It is hard to begin a chapter about the brain without waxing lyrical. The brain is the physical organ that makes all our mental life possible. It enables us to read these words, and to consider thoughts that we have never considered before—or even to create thoughts that no human has considered before. This book will scratch the surface of how this is all possible, but the purpose of this chapter is more mundane. It offers a basic guide to the structure of the brain, starting from a description of neurons and working up to a description of how these are organized into different neuroanatomical systems. The emphasis is on the human brain rather than the brain of other species.
All neurons have basically the same structure. They consist of three components: a cell body (or soma), dendrites, and an axon. Although neurons have the same basic structure and function, it is important to note that there are some significant differences between different types of neurons in terms of the spatial arrangements of the dendrites and axon.
The cell body contains the nucleus and other organelles. The nucleus contains the genetic code, and this is involved in protein synthesis (e.g. of certain neurotransmitters). Neurons receive information from other neurons and they make a “decision” about this information (by changing their own activity) that can then be passed on to other neurons. From the cell body, a number of branching structures called dendrites enable communication with other neurons. Dendrites receive information from other neurons in close proximity. The number and structure of the dendritic branches can vary significantly depending on the type of neuron (i.e. where it is to be found in the brain). The axon, by contrast, sends information to other neurons. Each neuron consists of many dendrites but only a single axon (although the axon may be divided into several branches called collaterals).
Ten Interesting Facts About the Human Brain
- (1) There are 86 billion neurons in the human brain (Azevedo et al., 2009).
- (2) Each neuron may connect with around 10,000 other neurons.
- (3) If each neuron connected with every single other neuron, our brain would be 12.5 miles in diameter (Nelson & Bower, 1990). This is the length of Manhattan Island. This leads to an important conclusion—namely, that neurons only connect with a small subset of other neurons. Neurons may tend to communicate only with their neighbors, and long-range connections are the exception rather than the rule.
- (4) The idea that we only use 10 percent of the cells in our brain is generally considered a myth (Beyerstein, 1999). It used to be thought that only around 10 percent of the cells in the brain were neurons (the rest being cells called glia), hence a plausible origin for the myth. This “fact” also turns out to be inaccurate, with the true ratio of neurons to glia being closer to 1:1 (Azevedo et al., 2009). Glia serve a number of essential support functions; for example, they are involved in tissue repair and in the formation of myelin.
- (5) The brain makes up only 2 percent of body weight.
- (6) It is no longer believed that neurons in the brain are incapable of being regenerated. It was once widely believed that we are born with our full complement of neurons and that new neurons are not generated. This idea is now untenable, at least in a region called the dentate gyrus (for a review, see Gross, 2000).
- (7) On average, we lose a net amount of one cortical neuron per second. A study has shown that around 10 percent of our cortical neurons perish between the ages of 20 and 90 years— equivalent to 85,000 neurons per day (Pakkenberg & Gundersen, 1997).
- (8) Identical twins do not have anatomically identical brains. A comparison of identical and nonidentical twins suggests that the three-dimensional cortical gyral pattern is determined primarily by non-genetic factors, although brain size is strongly heritable (Bartley et al., 1997).
- (9) People with autism have large brains (Abell et al., 1999). They also have large heads to accommodate them. There is unlikely to be a simple relationship between brain size and intellect (most people with autism have low IQ), and brain efficiency may be unrelated to size.
- (10) Men have larger brains than women, but the female brain is more folded, implying an increase in surface area that may offset any size difference (Luders et al., 2004). The total number of cortical neurons is related to gender, but not overall height or weight (Pakkenberg & Gundersen, 1997).
Key Terms
Neuron
A type of cell that makes up the nervous system and supports, among other things, cognitive function.
Cell body
Part of the neuron containing the nucleus and other organelles.
Dendrites
Branching structures that carry information from other neurons.
Axon
A branching structure that carries information to other neurons and transmits an action potential.
Synapse
The small gap between neurons in which neurotransmitters are released, permitting signaling between neurons.
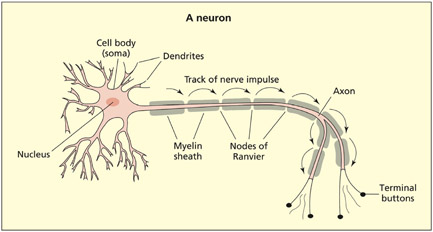
Neurons consist of three basic features: a cell body, dendrites that receive information and axons that send information. In this diagram the axon is myelinated to speed the conduction time.
