15
Ecology
Ecology is a theoretical, quantitative study of organisms, populations, species, communities, and ecosystems. Both locally and globally, this highly integrative discipline incorporates information, processes, techniques, and data for related fields represented in the previous chapters, as well as inorganic chemistry, organic chemistry, physics, meteorology, geology, soil science, evolution, genetics, immunology, pathology, bioengineering, behavior, and natural history. While all disciplines are interdisciplinary to some extent, few are as interdisciplinary as ecology.
The study of the interactions of organisms, with each other and with the environment, is far more complex than we can fully comprehend. However, that doesn't stop us from trying. Ecology began as a pure science, primarily the study of species in their natural habitats, and expanded into a science involved with ecological principles and mechanisms. Much of the theoretical aspect of this field has yet to be used in any practical way. Ecological studies provide a valuable base for all those who plan to enter a field that will potentially affect the environment. It would be much better if far more people studied ecology at some time in their life. Unfortunately, too many children grow up thinking ecology is all about recycling, period. Ecology is far more than just that, and for legislators to ensure that we don't destroy much that lives on our planet, it's important for them to learn that healthy people and healthy communities need far more than a robust economy. We also need to make sure that everyone has access to a healthy, beautiful, vibrant, natural environment. And for species to survive, we cannot just save a small number of parks where everyone goes on vacation. We need places where plants and animals, and fungi and bacteria can live on in perpetuity, without becoming threatened or endangered. I know, that's asking a lot of people.
Having laws to save species when they become endangered sounds like a good plan, but it doesn't work. By the time a species becomes endangered, the costs involved to fix things properly are more than most societies choose to bear. As a result, endangered species management should be a last resort. It's far wiser and costs much less to act early rather than late, by setting aside large swaths of land for long-term conservation purposes. Large wildlife preservation parks should be created that are off limits for recreation. Having large parks that include recreation too is also good. Unfortunately, mixed-use parks put too great a burden on too many habitats and on too many species to ensure their survival. So it's best to create conservation parks and then leave the conservation parks wild, without logging, skiing, hiking, fishing, biking, boating, and everything else people love to do. Leave wild places alone so we don't introduce aggressive invasive species, and so we don't take the turtles, snakes, and rare flowers out (people steal things all the time), and so we don't bring seeds into wild places in the treads of the tires of the vehicles we drive. Just leave the wild places alone (maybe some open places should be maintained by cutting woody vegetation every several years, and maybe some places should be burned over occasionally). Nature knows how to manage a place far better than any landscaper, environmental protection specialist, ecologist, or wildlife biologist I've ever met.
The field of ecology grew in response to our immediate need for a greater understanding about the interactions of organisms in their natural, as well as in what are increasingly, highly altered habitats. With human disturbances affecting ecosystems globally, we are creating methods to assess and remediate the effects of our actions. We also need to maintain healthy ecosystems, which can be done most easily by enabling nature to do what it does for free, when we provide enough undisturbed real estate, not just on land, but also in fresh, brackish, and salt water. In rivers, bays, on continental shelves, and in just about every habitat imaginable.
This chapter is an overview of the primary ecological concepts that are presented in most introductory biology courses, as well as in the media.
HABITAT AND NICHE
The physical environment affects organisms living there in many different ways. The climate, weather, and microclimates are influenced by temperature, humidity, rainfall, and other meteorological phenomena. Soils are extremely important in their effect on terrestrial habits. In addition, aquatic habitats are also affected by salinity, oxygen, and carbon dioxide in solution, pH, temperature, as well as by available minerals and by other nutrients. The amount of light penetrating through any environment is important, as are the seasonal fluctuations with regard to any of the above variables.
Just as aquatic organisms are adapted to specific sets of variables, certain plants grow only on special soil types, and some animals live only in areas where there are certain associated vegetation patterns. Air quality is also important.
An organism's habitat is the place where it lives. The variables mentioned above determine the characteristics of that habitat. An organism's niche is what the organism does. A niche is described by defining an organism in terms of its role in the ecosystem. Niches may be described using as many variables as one examines. For instance, a niche might be represented in terms of where an animal forages, when it forages, and what it forages on. This could be contrasted to the niches of related species living in the same general area to see if their niches vary according to these variables. The range of variables one examines is practically limitless.
