Chapter Eight: Fluvial geomorphic disturbances and life-history traits of riparian tree species
Futoshi Nakamuraa; Satomi Inahara Nakamurab a Hokkaido University, Sapporo, Japan
b Ebetsu, Hokkaido, Japan
Abstract
Riparian forest communities survive fluvial and geomorphic disturbances through physiological and structural adaptations, utilizing geomorphic surfaces as regeneration habitats that provide suitable opportunities for seedling establishment and maturation to allow further seed production. Here, we classify a river catchment in the Asian Monsoon Belt of Japan into headwaters, braided rivers, and low-gradient meandering rivers, and we characterize the disturbance regimes and the availability of regeneration habitats in the temporal and spatial dimensions. The severity and frequency of riparian disturbances range from sediment removal and changes in soil properties to inundation, with recurrence intervals from seasonal up to hundreds or thousands of years. The dominant species exhibit unique life-history strategies that often change over life stages and are successful in adapting to the spatially and temporally variable occurrences of regeneration habitats.
Keywords
Fluvial and geomorphic disturbance; Regeneration habitat; Life history; Riparian tree
Acknowledgments
We wish to thank Drs. Hitoshi Sakio, Mahito Kamada, Tohru Nakashizuka, Gregor Auble, Edward Johnson and Kiyoko Miyanishi for their critical and helpful comments on earlier drafts of this manuscript. This research was supported by Grants in Aid for Scientific Research (Nos. 13460061, 14506039) from the Ministry of Education, Science and Culture, Japan, and a grant from the Water Resources Environment Technology Center.
Introduction
The mechanism for maintaining species coexistence and richness in a forest is one of the central issues in plant community ecology. Early works identified two important concepts to explain forest structures and compositions: niche partitioning along environmental gradients (Whittaker, 1956, 1965) and the regeneration niche (Grubb, 1977). According to the first concept of niche partitioning, tree species are arranged along complex gradients of elevation, light and edaphic conditions and form stand patches and canopy layers, thus partitioning a given habitat among the species present in the forest. The second concept of regeneration niche focuses on niche partitioning in reproduction, seed dispersal, seedling establishment, and further development of the immature plant. According to Grubb (1977), such partitioning can explain the indefinite persistence of many coexisting species. In temperate mixed deciduous forest, Nakashizuka (2001) argued that life-history traits and their variations generate niche partitioning and tradeoffs, promoting tree coexistence in a community with heterogeneous abiotic and biotic conditions.
In riparian zones, where episodic disturbances provide spatial and temporal heterogeneity, life-history traits and the availability of regeneration habitat associated with disturbance regimes may be important. Located at transition zones between terrestrial and aquatic ecosystems, riparian zones preserve a dynamic state in their interaction with streams (Swanson et al., 1982). Energy and material flows can be concentrated during fluvial and geomorphic disturbances, which occur more frequently in riparian zones relative to adjacent hillslope ecosystems, and thus their influence on forest communities is likely to be more significant in riparian zones (Junk et al., 1989). Riparian forest communities survive such intensive and frequent disturbance regimes through physiological and structural adaptations, while utilizing the materials trapped by these disturbance events in a complex of river geomorphic surfaces (i.e., secondary channel, point-bar, backswamps) as habitat for their growth and maintenance.
Previous studies have consistently demonstrated that riparian forest structure is tightly linked to spatial and temporal patterns of fluvial and geomorphic disturbances (Niiyama, 1987, 1989; Ishikawa, 1988; Hughes, 1990; Aruga et al., 1996; Nakamura et al., 1997; Robertson and Augspurger, 1999; Nakamura and Shin, 2001; Shin and Nakamura, 2005). Intensive disturbance provides exposed sediment for seedling establishment and alters the edaphic environment (e.g., soil texture, moisture, organic content, and nutrients) of the floodplains. Inundation frequencies and durations, which differ at elevations from the low-lying active channels to the upper floodplain terraces, form the elevation gradient of soil conditions, resulting in the species distribution along the elevation gradient. This species distribution on the elevation gradient can be complicated by multiple disturbance regimes, with each different disturbance frequency and turnover time often forming a mosaic pattern of vegetation patches (Suzuki et al., 2002; Shin and Nakamura, 2005). Timing, duration and frequencies of disturbance influence the processes of erosion and deposition that promote the formation of channel bars and islands as regeneration habitats (Cordes et al., 1997; Dykaar and Wingington Jr., 2000). Accordingly, many studies have reported the alteration of forest communities in flow-regulated rivers, the lack of light-demanding early colonizers leading to dominance by shade-tolerant species, grasses, shrubs, and exotic species (Auble et al., 1994; Johnson, 1998; Merritt and Cooper, 2000). Thus, temporal and spatial patterns of disturbance regimes and species responses to these patterns are the key to understanding the mechanisms of riparian forest maintenance.
The disturbance regime in riparian zones changes from headwaters to low-gradient alluvial rivers (Nakamura et al., 2000; Nakamura and Swanson, 2003). A dominant disturbance in headwater streams is mass movement, such as landslide and debris flow, characterized by its intensiveness but rare occurrence. In downstream braided reaches, seasonal flood disturbances generate more frequent high flows which form extensively developed geomorphic surfaces on floodplains. Low-gradient alluvial rivers in developing peat marshes rarely experience disturbances that result in landform alteration, although prolonged inundation in the marsh can cause a great deal of physiological stress to trees. Thus, a variety of disturbance types, frequencies, and intensities can be found at a catchment scale and may have diverse ecological consequences for riparian communities. Yet, much of the past studies have focused mainly on seasonal or relatively frequent flood disturbances in large rivers and on the responses of one group of species, the Salicaceae (e.g., Rood and Heinze-Milne, 1989; Begg et al., 1998; Friedman and Lee, 2002). The regeneration of Salicaceae forests has been threatened in flow-regulated rivers by the lack of flow regimes that promote their seedling establishment (Auble et al., 1994; Johnson, 1998; Merritt and Cooper, 2000). Life-history traits characteristic of these short-lived Salicaceae species and of flood disturbances operating at the scale of seasonal to decades have now been reasonably understood; however, such understanding falls short of allowing any generalizations on the importance and role of geomorphic disturbances at the catchment scale. There may be a range of life-history strategies to accommodate a variety of disturbance regimes, such as those with long recurrence intervals up to hundreds or thousands of years. In addition, Nakashizuka (2001) asserts that variation in life-history strategies among different life stages of a species as well as among different species is important in explaining the relationships between environmental heterogeneity and the organization of temperate mixed deciduous forest communities.
