
Aside from deserts, with their distinctive hot and arid climate, rocky and sandy soil, and sparse vegetation, other arid and semiarid ecosystems exist. A steppe is an unusually dry and cold region where the dominant vegetation is grasses. The most prominent of these regions is the Eurasian Steppe, better known simply as the Steppe, which stretches in a band from Hungary in the west to Manchuria in the east. Scrubland is a dry region where the dominant vegetation is woody shrubs. These regions can be found from the Mediterranean to California to Australia.
The Eurasian Steppe is a belt of grassland that extends some 8,000 km (5,000 miles) from Hungary in the west through Ukraine and Central Asia to Manchuria in the east. Mountain ranges interrupt the steppe, dividing it into distinct segments; but horsemen could cross such barriers easily, so that steppe peoples could and did interact across the entire breadth of the Eurasian grassland throughout most of recorded history.
Nonetheless, the unity of steppe history is difficult to grasp; steppe peoples left very little writing for historians to use, and Chinese, Middle Eastern, and European records tell only what happened within a restricted range across their respective steppe frontiers. Archaeology offers real but limited help (grave relics from chieftains’ tombs abound but, of course, say little about everyday life and leave political, military, and linguistic alignments to inference). As a result, until about 1000 CE, information concerning the rise and fall of steppe empires and the relation between events in the eastern and western portions of the steppe remains fraught with great uncertainty.
The lay of the land divides the Eurasian Steppe into two major segments. The first of these may be called the Western Steppe. It extends from the grassy plains at the mouth of the Danube River along the north shore of the Black Sea, across the lower Volga, and eastward as far as the Altai Mountains.
The conventional division between Europe and Asia at the Ural Mountains is completely meaningless for steppe history and geography. The grasslands extend continuously south of the Ural Mountains on either side of the Ural River. The Western Steppe therefore constitutes one vast region, some 4,000 km (2,500 miles) from east to west and between 300 and 1,000 km (200 and 600 miles) from north to south. Within its bounds, a vast sea of grass made cross-country movement easy for anyone with a horse to ride. Rivers and streams cut through the grasslands, with trees growing along the banks. Streams flow slowly, trending, for the most part, either north or south and providing an easy mode of transport by river boat in summer and by sleigh in winter. Consequently, animal caravans and river transport made the steppe accessible to commerce even before modern roads and railroads transformed travel conditions.

On the Central Asian Steppe in Russia, a woman harvests wild grasses using a scythe. Lynn Johnson/National Geographic Image Collection/Getty Images
Hot summers and cold winters divide the year into sharply contrasting seasons. Temperatures are slightly more extreme in the east, but a more critical variable is rainfall, which diminishes as the rain-bearing winds from the Atlantic become increasingly erratic east of the Don. These temperature and precipitation gradients make the Ukraine and adjacent parts of Romania far richer natural pastureland than the land farther east. Peoples of the Western Steppe therefore tended to migrate westward along the steppe, seeking better grass and milder temperatures, whenever political conditions allowed them to do so.
The second major segment of the Eurasian Steppe extends from the Altai Mountains on the west to the Greater Khingan Range on the east, embracing Mongolia and adjacent regions. It is higher, colder, and drier than the Western Steppe, with greater seasonal extremes of temperature than are found anywhere else in the world. Some 2,500 km (1,500 miles) from east to west and about 650 to 800 km (400 to 500 miles) from north to south, the Eastern Steppe is in every way a harsher land for human habitation than the Western Steppe. All the same, lower temperatures counteract lower precipitation by reducing evaporation, so that sparse grass does grow, at least seasonally, even where rainfall is only between 250 and 500 mm (10 and 20 inches) a year. At higher elevations precipitation increases, and the mountaintops accumulate snow caps from which streams descend into the dry lands below. Irrigated cultivation is possible along such streams. Oasis dwellers, whose skills and goods complemented those of pastoralists, played important roles in steppe history.
These geographical conditions meant that nomads of the Eastern Steppe, living as they did in one of the most severe climates of the Earth, were under constant temptation to move in one of two directions: either southward and eastward toward Manchuria and northern China or westward, passing between the Altai and Tien Shan along the valley of the Ili River and the shores of Lake Balkhash, toward the more inviting grasslands of the Western Steppe. Migrations and conquests funneling through this Dzungarian Gate, as it is often called, gave the peoples of all the steppe a common history from the onset of horse nomadism. Warfare techniques, life-styles, religious ideas, artistic styles, languages, etc., spread widely across the steppes, never erasing local variations completely but making a single whole of the entire region in a more intimate way than the fragmentary records left by civilized scribes reveal.