POPULATIONS AND COMMUNITIES
All the members of a certain species living in a specific area at a specific time are defined as a population. Each of the populations living in a given area constitutes a community. Changes in a population's size (number of individuals in the population) are due to births, deaths, immigration, or emigration. Should births and immigrations exceed deaths and emigration, the population will grow. Likewise, the opposite is also true: If the total number of deaths and individuals emigrating from the population exceed the gain from births and immigrating individuals, the population will decrease in size.
The density of a population is calculated by counting the number of individuals per particular area (or volume). Dispersion is a parameter used for understanding the type of spacing of individuals within the population. The individuals may be distributed uniformly, randomly, or in clumps. The last two distribution patterns are more common than the first.
POPULATION GROWTH
Many factors affect the numbers of organisms in each population. Such numbers may vary from season to season or from year to year. Some populations are growing in numbers while others are declining. Many remain quite stable, sometimes having built-in fluctuations. When there are no checks controlling population growth, it may grow on a logarithmic scale. When plotted on a graph, such growth curves are often termed exponential or geometric (see Figure 15.1). Populations that grow in a more controlled manner usually have a growth curves that are linear. Such growth patterns are usually called arithmetic.
The rate of change in numbers of individuals in a population during a period of time is related to that species' innate capacity for increase or biotic potential. The actual rate of population increase at any given time may be calculated and is presented as a number that is called r. The biotic potential is higher for fruit flies than it is for people; therefore flies have a higher r.
A species' biotic potential is affected by any of the following:
- The number of offspring produced each time a member of the species reproduces.
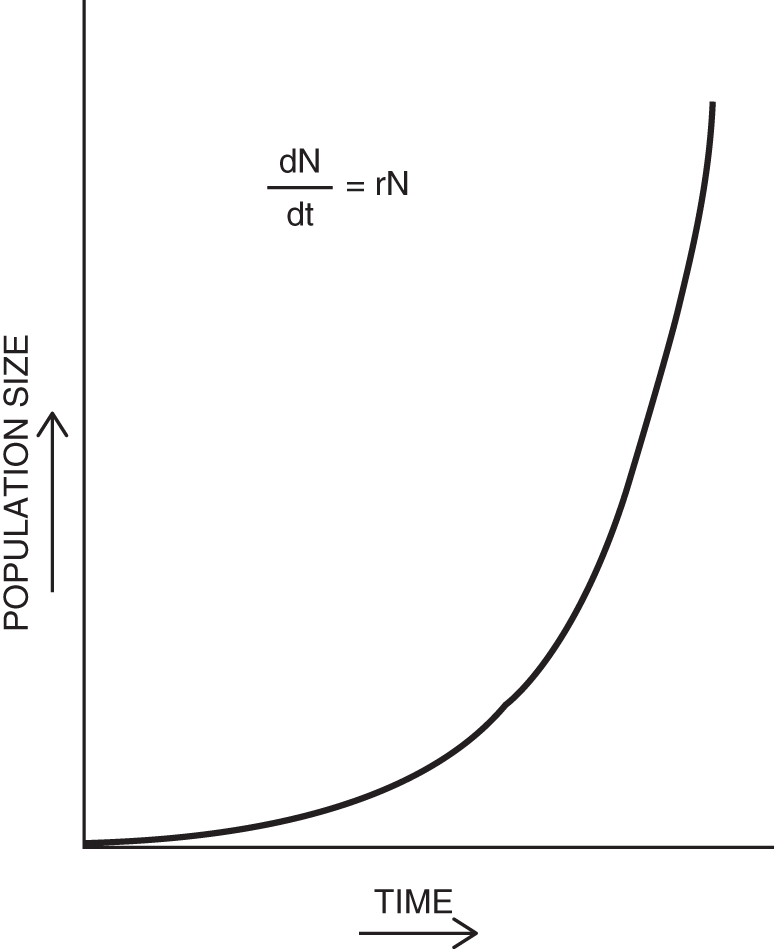
Figure 15.1 Exponential growth curve (N = population size; r = rate of increase; t = time; d = change).
- When the reproductive life of that organism begins (how old is the organism when it can begin to reproduce?).
- How long the reproductive life of that organism lasts.
In addition to representing survivorship with survivorship curves, life tables may also be constructed. These summarize, for each class as defined by the researcher, specific types of information, such as the number of organisms living at the beginning of that age interval, the number of organisms that die during that age integral, and the average number of years of life remaining at the beginning of that age interval. Researchers often use this information to project what the effect on a population might be if certain variables are changed very slightly. Life table data have made it possible to project a range of effects without having to conduct nearly as much time-consuming fieldwork. Some computer programs have been especially useful in interpreting reams of data in new ways that were far too difficult just a few years ago.
