All of the organisms we see today arose from earlier organisms. This process, known as evolution, can be described as a change in a population over time. Interestingly, however, the driving force of evolution, called natural selection, operates on the level of the individual. In other words, evolution is defined in terms of populations but occurs in terms of individuals.
What is the basis of our knowledge of evolution? Much of what we now know about evolution is based on the work of Charles Darwin. Darwin was a 19th-century British naturalist who sailed the world in a ship named the HMS Beagle. Darwin developed his theory of evolution based on natural selection after studying animals in the Galápagos Islands and other places.
He observed that there were similar animals on the various islands, but the beaks varied in length on finches, and necks varied in length on tortoises. There must be a reason why animals in different areas had different traits. Darwin concluded that it was impossible for the finches and tortoises of the Galápagos simply to “grow” longer beaks or necks as needed. Rather, those traits were inherited and passed on from generation to generation. So, he decided there must be a way for populations to evolve and change their traits (i.e., a population of finches developing longer beaks).
He decided that there must have been a variety of beak lengths originally, but only the longest one was particularly helpful. Since those finches could eat better, survive better, and reproduce better, they were more likely to contribute offspring to the next generation. Over many years, long-beak finches became more and more plentiful with each generation until long beaks were the “norm.” In another example, on the first island Darwin studied, there must once have been short-necked tortoises. Unable to reach higher vegetation, these tortoises eventually died off, leaving only those tortoises with longer necks. Consequently, evolution has come to be thought of as “the survival of the fittest”: only those organisms most fit to survive will survive and reproduce.
Darwin elaborated his theory in a book entitled On the Origin of Species. In a nutshell, here’s what Darwin observed:
-
Each species produces more offspring than can survive.
-
These offspring compete with one another for the limited resources available to them.
-
Organisms in every population vary.
-
The fittest offspring, or those with the most favorable traits, are the most likely to survive and therefore produce a second generation.
Darwin was not the first to propose a theory explaining the variety of life on Earth. One of the most widely accepted theories of evolution in Darwin’s day was that proposed by Jean-Baptiste de Lamarck.
In the 18th century, Lamarck had proposed that acquired traits were inherited and passed on to offspring. For example, in the case of Lamarck’s giraffes, Lamarck’s theory said that the giraffes had long necks because they were constantly reaching for higher leaves while feeding. This theory is referred to as the “law of use and disuse,” or, as we might say now, “use it or lose it.” According to Lamarck, giraffes have long necks because they constantly use them.
We know now that Lamarck’s theory was wrong: acquired changes—that is, changes at a “macro” level in an organism’s regular (somatic) body cells—will not appear in gamete cells. For example, if you were to lose one of your fingers, your children would not inherit this trait. The gametes that your body makes include copies of your regular old genome, not a new version of a genome that is determined by how careful you are with a table saw.
In essence, nature “selects” which living things survive and reproduce. Today, we find support for the theory of evolution in several areas:
-
Paleontology, or the study of fossils: paleontology has revealed to us both the great variety of organisms (most of which, including trilobites, dinosaurs, and the woolly mammoth, have died off) and the major lines of evolution. Fossils can be dated by:
i. The age of the rocks where a fossil is found
ii. The rate of decay of isotopes including carbon-14
iii. Geographical data
-
Biogeography, or the study of the distribution of flora (plants) and fauna (animals) in the environment: scientists have found related species in widely separated regions of the world. For example, Darwin observed that animals in the Galápagos have traits similar to those of animals on the mainland of South America. One possible explanation for these similarities is a common ancestor. As we’ll see below, there are other explanations for similar traits. However, when organisms share multiple traits, it’s pretty safe to say that they likely also shared a common ancestor.
-
Embryology, or the study of development of an organism: if you look at the early stages in vertebrate development, all embryos look alike! For example, all vertebrates—including fish, amphibians, reptiles, birds, and even mammals such as humans—show fishlike features called gill slits.