The objective of this chapter is to describe the relationship between disturbance regimes, ranging from montane headwater streams to low-gradient meandering rivers, and life-history traits of the dominant tree species from riparian forests in the Asia Monsoon Belt of Japan as an example. We define “regeneration habitat” as the light and edaphic conditions or opportunities suitable for successful seedling establishment and maturation of the young trees to allow seed production. As disturbances operate in time and space, the availability of regeneration habitat also varies in time and space. Riparian trees are specialized to disturbance regimes characteristic of their given geomorphic settings by developing life-history strategies that are adapted to the fluctuating availability of regeneration sites. Reproductive traits such as fruiting, modes of seed dispersal and their timing can be constrained by the times and locations of regeneration habitats (Clark, 1991; Strykstra et al., 2002). Structural adaptation and asexual regeneration, which can maintain the stages of post-seedling establishment, may be crucial during a period when regeneration habitat is not available. Thus, we propose that predictability in the occurrence of regeneration sites (referred to as “reliability” by Strykstra et al. (2002)), both in time and space, is the link between life-history traits and disturbance regimes at the catchment scale. We begin with a geomorphic classification of riparian zones and describe the disturbance regimes dominant in each class. The availability of regeneration habitat determined by the disturbance regimes is then characterized in temporal and spatial dimensions. We discuss how riparian trees are adapted to the temporally and spatially dynamic regeneration habitat, with a particular emphasis on life-history strategies of dominant species.
Geomorphic classification of riparian zones and disturbance regimes in a catchment
For many ecological phenomena, especially disturbance in stream networks, it is important to place the analysis in a geographic context. River landscape conditions can be classified into three dominant morphologies: headwater streams, braided rivers on alluvial fans, and low-gradient meandering rivers (Nakamura and Swanson, 2003). Headwater streams are boulder-dominated and are characterized by their high-gradient and very narrow valley floors. Braided rivers, generally observed on alluvial fans, have a relatively steep gravel bed and multiple channels. Low-gradient, meandering rivers are typical of sand-bed streams. In many regions of the world, such as central North America and central Eurasia, there is a transitional system between braided gravel-bed rivers and meandering sand-bed rivers that is characterized by meandering reaches with moderate gradients. Some studies that we cite in our discussion of life-history strategies may not fit within our classification because this transitional system is not usually developed in the Japanese river landscapes.
Headwater streams (0 ~ 3rd order streams)
A disturbance regime dominant in headwater streams is mass movement, such as landslide, debris flow, and earthflow, most of which are landform shifting due to gravity (Fig. 8.1). Mass movement is characterized by a deep layer of massive soil moving at high velocities but at extremely low frequency on the order of every 100 to 1000 years and therefore it is unpredictable. During the period of mass movement events, tree-fall and windthrow gaps are created more frequently. This type of gap formation may be localized, such as an individual tree-fall gap, or may be extended to a relatively large area by windthrow.

The depth of the soil layer in deep-seated landslides, initiated by episodic events such as an earthquake in a certain geological setting, can reach from a few to dozens of meters. These deep soil layers can move with trees that remain standing on the layers. The trees suffer from root cutting and may topple at slope failures and around landslide scars. The surviving individuals often exhibit architectural deformity and growth reduction, which can be visible as eccentric growth of their annual rings. This type of mass movement is definitely high in its magnitude and detrimental to riparian trees. While its occurrence is extremely episodic, the locations are geologically limited.
Shallow landslides may be more important for forest composition and structure because they occur more frequently and can extend over a large area. They occur in headwater basins after heavy rainfall events and generate soil mass movement of layers that range from tens of centimeters up to two meters in depth. Depending on geological and geomorphological settings, a landslide can occur at a particular site from once every few decades (Shimokawa, 1984; Yanai and Usui, 1989) to centuries (Nakamura et al., 1995). Shallow landslides typically form bedrock hollows (Tsukamoto, 1973; Dietrich and Dunne, 1978) and steep, convergent slopes (> 30 degrees) created by channel incision, then followed by being filled with colluvium (Hatano, 1974). Most of the established trees on the erosion surfaces of mass movements fall down, leaving few surviving individuals. However, where the energy of mass movement is small and the landslide deposits are shallow, trees may persist by forming adventitious roots and stems to anchor themselves on the deposits.
Shallow landslides often develop into debris flows, and their influences extend through stream networks to the lower reaches of the catchment (Nakamura et al., 2000). Consequently, a main tributary in a small basin with multiple landslide-prone headwaters would experience debris flow disturbances more frequently, i.e., at a time scale of several decades. Debris flows may stop at river confluences or in unconstrained reaches and extend over valley floors. Mass-movement deposits consist of large logs and the river-bottom substrate, and can reach boulder sizes, while creating undulating landforms that provide heterogeneous regeneration habitats.
Where geomorphic disturbances propagate through a stream network, the severity of the riparian disturbances becomes variable, thus complicating their influences on riparian forests (Nakamura et al., 2000). Debris flows initiate a sequence of geomorphological processes in the downstream direction over time, such as streambank erosion, wood transport, and flood surges. Large logs directly enhance the physical forces on riparian stands and increase the downstream flow energy by channel constriction (Swanson et al., 1998; Johnson et al., 2000). These logs can also form levees and jams, which may act as buffers that protect the existing stands. In larger channels, floating logs become mobilized (Seo and Nakamura, 2009), further influencing geomorphic processes in the downstream environments (Collins et al., 2012; Nakamura et al., 2017).
Braided Rivers on alluvial fan (4 ~ 5th order streams)
The most common type of disturbance in braided rivers is flooding (Fig. 8.1). In Asian Monsoon areas, heavy seasonal rainfalls cause flooding in both the monsoons (June to early July) and typhoons (August to early October). In the northern part of Japan, snowmelt high flow is also predominant, occurring every year in spring (April and May). The magnitude and frequencies of typhoon- and monsoon-induced flooding show greater year to year variation than snowmelt high flows.
