CELLS—THEY TALK ABOUT EVERYTHING!
CELLULAR COMMUNICATION is inherently complicated, with an immeasurable number of signals going in all directions at once. In the midst of billions of cells, a particular cell can rapidly make complex decisions and send signals that direct the efforts of many other cells to make our bodies work in amazing ways.
Before we delve into how specific types of cells—blood cells, gut and skin cells, cancer cells, brain cells, microbes, and more—use signals to perform their unique physiological functions, we’ll devote this chapter to discussing four areas that all cells appear to converse about, which was once unfathomable to even the most ardent researchers. These cellular conversations allow each cell to function with other cells in tissue throughout the body by knowing their appropriate size, their age, the time of day, and their own location.
While it is clear that individual cells are able to use the information that is described throughout this chapter in a variety of important ways, the mechanisms by which they do so are just now beginning to be discovered, and there is still much to learn. Finding individual minuscule molecules used as signals inside cells and tissues is extremely difficult, even as advanced imaging technologies are enabling us to view ever smaller details of cells. In further chapters, it will be seen that more detailed information is becoming available for many of the cells described.
DETERMINING THEIR PROPER SIZE
Cells come in many different shapes and sizes, but particular types of cells are usually produced within a narrow size range. For example, at least a thousand diverse types of neurons have specific sizes and shapes to fit into particular neural circuits. In other organs, it is not as obvious why cells are certain sizes.
Many influences determine cell sizes, such as input from the environment and signals from other cells. Cell sizes can be altered by signals from food particles or from molecules that are part of ordinary metabolic cycles. Cells can also increase size in new circumstances. Pancreatic cells, for instance, increase their size to make more insulin during pregnancy. But when these cells die because of the effects of diabetes, cell size stays the same, and the number of cells decreases.
Liver cells also increase in size during pregnancy. When fat cells enlarge, they signal changes in the extracellular matrix. Lymphocytes and microglia change size when immune cytokine signals trigger them to perform different functions. (Lymphocytes, microglia, and cytokine signals are all explained in the chapters ahead.)
Organs, too, have multiple techniques to maintain exact cell sizes, but these are not yet well understood. Even with differing environmental factors, they know what size the new cells should be. Cells produced from stem cells are not the size of the stem cell itself and can be ten times larger. The number of human cells, not the average cell size, makes one person larger than another. Even during rapid growth, organs maintain cell sizes. Two pancreatic cell types that are right next to each other are maintained at different sizes. Bone cells increase size by ten times when bone is growing.