Manchuria on the east and Hungary on the west are separated from the two main portions of the Eurasian Steppe by the Greater Khingan and Carpathian mountains, respectively, and are also distinguished by relatively benign climates favourable to agriculture. Hence, before modern times, a mixed economy of pastoral and agricultural activities had greater scope in Hungary and Manchuria than in the main areas of the steppe.
The same marginal participation in steppe history prevailed in the interior of Asia Minor, where open grassland, like that of the main portion of the steppe, was contiguous to similar grasslands in northern Syria and on southward into Arabia. On these southern grasslands arose another historically important style of nomad pastoralism that extended across the Red Sea deep into Africa as well. Since bypassing the Caucasus was easy for horsemen, movement from the northern to the southern grasslands occurred repeatedly. As a result, Eurasia’s two great pastoral traditions—Semitic in the south, Indo-European, Turkish, and Mongol in the north—met and mingled in Asia Minor and on steppe lands south of the Caucasus and therefore shared common traditions. In all likelihood, horses were first domesticated in the north, for example, but came to play important roles in Arabia and even in Africa; while the spread of Islam across the northern steppe attested to the impact of southern nomad ideals upon northerners.
Boundaries of the steppe are imprecise. Toward the north the Eurasian grasslands fade into forested landscapes, and because of long winters and short, cool summers, even scant rainfall can sustain scrub growth of spruce and other conifers. The Russian name for such forests is taiga, as steppe is the Russian word for grasslands; and it is convenient to use these terms to describe the two zones of vegetation that set narrow limits on human life in northern Eurasia even today.
The taiga was, for the most part, even more forbidding for human beings than the grasslands that lay to the south. In European Russia, a milder climate allowed deciduous forests to grow in some regions. There crops grew better than in the taiga, where agriculture could only be marginally successful, on account of poor soils and short growing seasons. In Asia, where taiga abutted directly on the steppe, hunters and gatherers of the forest were prone to migrate into the open grasslands. There they could establish themselves as nomads, and, being inured to the cold, heat, and hardship of the climate, they were in a good position to compete for a place on even the most forbidding steppe lands of Outer Mongolia. Similar migration from the forested north also occurred sporadically farther west. For example, Goths from southern Sweden penetrated the Ukraine in the early Christian centuries and swiftly adopted the habits and accoutrements of steppe nomads.
To the south the Eurasian Steppe fades into desert; but the deserts of Central Asia are dissected by mountain ranges in far more complicated fashion than the steppe proper. Since rainfall usually increases with elevation, mountains become islands of greenery in otherwise dry landscapes; and streams descending from mountaintops can sustain oasis cultivation in low-lying desert land. Grassland, sometimes merely seasonal, exists in all the mountainous areas of the Central Asian deserts. Complex, locally variable landscapes result. Hence the desert region that extends from the lower Volga and central Iranian plateau eastward through the Karakum and Kyzylkum deserts to the Takla Makan and Gobi in the east is uninhabitable only in some salt-encrusted lowlands. Even in the most barren reaches of unsalted soil, some herbage is occasionally available for animals to pasture on, and oases are often densely populated.
Interaction between steppe nomads and the various oasis dwellers of Central Asia was prolonged and intimate. Cities of the oases were often subjected to nomad rule; on the other hand, city dwellers’ superior skills regularly captivated unfriendly nomads, and suitably fortified cities could sometimes preserve their independence, even against nomad assault. Looked at from a steppe point of view, China and Europe, together with the cultivable areas of the Middle East, were no more than unusually large oases fed by moisture from adjacent oceans and from the Mediterranean and other inland seas.
Resources available for human life in these favoured regions were obviously more plentiful than in the steppe; and nomadic peoples, even when attached to their own ways of life, were strongly attracted by the wealth and ease that agricultural societies afforded. Movement southward from the steppe into one or another civilized zone was therefore a recurrent feature of Eurasian history. Nomads came as slaves, as traders and transport personnel, or as raiders and rulers. In this latter capacity, they played a politically prominent and often dominant part in Eurasian history. Because of their way of life, steppe peoples found it relatively easy to assemble large, mobile cavalry forces that could probe any weakness in civilized defenses and swiftly exploit whatever gaps they found. The political history of Eurasia consists very largely of nomad raids and conquests and the countervailing efforts by agricultural societies to defend themselves with an appropriate mix of armed force and diplomacy.