Valley floor landforms in alluvial fan systems are typically unconstrained with relatively steep bed gradients of braided rivers. Multiple active channels migrate laterally, forming abandoned secondary channels. Bed materials, which consist of pebble and gravel size particles, are typically low in moisture content. Flood frequency is in the order of years near active channels and ten to a hundred years for more stable floodplains (Nakamura, 1990; Nakamura et al., 1995). However, lateral channel migration can alter stable floodplains, changing them to highly unstable areas prone to intensive and frequent flood disturbances.
Flood disturbances in braided rivers influence riparian trees to varying degrees of severity. Some of the trees toppled during a flood may be completely uprooted and die, while others may survive by re-sprouting from their buried shoots and stems (Everitt, 1968; Karrenberg et al., 2002). With less intensive flooding, trees that are left standing may be killed due to the anoxic conditions under sediment depositions while others may survive by forming adventitious roots (Nakamura and Kikuchi, 1996). Disturbance intensity decreases along an elevation gradient from the active channel to higher terraces in a floodplain and is determined primarily by inundation frequency (Nakamura et al., 1997). Flooding rarely reaches the high elevations and, even if it does, removal of canopy trees is rare. Only understory vegetation on the high elevation surfaces is affected, resulting in the dominance of large canopy trees there. In contrast, geomorphic surfaces at low elevations near active channels are frequently subject to flood disturbance. On these low surfaces, fully mature trees are not established while seedlings and saplings of light-demanding species are common.
When flooding completely eliminates the ground surface in a braided reach, new floodplain surfaces are formed during periods of water recession. Such a stand-replacement disturbance produces many large wood pieces, and some of them are transported and then become stranded over gravel bars. The heterogeneous site conditions created by large wood provide diverse regeneration habitats for various plant species (Pettit and Naiman, 2006; Nakamura et al., 2012). Sediment and nutrients transported from upstream accumulate to form complex geomorphic surfaces that have a variety of substrate properties and nutrient conditions. Flood disturbances can occur at various frequencies and magnitudes, resulting in further topographically and edaphically diverse soil conditions (Wolman and Miller, 1960; Dollar, 2000; Shin and Nakamura, 2005). These new geomorphic surfaces are typical recruitment sites for wind-dispersed, light-demanding species, but may not be suitable as regeneration habitat where the seedlings can survive to reproductive maturity.
Low-gradient meandering rivers (6th or higher order streams)
Geomorphic disturbances in low-gradient alluvial rivers are less extreme in magnitude than those in headwater and braided rivers. The meandering process in alluvial rivers creates unique landforms over geologic time. Natural levees adjacent to the active channels are meander belts. Point bars, the areas of fresh, sandy sediment on the inside of the meander bends, are formed one after another to create ridge and swale topography of meander scrolls. Progressively older plant communities develop on the scrolls at increasing distance from the active channel. In back swamps behind the natural levees, a layer of peat soil develops in which water tables are constantly high (Nakamura et al., 2002).
Seasonal flooding is the major disturbance in low-gradient meandering rivers (Fig. 8.1). However, compared to braided rivers, both runoff-rise and recession in meandering systems are slower with a less sharp peak. This is due partly to the low gradient of the meandering rivers but also to the ability for water retention in back swamps. Storm waters are intercepted by the sponge-like peat soil in the swamps, which have extremely low hydraulic conductivities, and released gradually to the stream channels.
Within a low-gradient meandering river, the influences of flood disturbances are different between natural levee belts and back swamps. In natural levee areas that include meander scrolls, the concave side of bends is continuously eroded by the concentration of stream flows and the occurrence of secondary flows. Trees established on the meander scroll areas therefore gradually collapse. The convex sides of the bends are depositional zones, which develop point bars available for seedling establishment. In the back swamps behind the levees, flooding that destroys trees is unlikely to occur. Most of the flooding is not accompanied by sediment movement in the back swamps. Instead of physical damage by erosion forces, flooding in the back swamps has a physiological impact caused by prolonged inundation. The dominant substrate in the back swamps is not sandy soil, as in point bar areas, but leaf litter from reeds and sedges or peat soil. Thus, because of the unique hydrology and substrate properties, the regeneration process in back swamps is expected to be very different from that on natural levees.
Disturbance, reliability of regeneration habitat and life history of dominant tree species
To evaluate the reliability of regeneration habitat, this section characterizes the occurrence of regeneration sites in terms of frequency on a temporal scale and abundance and regularity on a spatial scale. The reliability of regeneration habitat in time is evaluated as high when regeneration sites are formed on a regular basis. We assumed that regularity of disturbances was closely related to their recurrence intervals (Fig. 8.1). For example, deep-seated landslides rarely occur and exhibit great variation in their recurrence intervals. Alternatively, snowmelt flood in braided rivers recurs more frequently and is seasonal. Lower frequency represents less predictability, and vice versa. The reliability of regeneration habitat in space is high where regeneration sites are abundant and distributed regularly over the space. The evaluation is arbitrary based on comparison among different disturbance regimes.
Headwater streams
In Japan, landslides occur mostly in 0-order streams or on steep slopes (greater than 30°) underlain by granite or tertiary geology. Mass movement can create a large-scale exposure of sediment as a potential regeneration site. However, because it is highly episodic, having a wide range of recurrence intervals from a few decades to several thousand years, a long life span is one of the most important strategies. We hereafter refer to species that are particularly adapted to mass movement as mass movement adapted (MMA) species.