-
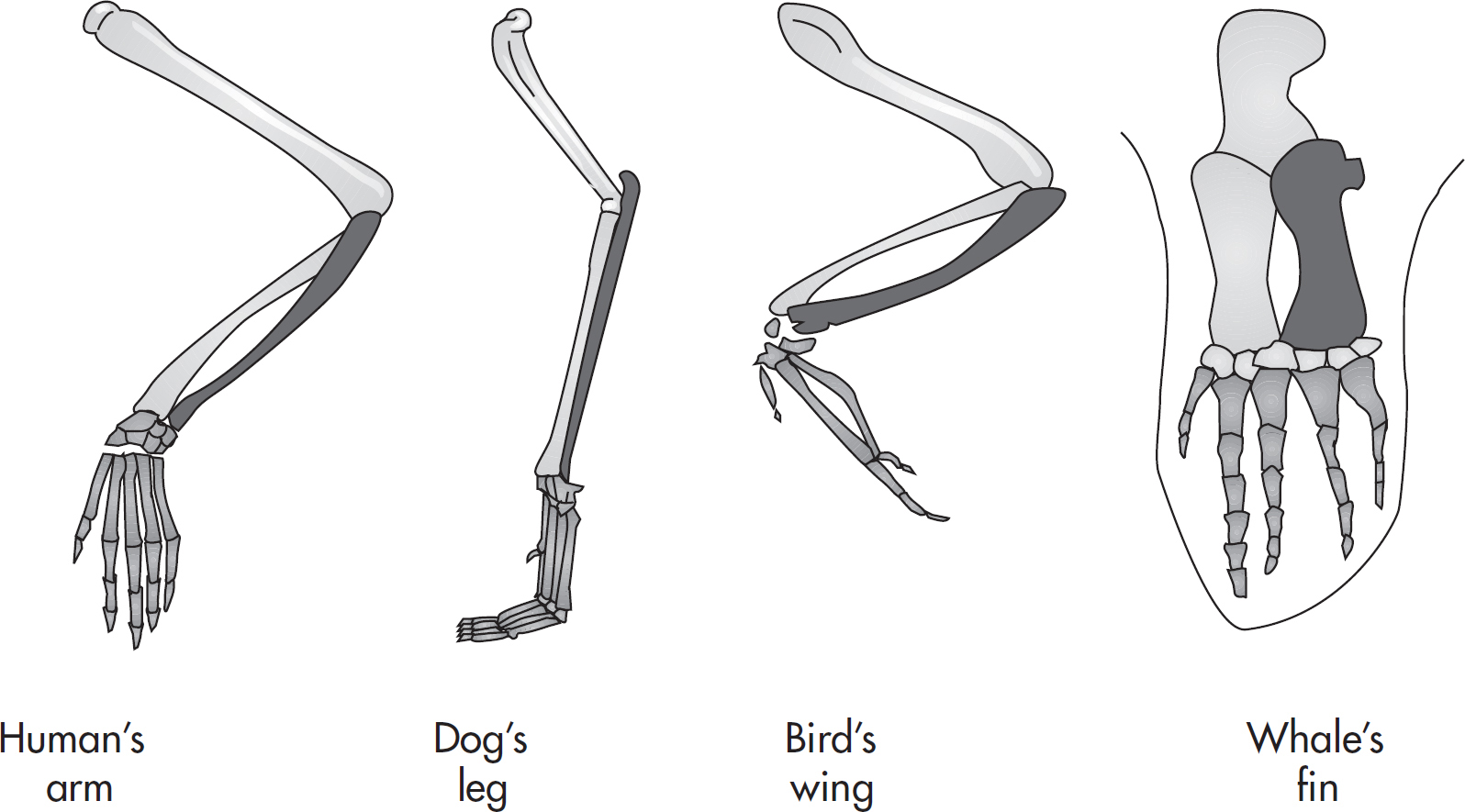
Comparative anatomy, or the study of the anatomy of various animals: scientists have discovered that some animals have similar structures that serve different functions. For example, a human’s arm, a dog’s leg, a bird’s wing, and a whale’s fin are all the same appendages, though they have evolved to serve different purposes. These structures, called homologous structures, also point to a common ancestor.
In contrast, sometimes animals have features with the same function but that are structurally different. A bat’s wing and an insect’s wing, for example, are both used to fly. They therefore have the same function, but they have evolved totally independently of one another. These are called analogous structures. Another classic example of an analogous structure is the eye. Though scallops, insects, and humans all have eyes, these three different types of eyes are thought to have evolved entirely independently of one another. They are therefore analogous structures.
-
Molecular Biology: Perhaps the most compelling proof of all is the similarity at the molecular level. Today, scientists can examine nucleotide and amino acid sequences of different organisms. From these analyses, we’ve discovered that organisms that are closely related have a greater proportion of sequences in common than distantly related species. For example, humans don’t look much like chimpanzees. However, by some estimates, as much as 99 percent of our genetic code is identical to that of a chimp. All eukaryotes have membrane-bound organelles, linear chromosomes, and genes that contain introns.
-
Continuing Evolution: There is evidence that evolution is constantly occurring. We can see consistent small changes in DNA and changes in the fossil record. We can see evolution of resistance to antibiotics, pesticides, herbicides, or chemotherapy drugs since these are particularly strong selective pressures. We can also see how fast replicating pathogens evolve and cause emergent diseases never before seen.
By using the evidence mentioned above, scientists can get a good handle on how evolution of certain species occurred. It all comes down to who has common ancestors. Life started somewhere. Some original life-form is the common ancestor to all life, but where things went from there can become quite convoluted.
Scientists use charts called phylogenetic trees, or cladograms, to study relationships between organisms. Phylogenetic trees are built using data from the fossil record or molecular record, but the molecular record provides more specific details. Trees show the amount of change over time, but cladograms just show the change. In other words, cladograms are often drawn with even spacing between species, but phylogenetic trees are often drawn with different distances between species and as a result they look more like a tree with uneven branches. Both types are hypotheses and thus they constantly change as more data is uncovered to establish the relationships between species.