Geography did much to shape the pattern of these interactions. In the east the Gobi, dividing Outer Mongolia from China proper, constituted a considerable barrier. Successful raiding across the Gobi required a larger scale organization and more centralized command than was needed further west, where no such geographical obstacles existed. Thus, nomad impact on China was both sporadic and drastic. In Central Asia the complex borderlands between the contiguous steppe in the north and Iran and Turan (i.e., modern Sinkiang and most of Central Asia), with their tangled mix of desert, mountain, grassland, and cultivated fields, made interpenetration between nomad populations and settled agriculturalists easy and inevitable. There more than elsewhere civilized traditions of life and those of steppe tribesmen blended through the centuries of recorded history down to the present. To the west, in Europe, the boundary between steppe and sown land was far clearer than in Central Asia so that massed agricultural populations were more often able to protect themselves effectively from nomad harassment. As a result, nomad impact on European history was far less significant than in Central Asia and the Middle East, where, of course, pressure from the northern steppe was compounded by raiders and conquerors coming from Tibet and the southern grasslands.
The earliest human occupants of the Eurasian Steppe seem not to have differed very much from neighbours living in wooded landscapes. As elsewhere in Eurasia, hunters and gatherers using Paleolithic tools and weapons were succeeded on the steppes by Neolithic farmers who raised grain, kept domesticated animals, and decorated their pottery with painted designs. A series of critical developments eventually distinguished life on the steppes from life in the wooded lands, and the first of these was the domestication of horses.
It is impossible to say when the domestication of horses first took place. Early Mesopotamian figurines showing equine animals pulling a cart probably record the domestication of donkeys and onagers, not horses. Only a few horse bones have been identified at early sites, and they may attest to successful hunting rather than domestication. However, sometime around 4000 BCE steppe dwellers learned to keep herds of horses in addition to raising cattle, sheep, and goats, which were the principal domestic animals in more southerly lands.
Maximizing the size of domesticated herds made it necessary to pursue a migratory way of life because animals kept together for protection and control consumed the grass faster than it could grow, especially in the semiarid regions of the steppe. This made it hard to combine grain-growing with herding, as had been customary among Neolithic food producers. Eventually a clear break occurred between those peoples who raised crops and animals and those who depended solely on the products of their flocks and herds and moved from pasture to pasture throughout the year.
In all probability nomadism developed into a fully independent way of life only after human beings had learned to live largely on animal milk and milk products, thus tapping a new food source and, in effect, discovering a new ecological niche by displacing male lambs, calves, and colts from their mothers’ teats. Lactating animals had to be tamed to allow human beings to milk them by hand, and human populations also had to adjust physiologically by continuing as adults to secrete the enzymes children need to digest their mother’s milk. Such adaptations surely took considerable time, but how they proceeded remains unknown.
In Mongolia, a horsebreeder and his daughter milk a mare. Dean Conger/National Geographic Image Collection/Getty Images
Nevertheless, by about 2000 BCE these fundamental adjustments had probably been made, since a notable movement of peoples off the steppe and into the forested regions of Europe was under way. Herdsmen and warriors, speakers of Indo-European tongues, carried a distinctive battle-axe and, with the milk of their cattle and other animals, enjoyed a food supply that was clearly superior to those of other peoples. These advantages allowed the newcomers to overrun older farming and fishing populations of the European peninsula, so that their languages and cultures became dominant among later generations of Europeans.
Other, related peoples remained on the steppe, occupying the grasslands as far east as the Altai Mountains. Some Indo-European tribes also penetrated the Eastern Steppe, where, however, they presumably shared the landscape with peoples of other tongues. Such remarkable migrations suggest that by about 2000 BCE the speakers of Indo-European languages had attained a formidably efficient nomadic way of life.
It is very likely that behaviour patterns observed only later date back to this great dispersal. At any rate, the critical feature of later steppe nomadism was that only small groups could conveniently manage flocks and herds. In emergencies, manpower might have to be concentrated to protect people and animals against raiders from afar; but in ordinary times to have more than 50 to 100 persons camped at the same location made daily travel between pastures unbearably lengthy for lactating animals. Accordingly, during most of the year, steppe pastoralists dispersed into small kinship groups. Hundreds of animals were tended by dozens of persons. Every few days or weeks the group had to move to a new location where the herbage had not yet been eaten down. Only portable goods, therefore, were of much value to nomads, though of course their animals allowed them to transport heavier loads than human strength alone could support. Still, tents and leather containers, compared to the mud huts and pottery of settled folk, leave little trace for archaeologists. The possibility of learning much about how and when nomad patterns of life arose and spread across the steppe remains correspondingly slim.