One MMA species comprising a cool temperate deciduous forest in Japan is Cercidiphyllum japonicum Sieb. & Zucc. ex J. Hoffman & H. Schult. This long-lived species has a very distinctive stand architecture, forming an enormous canopy with numerous shoots (Fig. 8.2). Canopy trees are distributed sporadically on landslide deposits and lower parts of V-shaped valleys. Abundant small seeds (3 mm long) are produced every year without a masting year and dispersed over a large area by wind (Kubo et al., 2000; Sakio et al., 2008). However, very few juvenile trees are found, suggesting low reliability of regeneration habitat for C. japonicum. The small seeds are incapable of germinating under leaf litter and are also susceptible to desiccation in too large open sites and in sandy soils with variable moisture contents (Seiwa and Kikuzawa, 1996; Kubo et al., 2000). While small seeds can easily attach themselves to steep slopes and logs and germinate on the exposed fine mineral soil without leaf litter, heavy rainfall and sediment erosion may wash away or bury the small seeds and seedlings on such unstable substrates. Thus, given these severe microsite constraints on seedling recruitment, the abundance of regeneration sites suitable for C. japonicum is limited and randomly distributed (Table 8.1). The occurrence of regeneration sites in time is also unreliable because of episodic disturbances in headwater streams. The annually constant seed production deals with low reliability of regeneration habitat in time, and fecundity and dispersal capability increase the chances of arriving at spatially unreliable regeneration habitat for C. japonicum. Moreover, clonal sprouting is a crucial strategy for sustaining a long life span. An individual tree of C. japonicum forms numerous basal sprouts around the main trunk and replaces the main trunk as it dies with surrounding shoots. This vegetative growth consequently maintains canopy trees for 500 to 1000 years. While sprouting appears to be encouraged by substrate instability (Kubo et al., 2001) and gap formation (Kubo et al., 2004), its ultimate result is longevity where reliability of regeneration habitat for C. japonicum is extremely low both in time and space.

Table 8.1
| blank cell | Regeneration habitat | Life-history strategy | ||||||
|---|---|---|---|---|---|---|---|---|
| Geomorphic classification | Disturbance regime | Dominant tree species | Landform and microsite requirement | Time | Space | |||
| Frequency | Abundance | Regularity | Reliability in time | Reliability in space | ||||
| Headwater streams (0–3rd order streams) | ||||||||
| Landslide, debris flow, earth flow | Cercidiphyllum japonicum | Landslide and debris flow deposits, fine mineral soil | Low | Low | Low | Sprouting, longevity, constant seed production | Abundant seed production, high dispersal capability | |
| Pterocarya rhoifolia | Landslide, debris flow deposit | Low to moderate | High | Moderate | Synchronous reproduction, fast growth | Wind dispersal, large seed | ||
| Euptelea polyandra | Talus cone, shallow landslide | Moderate | Moderate | Moderate | Sprouting, dormant bud | Wind dispersal | ||
| Fraxinus platypoda | Abandoned channel, landslide and debris flow deposit, canopy gap | Low to moderate | High and low | Moderate and low | Seedling bank | Generalist, wind dispersal, large seed | ||
| Tree-fall/windthrow gap | Aesculus turbinata | Canopy gap, terrace and toe slope | Low | Low | Low | Seedling bank | Large seed, secondary dispersal (animal dispersal) | |
| Braided rivers on alluvial fan (4–5th order streams) | ||||||||
| Large flood | Ulmus davidiana var. japonica | Upper floodplain, sand and gravel deposit | Moderate | High | High | Synchronous reproduction, longevity, seed dormancy | Wind dispersal | |
| Fraxinus mandshurica var. japonica | Synchronous reproduction, longevity, seed dormancy | Wind dispersal | ||||||
| Small flood | Salicaceae | Point bar, sand and gravel deposit | High | Low | Low | Constant seed production, timed seed dispersal, bending stability, sprouting, adventitious rooting, fast growth, early maturation | High dispersal capability (water and wind), abundant seed production | |
| Low-gradient meandering rivers (6th order streams) | ||||||||
| Flood | Saliaceae | Natural levee, sand and gravel deposit | High | High | High | Constant seed production, timed seed dispersal, bending stability, sprouting, adventitious rooting, fast growth, early maturation | High dispersal capability (water and wind), abundant seed production | |
| Waterlogged | Alnus japonica | Backwater swamp, small hummock (tussok) | Low | Low | Low | Sprouting, enhanced stem growth, hypertrophied lenticel, adventitious rooting, nitrogen fixation | Water and wind dispersal | |
| Fraxinus mandshurica var. japonica | Sprouting, enhanced stem growth, hypertrophied lenticel, adventitious rooting | Water and wind dispersal | ||||||

Pterocarya rhoifolia Sieb. & Zucc. is another MMA species that synchronously regenerates with the recurrence of mass movements (Sakio et al., 2002). Mature stands establish on large-scale debris flows and landslide deposits but can also be found in a wide range of landforms from floodplains to the adjacent terraces and slopes (Sato, 1992; Ann and Oshima, 1996; Sakio et al., 2002). Although vegetative growth and adventitious root formation have been reported (Kaneko, 1995), seedlings are the primary means of regeneration (Kaneko and Kawano, 2002). Seeds of P. rhoifolia are carried by wind and quickly colonize fresh exposed surfaces to form an even-aged stand (Oshima et al., 1990; Sato, 1992; Kaneko and Kawano, 2002; Sakio et al., 2002). Similar to C. japonicum, regeneration of P. rhoifolia relies on a low frequency of mass movement. However, its seeds are larger than those of C. japonicum and are less constrained in substrate requirements for germination and seedling establishment. Therefore, regeneration habitats suitable for P. rhoifolia should be much less limited in their spatial abundance and regularity, implying higher reliability in space. Larger seeds are more tolerant of leaf litter accumulation, thereby allowing seedling establishment on flat surfaces where the ground is less disturbed. Where young P. rhoifolia and C. japonicum co-occur, P. rhoifolia grows faster to reach the canopy (Sakio et al., 2002). The life span of P. rhoifolia is around 100 years and appears to be timed to disturbance frequencies ranging from a few decades to 100 years. Without a disturbance within this time scale, stands of P. rhoifolia are eventually replaced by species having longer life spans such as Fraxinus platypoda Oliver, C. japonicum and Aesculus turbinata Blume (Sato, 1995; Sakio et al., 2002).