They always begin with the common ancestor and then branch out. Anytime there is a fork in the road, it is called a common ancestor node. These common ancestors likely do not exist anymore, but they are the point at which evolution went in two directions. One direction eventually led to one species, and the other eventually led to another species.
For example, we have a common ancestor with chimps. This doesn’t mean that our great-great…great-grandfather was a chimp. It means that chimps and we have the same great-great…great-grandfather, who was neither a human nor a chimp. Instead, he was some other species completely, and a speciation event must have occurred where his species was split and evolved in different directions. One lineage became chimps, and the other linage became humans.
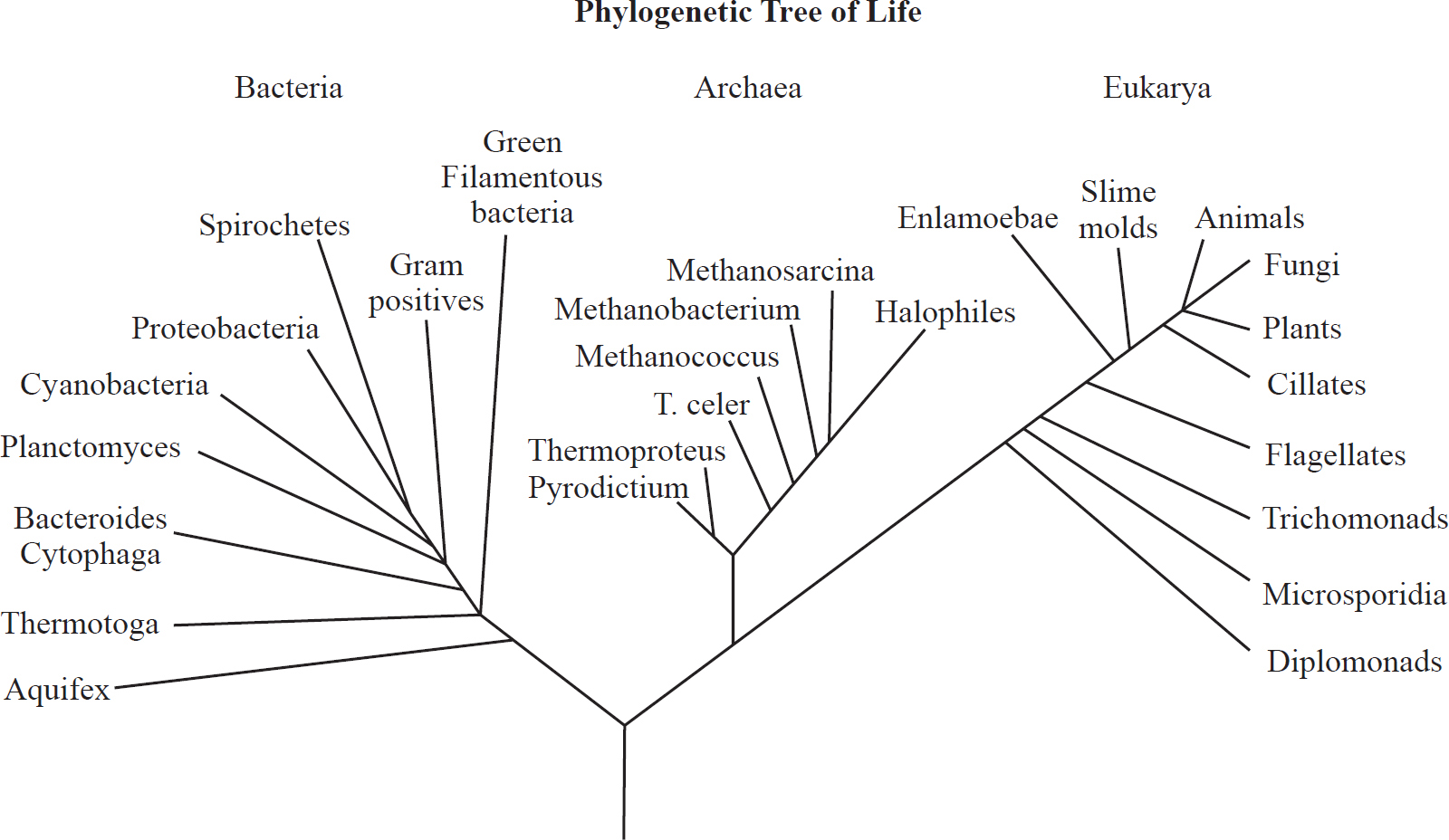
This is an example of a phylogenetic tree.
In this tree, the three main domains of life (bacteria, archaea, and eukarya) are seen. You can see that plants, animals, and fungi are much more closely related to each other than they are to bacteria. Archaea and eukarya have a common ancestor with bacteria, and then they also have another common ancestor that bacteria doesn’t have.
