CAPÍTULO 33
La genética
Las leyes que gobiernan la herencia son desconocidas. Nadie puede decir por qué algunas características se heredan a veces sí y a veces no; por qué un individuo se parece en ocasiones a sus abuelos o a antepasados aún más remotos, ni por qué algunas peculiaridades se transmiten a los descendientes de ambos sexos y otras sólo a los de un sexo.
DARWIN
En el campo de las ciencias biológicas, el siglo XIX había sido testigo de una gran revolución. La gran teoría de la evolución de Darwin se había convertido finalmente, con mucho esfuerzo, en el eje de toda la biología, gracias a una ley sencilla pero apabullantemente sintética que daba cuenta de la multiplicidad de la naturaleza: la selección natural. Era una ley impresionante, por cierto, pero tenía un delicadísimo problema. Recuerden que la selección natural actuaba sobre las variaciones naturales que aparecían en cada generación de una especie, conservando las «buenas» (las adaptativas) y descartando las «malas» (las menos adaptativas), en una acción que se arrastraba lentamente a lo largo de las eras. Pero ocurría que nadie —con una sola excepción— tenía la menor idea de cómo funcionaban los mecanismos de la herencia.
Darwin, por ejemplo, creía que los caracteres de un individuo eran una especie de mezcla o promedio de los de sus progenitores, lo cual no dejaba de plantear un problema: ¿qué pasaba cuando emergía un rasgo adaptativo, pero el poseedor de ese rasgo se apareaba con individuos que no lo poseían? ¿Los rasgos nuevos no terminarían diluyéndose en el promedio general? Para salvar este inconveniente (que no era el único), los evolucionistas abandonaron en parte la idea de la selección natural actuando sobre individuos y pensaron en poblaciones portadoras del nuevo rasgo que se apareaban entre sí, perpetuándolo. Pero la solución era más bien oscura, porque era difícil admitir que aparecieran de la nada poblaciones enteras con un rasgo nuevo.
La falta de una teoría sobre la herencia constituía sin duda el flanco débil de la concepción darwiniana. Al no poder explicar el origen de la diversidad de rasgos que permitía la acción de la selección natural, ni la manera en que esos rasgos, una vez seleccionados, se mantenían sin perderse a través de las generaciones, nuestro buen amigo se vio obligado a emparchar una y otra vez la teoría de la evolución, introduciendo incluso dudosos elementos lamarckianos.
Y sin embargo, mientras Darwin se enfrentaba infructuosamente con el problema, la solución empezaba a gestarse en un monasterio austríaco, donde un contemporáneo suyo estaba a punto de dar con las reglas básicas del mecanismo de la herencia de las características físicas.
El tema, en realidad, y como siempre, era muy antiguo.
Breve historia del asunto
Porque obviamente, desde siempre, se supo que los hijos se parecen a los padres, a los abuelos, a los tíos o incluso a parientes más lejanos. Pero la humanidad vivió durante muchos siglos —durante la mayoría de su historia, en realidad— sin comprender los mecanismos de la herencia, a pesar de que usaba algunos conocimientos intuitivos de manera práctica. Por poner sólo un ejemplo, hace más o menos 7 mil años, en el período conocido como Neolítico, los hombres comenzaron a domesticar animales, a cultivar plantas y a efectuar cruzas eligiendo los ejemplares que resultaban más beneficiosos para obtener mejoras en la descendencia. No quiero decir con esto que nuestros ancestros neolíticos estuvieran al tanto de la genética, desde ya, pero lo que está claro es que al menos operaban de manera básica con ella, sin tener ni la menor idea (por lo menos, desde el punto de vista científico) de lo que estaban haciendo.
Durante mucho tiempo nadie se preguntó cómo funcionaba el mecanismo. Y cuando se empezó a reflexionar de manera más o menos sistemática sobre el asunto (es decir, cuando surgió la ciencia) se pensó que la clave de todo el asunto debería residir en la «semilla».
Se trata de una viejísima metáfora: el viejo Hipócrates sostenía que las diversas partes del cuerpo producían unas partículas específicas, o «semillas», que se transmitían a la descendencia en el momento de la concepción haciendo que ciertas partes de la progenie se asemejaran a esas mismas partes de los padres.
Aristóteles, por su parte, negó que el semen proviniera de todas las partes del cuerpo: de hecho, los hijos muchas veces se parecen más a sus abuelos, o sus bisabuelos, que a sus padres. Si era así: ¿de qué manera estos parientes lejanos podrían haber contribuido con las «semillas» de la carne y de la sangre que eran transmitidas de los padres a los hijos? Para resolver este enigma, postuló que el semen del macho estaba formado por ingredientes mezclados en forma imperfecta, algunos de los cuales habían sido heredados de generaciones pasadas. El semen se producía en el cuerpo masculino a partir de la sangre, y era un vehículo adecuado para transportar el pneuma, aquel omnipresente principio vital encargado de iniciar el desarrollo fetal gracias a un calor que estimulaba el crecimiento, del mismo modo que el Sol estimula el crecimiento de las plantas. Por su parte, la materia del cuerpo era aportada en forma exclusiva por el fluido menstrual de la mujer. El macho, en todas las especies, aportaba el principio vital o alma; la hembra cooperaba con la materia, además de hospedar al niño en su vientre y darle alimento. En la fecundación, el semen masculino se mezclaba con el fluido menstrual, que para él era el «semen femenino». En esa mezcla, el principio masculino le daba forma a la sustancia amorfa femenina, a partir de la cual se desarrollaba la progenie.
Galeno se diferenció de Aristóteles al intentar explicar por qué los hijos se parecen tanto a su madre y ancestros maternos como al padre y ascendientes paternos. Para él, ambos progenitores aportaban forma y materia al mismo tiempo, y ambos eran capaces de contribuir al desarrollo del embrión, lo que se conoció como «teoría de las dos semillas». Semillas que, dicho sea de paso, no tenían la misma jerarquía: dado que las mujeres eran más «frías» que los hombres y, por ello, más pequeñas, recubiertas con menos pelo, y con órganos reproductivos internos, su aporte era más débil; en los hombres, el calor interior hacía que los órganos reproductivos fueran externos y su semilla más poderosa.