Euptelea polyandra Sieb. & Zucc. can also be classified as an MMA species which establishes even-aged stands on sandy gravel deposits in landslide sites and talus cones (Sakai et al., 1995). It has small seeds, but larger than C. japonicum, that are dispersed by wind over long distances (Sakio et al., 2008). While C. japonicum and P. rhoifolia are adapted to intensive, infrequent mass movements, such as a deep-seated landslide, E. polyandra regeneration can cope with more frequent and less intensive disturbances, such as a shallow landslide. The occurrence of regeneration sites for E. polyandra is qualified as moderate in time and space, with its abundant specific locations (landslide deposits and talus cones). However, talus cones and steep slopes, in which mature stands typically occur, are subject to frequent ground disturbance (Sakai et al., 1995). E. polyandra forms multiple shoots to cope with erosion, sediment burial, and soil creep on the unstable slopes. The species starts sprouting at relatively early stages of its life history and constantly forms dormant buds to respond quickly to the frequent soil disturbance. Sprouting certainly has an important role in repairing damage by disturbances. Roots also are well adapted to anchor on steep slopes, protecting the trees from uprooting. These structural adaptations consequently contribute to sustaining a life span while regeneration habitat is not available.
Some species in headwater streams may depend on gap formation (tree-fall and windthrow). Although gap formation can be more frequent than mass movement in a forest (Fig. 8.1), its abundance is low with irregular distribution in space (Table 8.1). Canopy gaps are low in light intensity, requiring shade tolerance of species in their seed and seedling stages. High moisture conditions of forest soil and thick leaf litter in the gaps make seeds and seedlings vulnerable to disease and herbivory (Hoshizaki et al., 1997). We refer to species that thrive in these conditions as gap phase (GP) species.
Aesculus turbinata is the most typical GP species in headwater riparian zones in Japan. This species preferentially occurs on stable surfaces of floodplain terraces and their adjacent slopes (Oshima et al., 1990; Sato, 1995; Kaneko et al., 1999). A prominent reproductive characteristic of A. turbinata is its large seed size (6.2 g dry weight) (Hoshizaki et al., 1997). The heavy seeds fall under parent crowns and are secondarily dispersed by rodents. This secondary seed dispersal plays a vital role in avoiding light-limited seedling mortality under parent trees as well as potentially increasing the chance of reaching a regeneration site (Hoshizaki et al., 1999). A large seed reserve enables the seedlings to grow above the leaf litter, which is crucial for successful seedling establishment (Sato, 1995; Hoshizaki et al., 1997; Kubo et al., 2000; Nakashizuka, 2001). A. turbinata seedlings also are shade-tolerant, growing in low light conditions (Ishida and Peters, 1998).
The last species representing headwater riparian zones is Fraxinus platypoda. The most prominent feature of this species is its generalist strategy in regeneration site requirements, exhibiting a characteristic of both MMA and GP species (Table 8.1). Similar to E. polyandra, a seed of F. platypoda is relatively large (30 mm in length) and capable of germinating on various substrates, ranging from sandy pebbles and logs to leaf litter (Kubo et al., 2000). Seedling regeneration forming even-aged stands often occurs on sandy gravel deposits in abandoned channels that are protected from frequent flooding (Sakio, 1997). F. platypoda can live around 180 years and is adapted to large-scale disturbances such as an earthquake-induced mass movement. Among the MMA species considered in this study, F. platypoda is the most shade tolerant and can form a seedling bank under a closed canopy. Seedlings grow into canopy trees when the forest canopy opens. F. platypoda shows strong dominance in headwater streams in central Japan, which may be a result of its generalist strategy between GP and MMA (Sakio, 1997; Sakio et al., 2002).
Although we found many riparian forest studies of headwater streams in Japan, no comparable examples were found for other regions. Gom and Rood (1999) compared reproductive strategies of two Populus sections and their hybrid between headwaters and the lower Great Prairie regions in Alberta, Canada. Clonal sprouting was encouraged in the headwater reach, partly by erosion, sedimentation, ice scouring and flow forces during high flows. By contrast, seedling reproduction appeared to be more common in meandering and braided rivers in the lower basins where frequent flood disturbances promote seedling regeneration. Flood disturbances in the headwater site seemingly do not eliminate the canopy trees and, thus, regeneration sites rarely occur. As inferred for C. japonicum, Gom and Rood concluded that clonal growth was important for the maintenance of cottonwood forests where regeneration habitat was unreliable both in time and space.
In summary, the species that are common in Japanese headwater riparian zones can be largely grouped as MMA or GP species. High dispersal potential either by wind or animals is a strategy adapted to increase the chances of arriving at regeneration habitat (Table 8.1). Generally, life spans are long, ranging from 100 to 500 years and reflecting the long interval between disturbances in headwater systems. The life span of P. rhoifolia appears to be synchronized with disturbance frequency, while this assumption does not appear to be applicable to C. japonicum. C. japonicum can reach the canopy only in sites that remain undisturbed beyond the life span of P. rhoifolia. Seed size is associated with microsite requirements (soil moisture, light availability, and substrate texture and stability). The ultimate function of sprouting is to sustain longevity on unstable substrates. For gap species, shade tolerance and the seedling bank strategy is vital. A facultative strategy between GP and MMA may have a role in adapting species to low reliability of regeneration sites in time and space.
Braided rivers on alluvial fans
One disturbance typical of braided rivers is large-scale flooding, which can extend across the floodplains and form various geomorphic surfaces that promote riparian forest regeneration. Compared to headwater streams, flood disturbance frequently occurs on a more regular basis in braided rivers because of a combination of seasonal snowmelt and more erratic Asian monsoon and typhoon high flows. It is important for species to increase their reproductive potential by timing periods of recruitment and maturation to the periodic floods. Because of the periodic creation of recruitment habitats, seedling regeneration with a short life span should be favored in braided rivers.
Depending on the elevation within a floodplain, the factors influencing reliability of regeneration habitat are different (Table 8.1). At high elevations reachable by relatively large floods, regeneration sites occur moderately often. Near active channels, the frequency of regeneration site occurrence is much higher because of shorter flood intervals. However, seedlings established in the active channel may be washed away by frequent and extensive channel migrations, thereby limiting the abundance and location of regeneration habitat. Therefore, seedling establishment in lower floodplains requires morphological tolerance to high flows; it is not useful to have strategies to escape in time, such as seedling bank formation, vegetative growth to reach maturation, or a long-life span. Species adapted to lower floodplain environments may extend their roots deep into the sediment to protect them from physical damage or from being flushed, or may bend themselves with flexible stems. They can also form adventitious roots and shoots after being buried by sediment.