SPECIES DIVERSITY
Ecological research has taught us about patterns in population size of individual species, as well as about systems involving the interactions of two or three species. But systems quickly become amazingly complex when more species are factored into the equation. There is still much to be learned about species interactions, but most of this information will be of theoretical interest, instead of being useful for conservationists. From my experience, organizations with conservation in their name, or as their mission, do very little innovative or useful conservation work beyond what was done each year previously. I wish this was not the case, but it is. Conservation organizations like to make it look like they do great work, but much of what they do is run an organization that has programming that looks good, might even do good, and of course, the organization has to raise money.
Ecological homeostasis, defined as well-regulated species population sizes combined with species diversity, is affected by many components, only some of which are understood. Species diversity can be defined as the number and kinds of species present in any specific community. In addition to species diversity, the population size of each species is very significant in understanding the dynamics of that community. Some species are dominant. In fact, in most communities, there are only a few species with significantly greater abundances than all the other species. From this comes the often repeated phrase referring to numbers of species and their relative abundances: “Few common, many rare.”
The factors contributing to what makes a species common in a particular community and what defines a “rare” species are continually changing. One example of what helps to explain a species' relative abundance has to do with whether it is a specialist or a generalist. What is meant by these terms is that some species specialize at something. For instance, a particular lizard may be observed feeing on insects during a particular time of day, when the temperature is within a well-defined range, at a certain height in the trees, on branches that are a certain width in diameter, eating only those insects that are within a narrow size range. There might be nine other closely related lizard species in that tree. However, each specializes in feeding at a different height, or on a different width branch, or at a different time, or on a different food. Repeatedly we find situations where species, usually closely related, are “packed” together, but each species' niche overlap is minimal. This appears to be due to genetic control. Each of these species could be said to be a specialist. And then there are generalists, those species that might be moving all over the tree, as well as on the ground, throughout the day (or night, or both), eating a range of foods.
SPECIES TURNOVER, RECRUITMENT, AND INTRODUCED SPECIES
Another factor relates to species turnover. The species' composition within a community is not always stable. In fact, of those species communities studied over time, it is common to find some species going extinct locally, a process usually referred to as extirpation, and, concurrently, new species moving in (recruitment). The rate of species turnover in a particular community can help tell an investigator much about the local interspecific dynamics (dynamics between species, as opposed to intraspecific interactions, those within a species).
Many species are being introduced to areas where they are not native. Such exotic species, often referred to as aliens, may compete with native species for otherwise scarce resources. The net result of the increasing numbers of introduced species has been a growing list of native species that are decreasing in number and range. Some may eventually become extinct.
COMPETITION
When two members of one species or of different species exploit a common resource, the interaction is often termed competition, though the behavior is rarely observed. Designing experiments that enable a researcher to show whether or not competition exists and, if so, to what extent usually requires great care and a rather simple system.
Competition may occur between different species (interspecific competition) or within a single species (intraspecific completion). When each individual directly competes for a resource without regard to other individuals that may also be using that resource, it is known as scramble competition. In cases where individuals compete indirectly for what may not be a very tangible resource, such as for social dominance or for territory, it is often called contest competition. Here, successful individuals usually get enough of the limited resource while the losers often go without.
The competitive exclusion principle states that when two or more species compete for the same resource necessary for their livelihood, and they are found or placed together in a situation where the resource is limited, they cannot coexist indefinitely. One will win out in the long run. Put another way, two species cannot occupy the same niche.
Sometimes the niche that a species might occupy differs according to the situation. That is, when only one species is there, the fundamental niche (what the species does when there is no competition) may be larger than the realized niche, the niche that is occupied when the environmental conditions are more restricted because one or more species are infringing on that otherwise larger, more expansive niche.
DENSITY
What causes the increased mortality of a species is not always clear-cut. As the population density of one species increases, the mortality within that species may increase. In such a case the mortality is said to be density dependent. However, if the mortality is due to causes other than the species' density, then it is said to be density independent.