Bastante después se planteó la oposición entre epigénesis y preformación, de la que ya hemos hablado en su momento, que se extendió a lo largo de varios siglos. Recuerden: la disputa era sobre si el individuo que iba a nacer ya estaba preformado en el semen masculino (o en el óvulo femenino) o si en verdad surgía de un tejido indiferenciado.
Los preformacionistas pensaban que el semen o los ovarios femeninos albergaban un hombre muy pequeño con todos sus órganos, y éste sólo necesitaba crecer en tamaño, lo cual, en los humanos, tenía lugar durante los nueve meses de embarazo. Se creía que sucedía lo mismo con todos los demás seres vivos; es más, estaban convencidos de que todos los organismos, los pasados, los presentes y los futuros, existían desde el primer acto creador y estaban a la espera de ser activados mediante la fecundación.
William Harvey, el descubridor de la circulación de la sangre, fue el representante más destacado del epigenetismo, la doctrina que negaba que el organismo estuviera preformado y recuperaba las ideas de Hipócrates y de Galeno de que todas las partes del cuerpo contribuían al desarrollo del futuro ser. Lo que sostenía esta teoría es que el embrión se formaba a partir de una mezcla más o menos homogénea de sustancias materna y paterna; a lo largo del desarrollo, esa materia se iba diferenciando progresivamente y los distintos órganos iban tomando forma. Harvey identificó al huevo como el rudimento de la vida («ex ovo omnia», «todo viene del huevo») y explicó la reproducción sexual en términos similares a lo que sucede en los animales ovíparos, como las aves y los reptiles. Pero no pudo responder la pregunta sobre el origen último del huevo en los organismos vivíparos, es decir, los que se desarrollan en el vientre materno (lo cual es bastante lógico: el descubrimiento de los ovocitos en los ovarios sería realizado dos siglos después, en 1827, por el biólogo alemán Karl Ernst von Baer).
En el siglo XVIII, la teoría preformacionista tenía una buena cantidad de adeptos, pero no tardarían en aparecer voces de discordancia. En 1745, el físico y astrónomo francés Pierre Louis Moreau de Maupertius (1698-1759), a través de su libro Venus Physique, se propuso desbaratar la hipótesis de la preformación y afirmó que, en la herencia, el padre y la madre realizan un aporte equivalente, determinado por el más ciego azar. El conde de Buffon, por su parte, postuló la existencia de lo que denominó «moléculas orgánicas», unidades primitivas que darían lugar a todos los seres vivos uniéndose a lo largo del desarrollo del embrión y que, a manera de un molde, mantenían las características de cada especie a través de las generaciones.
El siglo XIX heredó este panorama en el que nada estaba muy claro y en el que las dos hipótesis convivían. El mismísimo Darwin, que para completar su teoría de la evolución necesitaba una explicación coherente acerca del mecanismo mediante el cual las características de una generación pasan a la siguiente, postuló una hipótesis propia heredera de las ideas de Hipócrates, Galeno y Harvey. Efectuó la primera presentación de su conjetura en 1868 (casi diez años después de la publicación de El origen de las especies), en el capítulo final de un libro sobre la variación en animales y plantas bajo la domesticación. A su teoría le dio el nombre de «pangénesis», término que, en virtud del prefijo griego «pan», indicaba que todas las células del cuerpo contribuían a la herencia a través de unas partículas que bautizó como «gémulas», que eran capaces de sufrir y cambiar al compás del medio ambiente, y que se almacenarían en los óvulos o en los espermatozoides. Esta hipótesis proponía que cada órgano y estructura del cuerpo producía gémulas, que llegaban a los gametos a través de la sangre.
Lo cierto es que todos tanteaban en un terreno resbaladizo y oscuro, y Darwin seguía sin una buena explicación para aclarar el mecanismo de la herencia.
Las arvejas dan la clave
Si Darwin hubiera leído este libro, se habría enterado de que en esos mismos momentos, mientras él especulaba con inexistentes gémulas, un monje llamado Gregor Mendel (1822-1884), en medio del silencio de un monasterio austríaco, publicaba buena parte de la solución a este problema. Pero prácticamente nadie lo leería hasta el siglo siguiente, cuando comenzara a desarrollarse la teoría genética propiamente dicha.
Así eran las cosas y así son muchas veces: aislado de todos los científicos de su época, un clérigo desconocido elaboraba las respuestas que necesitaba Darwin y que, dicho sea de paso, no dependían del desarrollo de grandes tecnologías (si se excluye la compleja y mal repartida tecnología de la paciencia).
Mendel nació en un pequeño pueblo llamado Heinzendorf, en la región de Moravia, que en aquellos tiempos pertenecía al Imperio Austro-húngaro (y hoy forma parte de la República Checa). Dado que sus padres eran campesinos pobres, la idea de realizar estudios universitarios parecía ciertamente utópica, hasta que vino la ayuda en la forma de unos dinerillos que le prestó su hermana menor y que le permitieron asistir durante dos años al Instituto Filosófico de la ciudad de Olmütz (actualmente Olomouc, República Checa). La vida no era fácil allí para un estudiante sin recursos, por lo cual optó por la única salida viable para alguien que en aquella época buscara una educación pero no tuviera el dinero para solventarla: con el nombre de Gregor, entró como fraile en un convento agustino en la ciudad checa de Brno (entonces Brünn), en el centro del Imperio Austro-húngaro.
Dado que, a diferencia de otras órdenes, los agustinos daban mucha importancia a la enseñanza y a la investigación, Gregor pudo estudiar todas las disciplinas que lo entusiasmaban y —lo más importante para nuestra historia— tuvo libre acceso al invernadero. Allí realizaría, entre 1854 y 1862, sus famosos experimentos sobre la reproducción y la herencia, cuyos resultados presentó tres años después en las reuniones de la Sociedad de Historia Natural de Brünn.