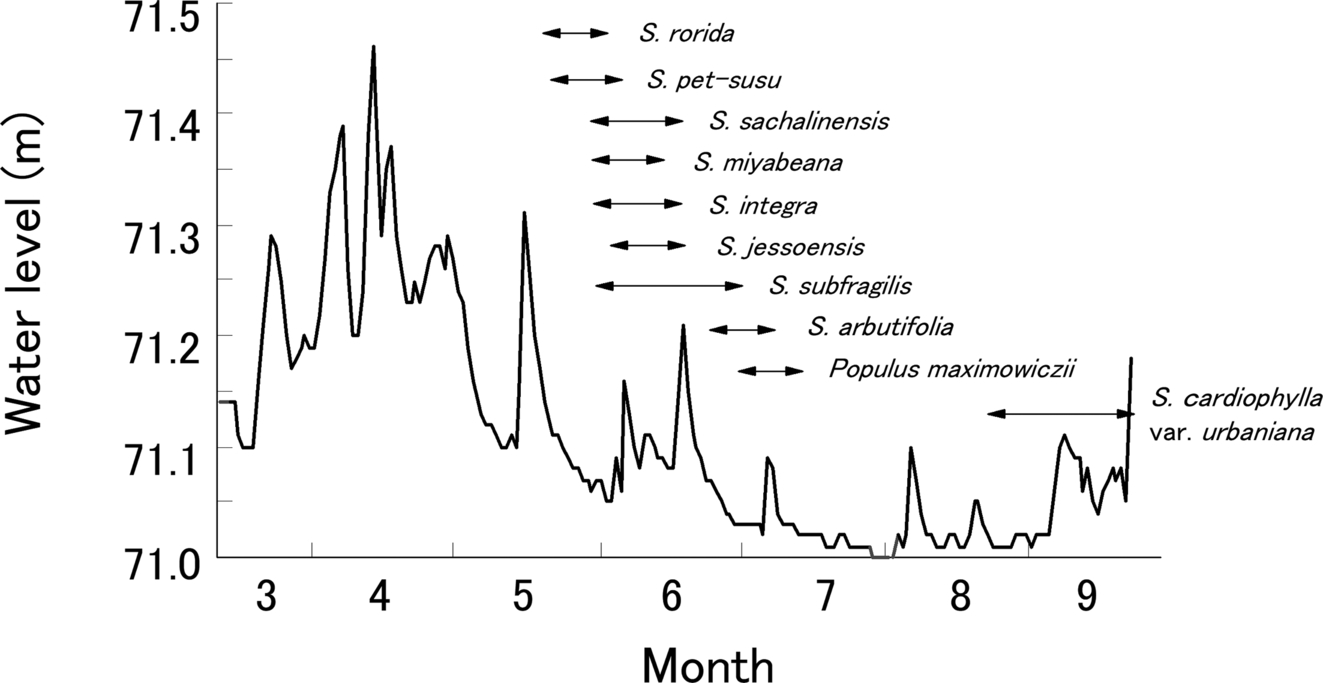
These life-history strategies can be seen in most species in riparian zones of braided reaches in Japan. The Salicaceae is dominant in braided rivers in Japan and comprises four genera and 17 species (Niiyama, 2002). Each species segregates its habitat along a stream with some overlapping. Fig. 8.3 shows a Salicaceae-dominant forest on the low floodplain of a braided river in an alpine region of Japan, where frequent channel migration and multiple channel development occur. Salicaceae trees reach reproductive maturity quickly, within a period that coincides with flood recurrence in the order of years to a decade (from 7.8 to 13.9 years on average, Nakamura et al., 2007). Their extremely light seeds (38–600 mg for 1000 seeds) are produced prolifically every year and are easily dispersed over a long distance by wind (Satoh, 1955; Niiyama, 2002). Seed germination requires an exposed surface with mineral soil consisting of sandy pebbles, which is formed extensively by flood disturbances. Seedling establishment requires abundant light and high soil moisture content; thus, leaf litter under forest canopies and herbaceous layers do not provide suitable regeneration habitat. Seeds are released during the period of the descending limb in a snowmelt flood, between May and September, with slightly different timing among the Salicaceae species (Niiyama, 1990; Nagasaka, 1996; Fig. 8.4). Salix rorida Lacksch. begins seed dispersal the earliest (mid-May) and S. subfragilis Anders. the latest (June to early July). These differences in dispersal times by genus are rather distinct. Seed dispersal of Salix arbutifolia Pallas and Populus maximowiczii Henry occurs from late June to early July, which is later than those of Salix spp.; that of Salix cardiophylla Trautv. & Mey. var. urbaniana (Seemen) Kudo is even later, from August to September. Species with seed dispersal in the early periods of a snowmelt flood may take advantage of less competition, but may also be flushed away during small flood peaks following the major flooding (Fig. 8.4). Contrastingly, the seeds dispersed in the later periods may be protected from high flows but are subject to higher competition from early colonizers. Thus, there is a trade-off in the seed dispersal strategy associated with reliability of regeneration habitat.

The Salicaceae has extremely short seed longevity, and thus the period of seed dispersal is critical (Karrenberg et al., 2002); it is known that spring floods promote its recruitment in Europe and North America (Fenner et al., 1985; Rood and Mahoney, 1990; Nilsson et al., 1991). For other species, Pettit and Froend (2001) reported seed dispersal adapting to flow dynamics in western Australia. They compared the reproductive characteristics of Eucalyptus and Melaleuca species in two rivers with contrasting flow dynamics (the Ord River and Blackwood River). Seed fall occurred in the periods of receding high flows in both rivers for both species. This timed seed fall can increase the chance of reaching regeneration habitat in time, as commonly seen in braided and meandering rivers.