Dependence on animals meant that relatively few human beings could make a living from the vast expanse of the Eurasian Steppe. Just how numerous ancient pastoralists may have been is impossible to say. The sudden appearance of large numbers of raiding horsemen often gave agricultural peoples the impression that vast hordes roamed the steppelands, waiting to pounce on undefended villages and towns. Because of the mobility of horsemen, raiding parties could gather from great distances, and thus the size of these groups (even if that were known) did not provide a reliable index of population density because their origins were unknown. The fact that a very large number of kurgans—i.e., mounds of earth raised atop chieftains’ graves—exist in the Western Steppe attests to the availability of relatively abundant manpower in ancient times, but, again, such monuments were raised by a chieftain’s followers, who were gathered for the purpose from afar and temporarily.
In general, there can be no doubt that nomadic populations always remained far sparser than agricultural populations. Nomad conquerors, however numerous they seemed at the moment of attack, were always far fewer than the settled populations they overran and, partly for that reason, were nearly always absorbed into the conquered society within a few generations.
In historic times yearly migrations followed a more or less fixed pattern—up and down mountain slopes with the season or north and south across open country for as much as 650–800 km (400–500 miles). When migratory herdsmen lived near cultivators, they often were able to pasture their animals on the stubble left behind after grain had been harvested. Exchanging grain for cheese and other animal products could also be mutually advantageous, even when rents or tribute payments skewed the simple economic symmetry of the relationship.
Dispersal across the steppe to maximize milk and meat production could be, and in historic times was, punctuated by occasional assemblages of large numbers of nomads for an annual roundup and slaughter of wild animals, for warlike undertakings against other communities, and for various ceremonial purposes as well. Real or fictitious kinship bonds united adjacent families of herdsmen into tribes; and tribal confederations, built upon ceremonial recognition of the primacy of a high king, were constructed and confirmed at such periodic assemblages.
Prowess in protecting the herds from animal predators had been necessary from the beginning of domestication, but, as pastoralism became an established way of life, prowess was redirected toward rival herdsmen. Quarrels over rights to grass and water were perennial, since boundaries between adjacent herding groups were necessarily imprecise and unpoliced. Infringement invited retaliation in the form of raids, and raids provoked counterraids. Warfare skills were thus inculcated by the nomads’ way of life, and their mobility made it possible to concentrate large numbers of experienced warriors wherever a tribal chieftain or high king might decree.
On the other hand, tribes and tribal confederations were always liable to break apart if the constituent groups felt aggrieved or merely distrusted the leader’s luck or military skill. Grounds for quarrels over precedence and dignity as well as over grass and water were always present within every steppe polity, and diplomats from civilized states were often able to exploit such weaknesses by pursuing a policy of “divide and rule.”
Nomadic customs and institutions thus superimposed fragile political structures on the migratory herding of small kinship groups. The formation of a far-flung war federation around the charismatic figure of a successful captain could occur very quickly. Division came even faster, since the passing of a high kingship from father to son was always precarious. Great men consolidated their power by marrying as many wives as the diversity of their following required, so whenever a great chieftain died, competition to the death among sons of different wives was likely. In effect, tribal confederations had to be reestablished every few generations.
Perhaps not all these features of nomad life were evident when civilized peoples first experienced the military might that steppe peoples could exert. For example, occupation of the Eastern Steppe by skilled horse nomads had not yet occurred when Indo-European conquerors first arrived in the Middle East, about 1700 BCE. Thereafter, scattered texts allow us to infer something about what happened on the Eurasian Steppe, although steppe peoples themselves did not begin to keep records in writing until about 2,300 years later.

Horsemen carry a ballot box back from the steppe where they collected votes for the general election from herdsmen in the district of Gachu outside Ulan Bator, Mongolia. C. Henriette/AFP/Getty Images
Also called shrubland, heathland, or chaparral, scrubland is a region with a diverse assortment of vegetation types sharing the common physical characteristic of dominance by shrubs. A shrub is defined as a woody plant not exceeding 5 metres (16.4 feet) in height if it has a single main stem, or 8 metres (26.2 feet) if it is multistemmed. The world’s main areas of scrubland occur in regions that have a Mediterranean climate—i.e., warm temperate, with mild, wet winters and long, dry summers. These areas include southern Australia, the Mediterranean region, California, Chile, and South Africa. Other scrublands are found in the semiarid tropics and in the Arctic, but smaller areas also occur in many other places. Australia, primarily because of its dry, variable climates, probably has the greatest expanse and range of scrublands.
Considering their variable presentation, it is not surprising that scrublands have diverse origins, which may be natural, anthropogenic, or both. Even the natural scrublands located in Mediterranean climates exhibit great interregional differences in plant species. This lends support to the view that the vegetation from scrublands of different areas evolved convergently—i.e., different ancestral plant species developed similar characteristics in response to similar climatic conditions. Thus, many plants that resemble each other from region to region are not closely related. Instead, they are said to be ecological equivalents.