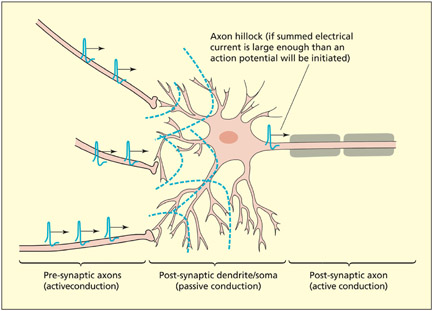
Electrical currents are actively transmitted through axons by an action potential. Electrical currents flow passively through dendrites and soma of neurons, but will initiate an action potential if their summed potential is strong enough at the start of the axon (called the hillock).
The terminal of an axon flattens out into a disc-shaped structure. It is here that chemical signals enable communication between neurons via a small gap termed a synapse. The two neurons forming the synapse are referred to as presynaptic (before the synapse) and postsynaptic (after the synapse), reflecting the direction of information flow (from axon to dendrite). When a presynaptic neuron is active, an electrical current (termed an action potential) is propagated down the length of the axon. When the action potential reaches the axon terminal, chemicals are released into the synaptic cleft. These chemicals are termed neurotransmitters. (Note that a small proportion of synapses, such as retinal gap junctions, signal electrically and not chemically.) Neurotransmitters bind to receptors on the dendrites or cell body of the postsynaptic neuron and create a synaptic potential. The synaptic potential is conducted passively (i.e. without creating an action potential) through the dendrites and soma of the postsynaptic neuron. If these passive currents are sufficiently strong when they reach the beginning of the axon in the postsynaptic neuron, then an action potential (an active electrical current) will be triggered in this neuron. It is important to note that each postsynaptic neuron sums together many synaptic potentials, which are generated at many different and distant dendritic sites (in contrast to a simple chain reaction between one neuron and the next). Passive conduction tends to be short range because the electrical signal is impeded by the resistance of the surrounding matter. Active conduction enables long-range signalingsignaling between neurons by the propagation of action potentials.
Key Terms
Action potential
A sudden change (depolarization and repolarization) in the electrical properties of the neuron membrane in an axon.
Neurotransmitters
Chemical signals that are released by one neuron and affect the properties of other neurons.
Electrical signaling and the action potential
Each neuron is surrounded by a cell membrane that acts as a barrier to the passage of certain chemicals. Within the membrane, certain protein molecules act as gatekeepers and allow particular chemicals in and out under certain conditions. These chemicals consist, among others, of charged sodium (Na+) and potassium (K+) ions. The balance between these ions on the inside and outside of the membrane is such that there is normally a resting potential of –70 mV across the membrane (the inside being negative relative to the outside).
Voltage-gated ion channels are of particular importance in the generation of an action potential. They are found only in axons, which is why only the axon is capable of producing action potentials. The sequence of events is as follows:
- If a passive current of sufficient strength flows across the axon membrane, this begins to open the voltage-gated Na+ channels.
- When the channel is opened, then Na+ may enter the cell and the negative potential normally found on the inside is reduced (the cell is said to depolarize). At about –50 mV, the cell membrane becomes completely permeable and the charge on the inside of the cell momentarily reverses. This sudden depolarization and subsequent repolarization in electrical charge across the membrane is the action potential.
- The negative potential of the cell is restored via the outward flow of K+ through voltage-gated K+ channels and closing of the voltage-gated Na+ channels.
- There is a brief period in which hyperpolarization occurs (the inside is more negative than at rest). This makes it more difficult for the axon to depolarize straight away and prevents the action potential from traveling backwards.
An action potential in one part of the axon opens adjacent voltage-sensitive Na+ channels, and so the action potential moves progressively down the length of the axon, starting from the cell body and ending at the axon terminal. The conduction of the action potential along the axon may be speeded up if the axon is myelinated. Myelin is a fatty substance that is deposited around the axon of some cells (especially those that carry motor signals). It blocks the normal Na+/K+ transfer and so the action potential jumps, via passive conduction, down the length of the axon at the points at which the myelin is absent (called nodes of Ranvier). Destruction of myelin is found in a number of pathologies, notably multiple sclerosis.
