installed for other taxa, may function as a means to avoid habitat fragmentation. A key element of effective management is to maintain large, healthy populations of Aspidoscelis hyperythra and Aspidoscelis tigris stejnegeri, since they appear to form the primary prey base of this snake. As declines in those species are also linked to habitat loss, management efforts among these species should be coordinated.
Monitoring, Research, and Survey Needs
Much remains to be learned about the life history and ecology of Salvadora hexalepis virgultea in California. Surveys employing pitfall traps, snake traps, and daytime surveys should be undertaken to establish baseline abundance data in remaining populations, and to ascertain whether these or other survey methods are potentially biased for this species, as has been suggested by some biologists. Because these snakes are infrequently encountered, power analyses are particularly important to determine how sensitive the surveys would be in detecting declines.
Autecological research focusing specifically on diet requirements, habitat utilization, and their interaction are badly needed, and the lack of this basic knowledge undermines our ability to effectively manage this snake. Specifically, the extent to which this taxon specializes on Aspidoscelis, as opposed to utilizing alternative prey, needs to be determined. Basic data on home range size and movement patterns, and how they may vary as a function of vegetation, are entirely lacking, and some anecdotal observations suggest that they may vary across the species’ range. These data are necessary to determine what habitat blocks are most important and how large they should be to effectively conserve this taxon.
Variation within the S. hexalepis complex has not been examined since the initial species description, and a genetic analysis has never been undertaken. A three-pronged genetic analysis is critical. First, a systematic characterization of the Salvadora species complex should be undertaken utilizing multiple independent nuclear markers. This will serve to clarify the reality of species and subspecies as valid evolutionary units and confirm their respective boundaries and range limits. Second, a phylogeographic study, using many independent markers, within S. h. virgultea is needed to identify large-scale management units within the taxon. Finally, landscape genetic studies that identify migration corridors between fragmented blocks of habitat will inform our understanding of movement ecology in this species. These would also provide an estimate of effective population sizes, augmenting ecological studies of census population sizes in protected and unprotected areas.