In the phylogenetic tree above species A is an out-group because it is the least related to all of the other species. Species B, C, D, and E all have the shared character of 8 legs. Wings are a shared character for species D and E, but they could also be considered a derived character since they separate species D and E from the rest of the species that do no have wings.
As you know, no two individuals are identical. The differences in each person are known as genetic variability. All this means is that no two individuals in a population have identical sets of alleles (except, of course, identical twins). In fact, survival of a species is dependent on this genetic variation; it allows a species to survive in a changing environment. If a population were all the same, then they would have the same weaknesses and the same strengths. Natural selection occurs only if some individuals have more evolutionary fitness and can be selected. The more variations there are among a population, the more likely that a trait will exist that might be the perfect lifesaver. One famous example is Rudolph the Red-Nosed Reindeer. His red nose was a random mutation, but under the right conditions, it became the best nose to have. A more diverse population is more likely to survive and evolve when things are constantly changing around them.
How did all this wonderful variation come about? Well, the simple answer is that random mutations are occurring all the time. This can be because of errors by DNA polymerase, changes to DNA caused by transposons, or other types of DNA damage. Either way, mutations create new variations and alleles.
However, just the occurrence of mutations does not account for the great amount of genetic variation in a population. The mixing of genes through sexual reproduction also contributes to genetic variation. During meiosis, crossing-over mixes alleles among homologous chromosomes, and independent assortment when chromosomes are packaged further adds to the genetic uniqueness of each gamete.
It might be hard to think of it in this way, but genetic variation is the very foundation of evolution, as we’ll soon see. Now that we’ve reintroduced genes, we can refine our definition of evolution. More specifically:
Evolution is the change in the gene pool of a population over time.
Let’s look at an example. During the 1850s in England, there was a large population of peppered moths. In most areas, exactly half of them were dark and carried alleles for dark coloring. The other half were light and carried alleles for light coloring. This 1:1 ratio of phenotypes was observed until air pollution, due primarily to the burning of coal, changed the environment. What happened?
Imagine two different cities: City 1 (in the south) and City 2 (in the north). Prior to the Industrial Revolution, both of these cities had unpolluted environments. In both of these environments, dark moths and light moths lived comfortably side by side. For simplicity’s sake, let’s say our proportions were a perfect fifty-fifty, half dark and half light. However, at the height of the Industrial Revolution, City 2, our northern city, was heavily polluted, whereas City 1, our southern city, was unchanged. In the north, where all the trees and buildings were covered with soot, the light moths didn’t stand a chance. They were impossible for a predator to miss! As a result, the predators gobbled up light-colored moths just as fast as they could reproduce, sometimes even before they reached an age in which they could reproduce. However, the dark moths with their dark alleles were just fine. With all the soot around, the predators couldn’t even see them; the dark moths continued doing their thing—above all, reproducing. And when they reproduced, they had more and more offspring carrying the dark allele.
After a few generations, the peppered moth gene pool in City 2 changed. Although our original moth gene pool was 50 percent light and 50 percent dark, excessive predation changed the population’s genetic makeup. By about 1950, the gene pool reached 90 percent dark alleles and only 10 percent light alleles. This occurred because the light moths didn’t stand a chance in an environment where they were so easy to spot. The dark moths, on the other hand, multiplied just as fast as they could.
In the southern city, you’ll remember, there was very little pollution. What happened there? Things remained pretty much the same. The gene pool was unchanged, and the population continued to have roughly equal proportions of light moths and dark moths.
Natural selection, the evolutionary mechanism that “selects” which members of a population are best suited to survive and which are not, works both “internally” and “externally”: internally through random mutations and externally through environmental pressures. Biotic and abiotic factors can fluctuate and this can affect the direction of evolution. With each generation, different traits might be selected for depending on environmental fluctuations.
Natural selection requires genetic variation and an environmental pressure that gives some individuals an advantage.
To see how this process unfolds in nature, let’s return to the moth case. Why did the dark moths in the north survive? Because they were dark-colored. But how did they become dark-colored? The answer is, through random mutation. One day, long before the coal burning, a moth was born with dark-colored wings. As long as a mutation does not kill an organism before it reproduces (most mutations, in fact, do), it may be passed on to the next generation. Over time, this one moth had offspring. These, too, were dark. The dark- and the light-colored moths lived happily side by side until something from the outside—in our example, the environment—changed all that.
The initial variation came about by chance. This variation gave the dark moths an edge. However, that advantage did not become apparent until something made it apparent. In our case, that something was the intensive pollution from coal burning. The abundance of soot made it easier for predators to spot the light-colored moths, thus effectively removing them from the population. Therefore, dark color is an adaptation, a variation favored by natural selection.
Survival of the fittest is the name of the game, and any trait that causes an individual to reproduce better gives that individual evolutionary fitness.
These traits are often things that simply help something to survive. After all, survival is essential for reproduction to occur. Strength, speed, height, camouflage, and many other things can be helpful. Sometimes odd things can be helpful as well. For example, a peacock’s tail has been selected for because females choose to mate with males that have a large and beautiful tail. The tail doesn’t help them to survive because it actually makes it easier for predators to catch them, but it is essential for the females to find them attractive. This is an example of sexual selection.