Key Term
Myelin
A fatty substance that is deposited around the axon of some neurons that speeds conduction.
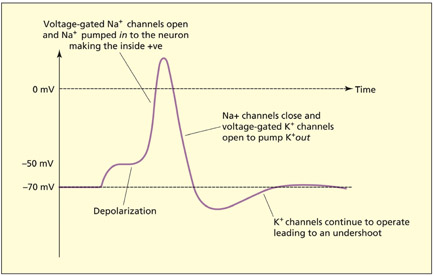
The action potential consists of a number of phases.
Chemical signaling and the postsynaptic neuron
When the action potential reaches the axon terminal, the electrical signal initiates a sequence of events leading to the release of neurotransmitters into the synaptic cleft. Protein receptors in the membrane of the postsynaptic neurons bind to the neurotransmitters. Many of the receptors are transmitter-gated ion channels (not to be confused with voltage-gated ion channels found in the axon). This sets up a localized flow of Na+, K+, or chloride (Cl–), which creates the synaptic potential. Some neurotransmitters (e.g. GABA) have an inhibitory effect on the postsynaptic neuron (i.e. by making it less likely to fire). This can be achieved by making the inside of the neuron more negative than normal and hence harder to depolarize (e.g. by opening transmitter-gated Cl– channels). Other neurotransmitters (e.g. acetylcholine) have excitatory effects on the post-synaptic neuron (i.e. by making it more likely to fire). These synaptic potentials are then passively conducted as already described.
How do neurons code information?
The amplitude of an action potential does not vary, but the number of action potentials propagated per second varies along a continuum. This rate of responding (also called the “spiking rate”) relates to the informational “code” carried by that neuron. For example, some neurons may have a high spiking rate in some situations (e.g. during speech), but not others (e.g. during vision), whereas other neurons would have a complementary profile. Neurons responding to similar types of information tend to be grouped together. This gives rise to the functional specialization of brain regions that was introduced in Chapter 1.
Key Terms
Gray matter
Matter consisting primarily of neuronal cell bodies.
White matter
Tissue of the nervous system consisting primarily of axons and support cells.
Glia
Support cells of the nervous system involved in tissue repair and in the formation of myelin (among other functions).
Corpus callosum
A large white matter tract that connects the two hemispheres.
Ventricles
The hollow chambers of the brain that contain cerebrospinal fluid.
If information is carried in the response rate of a neuron, what determines the type of information that the neuron responds to? The type of information that a neuron carries is related to the input it receives and the output it sends to other neurons. For example, the reason neurons in the primary auditory cortex can be considered to carry information about sound is because they receive input from a pathway originating in the cochlea and they send information to other neurons involved in more advanced stages of auditory processing (e.g. speech perception). However, imagine that one were to rewire the brain such that the primary auditory cortex was to receive inputs from the retinal pathway rather than the auditory pathway (Sur & Leamey, 2001). In this case, the function of the primary “auditory” cortex would have changed (as would the type of information it carries) even though the region itself was not directly modified (only the inputs to it were modified). This general point is worth bearing in mind when one considers what the function of a given region is. The function of a region is determined by its inputs and outputs. As such, the extent to which a function can be strictly localized is a moot point.
Gray matter, white matter, and cerebrospinal fluid
Neurons are organized within the brain to form white matter and gray matter. Gray matter consists of neuronal cell bodies. White matter consists of axons and support cells (glia). The brain consists of a highly convoluted folded sheet of gray matter (the cerebral cortex), beneath which lies the white matter. In the center of the brain, beneath the bulk of the white matter fibers, lies another collection of gray matter structures (the subcortex), which includes the basal ganglia, the limbic system, and the diencephalon.
White matter tracts may project between different cortical regions within the same hemisphere (called association tracts), may project between different cortical regions in different hemispheres (called commissures; the most important commissure being the corpus callosum) or may project between cortical and subcortical structures (called projection tracts).
The brain also contains a number of hollow chambers termed ventricles. These were incorrectly revered for 1,500 years as being the seat of mental life. The ventricles are filled with cerebrospinal fluid (CSF), which does serve some useful functions, albeit non-cognitive. The CSF carries waste metabolites, transfers some messenger signals, and provides a protective cushion for the brain.