En 1866 los publicó en las actas de la Sociedad con el título de «Experimentos sobre hibridación de plantas», y ciertamente hubieran generado una revolución impresionante en el campo de la biología. Sin embargo, la publicación fue ignorada por completo. Sólo se captaría su relevancia en la primavera de 1900, treinta y cinco años después de la publicación, y dieciséis años luego de su muerte.
Nada de lo que vio Mendel hubiera sido posible sin la ayuda de las generosas arvejas (o guisantes), plantas útiles para el experimento, ya que son fáciles de cultivar y se hacen adultas en una estación. Tienen además la particularidad de ser autopolinizantes, es decir, contienen todo lo necesario para su autorreproducción, sin necesidad de interactuar con otras.
Algo le llamaba la atención a nuestro monje en la homogeneidad entre sus arvejas, porque comenzó a experimentar abriendo las flores antes de que se desarrollaran del todo para quitarles el polen propio e intercambiarlo con el de otra flor seleccionada para sus experimentos. De esta manera forzaba su cruzamiento.
La historia es que reunió 34 cepas de arvejas de toda Europa y afinó sus selecciones por medio de cruces forzados hasta obtener líneas o variedades puras, es decir, que durante generaciones venían dando una característica particular sin variaciones: altas, bajas, verdes, amarillas, gruesas, delgadas. Centrémonos en el color. Gregor, gracias a sus entrecruzamientos, había llegado a tener dos cepas de características puras: unas eran completamente verdes y daban siempre arvejas verdes; otras eran completamente amarillas y daban siempre arvejas amarillas. En uno de sus experimentos cruciales, mezcló plantas de semillas que habían sido exclusivamente amarillas con otras similares, pero verdes. El resultado fue una generación (llamémosla G1) de plantas totalmente amarillas. Pero cuando mezcló las plantas de esta nueva generación entre sí, observó con sorpresa que en la segunda generación (G2) aparecían plantas con flores verdes en una proporción de 1 a 3. Obviamente, el carácter «verde» había permanecido oculto y latente en la generación amarilla (G1).
La paciencia de Mendel era realmente monástica: después de generaciones de experimentación obtuvo una muestra de 8.023 plantas que dieron una proporción de 6.022 amarillas y 2.001 verdes, y después de realizar muchas observaciones más, sacó algunas conclusiones, que van a resultar más claras si miran las siguientes figuras:
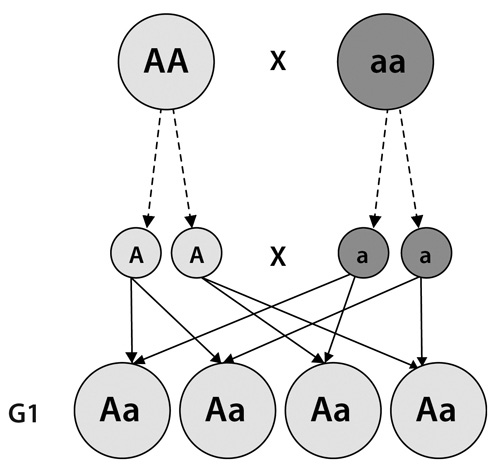
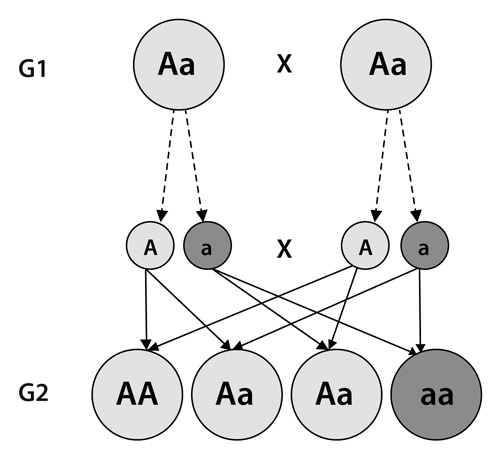
• En primer lugar, había una especie de «unidades de herencia», capaces de transportar los rasgos de una generación a otra, que Mendel llamó «factores hereditarios» y que se presentaban de a pares: los padres contribuyen cada uno con uno.
• Hay algunos «factores hereditarios» dominantes y otros recesivos. Cuando ambos rasgos heredados indican que el descendiente sea verde, la planta será verde, y si ambos indican que sea amarilla, será amarilla. Pero ¿qué pasa si uno indica verde y el otro amarillo? Mendel comprobó que en estos casos la planta sería amarilla porque este color, de alguna manera, era más «fuerte» o «dominante» por así decirlo, que el verde, que es recesivo.
• Los factores recesivos no desaparecen, sino que quedan ahí, y pueden ser transmitidos a la descendencia. Y así estos ejemplares, que tenían un rasgo de cada tipo heredado de sus progenitores, pasarían en partes iguales los rasgos verdes y amarillos a su descendencia, lo que permitiría que éstos tuvieran en un caso los dos genes dominantes (como se llama a los más «fuertes»), en otro los dos recesivos (los «débiles») y en otros una mezcla. Sólo en el que tienen los dos recesivos la planta sería verde.
Mendel había llegado a este secreto de la naturaleza sólo por la estadística: sin microscopios, ni conocimientos de una ciencia genética que simplemente no existía. En los años siguientes extendió sus experimentos a otros fenotipos (rasgos observables), como plantas con semillas redondas o arrugadas, y elaboró toda una serie de «Leyes descubiertas por los guisantes».