Let’s go back to our moth example. We ended up with different gene pools in the two populations (north vs. south). Eventually, over long stretches of time, these two different populations might change so much that they could no longer reproduce together. At that point, we would have two different species, and we could say, definitively, that the moths had evolved. Evolution occured as a consequence of random mutation and the pressure put on the population by an environmental change. Catastrophic events can speed up natural selection and adaptation. When the rules change, sometimes odd variations are selected for. Earth has undergone several mass extinction events. The tree of life would look much different if these events had not occurred.
Genetic drift is something that causes a change in the genetics of a population, but it is not natural selection. Instead, it is caused by random events that drastically reduce the number of individuals in a population. Often called the bottleneck effect or the founder effect, this occurs when only a few individuals are left to mate and regrow a population. If those individuals were the only survivors due to random luck, that means that the alleles that are present to regrow the population are just random alleles. They are not necessarily the fittest alleles. For example, if a hurricane swept through and only 4 birds were left on an island, the traits that they have will be the traits of the population going forward, even if those traits were not previously being selected for.
Gene flow can occur between different populations of the same species if individuals migrate to/from the populations. In other words, new alleles are entering/exiting and the genetic diversity will change. Small populations are more susceptible than large populations to this type of change.
The situation with our moths is an example of directional selection. One of the phenotypes was favored at one of the extremes of the normal distribution.
In other words, directional selection “weeds out” one of the phenotypes. In our case, dark moths were favored, and light moths were practically eliminated. Here’s one more thing to remember: directional selection can happen only if the appropriate allele—the one that is favored under the new circumstances—is already present in the population. Two other types of selection are stabilizing selection and disruptive selection.
Stabilizing selection means that organisms in a population with extreme traits are eliminated. This type of selection favors organisms with average or medium traits. It “weeds out” the phenotypes that are less adaptive to the environment. A good example is birth weight in human babies. An abnormally small baby has a higher chance of having birth defects; conversely, an abnormally large baby will have a challenge in terms of a safe birth delivery.
Disruptive selection, on the other hand, does the reverse. It favors both the extremes and selects against common traits. For example, females are “selected” to be small and males are “selected” to be large in elephant seals. You’ll rarely find a female or male of intermediate size.
A dog and a bumblebee obviously cannot produce offspring. Therefore, they are different species. However, a chihuahua and a Great Dane can reproduce (at least in theory). We would not say that they are different species; they are merely different breeds. In order for these to become different species of dogs, they would have to become reproductively isolated from each other. This would then allow the two groups to undergo natural selection and evolve differently. With different variation and different environmental pressures, they could each change in different ways and no longer be able to mate. This is called divergent evolution. Divergent evolution that occurs quickly after a period of stasis (little evolution) is called punctuated equilibrium. This is often due to a major event. A change that comes about after many smaller changes is called gradualism and it takes hundreds or millions of years. You should also be aware of adaptive radiation, which occurs when a species rapidly diversifies due to an abundance of available ecological niches suddenly opening up.
Divergent evolution results in closely related species with different behaviors and traits. As with our example, these species often originate from a common ancestor. More often than not, the “engine” of evolution is cataclysmic environmental change, such as pollution in the case of the moths. Geographic barriers, new stresses, disease, and dwindling resources are all factors in the process of evolution. Pre- and post-zygotic barriers also prevent organisms of two different species from mating to produce viable offspring. Pre-zygotic barriers prevent fertilization. Examples of this kind of barrier include temporal isolation, which occurs when two species reproduce at different times of the year. A post-zygotic barrier is related to the inability of a hybrid organism to produce offspring. For example, a horse and a donkey can mate to produce a mule, but mules are sterile and therefore cannot produce a second generation.
Convergent evolution is the process by which two unrelated and dissimilar species come to have similar (analogous) traits, often because they have been exposed to similar selective pressures. Examples of convergent evolution include aardvarks, anteaters, and pangolins. They all have strong, sharp claws and long snouts with sticky tongues to catch insects, yet they evolved from three completely different mammals.
There are two types of speciation: allopatric speciation and sympatric speciation. Allopatric speciation simply means that a population becomes separated from the rest of the species by a geographic barrier so that the two populations can’t interbreed. An example would be a mountain that separates two populations of ants. In time, the two populations might evolve into different species. If, however, new species form without any geographic barrier, it is called sympatric speciation. This type of speciation is common in plants. Two species of plants may evolve in the same area without any geographic barrier. Often plants undergo speciation due to polyploidy, which occurs when they get doubles of their chromosomes.
Mendel’s laws can extend to the population level. Suppose you caught a bunch of fruit flies—about 1,000. Let’s say that 910 of them were red-eyed and 90 were green-eyed. If you allowed the fruit flies to mate and counted the next generation, we’d see that the ratio of red-eyed to green-eyed fruit flies would remain approximately the same: 91 percent red-eyed and 9 percent green-eyed. That is, the allele frequency would remain constant. At first glance you may ask, how could that happen?