Some ancestral plants, such as those of the scrublands of California and Australia, appear to have originated in more tropical environments. The overall climatic changes that led to the early evolution of scrubland plants from ancestors adapted to moister environments can be traced back to the gradual cooling and aridification of world climates that occurred during the Cenozoic Era (65.5 million years ago to the present). In California a significant climate change—elimination of reliable summer rainfall—took place about six million years ago. Even scrublands that appear to be entirely natural in their present form—for example, flora in regions of Mediterranean climate—probably have a geologically recent history. This is because the regions in which scrublands occur had radically different climates not long ago—at the end of the last Pleistocene Glacial interval about 11,700 years ago.
In areas in which climate clearly has been influential in the development of scrubland, human impact in such forms as fire or grazing also has been important. Anthropogenic scrublands—those arising from human impact on the vegetation—may be at least as widespread as natural scrublands. They occur where humans have altered an environment formerly dominated by trees to such an extent that it is no longer able to support them; this development is usually brought about through some combination of tree clearance, burning, and grazing that leads to soil degradation. In some cases, deforestation has led to the vigorous growth of shrubby plants that form a scrubland so dense that the originally dominant trees cannot return. This situation occurred in areas of former deciduous forest in Japan that have now been replaced by thickets of dwarf bamboo (Sasa). Rarely, scrublands may result from the introduction and establishment of a vigorous alien shrub in an area of grassland or in another shrub-free region.
Most natural scrublands occur under relatively harsh conditions. Even when a scrubland’s origin is attributable to human activity, however, the transition occurs most easily in areas where aspects of the natural environment are less than optimal for trees. By contrast, in habitats more suitable for trees even severe and repeated degradation of the vegetation is likely to be followed by some tree regeneration.
Some of the most extensive natural scrublands are found in regions of semiarid or Mediterranean climate between 20° and 40° latitude in both hemispheres. The most typical scrublands are found here, especially where fires are a regular feature of the environment. They occur mainly on the west (windward) coasts of continents between about 30° and 40° latitude. Where suitable land surfaces occur, as they do on either side of the Mediterranean basin, scrublands are fringed by deserts at lower latitudes and by temperate forests at higher latitudes. The climate is characterized by mild, moist winters and hot, dry summers. Trees are unable to grow in these regions and the vegetation is dominated by woody plants with a shrubby habit. These scrublands are very diverse, both in structure and in the identity and relationship of their component plants.
Scrublands in areas with Mediterranean climates experience drought stress in late summer when high temperatures lead to high rates of evaporation of water from both the soil and the plants. As there is usually little or no rain at this season, scrubland shrubs commonly become dormant. Most develop new leaves and experience a burst of flowering in spring when temperatures rise at the end of the wet winter season. In some areas, such as Western Australia, the display of spring blossoms is so spectacular that it has become a major tourist attraction.
In California scrubland vegetation growing under Mediterranean conditions occurs between 31° and 41° N and is called chaparral. Similar scrublands occupy areas of similar climate between 30° and 36° S in Chile, where they are called chaco and matorral. In both areas more than 80 percent of rain falls in winter. In areas of Mediterranean climate in Europe and Asia, scrublands occur over a wider climatic range, including areas receiving a larger proportion of average annual rain in summer—up to 40 percent of the annual 300 to 900 mm (12 to 35 inches). In these cases the wider distribution of scrubland results from a combination of climate and centuries of human influence.
In parts of the dry tropics, tall, open scrublands in which the dominant shrubs are more than five metres (16 feet) high—sometimes called thorn scrubs—grow across wide areas, to both the north and south of the Equator. In general they occupy the area between savannas at lower latitudes and deserts at higher latitudes. In these regions rainfall is low and erratic, and its effectiveness is further reduced by extremely high rates of potential evapotranspiration. Particularly large areas of such vegetation occur in Africa and Australia.
Scrublands also can be found in semiarid regions at temperate latitudes where the predominant season of low rainfall is typically, but not invariably, winter. A scrubland type found in several such regions is chenopod scrubland, named for its dominant shrubs, which belong to the chenopod, or saltbush, family. In Australia, for example, in areas south of the Tropic of Capricorn (23°27' S), which receive an average annual rainfall of 150 to 250 mm (6 to 10 inches), chenopod scrublands cover about 6 percent of the continent.