Pero aunque el descubrimiento era extraordinario, nadie parecía notarlo. Para colmo se frustró en sus siguientes experimentos, en los que sus teorías no pudieron avanzar. Había enviado su artículo a muchos naturalistas importantes, pero sólo obtuvo respuesta de Karl Naegeli (1817-1891), un famoso botánico de la época, que se mostraba escéptico y no podía entender cómo un híbrido Aa transmitía el carácter A y perdía el recesivo, ya que defendía la idea de la herencia por mezcla, por completo contraria a los resultados de Mendel. Naegeli le recomendó experimentar con otra planta llamada «diente de león». Por desgracia, Mendel no encontró rastros de sus leyes al experimentar con estas plantas que, como se descubrió años más tarde, se reproducen sin fertilización ni entrecruzamiento de distintos ejemplares, con lo cual cada descendiente es un clon de la planta madre. Además, en 1868 fue elegido abad de su monasterio y quedó absorbido por las tareas administrativas. Murió en enero de 1884, a los 63 años, dos años después que Darwin, quien nunca supo de sus trabajos.
Poco después de su muerte, en el patio del monasterio, en el lugar que había ocupado el invernadero, se hizo una hoguera con todos sus papeles personales y científicos. Es posible que el nuevo abad, por pura competencia con su predecesor, quisiera eliminar todo recuerdo de su antecesor, unos papeles en los que nadie parecía estar interesado.
Las tres leyes
Las conclusiones de Mendel dieron lugar a lo que se conoce como leyes de la herencia, principios que funcionan de manera general en todas las especies.
La primera ley, de la «uniformidad de los híbridos de la primera generación», afirma que cuando se cruzan dos individuos de dos variedades puras de la misma especie, que difieren en el aspecto que presenta un mismo carácter (por ejemplo, semillas amarillas versus verdes), los descendientes serán iguales entre sí e iguales a uno de los progenitores, el que aporta el factor dominante (semillas amarillas).
La segunda ley es la de la «segregación de los caracteres» y demuestra que los factores hereditarios constituyen unidades independientes, que pasan de una generación a otra sin sufrir alteración alguna. Si se cruzan entre sí los descendientes de la primera generación (a partir de dos líneas puras), el carácter recesivo transmitido por uno de los progenitores (semilla verde), que no se manifestaba, se hace evidente en una cuarta parte, y el dominante en las tres cuartas partes de los descendientes.
La tercera ley, de la «independencia de los caracteres», afirma — simplificando muchísimo— que cada carácter se hereda en forma independiente de los demás.
Las tres leyes de Mendel permiten explicar y predecir cómo serán los rasgos físicos (fenotipo) de un nuevo individuo. Ahora bien, sólo la segunda y la tercera se refieren a cómo se transmiten los caracteres a la descendencia. La primera, en cambio, no trata acerca de la transmisión, sino de la expresión de los caracteres.
Suele afirmarse que, si Darwin hubiera conocido el informe de Mendel, habría podido pensar en un mecanismo para la selección natural y su teoría habría quedado mejor respaldada frente a los adversarios. Sin embargo, quienes se oponían a esas ideas estaban menos preocupados por el mecanismo de la selección natural que por el hecho mismo de que esa selección hubiera acontecido: la teoría darwiniana atacaba las creencias más básicas acerca del lugar del hombre en el universo y del papel de cualquier dios en la creación.
Una sustancia cualquiera
Para que en algún momento se llegara a la formidable síntesis que fue la genética, primero fue necesaria otra síntesis de la que ya hablamos hace algunos capítulos: la teoría celular, mediante la cual la biología alcanzó su tan ansiada unidad fundamental.
Gracias a la formulación de la célula como unidad de la vida (es decir, gracias a un concepto teórico) y al mejoramiento de los microscopios (es decir, gracias a un progreso observacional), comenzaron a verse cosas que nunca antes se habían visto o, si se habían visto, nunca se habían podido interpretar. En 1869, Johann Friedrich Miescher (1844-1895), que no sólo era suizo sino también químico y trabajaba en un hospital de Tubinga, al sudoeste de Alemania, al que llegaban heridos de la Guerra de Crimea, empezó a estudiar la secreción que se juntaba en los vendajes. Rompió con enzimas digestivas la membrana celular de los glóbulos blancos en el pus de las heridas y encontró una sustancia que contenía fósforo y nitrógeno, hecha de moléculas en apariencia muy grandes y que se encontraban en el interior del núcleo de los glóbulos blancos. La llamó «nucleína» y, como una década más tarde se comprobó que tenía propiedades ácidas, se la redesignó como «ácido nucleico».
O mejor dicho, «ácidos nucleicos», porque había de dos tipos: uno que contenía el azúcar ribosa y el otro el azúcar desoxirribosa. Y por lo tanto, se llamaron ácido ribonucleico (ARN) y ácido desoxirribonucleico (ADN). Miescher no abandonó sus ácidos y en 1889 aisló por primera vez el ADN del esperma de un salmón. Eran moléculas grandes, pesadas, gomosas, molestas e incluso poco simpáticas, con una indescifrable estructura y que, en principio, no servían para nada. ¿A quién podían interesarle dos aciduchos que parecían estar ahí sin ningún motivo?
Había, sin embargo, algo que resultaba curioso: la cantidad de ADN en los espermatozoides era la mitad del que había en las células normales. Cosa rara, es cierto, pero nada más para una época que todavía no estaba lista para interpretar la importancia del hecho.
En 1880, un biólogo alemán llamado Walter Flemming (1843-1905), trabajando con las células, utilizó tintura roja y vio que se adhería a unas tiritas que aparecían en el núcleo y a las que, justamente por su capacidad de recibir el color, llamó «cromatina» (por chromo, color en griego). Y que jugaban, en apariencia, algún papel importante en el proceso de la división celular: la cromatina, observó, se agrupaba en forma de filamentos que emigraban a ambos lados de la célula mientras ésta se estrechaba por el medio y se dividía en dos —eso al menos era lo que se veía en el microscopio—. Ocho años más tarde, el biólogo alemán Wilhelm von Waldeyer (1836-1921) llamó a los filamentos «cromosomas» y enseguida se pudo comprobar que cada especie tenía un número fijo y específico de esa especie (por ejemplo, el hombre tiene 23 pares, es decir, 46 cromosomas). Pero además —¡oh sorpresa!— resultaba que en las células germinales había exactamente la mitad.