CARRYING CAPACITY
The carrying capacity, often referred to by the letter K, is the number of individuals of a particular species that a specific environment can support indefinitely. Change the environment, and K changes. Add a parasite, remove a competitor, add a predator – each of these factors may affect the carrying capacity of the species in question. The curve often used to show the exponential increase in number of a species until it reaches its carrying capacity in that environment is called a logistic curve (see Figure 15.2).
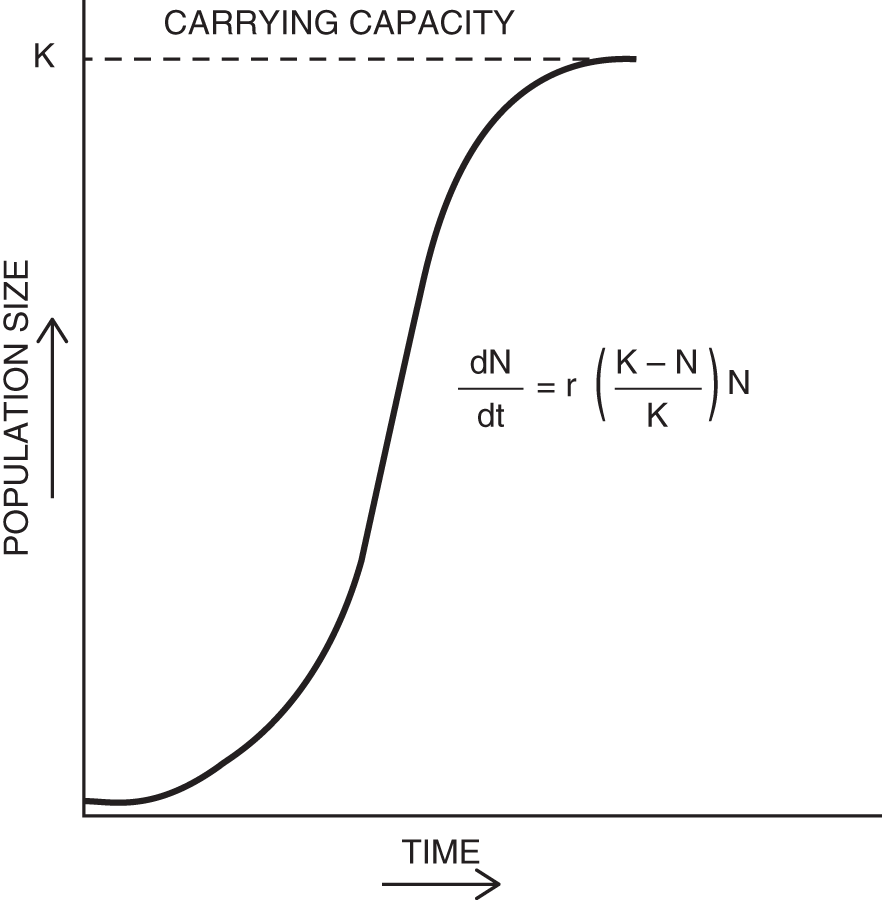
Figure 15.2 Population growth as described by the logistic equation (K = carrying capacity; other variables as defined in Figure 15.1).
SUCCESSION
The population size of each species and the species diversity in many communities are stable and persistent, often remaining predictably uniform for centuries or more. Other populations are far more fluid, or dynamic, entities in the constant process of change from what may appear to be one distinct community to another. Such a sequence of communities replacing one another is known as succession, which is usually far more gradual than is commonly recognized. Successional events that occur on land are known as terrestrial succession; successional events that occur underwater are known as aquatic succession.
Succession may be triggered by a cataclysmic event such as a flood, fire, or even a plow or bulldozer. But sometimes, the changes are triggered by a far subtler influence, such as a disease, a slight increase in average summer temperatures, or the invasion of a new insect.
The exact species composition is the result of such factors as soil, climate, which seeds arrive first, and which seed germinate first. When a cataclysm is initiated by a plow, for instance, seeds germinate on the cleared soil. Some seeds are blown in, others deposited in animal feces, so which animals live nearby or migrate through can be as important as any other single factor. The initial colonizers also impose certain characteristics: They act as a filter, enabling only certain new species to prosper.