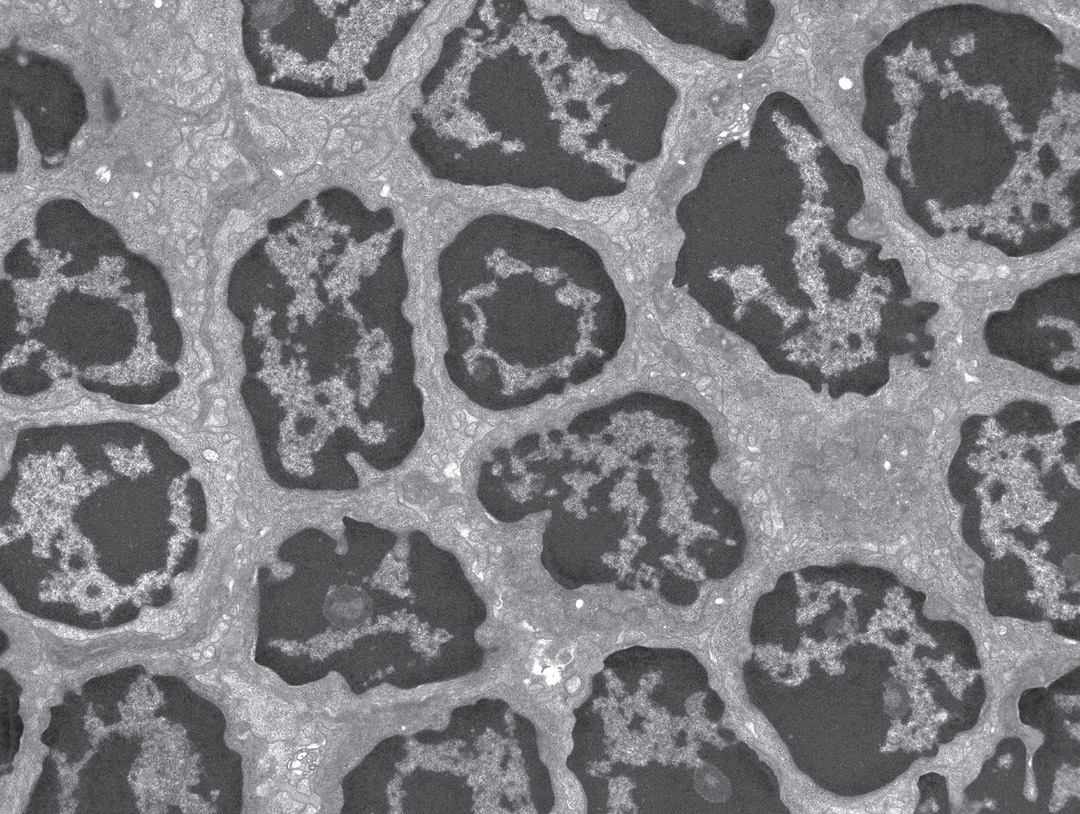
Lymphocytes in a lymph node. Electron micrograph. (Steve Gschmeissner/Science Source)
For an individual cell, size is determined by activity in phases of the multi-staged reproduction cycle for new cells. When new cells emerge, only certain-sized cells can proceed to the next stage in the process. For example, cells measure protein production as they go through different phases of the cycle. These levels of proteins become signals to tell a too-small cell that it has to stay in a particular phase longer to catch up in size. Cellular size is often increased between stages of DNA copying and the separation of the two DNA strands, for instance.
Sending secreted signal molecules from one cell to another is another technique used to alter cell size. These signals activate receptors on the second cell that signal internally to the nucleus to adjust the cell’s size. Signals include immune cytokine signals and growth factors (often proteins or hormones) that trigger cells to divide in order to produce new, smaller cells. Some signals increase growth and others decrease it. These factors have varied effects in different organs and are not yet well understood.
CELLS INFLUENCE THEIR OWN AGING PROCESS
Cells also can directly influence their own aging process. Cell aging has multiple distinct steps, and cells can make choices along the way. They can proactively use different rates of aging for various purposes. When healing a wound, for example, particular cells can age rapidly and die off to avoid producing severe body scars. In the fetus and during regeneration in adult organs, cells can use a rapid aging process to avoid making too many cells as a particular structure, such as an organ, is being developed. They can also preplan a tidy cleanup of dead-cell debris.
Cells use different genetic pathways to alter aging, including modifying how they reproduce. Cells can increase the rate of aging by shortening appendages (called telomeres) at the ends of DNA molecules. Cancer cells do the opposite—they enlarge the appendages to make themselves grow out of proportion.
Cells also produce an enzyme that can make the appendages longer or shorter, and new research has found a switch that turns this enzyme on and off. Multiple internal signaling pathways trigger the switch. These are pathways related to the repair of damaged DNA and they provide protection from destructive oxygen-based molecules produced in various metabolic processes that are related to cell stress.
Other major influences on cellular aging are pathways related to preprogrammed cell suicide, and these pathways are housed in mitochondria, the subcompartment of cells that provides energy and other important functions. The preplanned cell suicide pathway is used in a variety of circumstances in which maintaining the cell would be dangerous to the organism, such as a cell that has been infected by too many viruses and could spread infection.