A principios del siglo XX, el rompecabezas se estaba armando. Pero, por el momento, todo permanecía en el estrecho ámbito de lo puramente observacional.
Resucitan las leyes de Mendel
Los cada vez más detallados estudios empíricos sobre las células confluyeron con la resurrección de las investigaciones de Mendel por parte de tres hombres al mismo tiempo, para trazar el sinuoso camino de la genética.
En los últimos años del siglo XIX, un botánico holandés llamado Hugo de Vries (1848-1935) llevaba cerca de veinte años cruzando un tipo especial de flores y separando a aquellas que presentaban alguna particularidad llamativa. Después de algunas generaciones, estas últimas habían sufrido lo que él llamó «mutaciones» y resultaban imposibles de cruzar con las originarias. Era lo que había imaginado Darwin, aunque sin poder demostrarlo experimentalmente.
Sus conclusiones se publicaron sobre el final del siglo en un libro que cayó en manos de Karl Correns (1864-1933), quien había comenzado a experimentar con arvejas y había llegado a conclusiones similares a las de Mendel. Al revisar los archivos de su maestro Karl Naegeli, cuyo papel en esta historia les conté un poco más arriba, se topó con la correspondencia que Mendel le había enviado, y cuando leyó la obra de De Vries rápidamente encontró una similitud… una enorme similitud con los resultados de nuestro monje, a tal punto que acusó a De Vries de plagio.
El susodicho tuvo que «aceptar» que había leído a Mendel poco antes de publicar su libro y que no lo había citado porque «no se había dado cuenta», aunque aseguró que había llegado a las conclusiones por sí mismo. Haya sido como haya sido, en las siguientes publicaciones reconoció explícitamente la primacía del abad.
El tercer hombre fue el botánico austríaco Erich von Tschermak (1871-1962). También él realizó experimentos con arvejas y también él llegó a las mismas conclusiones que su compatriota había registrado unas décadas antes. Mientras escribía los resultados de sus experimentos, Tschermak encontró una referencia al trabajo de Mendel que lo llevó a buscar la obra y rápidamente descubrió que el abad no sólo había llegado a los mismos resultados que él, sino que lo había superado. En 1900, los tres investigadores publicaron una suerte de reivindicación de la obra del monje.
Las leyes de Mendel experimentaron una verdadera resurrección. Pero, así y todo, seguían sin dar ninguna respuesta (y ni siquiera una pista) acerca de la naturaleza de lo que Mendel había llamado «factores hereditarios». El principio organizador ya estaba; ahora el asunto era avanzar para describir esos paquetitos de información que hacían que una arveja fuera verde o amarilla, que un pájaro tuviera el pico de tal o cual forma o que un chico fuera rubio o morocho, y no hubiera siempre una regresión al promedio, como temía Darwin.
Todo apuntaba a los cromosomas: al fin y al cabo, como los factores hereditarios, aparecían de a pares. En 1902, el estadounidense Walter Sutton (1877-1916), cuando aún era un estudiante, escribió un par de trabajos que daban la demostración más temprana de cómo es que los cromosomas funcionan dividiéndose y volviéndose a juntar en un individuo nuevo y sugirió que este mecanismo podía dar cuenta de las leyes mendelianas. Pero había una objeción casi de sentido común: los cromosomas eran muy pocos (23 pares, como recordarán), y, si verdaderamente transportaban los factores hereditarios de los rasgos, tenían que estar sobrecargadísimos.
«Factores hereditarios»: la terminología empezaba a resultar pesada. El 31 de julio de 1906, durante la 3ª Conferencia de Hibridación y Reproducción de Plantas, se aceptó la palabra «genética» (de la palabra griega genos, que significa, entre otras cosas, «generación») como nombre para el estudio de los fenómenos de la herencia y la variación y, en 1909, Wilhelm Johannsen (1857-1927), un fisiólogo vegetal, decidió que los factores hereditarios deberían recibir un nuevo nombre y los llamó «genes».
Aparecen candidatos firmes
Todos corrían la carrera en busca de esos «genes» hasta entonces invisibles. Y la cosa transcurría con cierta vertiginosidad. Hacia el año 1908, Thomas Morgan (1866-1945) instauró un nuevo método de experimentación con moscas del vinagre, como se conoce habitualmente a la Drosophila melanogaster, bicho que tiene la ventaja (para los investigadores, por supuesto, no para ellas) de hacer todo su ciclo vital —desarrollo, reproducción y muerte— en dos semanas. Era una gran mejora cronológica frente a las arvejas, con las cuales conseguir una nueva generación llevaba meses.
Morgan estaba seguro, como tantos otros, de que los cromosomas tenían mucho que ver con las leyes de la herencia, pero era obvio que no podían ser ellos mismos los factores hereditarios, sino que, en todo caso, debían transportar gigantescos paquetes de esos factores. A esos segmentos de los cromosomas que efectivamente estaban encargados de almacenar la información hereditaria los llamó, siguiendo la terminología de Johannsen, genes.
Herman Müller (1890-1967), biólogo estadounidense, se dedicaba mientras tanto a preguntarse cómo es que si sólo se heredan rasgos paternos y maternos, cada tanto pueden aparecer mutaciones que no tienen antecedentes en la especie. En 1923, tras numerosos experimentos en los que irradiaba los cromosomas con rayos X, concluyó que las mutaciones que se observan habitualmente son producto de una alteración aleatoria en los genes, cuyas consecuencias no se pueden pronosticar (al menos hasta ese momento).
Por otra parte, entre 1926 y 1930, James Sumner (1887-1955) y John Northrop (1891-1987) explicaron que todas las enzimas responden a las leyes de la química y que no son otra cosa que proteínas que permiten la producción de otras proteínas con distintas funciones para la vida.