However, a study in a regulated braided river, the Platte River in Colorado, demonstrated that small spikes of flows after cottonwood (Populus) germination greatly impacted their survival within a year (Johnson, 2000). Although seedling recruitment could occur extensively on alluvial surfaces after major floods, trees surviving for a year were very few. Regeneration sites occurred seasonally but with irregular and sparse distribution because of frequent high flows; these results are in agreement with the characteristics of regeneration habitat in low floodplains of Japanese braided rivers (Table 8.1).
On upper floodplains in the same braided rivers, although regeneration habitats may be abundant, their occurrence is not frequent because of high elevations (Table 8.1). Dominant species are relatively shade-tolerant, such as Ulmus davidiana var. japonica (Sarg. ex Rehder) Nakai and Fraxinus mandshurica Rupr. var. japonica Maxim. Seedlings of these species are less light-demanding and can establish under canopy trees. However, in most cases, the opportunities for germination under canopy trees are limited because of shading, disease, herbivory, and in some cases, desiccation above the leaf litter. Successful seedling establishment is more likely to occur on exposed mineral soil, thus relying on flood disturbances that can eliminate the forest canopy or, at least, the forest floor vegetation. Similar to the Salicaceae, time to reproductive maturity of these species seems to coincide with the longer interval flood frequencies of the upper floodplains (e.g., 47 years for F. mandshurica var. japonica (Nakamura et al., 2007; Fig. 8.1). Their seeds are heavier than those of the Salicaceae and dispersed by wind to a limited distance from the parent trees. These species grow more slowly but live longer than the Salicaceae; e.g., U. davidiana var. japonica can live for about 200 to 300 years (Kon and Okitsu, 1999).
Given less frequent recruitment opportunity on upper floodplains, a strategy to thrive within canopy stands may be important to some extent. It has been shown that U. davidiana var. japonica exhibits delayed germination (Seiwa, 1997). Its seeds are dispersed in summer and germinate in the same growing season. However, some of the seeds undergo dormancy during the winter and germinate early the following spring. This delayed germination takes advantage of the better light conditions before the leaves of the deciduous canopy trees are developed. It can also dilute the variation in seed production (mast seeding) and avoid sibling competition (Shibata and Nakashizuka, 1995).
Timed dispersal in the abovementioned Australian study by Pettit and Froend (2001) may be considered as another example of reducing seedling mortality under parent trees. Seasonal high flows in the Blackwood River were less intensive, and seed dispersal of Eucalyptus rudis Sm. and Melaleuca rhaphiophylla Schauer depended upon wind; seed density was a function of the distance from the parent trees. However, some portions of the seeds tended to be retained in the parent trees and released throughout the year. By contrast, wind-dispersed seeds of E. camaldulensis Dehn. and M. leucadendra (L.) L. in the Ord River, with its erratic and intensive flooding, appeared to be further transported by currents, presumably due to much greater flow forces (about 30 times greater average maximum discharge than the Blackwood River). Seed dispersal by river flows is less efficient in the Blackwood River and, therefore, “delayed dispersal” by retention in the parent trees can contribute to avoidance of sibling competition and seedling mortality under canopy trees. This “delayed dispersal” may be an adaptation to temporally unreliable regeneration sites near parent trees, which is also assumed for “delayed germination” by Ulmus davidiana var. japonica.
In summary, both the Salicaceae and shade-tolerant species depend upon flood disturbances for their seedling establishment. Both groups exhibit a strategy to increase their chances of successful regeneration by synchronizing their maturation time to flood frequencies. Life span is longer for the shade-tolerant species than the Salicaceae, reflecting the difference in flood frequencies between the upper and lower floodplains. Furthermore, seed dispersal of the Salicaceae is timed to seasonal snowmelt flooding to increase their reproductive potential.
Low gradient meandering rivers
Flood disturbance in low-gradient meandering rivers is generally less destructive and more predictable in its spatial patterns. Intensive flood disturbances that cause large-scale landform alteration rarely occur. As described previously, two distinctive types of riparian habitats are formed: natural levees and backwater swamps. On natural levees, seedlings on the concave parts of a meander reach are subject to erosion and eventually being washed away. On the contrary, the convex sides develop point-bars and become regeneration sites because severe erosion is unlikely to occur there. Thus, the location of a regeneration site is predictable in natural levees, and abundance of regeneration sites is moderate. Overall, reliability of regeneration habitat in space is high in natural levees (Table 8.1). Seasonal high flows, which occur by snowmelt and monsoon rainfall, increase the temporal opportunity for successful regeneration in natural levees.
In contrast, backwater swamps behind the natural levees are less likely to support reliable regeneration habitat because of long-term inundation. Swamp habitats are characterized by high water tables synchronizing with adjacent river stages and by fine sediment deposition due to overbank flooding. Vegetation in the backwater swamps, which typically is a cover of herbaceous species such as sedges and reeds, is adapted to anoxic conditions. There are few open sites available for tree seedling establishment. Intensive high flows that can eliminate the herbaceous layer are rare. Accordingly, the occurrence of regeneration sites in backwater swamps is extremely rare both in time and space (Table 8.1). As for MMA species in headwater riparian forests, high dispersal ability and long life spans might be important.
Dominant species on natural levees are members of the Salicaceae, in particular, Salix pet-susu Kimura and S. subfragilis in cool temperate zones in Japan (Nakamura et al., 2002). Seed dispersal of these two species is timed to water receding periods after snowmelt high flows, by which seedlings successfully establish on point bars. Abundant seeds are dispersed by stream flow and wind. At the margins between point-bars and the stream current, water depth and velocity are greatly reduced, allowing the floating seeds to colonize on the point bars. Therefore, the Salicaceae seedlings are distributed in a belt on point-bars along an active channel. This spatial pattern of cottonwood stands associated with lateral point-bar formation has also been described for meandering rivers in the USA (Everitt, 1968; Auble and Scott, 1998; Robertson and Augspurger, 1999; Merritt and Cooper, 2000). As periodical and moderate flooding grow convex point-bars, cottonwood seedlings become established, forming a predictable stand structure with increasing age with distance from the active channel.