A hierarchical view of the central nervous system
Brain evolution can be thought of as adding additional structures onto older ones, rather than replacing older structures with newer ones. For example, the main visual pathway in humans travels from the retina to the occipital lobe, but a number of older visual pathways also exist and contribute to vision (see Chapter 6). These older pathways constitute the dominant form of seeing for other species such as birds and reptiles.
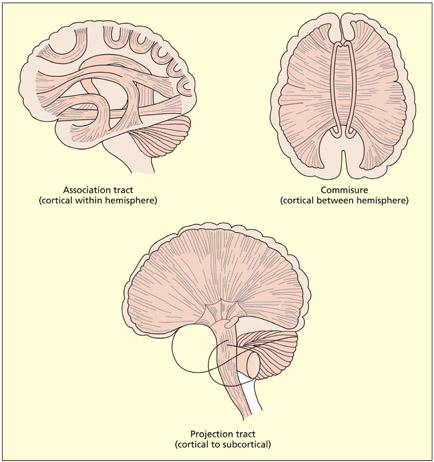
There are three different kinds of white matter tract, depending on the nature of the regions that are connected.

The brain consists of four ventricles filled with cerebrospinal fluid (CSF): the lateral ventricles are found in each hemisphere, the third ventricle lies centrally around the subcortical structures, and the fourth ventricle lies in the brainstem (hindbrain).
Terms of reference and section
Key Terms
Anterior
Towards the front.
Posterior
Towards the back.
Superior
Towards the top.
Inferior
Towards the bottom.
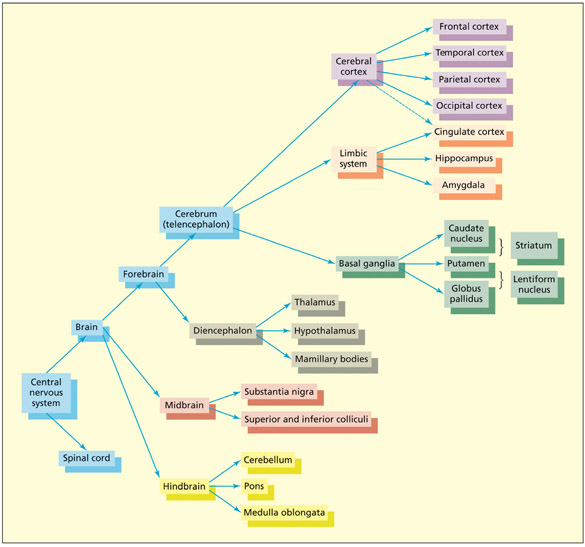
The central nervous system (CNS) is organized hierarchically. The upper levels of the hierarchy, corresponding to the upper branches of this diagram, are the newest structures from an evolutionary perspective.
There are conventional directions for navigating around the brain, just as there is a north, south, east, and west for navigating around maps. Anterior and posterior refer to directions toward the front and the back of the brain, respectively. These are also called rostral and caudal, respectively, particularly in other species that have a tail (caudal refers to the tail end). Directions toward the top and the bottom are referred to as superior and inferior, respectively; they are also known as dorsal and ventral, respectively. The terms anterior, posterior, superior, and inferior (or rostral, caudal, dorsal, and ventral) enable navigation in two dimensions: front–back and top–bottom. Needless to say, the brain is three-dimensional and so a further dimension is required. The terms lateral and medial are used to refer to directions toward the outer surface and the center of the brain, respectively; although “medial” is ambiguous, because it is also used in another context. Although it is used to refer to the center of the brain, it is also used to refer to the middle of structures more generally. For example, the medial temporal gyrus lies on the lateral surface of the brain (not the medial surface). It is labeled medial because it lies midway between the superior and inferior temporal gyri.
Key Terms
Ventral
Towards the bottom.
Lateral
The outer part (cf. medial).
Medial
In or toward the middle.
The brain can be sectioned into two-dimensional slices in a number of ways. A coronal cross-section refers to a slice in the vertical plane through both hemispheres (the brain appears roundish in this section). A sagittal section refers to a slice in the vertical plane going through one of the hemispheres. When the sagittal section lies between the hemispheres it is called a midline or medial section. An axial (or horizontal) section is taken in the horizontal plane.