Las proteínas, una suerte de comodín biológico, eran consideradas por todo el mundo como las mejores candidatas para ser los genes, los mensajeros de la herencia. Frederick Griffith (1881-1941) llegó a esa conclusión al intentar aislar el principio gracias al cual la información de las células puede intercambiarse, y lo cierto es que si en ese momento hubiera habido una elección, las proteínas hubieran ganado por goleada.
Y entonces vino un alumno de Griffith, el canadiense Oswald Avery (1877-1955), y mandó a parar. Con la intención de probar, como lo habían hecho Sumner y Northrop con las enzimas, que los genes no eran más que proteínas, hizo el experimento decisivo: en 1944 aisló extractos de células y comenzó a separar proteínas, lípidos, ácidos nucleicos y otros a fin de ver cuál era el principio fundamental que permitía seguir reproduciéndose a la bacteria que estudiaba. Trabajaba con neumococos, los causantes de la neumonía. Y aquí viene el asunto, porque había dos cepas: una era suave, rodeada por una cápsula protectora; la otra, rugosa, sin cápsula. Por alguna razón, las bacterias rugosas no podían sintetizar la cápsula. Avery le agregó a la cepa rugosa un extracto de la suave; instantáneamente las rugosas empezaban a fabricar la cápsula. Obviamente, el extracto transportaba las instrucciones para fabricarla, es decir, conducía la información genética que le indicaba a la célula rugosa cómo encapsularse. Ahí tenían que estar las ansiadas proteínas que, supuestamente, acarreaban la información…
Era un momento vital para el desarrollo de la teoría biológica, uno de esos momentos destinados a marcar un antes y un después en el ámbito científico, y es lógico que Avery se pusiera a buscar con afán y ansiedad dentro de ese extracto que daba la pauta para fabricar la cápsula. El problema es que, por más que hurgó… ¡no había allí ninguna proteína! Había, eso sí, ácidos nucleicos. La conclusión estaba a la vista: no eran las proteínas sino las viejas y queridas moléculas de Miescher, el ARN y el ADN, esas inútiles, los vehículos de la información hereditaria.
Empezaba una nueva época.
El asalto final: la doble hélice
Sabiendo que el secreto de la herencia se encerraba en el ARN y el ADN, en 1953 el estadounidense James Watson (n. 1928) y el británico Francis Crick (1916-2004) que llegarían a ser dos de los científicos más reconocidos del siglo XX, se prepararon para el asalto final: detectar la estructura del ADN.
El modelo que finalmente fabricaron indicaba una estructura en espiral de dos tiras; cada tira es una larga cadena de fosfatos de los cuales cuelgan azúcares y bases: cada par azúcar-base es un nucleótido. Hay sólo cuatro nucleótidos diferentes, según la base que contengan (el azúcar es siempre el mismo). Las dos tiras de la doble hélice, por su parte, están unidas por hidrógenos. Cuando llega el momento de la reproducción, ambas tiras se separan y cada una fabrica una réplica de sí misma.
Watson y Crick creyeron que las tiras principales estaban separadas y que cada una servía como molde para la formación de otras moléculas. Este proceso complejo de copiado explicaba la replicación de los genes y, eventualmente, de todo el cromosoma.
Las cadenas de ADN tienen la forma de una escalera enroscada. Si tomáramos todas las cadenas que se encuentran en una sola célula humana y las uniéramos, obtendríamos un cordón de cerca de dos metros de largo y de 20 mil millonésimas de centímetros de ancho.
Los laterales están hechos de grupos de azúcar y fosfato alternados. Los «escalones» están formados por nucleótidos (bases nitrogenadas) de Adenina, Guanina, Citosina o Timina, a las que, para facilitar, se llama A, G, C y T. Por sus características químicas, la A sólo se puede unir con la T y la G con la C, lo cual no deja tantas opciones de combinaciones.
Toda la información hereditaria, que especifica las características de un individuo, está contenida en los genes y los genes no son sino trozos de ácido desoxirribonucleico (ADN). Lo impresionante es que con apenas cuatro nucleótidos, como si fueran las letras de un alfabeto mínimo, se escriben todas las instrucciones necesarias para la materia viviente, desde los virus a los elefantes, en secuencias como ATGTGAGGGGG, que especifican la forma en que cada célula fabrica proteínas.
La longitud de los genes es variable, según la especie y la función: puede ir de unos pocos cientos a varios miles de nucleótidos seguidos, que se aprietan en mucho menos que una milésima de milímetro. Una señal (una cierta combinación de nucleótidos) en la hebra indica que el gen comienza; una segunda señal anuncia que empieza el mensaje genético (es decir, la sucesión de nucleótidos relevante para la fabricación de una determinada proteína), hasta que una tercera señal anuncia el fin del mensaje. Una última señal informa a quien corresponda que allí termina el gen.
Puede parecer increíble, pero en esas secuencias está la información que permite que se forme un ojo en toda su complejidad, que se produzcan jugos gástricos o que indica la forma en que una neurona debe contactarse con otras para, por ejemplo, leer estas líneas. Es un recetario para crear proteínas que se combinan en una complejidad pasmosa. Es mucha información y cada célula la tiene absolutamente toda porque, de hecho, nuestro cuerpo entero proviene de una sola célula, formada por la unión de un óvulo y un espermatozoide que sufrió millones de divisiones celulares.
El trabajo de Watson y Crick no fue un final sino un comienzo: con el tiempo se fue revelando la gran cuestión pendiente, que consistía en determinar de qué manera el orden de las unidades que componen el ADN (recuerden: ATCG) podía especificar la secuencia de aminoácidos en una proteína. El mecanismo hallado asombra por lo maravilloso. Dicho de manera simplísima: el ADN o, mejor, una parte del ADN correspondiente a uno o varios genes, se copia en una molécula de ácido ribonucleico (ARN), que se llama ARN mensajero. Este ARN mensajero sale del núcleo y es llevado a los ribosomas, unos pequeños orgánulos que se encuentran en el citoplasma de la célula, donde es «leído», y se ensamblan los aminoácidos que formarán las proteínas.