Although the main environmental condition responsible for the growth of scrublands in dry regions is moisture shortage, other factors may play important roles. In some cases soil is a primary factor in inducing development of scrubland vegetation. An example is provided by the fynbos, a type of scrubland found between latitudes 32° and 34° S in southwestern South Africa. Although rainfall in this region has the pronounced summer deficit typical of Mediterranean climates, nutrient-poor soils developed on various ancient rocks are thought to be primarily responsible for this unusual scrubland, famous for its great diversity of plant species.
Other natural stresses producing scrubland vegetation include salt-laden winds on coastal promontories and seasonal or yearlong cold. Where climates at high latitudes are too cool or offer too brief a warm season to permit tree growth, shrubs become dominant species, forming large areas of Arctic scrubland. Comparable but smaller areas of alpine scrubland are found on many high mountains.
The biota of scrublands, as might be anticipated from their broad geographic distribution and diverse environments, varies considerably from place to place.
In areas where scrublands occur naturally, especially as a result of nutrient-poor soils, and have existed for a long period, plant diversity can be astonishingly high. For example, more than 8,500 plant species are known from the fynbos of southwestern South Africa, and similar floristic richness occurs under comparable climatic and soil conditions in the scrublands of Western Australia. In each of these areas, a large proportion of the flora is endemic—that is, many plant species grow nowhere else and are known only as small populations at single locations.
Among the fynbos, the most diverse plant genera are Erica, Aspalathus, and Senecio, shrubs respectively in the heather, bean, and daisy families. Other richly represented families include the sedges, irises, grasses, lilies, and orchids, all of which consist of small, herbaceous plants that grow beneath the shrub canopy. The most colourful and conspicuous shrubs are heathers (Erica) and proteoids, especially Leucadendron and Protea. The flowers of this extraordinarily diverse flora are pollinated by both insects—but few butterflies—and nectar-eating birds such as sunbirds (Nectarina) and the Cape Sugarbird (Promerops cafer)—animals with which they have coevolved. Seed dispersal by ants occurs in an unusually large number of the plant species of the fynbos.
Many of the same plant families that are prominent in the fynbos also grow in scrublands in the temperate regions of Australia that receive winter rainfall. Members of the bean family are common, especially wattles (Acacia), as are many others—e.g., Phyllota and Pultenaea; proteoids such as Banksia, Conospermum, and Persoonia are also diverse and conspicuous. However, despite family resemblances, identical species rarely occur in both African and Australian heathlands, except through human introduction. For example, Australian wattles have been introduced into the South African fynbos where they pose substantial conservation problems. Heathers are virtually absent from Australia, their place being taken by epacrids (e.g., Lissanthe and Melichrus) and myrtles (e.g., Homoranthus and Kunzea). Again, bird pollination, primarily by various honeyeaters (family Meliphagidae), and seed dispersal by ants are common.
The scrublands of drier, hotter regions are poorer in plant species than most Mediterranean scrublands. Chenopod scrublands are dominated by shrubs in the family Chenopodiaceae; these bushes also are called saltbushes because they typically grow in dry, saline environments and sometimes store salt in their leaves, which have a characteristically salty taste. The mallees grow as multistemmed shrubs, frequently more than five metres (16 feet) tall, although some can assume tree form when conditions are not stressful. In Chilean scrublands members of the daisy family, such as Baccharis and Haplopappus, and members of the bean family, such as Acacia, Dalea, and Prosopis, are particularly well represented, together with cacti and many other plants.
The biota of Northern Hemisphere scrublands is distinct from that of the Southern Hemisphere. The wide variety of shrubs excludes proteoids, while some common groups of plants in the north such as oaks (Quercus) do not occur in the south. Other, more widespread, plant groups tend to be represented in northern scrublands by distant relatives of southern plants.
Scrublands in Mediterranean Europe are dominated by a variety of shrubs in such families as oak, heather (Erica and Arbutus), bean (Cytisus), olive (Olea), and pistachio (Pistacia). Oaks and heathers (Arctostaphylos) are also prominent in California, as are many other shrubs including Garrya, Heteromeles, Rhus, the blue-flowered Ceanothus, and the common chamiso (Adenostoma fasciculatum).
Many shrubs that dominate scrublands in all regions are thorny, and their foliage, which is usually not very nutritious, contains pungent and unpalatable substances that discourage browsing. Larger mammals are typically absent. Small mammals may be common, however, and they play an important role in influencing regeneration of the vegetation through eating seeds. Birds may be varied and abundant, but in Northern Hemisphere scrublands few are reliant on nectar as a food source and thus they only incidentally pollinate flowers, unlike the coevolved fauna of the Australian and South African scrublands.