The Hardy-Weinberg law states that even with all the shuffling of genes that goes on, the relative frequencies of genotypes in a population are constant over time. Alleles don’t get lost in the shuffle. The dominant gene doesn’t become more prevalent, and the recessive gene doesn’t disappear.
Hardy-Weinberg law says that a population will be in genetic equilibrium only if it meets these five conditions: (1) a large population, (2) no mutations, (3) no immigration or emigration, (4) random mating, and (5) no natural selection.
When these five conditions are met, the gene pool in a population is pretty stable. Any departure from them results in changes in allele frequencies in a population. For example, if a small group of your fruit flies moved to a new location, the allele frequency may be altered and result in evolutionary changes.
Let’s say that the allele for red eyes, R, is dominant over the allele for green eyes, r. Red-eyed fruit flies include homozygous dominants, RR, and heterozygous, Rr. The green-eyed fruit flies are recessive, rr. The frequency of each allele is described in the equation below. The allele must be either R or r. Let “p” represent the frequency of the R allele and “q” represent the frequency of the r allele in the population.
p + q = 1
This sum of the frequencies must add up to one. If you know the value of one of the alleles, then you’ll also know the value of the other allele. This makes sense because Hardy-Weinberg works ONLY for populations with 2 alleles that have normal dominant-recessive behavior. So, if you know there are only 2 alleles possible and you know that 70% are the dominant allele, then the remaining 30% must be the recessive allele.
We can also determine the frequency of the genotypes in a population using the following equation.
p2 + 2pq + q2 = 1
In this equation, p2 represents the homozygous dominants, 2pq represents the heterozygotes, and q2 represents the homozygous recessives.
Both of these equations are listed on the AP Biology Equations and Formulas sheet. But how do you use them? Use proportions in the population to figure out both the allele and genotype frequencies. Let’s calculate the frequency of the genotype for green-eyed fruit flies. If 9% of the fruit flies are green-eyed, then the genotype frequency, q2, is 0.09. You can now use this value to figure out the frequency of the recessive allele in the population. The allele frequency for green eyes is equal to the square root of 0.09—that’s 0.3. If the recessive allele is 0.3, the dominant allele must be 0.7. That’s because 0.3 + 0.7 equals 1.
Using the second equation, you can calculate the genotypes of the homozygous dominants and the heterozygotes. The frequency for the homozygous dominants, p2, is 0.7 × 0.7, which equals 0.49. The frequency for the heterozygotes, 2pq, is 2 × 0.3 × 0.7, which equals 0.42. If you include the frequency of the recessive genotype—0.09—the numbers once again add up to 1.
There have been many extinctions throughout Earth’s history. The rate of extinction can vary and is usually accelerated during ecological stress. Humans have been a large ecological stressor and have likely contributed to extinction events. A population with high genetic diversity is more protected from extinction events because there is a higher chance that an individual will have the characteristics required to survive the ecological stress leading to the extinction event. When a species becomes extinct, the niche it occupied can become available to other species.
The Earth formed approximately 4.6 billions years ago. Prior to 3.9 billion years ago, the Earth was likely an extremely hostile place. The earliest evidence of life is approximately 3.5 billions years ago, but how did life arise? This is still a hotly debated topic among scientists. Most scientists believe that the earliest precursors of life arose from nonliving matter (basically, gases) in the primitive oceans of the earth. But this theory didn’t take shape until the 1920s. Two scientists, Alexander Oparin and J. B. S. Haldane, proposed that the primitive atmosphere contained mostly inorganic molecules and was rich in the following gases: methane (CH4), ammonia (NH3), hydrogen (H2), and water (H2O). Interestingly enough, there was almost no free oxygen (O2) in this early atmosphere. They believed that these gases collided, producing chemical reactions that eventually led to the organic molecules we know today.
This theory didn’t receive any substantial support until 1953. In that year, Stanley Miller and Harold Urey simulated the conditions of primitive Earth in a laboratory. They put the gases theorized to be abundant in the early atmosphere into a flask, struck them with electrical charges in order to mimic lightning, and organic compounds similar to amino acids appeared!
But how do we make the leap from simple organic molecules to more complex compounds and life as we know it? Since no one was around to witness the process, no one knows for sure how (or when) it occurred. It is likely that the original life-forms were simply molecules of RNA. This is called the RNA-world hypothesis. RNA can take many shapes because it is not restricted to be a double helix. It is possible that RNA molecules capable of replicating and thus passing along themselves (their genome) were the first life-forms. Complex organic compounds (such as proteins) must have formed via dehydration synthesis. Simple cells then used organic molecules as their source of food. Over time, simple cells evolved into complex cells.