The field may be composed of grasses and herbs, annuals, biennials, and perennials. The precise mix changes with time, and eventually woody species take hold and shade out many of the field species. In addition to the woody shrubs, pines and cedars are good colonizers that grow quickly when out in the open. However, if fires don't burn them back, perhaps every 15 years or so, these evergreens may soon be overtaken by deciduous hardwoods, species such as oaks and maples. The reason pines may not be overtaken by hardwoods if fires persist with regularity is that the pines are often fire resistant. Although they suffer considerable damage, they also grow back quickly, and in a number of years the area will once again be a pine forest. But without the fires, the hardwoods overtake the softwoods (pines) because they grow higher, shading out and eventually causing the pines to die off.
In this continuum of different communities that gradually change into one another, the concept of the final stable community is often evoked. That is, in some places, an oak-hickory forest might be the last natural successional community. Elsewhere it might be a beech-maple forest, depending on the weather, soils, and a number of other factors. Other final, stable communities, also known as climax communities, are deserts and grasslands. Climax communities occur only after the gradual series of continual changes. Preceding communities are lost to better-adapted competitors that thrive under these different conditions, made different by the new plants and animals living there.
Once in place, a climax community lasts until the next flood, fire, or bulldozer comes along, when the successional processes are reinitiated.
REPRODUCTIVE STRATEGY
Natural selection favors organisms that tend to pass on more offspring, which in turn pass on more offspring, and so contribute to succeeding generations. Included among the characteristics that natural selection appears to affect are those that shape an organism's reproductive strategy. Such strategies are not conscious. Rather, they evolve over time. There are specific categories of reproductive strategies, with many variations. For instance, some organisms produce great numbers of minute offspring, while other species invest more in each offspring; they have only a few, but each is large, with considerable survival value. One of the primary differences among most reproductive strategies is the survivorship of the young. If each parent can have a few young but still pass on enough offspring to maintain the population, that might be sufficient. However, if there is extremely high mortality, and only a very small percentage of the young survive to maturity, it is sometimes best to bear very many young, investing little in each. Each species has its characteristic reproductive strategy.
Recording and plotting the fate of the young and their chances of survival at key age categories enable the researcher to construct survivorship curves. Birth rates, death rates, and longevity (how long the organisms may live) are significant factors affecting all populations. Recording the numbers of births and deaths over a period of time and determining the average longevity of the organisms during each age class tell the scientist much about that population. Should the number of births rise, the number of deaths fall, or the survival time of the organisms increase, the population will be affected positively; that is, the population should grow. Conversely, should the births decrease, the death rate increase, or the average longevity decrease, the population would be affected adversely.