Natural scrublands occur in areas in which factors such as climate or soil are sufficiently challenging to preclude the growth of trees but are adequate to allow the growth of shrubs. In many areas the shrubs belong to species that are able, under more favourable conditions, to develop into trees, but in others they always grow into shrub form regardless of the environment.
In scrublands with Mediterranean climate similar vegetation structure and ecological processes have evolved from different ancestral plants and animals in mutual isolation but in response to similar climatic environments. The extent to which this has occurred has been investigated substantially in the scrublands of California and Chile. These areas were chosen in part because natural patterns have not been obscured by long histories of intensive human agricultural impacts. This research has revealed remarkable similarities in the morphology of individual plants, the structure of the overall vegetation, the patterns of plant physiology and distribution along aridity gradients, and the diversity and adaptations of associated fauna.
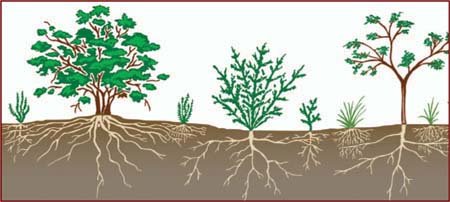
Vegetation profile of a scrubland. Encyclopædia Britannica, Inc.
The natural development of semiarid scrublands is affected by cultivation of the land and by grazing by domestic animals, a destructive form of land use that can substantially alter vegetation. Even regions that have been used for farming only for relatively short periods have been rapidly and substantially altered. Scrublands that are dependent on nutrient-poor soil can be permanently destroyed by the addition of nutrients, such as fertilizers used for agriculture or that have entered the ecosystem as pollution. The amelioration of soil conditions permits invasion by plants from other vegetation types that can grow more vigorously than the original scrubland plants.
In most regions of Australia, agriculture, including the grazing of domestic animals, has been practiced for less than 200 years. The semiarid tropical scrublands are reasonably intact across large areas, but the more southerly chenopod scrublands have been altered markedly during the past 150 years by intense sheep grazing. Ninety percent of these valuable rangelands have degenerated to some extent, and 25 percent have been severely affected. During this process, many plant species have become so rare that they are considered to be in danger of extinction if land-use practices are not changed. The number of palatable species seems to be declining, especially near watering points, although by contrast some unpalatable shrub species have become more common. This effect has greatly reduced the value of the vegetation as rangeland.
What was once Lake Natimuk in Australia is now dry and overgrown with weeds. Mark Dadswell/Getty Images
Large areas of mallee have been cleared to grow wheat, although the climate is so marginal that the crop often fails completely. Other Australian scrublands also continue to be cleared and fertilized for various agricultural purposes.
Broad-scale farming activity may create as well as destroy scrublands. This has happened in large areas of semiarid eastern Australia where grasslands were formerly dominant. Previously, when thick grass grew after a rain, it became fuel for wildfires, a process that favoured regrowth of the grass after the next rain but killed many shrub seedlings. After introduction of large numbers of domestic stock and eviction of the area’s Aboriginal inhabitants who had maintained the fire regime, the grass was mostly eaten by sheep and fires became less frequent. As a consequence, very large areas were invaded by “woody weeds”—unpalatable native shrubs that have created new areas of scrublands and greatly diminished the area’s grazing value.
Wherever climatic conditions are marginal for tree growth, vegetation is vulnerable to alteration by human activities. Trees disappear as a result, often to be replaced by shrubs. This vegetational degradation may have occurred so long ago as to be effectively permanent, leading modern observers to believe initially that these scrublands represent the natural, climate-induced vegetation of those areas. Scrublands of the Mediterranean region that have been affected by human activity for centuries, called by local names such as garigue, maquis, and macchia, are examples. Many areas have been converted from forest to scrubland by tree clearance, heavy grazing especially by goats, frequent burning, and consequent soil erosion. In these ways, evergreen forests of pine and oak have been replaced by scrublands in places throughout the northern Mediterranean fringe from Spain to Lebanon. Similar changes can be observed at similar latitudes as far eastward as China.
Occasionally, scrublands may result from the introduction of, and subsequent dominance by, an alien shrub species in a region in which woody plants are absent or rare. This has occurred in the grass-dominated, seasonally flooded areas of northernmost Australia to which the giant sensitive plant (Mimosa pigra) was introduced from Mexico; it since has multiplied to form extensive areas of dense, unnatural scrubland. A similar process is occurring in the Mitchell grasslands in inland northeastern Australia, an area being invaded by the introduced African shrub Acacia nilotica.