Let’s finish by defining a few more key terms. All organisms capture and store free energy for use in biological processes. This can be done two different ways. Living organisms that rely on organic molecules for food are called heterotrophs (consumers). For example, we’re heterotrophs, but the earliest heterotrophs were simple unicellular life-forms. Heterotrophs metabolize carbohydrates, lipids, and proteins, and then hydrolyze them as sources of free energy. Eventually, some life-forms found a way to make their own food—most commonly through photosynthesis. These organisms capture free energy present in sunlight and are called autotrophs (producers). Early autotrophs (most likely cyanobacteria) are responsible for Earth’s oxygenated atmosphere.
The heterotroph hypothesis suggests that the first cells were likely heterotrophic and would have fed on organic molecules that had been made without cells. These cells likely survived by performing processes similar to glycolysis and fermentation. Once autotrophs were producing and releasing oxygen, aerobic respiration followed.
evolution
natural selection
Charles Darwin
Jean-Baptiste de Lamarck
paleontology
biogeography
flora
fauna
embryology
comparative anatomy
homologous structures
analogous structures
molecular biology
continuing evolution
phylogenetic tree (cladogram)
out-group
genetic variability
peppered moths
environmental pressure
random mutation
adaptation
evolutionary fitness
sexual selection
genetic drift
bottleneck (founder effect)
gene flow
directional selection
stabilizing selection
disruptive selection
species
reproductively isolated
divergent evolution
punctuated equilibrium
adaptive radiation
pre-zygotic barriers
post-zygotic barriers
convergent evolution
speciation
allopatric speciation
sympatric speciation
polyploidy
Hardy-Weinberg law
Alexander Oparin and J. B. S. Haldane
Stanley Miller and Harold Urey
RNA-world hypothesis
heterotrophs (consumers)
autotrophs (producers)
-
Charles Darwin made the following key observations that led to his theory of natural selection:
-
Each species produces more offspring than can survive.
-
Offspring compete with each other for limited resources.
-
Organisms in every population vary.
-
The offspring with the most favorable traits are most likely to survive and reproduce.
-
Evidence for evolution includes:
-
fossils
-
biogeography
-
comparison of developmental embryology
-
comparative anatomy, including homologous and analogous structures
-
molecular biology (sequences of genes are conserved across many types of species)
-
Members of a species are defined by the ability to reproduce fertile offspring.
-
Evolution and speciation occur when environmental pressures favor traits that permit survival and reproduction of selected individuals in a varied population. The favored traits may evolve to cause divergent or convergent evolution.
-
Adaptations are random variations in traits that end up being selected for until they become prominent in the population.
-
Genetic drift occurs when a population’s traits change due to random events rather than natural selection.
-
Selection can be stabilizing if an average phenotype is preferred, disruptive if the extremes are preferred, and directional if one extreme is preferred.
-
Hardy-Weinberg equations can be used to determine genetic variation within a population. These equations are as follows:
-
p + q = 1
Frequency of the dominant (p) and recessive (q) alleles
-
p2 + 2pq + q2 = 1
Frequency of the homozygous dominants (p2), heterozygotes (2pq), and homozygous recessives (q2)
-
Hardy-Weinberg equations describe a population that is not evolving but is instead said to be in genetic equilibrium. Such a population will be so only if it is not violating any of the following five conditions (almost all populations are evolving due to a violation of one or more of these factors):
Answers and explanations can be found in Chapter 15.
1. The eye structures of mammals and cephalopods such as squid evolved independently to perform very similar functions and have similar structures. This evolution is an example of which of the following?
(A) Allopatric speciation
(B) Sympatric association
(C) Divergent evolution
(D) Convergent evolution
Questions 2–5
refer to the following graph and paragraph.
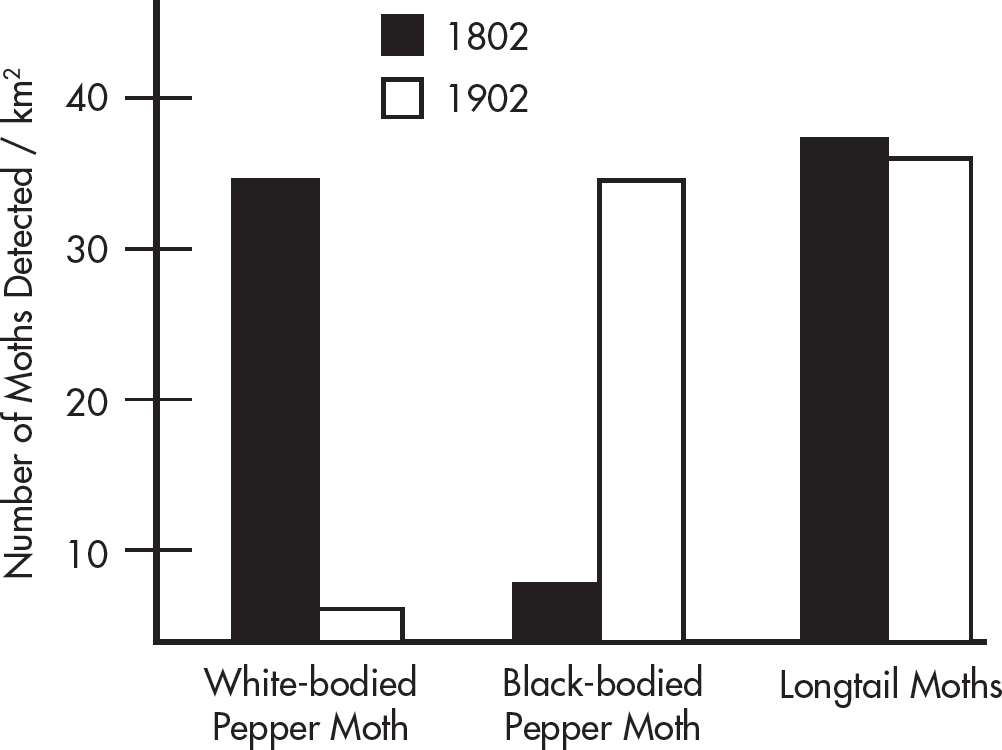
During the Industrial Revolution, a major change was observed in many insect species due to the mass production and deposition of ash and soot around cities and factories. One of the most famous instances was within the spotted moth population. An ecological survey was performed in which the number of spotted moths and longtail moths were counted in 8 different urban settings over a square kilometer in 1802. A repeat experiment was performed 100 years later in 1902. The results of the experiment are shown below.