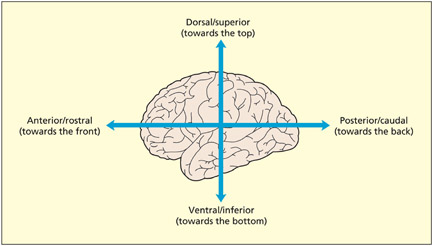
Terms of reference in the brain. Note also the terms lateral (referring to the outer surface of the brain) and medial (referring to the central regions).
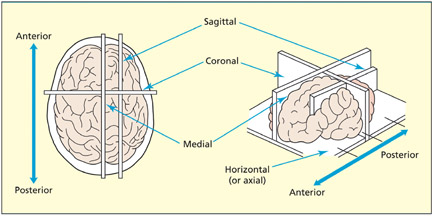
Terms of sections of the brain.
The cerebral cortex consists of two folded sheets of gray matter organized into two hemispheres (left and right). The surface of the cortex has become increasingly more convoluted with evolutionary development. Having a folded structure permits a high surface area to volume ratio and thereby permits efficient packaging. The raised surfaces of the cortex are termed gyri (or gyrus in the singular). The dips or folds are called sulci (or sulcus in the singular).
Key Terms
Gyri (gyrus = singular)
The raised folds of the cortex.
Sulci (sulcus = singular)
The buried grooves of the cortex.
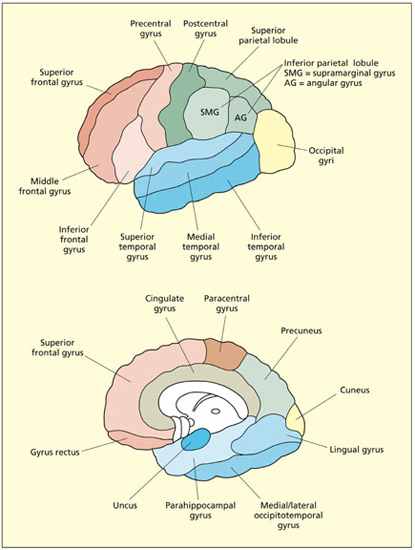
The main gyri of the lateral (top) and medial (bottom) surface of the brain. The cortical sulci tend to be labeled according to terms of reference. For example, the superior temporal sulcus lies between the superior and medial temporal gyri.
The cortex is only around 3 mm thick and is organized into different layers that can be seen when viewed in cross-section. The different layers reflect the grouping of different cell types. Different parts of the cortex have different densities in each of the layers. Most of the cortex contains six main cortical layers, termed the neocortex (meaning “new cortex”). Other cortical regions are the mesocortex (including the cingulate gyrus and insula) and the allocortex (including the primary olfactory cortex and hippocampus).
Key Term
Brodmann’s areas
Regions of cortex defined by the relative distribution of cell types across cortical layers (cytoarchitecture).
The lateral surface of the cortex of each hemisphere is divided into four lobes: the frontal, parietal, temporal and occipital lobes. The dividing line between the lobes is sometimes prominent, as is the case between the frontal and temporal lobes (divided by the lateral or sylvian fissure), but in other cases the boundary cannot readily be observed (e.g. between temporal and occipital lobes). Other regions of the cortex are observable only in a medial section, for example the cingulate cortex. Finally, an island of cortex lies buried underneath the temporal lobe; this is called the insula (which literally means “island” in Latin).
There are three different ways in which regions of cerebral cortex may be divided and, hence, labeled:
- Regions divided by the pattern of gyri and sulci. The same pattern of gyri and sulci is found in everyone (although the precise shape and size varies greatly). As such, it is possible to label different regions of the brain accordingly.
- Regions divided by cytoarchitecture. One of the most influential ways of dividing up the cerebral cortex is in terms of Brodmann’s areas. Brodmann divided the cortex up into approximately 52 areas (labeled from BA1 to BA52), based on the relative distribution of cell types across cortical layers. Areas are labeled in a circular spiral starting from the middle, like the numbering system of Parisian suburbs. Over the years, the map has been modified.
- Regions divided by function. This method tends only to be used for primary sensory and motor areas. For example, Brodmann areas 17 and 6 are also termed the primary visual cortex and the primary motor cortex, respectively. Higher cortical regions are harder (if not impossible) to ascribe unique functions to.