One representative species in cool temperate backwater habitats in Japan is Alnus japonica (Thunb.) Steud. This species often grows in a dwarfed form with numerous shoots in sedge-reed communities (Fig. 8.5). The above ground plant consists of a single shoot in the first generation which dies in a few decades. The remaining trunk again sprouts numerous shoots (Yamamoto, 2002) and continues increasing the number of shoots outward, forming an alder thicket in a dwarfed form (Yabe and Onimaru, 1997). Recent studies suggested that anoxic conditions and/or nutrient poor conditions caused this vegetative growth (Yamamoto, 2002; Nakamura et al., 2002). In such a stressful environment, plants could survive by keeping their aboveground biomass at a low level with a high turnover of repeating asexual regeneration (Yamamoto, 2002). This vegetative growth also complements poor seedling regeneration in backwater habitats where regeneration habitat is highly unreliable both in space and time. Although seed dispersal of A. japonica occurs by both wind and water, seedling establishment is difficult due to shading and constant inundation within a sedge-reed community. In deepwater swamps in the southeastern USA, viable bald cypress (Taxodium distichum (L.) L.C. Rich.) seeds are transported by water potentially for their buoyancy periods up to nearly 80 days and trapped by emergent substrates (Schneider and Sharitz, 1988). Seed germination requires occasional drawdowns, and otherwise floating vegetation mats can be important regeneration sites (Huffman and Lonard, 1983). Likewise, it has been proposed that floating peat mats provide regeneration sites for A. japonica (Yabe and Onimaru, 1997). In a mesotrophic mire of northern Japan, A. japonica seedlings establish on small hummocks (tussock), where leaf litter is thinner and light and temperature conditions are favorable (Shida and Nakamura, 2011). The species exhibits various structural adaptations to anoxic conditions of backwater swamps, including enhanced stem growth, development of hypertrophied lenticels and adventitious root formation on submerged stems (Yamamoto et al., 1995a, 1995b). Hypertrophied lenticels and adventitious roots belowground can compensate for reduced oxygen supply and increase the efficiency of water absorption (Yamamoto et al., 1995b). The ability for nitrogen fixation has also been known for this species.

Fraxinus mandshurica var. japonica often occurs with A. japonica in backwater swamps in cool temperate forests in Japan. Although the species exhibits similar traits such as adventitious root formation and vegetative growth to overcome anoxic conditions (Yamamoto et al., 1995a) and probably is adapted to unreliable regeneration habitat, the extent of its functional ability seems to be limited. For example, while A. japonica can continuously develop adventitious roots from spring through early fall, F. mandshurica var. japonica stops forming them in summer (Yamamoto, 2002). In addition, the larger seeds of F. mandshurica var. japonica may restrict its dispersal potential by water. Limited structural adaptation and ineffective seed dispersal may confine F. mandshurica var. japonica to being subdominant to A. japonica in backwater swamps.
In summary, the reliability of regeneration habitat is distinctly different between natural levees and backwater swamps. In natural levee habitats, regeneration sites (point-bars) are formed constantly in a predictable manner, both in time and space. The Salicaceae species dominate natural levee habitats using their strategies of fast growth, timed reproduction and seed dispersal that utilize the directional flood disturbance in meandering alluvial rivers, as well as their structural adaptation to tolerate the high flows. A. japonica thrives in backwater swamps where sedge-reed communities are dominant. Structural adaptation and vegetative growth, particularly in a dwarfed form, enable the species to persist in waterlogged and oligotrophic wetlands where regeneration habitat reliability is extremely low, both in time and space.
Concluding remarks
The riparian forests in the Asia Monsoon Belt of Japan are subject to a variety of geomorphic and fluvial disturbances that can vary longitudinally at a catchment scale. This is because of combined seasonal and extreme floods caused by snowmelt, heavy rainfalls initiated by monsoons and typhoons, as well as the high gradient topography and extensive tectonic area. The temporal and spatial availability of regeneration sites can also vary depending upon the disturbance regime. Dominant species thus exhibit various life-history strategies to cope with the temporal and spatial reliability of regeneration habitat, which is determined by disturbance regimes in a dynamic state. The specific strategies for each life-history stage that were identified in this study are listed in Table 8.2. These strategies are found in three geomorphic types of habitat that are distinguished by their disturbance regime (Table 8.1). For all types of riparian systems, longevity and wind dispersal are the prominent strategies to cope with regeneration habitat that is unreliable in time and space. In habitats where reliability of regeneration habitat is extremely low both in time and space, vegetative growth is a vital strategy to complement seedling regeneration. Structural adaptation is common for most species. For some species, the periods of maturation and seed dispersal coincide with disturbance frequencies and timing to increase their reproductive potential both in time and space.
Table 8.2
| Life-history stage | Strategies |
|---|---|
| Seed dispersal | Dispersal by wind over an extensive area |
| Secondary dispersal by animals | |
| Seed provision on point bars by water | |
| Synchronization between the timings of seed dispersal and snowmelt flood | |
| Germination, seedling and sapling | Large seed |
| Seed dormancy | |
| Fast growth | |
| Seedling/sapling bank | |
| Adventitious roots | |
| Bending ability | |
| Enhanced stem growth | |
| Hypertrophied lenticels | |
| Reproductive mature | Synchronization between maturation time and disturbance frequency |
| Longevity | |
| Sprouting | |
| Enhanced stem growth | |
| Hypertrophied lenticels | |
| Adventitious roots |

As predicted by Grubb's (1977) concept of regeneration niche, species colonize riparian zones by adapting to the reliability of regeneration habitat with different strategies at each developmental life stage. The variety of disturbance regimes promotes a diverse array of life-history strategies that differ among species and life stages, thus contributing to species coexistence in riparian forests (Nakashizuka, 2001).
Focusing on life-history strategies and the likelihood of regeneration habitat occurrence, we were able to describe the relationships between a number of species and disturbance regimes. While we included various disturbance regimes at a catchment scale, our review heavily relied upon examples of the Japanese rivers. Some of these examples may be comparable to the studies in other regions. However, much of the riparian studies in other regions, particularly in North America, lack variation in tree species and are limited to low gradient alluvial rivers. Thus, roles of mass-movement type disturbances have been poorly studied. Further efforts in these gaps are required to develop a generality of species-disturbance relationships in riparian zones.