2. What type of selection is represented by the results of this study?
(A) Stabilizing selection
(B) Directional selection
(C) Disruptive selection
(D) Divergent selection
3. Which of the following statements best explains the data?
(A) As time passed from 1802 to 1902, the frequency of white-bodied pepper moths increased and black-bodied pepper moths decreased.
(B) As time passed from 1802 to 1902, the frequency of white-bodied pepper moths decreased and black-bodied pepper moths increased.
(C) As time passed from 1802 to 1902, the frequency of white-bodied pepper moths and black-bodied pepper moths both increased.
(D) As time passed from 1802 to 1902, the frequency of white-bodied pepper moths and black-bodied pepper moths both decreased.
4. Why was the population of longtail moths also surveyed in this study?
(A) Variations in the environment were expected to alter the population of longtail moths.
(B) Longtail moths were included as a control because they were not expected to change appreciably due to changes associated with the Industrial Revolution.
(C) The peppered moth did not have a long enough tail to visualize.
(D) Longtail moths were poisoned by the coal dust and suffered drastic population loss.
5. How would the results of this study have been different if factories produced white or light gray ash and soot rather than black?
(A) There would be no change to the results of the experiment.
(B) There would have been added selection pressure for more white-bodied spotted moths and against black-bodied spotted moths.
(C) There would have been added selection pressure for more black-bodied spotted moths and against white-bodied spotted moths.
(D) There would have been an increase in the frequency of both black-bodied and white-bodied spotted moths.
6. The Middle East blind mole rat (Nannosplalax ehrenbergi) lives in the Upper Galilee Mountains of Israel. Two groups of these mole rats live in the same region, but scientists discovered that there is a 40% difference in mitochondrial DNA between these two groups. These rats do not seem to interbreed in the wild. This is an example of
(A) allopatric speciation
(B) sympatric speciation
(C) pre-zygotic barrier
(D) hybrid zone
Questions 7 and 8
refer to the following scenario.
A cruise ship with 500 people on board crashes on a deserted island. They like the island so much that they decide to stay. There is a small number of people on board with red hair, an autosomal recessive trait. Assume that red hair provides no increase in fitness.
7. What is most likely to occur with respect to the number of red-haired individuals on the island?
(A) There will be no change in the overall number of red-haired people in the population over time.
(B) Convergent evolution will occur, with all people eventually becoming red-haired.
(C) It is likely that over many generations, the proportion of red-haired people in the population will increase.
(D) Red-haired people will probably decrease in the population over many generations.
8. Evolution does not occur when a population is said to be in equilibrium by the Hardy-Weinberg definition. In the situation above, which of the following does NOT support the claim that evolution will occur in this population?
(A) The situation describes a small population.
(B) Mutations occur in this population.
(C) People do not usually mate randomly.
(D) Natural selection pressures do not exist on this island.
9. Peacock males have evolved to have huge, beautiful tails with numerous large eye spots. Long, colorful tails are difficult to carry when you are running from a predator. However, peahens (female peacocks) have been shown by researchers to mate preferably with males with more eye spots and longer tails, despite these traits making the males more susceptible to predation. Over time, this preference has resulted in peacocks with huge tails due to
(A) sexual selection
(B) disrupted selection
(C) divergent evolution
(D) sympatric speciation
Respond to the following questions:
-
Which topics from this chapter do you feel you have mastered?
-
Which content topics from this chapter do you feel you need to study more before you can answer multiple-choice questions correctly?
-
Which content topics from this chapter do you feel you need to study more before you can effectively compose a free response?
-
Was there any content that you need to ask your teacher or another person about?