With internal signals, cancer cells avoid triggers from mitochondrial metabolic pathways that stimulate the suicide pathway, enabling them to live much longer. With chronic stress, the opposite occurs in which cells use alternative metabolic pathways, triggered by mitochondria, to self-destruct. Also, immune signals can alter the normal cell-suicide mechanisms.
Two types of cell aging exist—acute and chronic. Acute aging is highly regulated during wound healing and in embryos to eliminate cells when their jobs are done. Signals are produced that stimulate aging for a particular group of cells in a section of tissue, but not the entire organ. They can trigger programmed cell-suicide pathways. Targeted cells rapidly age and die. This stops excessive scarring and other problems in the repair of organs, such as the liver.
Chronic aging occurs with cellular distress over time. It is random and generally considered to be “natural” cellular aging. For example, neurons that do not divide and can live for a century gradually accumulate random DNA damage and are eventually hurt by immune cytokine signals and inflammation. Aging cells gradually increase overall tissue aging by making it less functional. Then the entire body ages when cells stop multiplying, causing problems for the entire organism. Aging cells damage stem cell niches and destroy extracellular matrixes. Faulty cells impair structures. Aging cells stimulate damaging inflammation and send signals to make other cells begin to age. Signals for cellular division can stem this aging briefly. However, in the end, many stressors cause deterioration.
CELLS KNOW THE TIME OF DAY
Every cell has its own clock, and each type of tissue has its own specific set of internal clocks. Signals from the central brain clock coordinate physiological functions, such as metabolism and immune responses, with clocks in cells and tissue.
Single cells coordinate with the brain’s central clock as it responds to light and darkness cycles, bodily movements, and cycles of eating and fasting. Genetic loops in individual cells create oscillations that sync with other bodily rhythms. The brain coordinates and plans for specific activity related to the environment with these signals. Signals from the central brain clock to all cellular clocks anticipate the major activities of the total organism, such as eating and sleeping.
The first individual cell clock in evolution was developed in bacteria two billion years ago, and this was based on sun availability. In addition to enabling these bacteria to produce energy by photosynthesis, sun rays break DNA. At the same time, most cellular DNA repairs occur when the sun is bright. The first clock allowed microbes to plan ahead with resources for DNA repair when the sun was brightest.
Clock mechanisms and signals are complex and not yet fully understood. A mechanism in the gut that coordinates the cycles of two cells was discovered recently. Friendly microbes living near gut cells move in a timed pattern—a micrometer to the left, then right, then back. Back-and-forth signals from each position keep the microbes in sync with the cellular rhythms of the nearby gut lining.
An individual cell’s clock mechanism is based on timed feedback loops of interacting genes. Clock genes, components of the body’s internal timekeeping system, are both stimulated and inhibited by RNA and protein molecules. A gene is triggered, producing a protein or RNA, which then triggers a second gene in the circuit. The second gene product stimulates a third gene, and so on. These events form a cycle that lasts twenty-four hours.
Molecular tags are an important type of signaling device described throughout the book. Tags placed on proteins to protect DNA are also part of these clock loops. Tags can open or close the availability of particular genes that produce RNAs and proteins related to clock functions.
While all cells have the same basic genetic clock machinery, various RNAs and proteins specific to each type of cell and organ are signals that produce various clock functions. A huge amount of all RNAs—at least 10 percent—are related to tags and signals for clocks. Multiple layers of genetic regulation influence these cycles. For example, very recently a new form of regulation was discovered that alters three-dimensional structures of the DNA molecule in the nucleus. When the structure changes, it alters how physically close particular genes are to each other. Bringing certain genes near each other can synchronize clock functions.
Multiple influences affect clock rhythms. Signals from metabolic cycles alter specific RNAs and proteins to influence clock genes. Various chemicals in particular organs affect clocks in different ways. Global factors, such as temperature and other environmental conditions, alter gene function. Many of these complex clock signals for individual cells are not yet understood.