Fire is essential to the health of most scrublands. Without periodic burning, many scrublands would alter in composition; some would gradually develop into tree-dominated vegetation. Fires serve to kill young trees or to keep them in a shrub form. They also permit reproduction by the fire-adapted scrubland flora. Most scrubland plants are well adapted to survive fire, generally in one of two ways: Some resprout from the stem base or from underground organs after the aboveground parts have been destroyed; others regenerate from seeds that have been shed on the burned ground or that lie dormant in the soil. Many scrubland plants produce seeds that can remain dormant for long periods, a useful adaptation in an environment in which periods suitable for the establishment of seedlings are infrequent and unpredictable.
Many scrubland shrubs develop a swollen stem base or an underground woody organ called a lignotuber, which, because it is at or just below the soil surface, is protected from damage even during fires that completely remove the shrub’s crown. After a fire, buds under the woody outer layers of this organ quickly grow, using the undamaged original root system; a new crown develops within a few months. Similarly, many herbaceous plants have fleshy bulbs, rhizomes, or other types of underground stem from which green shoots rapidly develop in the wake of a fire that destroyed earlier shoots. Several species are stimulated to flower by fire, so that all individuals bloom simultaneously a few weeks after being burned. An example of this behaviour is the conspicuous Australian grass trees (Xanthorrhoea).
Other plants hold their seeds in woody fruits that remain closed until being opened by fire. Only then do the seeds fall to the ground, which has been temporarily fertilized by ash and cleared of dead leaf litter, competing plants, seed-harvesting ants, and parasitic molds. Such plants typically depend on fire to provide the conditions necessary for reproduction and are unable to regenerate without appropriate intervals of burning. For example, a common and attractive shrub of coastal scrublands in eastern Australia, Banksia ericifolia, is eliminated not only if an area is burned more often than every fifth year—the time taken for seedlings to set their first seed—but also if it is burned less often than every 40 years—the plant’s life span.
Many scrubland shrubs shed seeds as they ripen, even in years when fire does not occur. The seeds remain dormant in the soil until stimulated to germinate by heat or, in some cases, by chemicals in the smoke released by a fire. Heat stimulation is required for plant germination in several families, including the bean family (many species of Acacia) and the rose family (the Californian chamiso Adenostoma fasciculatum, which is stimulated to germinate by exposure to an air temperature of 90 to 95 °C [194 to 203 °F]).
Scrublands typically grow under conditions of high environmental stress. The typical climatic environment experienced by scrublands includes long periods of hot, dry weather in which lack of moisture is a limiting factor for plant growth. Furthermore, soil nutrient levels typically are very low. These factors restrict rates of plant photosynthesis. There may be a burst of growth during briefly favourable conditions, such as in spring in Mediterranean climatic regions, but productivity overall is not high.
The dominant shrubs may be deciduous, losing their leaves for significant periods of time when moisture is in short supply; if they are evergreen their leaves are small and tough and total leaf area is relatively low. They also typically have a substantial proportion of their biomass (dry weight of organic matter in an area) in the form of extensive root systems or lignotubers or other large underground organs. Only about half of the living vegetation comprises aboveground shoots, and only a small part of that is in the form of leaves or other green parts such as stems in which photosynthesis takes place. It is therefore easy to see why production of new plant material is slow.
Some of the most complete information on scrubland productivity has been assembled from data compiled on Californian chaparral. While values derived from studies of scrublands in climatically comparable parts of France and Greece are similar, greater variation occurs in other shrub-dominated vegetation in less similar environments.
Chaparral 1.5 metres (4.9 feet) tall had a biomass of 46.2 metric tons per hectare (51 tons per hectare), of which 16.7 (18.4 tons) was woody shoots, 3.7 (4 tons) leaves, 12.2 (13.4 tons) roots, and 13.6 (15 tons) litter on the ground. The annual accumulation of aboveground biomass in that system was 4.12 metric tons (4.5 tons) per hectare, of which 2.82 metric tons (3 tons) consisted of litter, with 2.64 metric tons (2.9 tons) of litter decomposing in the same period.
While such values typically demonstrate steady accumulation of biomass—in this case an annual aboveground increment of 0.13 metric tons (0.14 tons) per hectare—they do not take into account the occasional removal of aboveground parts through episodic disturbance, most commonly through fire. Productivity is more rapid during early stages of post-fire regrowth. For example, in one stand of French garigue, as the scrubland increased in height from 0.2 metres (0.66 feet) to 1.5 metres (4.9 feet) over a period from 1 to 10 years after a fire, its productivity declined from 4.7 to 3.2 metric tons (5.1 to 3.5 tons) per hectare per year.