Reproducción y evolución: la teoría sintética
Recién con el desarrollo de la teoría cromosómica, que explica la herencia y la genética en general, se elaboró en la década del treinta la gran síntesis neodarwiniana que dio al evolucionismo por selección natural una solidez absolutamente indiscutible.
La fundación de la genética echó una nueva luz sobre el darwinismo: si los caracteres pasan de generación en generación mediante unidades de herencia discretas, como los genes, el punto oscuro de la teoría de Darwin quedaba aclarado. Los genes sufren cada tanto leves cambios al azar (mutaciones), que implican pequeñas modificaciones de los rasgos que transportan. Cuando se produce la mezcla de portadores de caracteres «buenos» y «malos» (con todas las salvedades del caso: recuerden que no hay caracteres buenos o malos de manera absoluta, sino que dependen siempre del contexto), éstos no se mezclan en la descendencia, sino que aparecen puros en el nuevo individuo. La selección natural desecha a los portadores de genes con modificaciones «malas» y conserva a los que tienen genes «buenos», expandiendo, de esta manera, estos últimos genes, que sí se reproducen. Así, con la guía aportada por la genética —y no sin duro trabajo—, entre 1930 y 1940 se elaboró la Teoría Sintética (o síntesis neodarwiniana), que nuevamente dio una explicación acabada de la evolución natural mediante los mecanismos mendelianos de la herencia y que uno de sus notorios constructores, Theodosius Dobzhansky (1900-1975), resumió así: «Evolución es un cambio en la composición genética de las poblaciones». Con la nueva síntesis, otra vez pareció que el problema de la evolución estaba terminado y que los problemas restantes serían solucionados por la nueva genética de poblaciones.
Intervalo siniestro: el caso Lysenko
Al compás del acordeón
canto en honor del gran Lysenko.
Gracias a sus enseñanzas
sembramos y recogemos el trigo
y hemos echado al mal tiempo de los campos
trabajando duro en ellos.
Todo miembro de las granjas colectivas
está orgulloso del gran Lysenko
que enseña a todo el país
cómo cultivar un jardín.
Hay una fiesta en la naturaleza
que cruza todo el país
y hará fructificar
la vida de la Unión Soviética.
Al compás del acordeón
canto en honor del gran Lysenko.
«El canto del koljós», letra de A. SALNIKOVA, música de K. MASALITAINOVA, canción de los tiempos de Stalin en honor de Lysenko
Ésta es una historia trágica. De la misma manera que el juicio a Galileo en el siglo XVII, el «caso Lysenko» muestra en el XX las nefastas y explosivas consecuencias de la mezcla entre ciencia, política e intolerancia. En febrero de 1935, en plena época estalinista, y durante el Segundo Congreso Soviético de Granjas Colectivas de la URSS, un tal Trófim Denisovich Lysenko habló y denunció a los genetistas que trabajaban científicamente y con rigor mendeliano diciendo que eran «enemigos del pueblo». El mismísimo Stalin, que estaba presente, interrumpió el discurso y gritó: «¡Bravo, camarada Lysenko, bravo!». Fue suficiente. A partir de ese momento, Lysenko inició un ascenso meteórico que le permitió transformarse durante treinta años en el dictador de la biología soviética. Y un dictador nada blando, por cierto.
¿Quién era Lysenko? Un biólogo del montón, que en los años veinte se había interesado por la adaptación de ciertas variedades de plantas a climas rigurosos. En diversos artículos aseguró haber obtenido fabulosos rendimientos por hectárea, y reclamó que se extendieran sus «nuevos métodos» al conjunto de la agricultura, ante el escepticismo de los biólogos soviéticos, que cuestionaban el rigor de su metodología, la veracidad de sus cifras, o la temeraria afirmación de que un experimento a escala reducida pudiera generalizarse a un territorio tan vasto y complejo como el de la URSS. Otros, simplemente, lo ignoraban.
Pero Lysenko, entre tanto, no se quedó con el resultado de sus «experimentos». Entre 1931 y 1934 elaboró una «teoría» de la herencia que negaba todos los principios de la genética mendeliana. Según Lysenko, los genes no existían y la transmisión hereditaria era una propiedad general interna de la materia viva, que no necesita de ningún mecanismo especial. La intención era clara: volver a la ya refutada teoría lamarckiana según la cual los caracteres adquiridos por adaptación al medio ambiente pueden ser transmitidos a la descendencia.
La ciencia soviética reaccionó, y con una catarata de trabajos refutó las tesis de Lysenko. Lo cual no hubiera sido más que una mera polémica entre tantas, si se recuerda que en aquellos años la genética, aunque ya bien establecida, no tenía aún el formidable aparato de la biología molecular para respaldarla. Pero en el medio se impuso el apoyo explícito de Stalin a las «innovaciones» de don Lysenko, y una simple controversia académica se convirtió en una sangrienta tragedia.
En 1937, una revista dirigida por Lysenko acusó con nombre y apellido a muchos genetistas de bujarinistas y trotskistas (Bujarin acababa de ser ejecutado poco antes). En febrero de 1938, Lysenko fue nombrado presidente de la Academia Nacional de Ciencias Agrícolas. Su poder creció ininterrumpidamente con el apoyo explícito de Stalin, que era su ardiente partidario, a pesar de los fracasos que la aplicación de las tesis lysenkianas producía una y otra vez en la agricultura y que eran sistemáticamente atribuidos a saboteadores, con la consiguiente ola de denuncias y arrestos.
Buena parte de la plana mayor de la biología soviética fue eliminada: N. V. Agol, acusado de trotskista, fue ejecutado; Levitt fue arrestado también y murió pronto en prisión. Nikolai Ivanovich Vavilov, uno de los botánicos más eminentes de su tiempo, fue enviado a la cárcel en 1940 y condenado a muerte. Aunque la sentencia fue pospuesta, Vavilov murió en 1943 en la prisión de Saratov.