When tissue cannot sync with the central brain mechanisms, illness can occur. One issue that needs to be addressed is our twenty-fourhours-a-day, seven-days-a-week online culture, which pays no attention to the rhythms corresponding to daylight that were established in our distant evolution. Multiple other influences on our bodily clock functions are not yet well understood. For example, we don’t understand the clock cycles in the liver and pancreas, which operate on opposite schedules. We also don’t know how cancer cells are able to respond to particular rhythms to help them grow.
CELLS TALK ABOUT THEIR LOCATION
Cells need to know where they are in order to make multiple decisions. For example, a white blood cell needs to know its current location when traveling to an infection at another location. Cells near an infection send signals to these traveling immune cells along blood vessels, which then provide the directions for travel. This type of signaling for traveling white blood cells is described in chapter two.
Importance of Gradients
For cells in a developing tissue, location is often derived by using a chemical gradient as a measuring tool. In a fetus, for instance, a traveling neuron or stem cell must know where it is and where it must end up in the developing brain. Also, when cells participate in the growth of an organ or a limb, they need to know how they fit into the final shape, such as determining where the edges are.
When we measure something, we use a measuring rod that spans a distance. Cells can measure the distance that certain molecules have traveled, such as measuring these chemicals across groups of cells and the spaces in between. Cells located throughout the gradient determine where they are by picking up the gradient molecule with receptors, which measure the concentration of the molecule at their position. But for this measurement to be accurate, the gradient has to be steady and not fluctuating.
Establishing a steady state in a molecular gradient is based on many different factors, including the rate of molecular production, how rapidly the molecule diffuses through the tissue medium, and the rate of elimination of the gradient molecule when it is picked up by various receptors on cells. Other influences, such as temperature, metabolism, and inflammation, can affect gradient levels as well.
For a growing organ, a row of cells can produce gradient molecules at the same rate. Every cell in the path of the gradient takes up the molecules with receptors. Signaling molecules that form chemical gradients trigger particular genes inside the cell to determine the cell’s actions in relation to the growing tissue. This technique is used in forming a fly’s wing, for example, where signals from cells at the center of the tissue continue to stimulate new cells until there is a steep drop-off of the gradient that determines the edge. Gradients are an important way that body organs and types of tissue of very specific shapes are formed, but scientists are just learning exactly how this happens.
Gradients, Retinoic Acid, and Signaling
One important gradient that is beginning to be understood forms the structure of the fetal brain. It is based on various levels of retinoic acid, which is produced in two steps from dietary vitamin A picked up by cells. This metabolic pathway that produces retinoic acid is highly regulated by molecular signals, which makes maintaining the gradient and the cell’s calculations easier.
Regulation involves feedback loops of various proteins that regulate the multiple enzymes synthesizing the gradient molecule. In addition, cells produce a variety of diverse protein receptors with different sensitivities to eliminate the gradient molecule. These varied receptors are also regulated by feedback loops. In laboratory experiments, when the amounts of ingredients are altered, these cellular pathways adjust to maintain the gradient and produce the precise receptors needed. Somehow, neurons use this information to find their place in the developing fetus and build the brain precisely.
Stem cells in the fetus also compare the retinoic acid gradients with gradients of several other molecules. Cells can switch between measuring the two different molecular gradients. One cellular process is maintained until the switch occurs, and then the other begins. This recurs until the cell eventually makes a decision based on information from both. Stem cells use this mechanism to decide whether to divide in a particular spot in the developing brain. It is not yet clear how the switching technique is regulated.
It is quite remarkable that cells are able to know the exact shapes of organs, perform these location measurements, and send signals about these measurements. It is not yet understood how all of this works. But what is clear is that the daily maintenance of all tissue structures in the body is dependent on measurements and signals. As more is learned, perhaps we will find a way to exploit the stimulation of these signals to rebuild damaged organs.