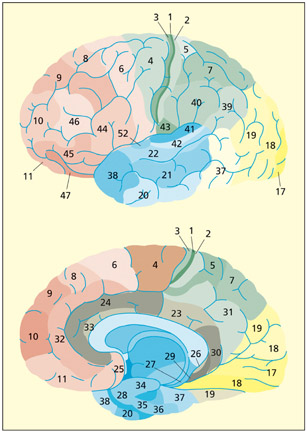
The Brodmann areas of the brain on the lateral (top) and medial (bottom) surface.
Beneath the cortical surface and the intervening white matter lies another collection of gray matter nuclei termed the subcortex. The subcortex is typically divided into a number of different systems with different evolutionary and functional histories.
Key Terms
Basal ganglia
Regions of subcortical gray matter involved in aspects of motor control and skill learning; they consist of structures such as the caudate nucleus, putamen, and globus pallidus.
Limbic system
A region of subcortex involved in relating the organism to its present and past environment; limbic structures include the amygdala, hippocampus, cingulate cortex, and mamillary bodies.
Thalamus
A major subcortical relay center; for instance, it is a processing station between all sensory organs (except smell) and the cortex.
Hypothalamus
Consists of a variety of nuclei that are specialized for different functions that are primarily concerned with the body and its regulation.
The basal ganglia
The basal ganglia are large rounded masses that lie in each hemisphere. They surround and overhang the thalamus in the center of the brain. They are involved in regulating motor activity, and the programming and termination of action (see Chapter 8). Disorders of the basal ganglia can be characterized as hypokinetic (poverty of movement) or hyperkinetic (excess of movement). Examples of these include Parkinson’s and Huntington’s disease, respectively (see Chapter 8). The basal ganglia are also implicated in the learning of rewards, skills, and habits (see Chapters 9 and 15). The main structures comprising the basal ganglia are: the caudate nucleus (an elongated tail-like structure), the putamen (lying more laterally) and the globus pallidus (lying more medially). The caudate and putamen funnel cortical inputs into the globus pallidus, from which fibers reach into the thalamus. Different circuits passing through these regions either increase or decrease the probability and intensity of certain behaviors (e.g. voluntary movements).
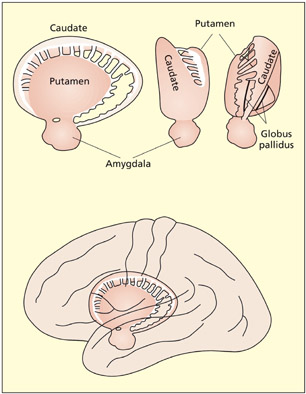
The basal ganglia are involved in motor programming and skill learning.
The limbic system
The limbic system is important for relating the organism to its environment based on current needs and the present situation, and based on previous experience. It is involved in the detection and expression of emotional responses. For example, the amygdala has been implicated in the detection of fearful or threatening stimuli (see Chapter 15), and parts of the cingulate gyrus have been implicated in the detection of emotional and cognitive conflicts (see Chapter 14). The hippocampus is particularly important for learning and memory (see Chapter 9). Both the amygdala and hippocampus lie buried in the temporal lobes of each hemisphere. Other limbic structures are clearly visible on the underside (ventral surface) of the brain. The mamillary bodies are two small round protrusions that have traditionally been implicated in memory (Dusoir et al., 1990). The olfactory bulbs lie on the under surface of the frontal lobes. Their connections to the limbic system underscore the importance of smell for detecting environmentally salient stimuli (e.g. food, other animals) and its influence on mood and memory.
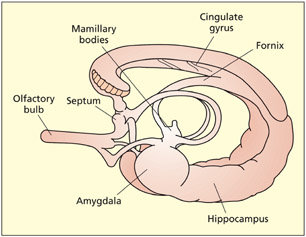
The limbic system.
The diencephalon
The two main structures that make up the diencephalon are the thalamus and the hypothalamus.
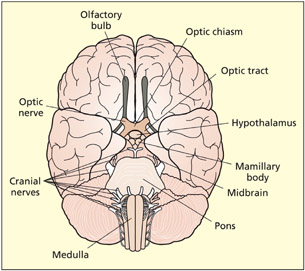
The ventral surface of the brain shows the limbic structures of the olfactory bulbs and mamillary bodies. Other visible structures include the hypothalamus, optic nerves, pons, and medulla.