En 1948, un decreto del Ministerio de Educación ordenaba la abolición de todos los cursos sobre genética mendeliana, la eliminación de todos los proyectos de investigación que tuvieran esa orientación, la erradicación de los libros que la apoyaran y la reorganización de todos los departamentos de ciencias biológicas de las universidades, destituyendo a los biólogos mendelianos y reemplazándolos por lysenkistas (y esto ocurría sólo cinco años antes de que Watson y Crick descifraran la estructura del ADN).
Desde 1948 hasta 1964, Lysenko reinó soberano en la biología y la agricultura soviéticas (ya que, tras la muerte de Stalin, Krushov también le prestó su apoyo, para desesperación de su hijo, Serguei Nikítich Krushov, que era científico aeronáutico y no dejaba de decirle a su padre que el lysenkismo era un disparate sin ninguna base racional). La investigación y el desarrollo en una de las ramas más dinámicas de la ciencia contemporánea lentamente fue deslizándose al marasmo y finalmente se detuvo, con sus lógicas consecuencias en la agricultura: en 1963, por primera vez en su historia, la Unión Soviética debió importar granos para alimentar a su población.
El desastre agrícola que el lysenkismo produjo en la Unión Soviética no fue para nada ajeno a la caída de Krushov, el 14 de octubre de 1964. El retraso científico en un área central como la biotecnología fue tal que la entonces Unión Soviética nunca pudo recuperar el tiempo perdido.
El proyecto genoma humano
Pero volvamos a hablar de cosas felices. El desarrollo de la genética fue espectacular, y en el año 1990 nació el ambicioso Proyecto Genoma Humano (PGH), un programa internacional originariamente dirigido por James Watson, destinado a determinar la secuencia completa de pares de bases químicas que componen el ADN humano, es decir, cartografiar la totalidad de nuestra información genética.
Inicialmente, se trató de un emprendimiento público, que se desarrollaba en centros públicos de investigación y universidades, pero a partir de 1995 entraron en carrera empresas privadas como Celera Genomics, creada en 1998 por el biólogo estadounidense Craig Venter, que ocultándose detrás de la intención de poner en marcha un nuevo método de secuenciamiento, mucho más rápido, reveló finalmente su intención de patentar genes (lo cual desató una verdadera batalla económica y ética entre quienes sostenían que la información tenía que ser de acceso público y los empresarios, que abogaban por un acceso restringido y estaban decididos a recuperar el tiempo y el dinero invertidos).
Finalmente, con el acuerdo de ambos grupos, en junio de 2000 se dio a conocer un borrador inicial del genoma. Un año más tarde, también ambos grupos publicaron informes en las revistas Nature y Science, donde revelaban la secuencia del genoma y finalmente, en abril de 2003, el NHGRI (National Human Genome Research Institute) y el Departamento de Energía de los Estados Unidos anunciaron oficialmente la terminación exitosa del PGH. En un artículo publicado en Nature el 24 de abril de 2003, justo cuando se cumplían cincuenta años de la comunicación de la estructura del ADN realizada por Watson y Crick, presentaron lo que denominaron «una visión para el futuro de la investigación sobre el genotipo». Venter, por su parte, debió admitir que la información fuera ofrecida al dominio público. De hecho, los datos de la secuencia generados por el PGH fueron depositados en bases de datos públicos y puestos a disposición de los científicos de todo el mundo, sin restricciones en cuanto a su uso o redistribución.
El secuenciamiento del genoma generó algunas sorpresas: si bien se creía que estábamos conformados por alrededor de 100 mil genes, resultó que no poseemos más que unos 30 mil, un valor no muy lejano de los que poseen algunos gusanos. Por otra parte, la evidencia mostró que la especie Homo sapiens, según las evidencias arqueológicas y genéticas, tuvo un único origen en África y de allí se dispersó a otras regiones del planeta. Quedó demostrado, al mismo tiempo, que los genes que determinan ciertas características pueden ser muy diferentes entre dos personas cuyos rasgos externos hacen que sean considerados de la misma raza: dos individuos que pertenecen a distintas razas pueden presentar más similitud genética en ciertas características que dos personas que pertenecen a la misma. Lo cual, felizmente, transformó en obsoleto el concepto de raza.
El secuenciamiento del ADN planteó, obviamente, nuevos desafíos. Uno de ellos, por ejemplo, fue el del denominado «ADN basura». En un principio, se puso en evidencia que sólo el 1,2 por ciento del ADN total está constituido por genes, es decir, por secuencias que contienen claves para la fabricación de proteínas; el resto no es más que una sucesión de «letras» en apariencia sin sentido, que, según se pensaba, no servía para nada. Era un dato algo incómodo, pero la investigación científica parecía confirmarlo: una buena parte del ADN, esa molécula central para la vida, era absolutamente inútil. Sin embargo, un informe de 2012 realizado por un proyecto internacional que involucra a más de 400 científicos (ENCODE) reveló que un porcentaje importante de ese conjunto aparentemente inútil (al menos el 80 por ciento) cumple un papel central a la hora de determinar el comportamiento de las células, los órganos y otros tejidos: funcionan como una suerte de «interruptores» que activan o desactivan a los genes y cuyo mal funcionamiento puede ser responsable de enfermedades conocidas.
En fin: como ocurrió en su momento con la microbiología de Pasteur y Koch, la genética despertó (y sigue despertando) expectativas desmesuradas sobre la futura cura de enfermedades hasta entonces (y hasta ahora) rebeldes, como el cáncer u otras. Muchas de esas ilusiones ya se revelaron como lo que eran (justamente, ilusiones) y se vio que no guardaban relación con lo que efectivamente podía lograrse en su momento. Si aún hoy mantenemos las esperanzas de que la genética sea un puntal de la medicina, que en cierto sentido lo es, todavía habrá que esperar e incorporar otros conocimientos y técnicas, muchas de ellas novedosas e inimaginables por ahora.