The thalamus consists of two interconnected egg-shaped masses that lie in the center of the brain and appear prominent in a medial section. The thalamus is the main sensory relay for all senses (except smell) between the sense organs (eyes, ears, etc.) and the cortex. It also contains projections to almost all parts of the cortex and the basal ganglia. At the posterior end of the thalamus lie the lateral geniculate nucleus and the medial geniculate nucleus. These are the main sensory relays to the primary visual and primary auditory cortices, respectively.
The hypothalamus lies beneath the thalamus and consists of a variety of nuclei that are specialized for different functions primarily concerned with the body. These include body temperature, hunger and thirst, sexual activity, and regulation of endocrine functions (e.g. regulating body growth). Tumors in this region can lead to eating and drinking disorders, precocious puberty, dwarfism, and gigantism.
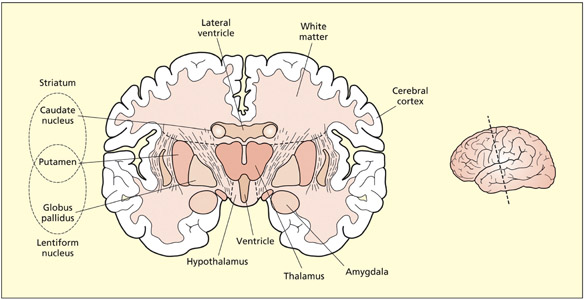
A coronal section through the amygdala and basal ganglia shows the thalamus and hypothalamus as prominent in the midline.
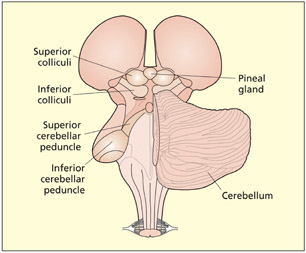
A posterior view of the midbrain and hindbrain. Visible structures include the thalamus, pineal gland, superior colliculi, inferior colliculi, cerebellum, cerebellar peduncle, and medulla oblongata (the pons is not visible but lies on the other side of the cerebellum).
Key Terms
Superior colliculi
A midbrain nucleus that forms part of a subcortical sensory pathway involved in programming fast eye movements.
Inferior colliculi
A midbrain nucleus that forms part of a subcortical auditory pathway.
The midbrain region consists of a number of structures, only a few of which will be considered here. The superior colliculi and inferior colliculi (or colliculus in singular) are gray-matter nuclei. The superior colliculi integrate information from several senses (vision, hearing, and touch), whereas the inferior colliculi are specialized for auditory processing. These pathways are different from the main #x003C;pg>cortical sensory pathways and are evolutionarily older. They may provide a fast route that enables rapid orienting to stimuli (flashes or bangs) before the stimulus is consciously seen or heard (Sparks, 1999). The midbrain also contains a region called the substantia nigra, which is connected to the basal ganglia. Cell loss in this region is associated with the symptoms of Parkinson’s disease.
Key Terms
Cerebellum
Structure attached to the hindbrain; important for dexterity and smooth execution of movement.
Pons
Part of the hindbrain; a key link between the cerebellum and the cerebrum.
Medulla oblongata
Part of the hindbrain; it regulates vital functions such as breathing, swallowing, heart rate, and the wake–sleep cycle.
The cerebellum (literally “little brain”) is attached to the posterior of the hindbrain via the cerebellar peduncles. It consists of highly convoluted folds of gray matter. It is organized into two interconnected lobes. The cerebellum is important for dexterity and smooth execution of movement. This function may be achieved by integrating motor commands with online sensory feedback about the current state of the action (see Chapter 8). Unilateral lesions to the cerebellum result in poor coordination on the same side of the body as the lesion (i.e. ipsilesional side). Bilateral lesions result in a wide and staggering gait, slurred speech (dysarthria), and eyes moving in a to-and-fro motion (nystagmus). The pons is a key link between the cerebellum and the cerebrum. It receives information from visual areas to control eye and body movements. The medulla oblongata protrudes from the pons and merges with the spinal cord. It regulates vital functions such as breathing, swallowing, heart rate, and the wake–sleep cycle.