Average longevity varies from species to species. When the survivorship curves are plotted, much can be determined about the reproductive strategy of that species. The three basic curve shapes are presented in Figure 15.3, although there are many variations.
A Type I survivorship curve (e.g. humans in Figure 15.3) depicts a species that has offspring with higher survival rates; most live to a certain age and then die. A Type II survivorship curve (e.g. deer in Figure 15.3) depicts organisms with a steady death rate stretching from the time they are born or hatch until the time they die. Their survivorship curve usually varies along a straight line. And a Type III survivorship curve (e.g. maples) is the inverse of a Type I curve; it depicts a low survivorship shortly after being born, but with high longevity for the individuals that survive.
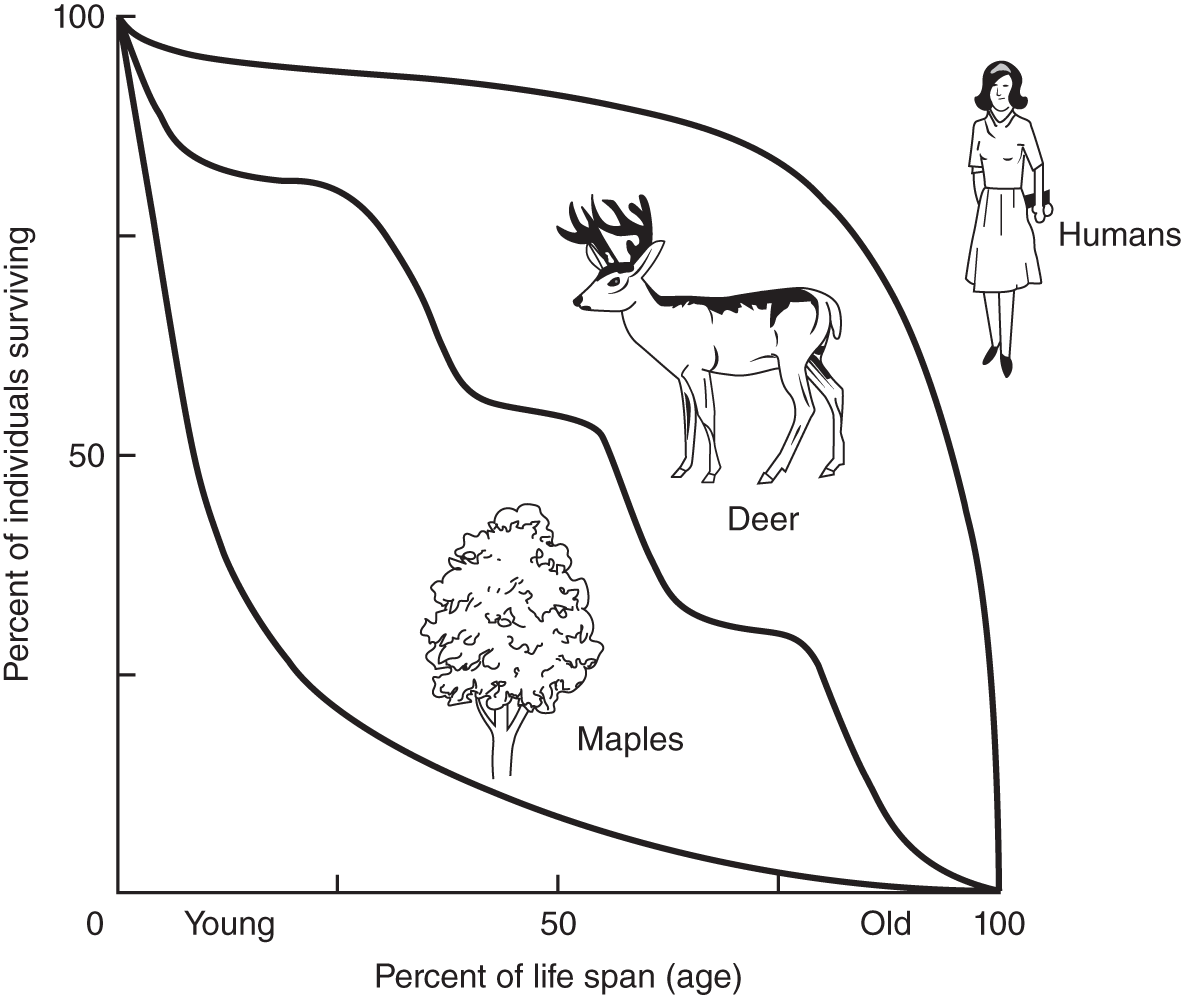
Figure 15.3 Survivorship curves for humans, deer, and maples.
ECOSYSTEMS
Ecosystems are the sum total of the organisms and their environment in a given area; all require energy. The sun is the ultimate energy source in most ecosystems. During the process we call photosynthesis, plants use light energy to convert carbon dioxide and water into carbohydrates. Plants also use additional inorganic nutrients and ions to manufacture nucleic acids, proteins, photosynthetic pigments, and other necessary components.
Organisms such as plants, which manufacture their own organic food molecules from inorganic substances, are the producers and are often called autotrophs. Organisms that acquire food from dead plants and animals are called decomposers. In some ecosystems, the metabolic pathways of certain bacteria use chemicals rather than sunlight to synthesize organic materials. Such alternative methods of manufacturing organic food molecules are termed chemosynthesis.
Ecosystems require an energy source, producers, decomposers, and abiotic (nonliving) nutrients. These are just the basics; most ecosystems have more components. When there are organisms present that eat the plants, such animals are called herbivores, or primary consumers. When these die, like all dead material, decomposers break them down. However, when living herbivores are eaten, their predators are termed carnivores (animals that eat other animals or animal parts); they are the secondary consumers. Depending on the complexity of the particular ecosystem, sometimes there are tertiary, or even quaternary consumers.
The autotrophs, or plants, represent the first level in the series of organisms that together are sometimes thought of as creating a food chain. Because of all the connections and complexities within most food chains, they are also called food webs. The plants are the first and most basic form of energy storage in an ecosystem. They are called the first trophic level. The second contains those animals that eat plants, and the third, when present, consist of animals that eat herbivores. Organisms that eat both plants and animals belong to more than one trophic level and are called omnivores.
When the energy is passed from one trophic level to another through the consumers, a certain percentage of energy is lost through the processes involved in hunting, eating, digesting, reproducing, growing, and maintaining. There is less energy available in each succeeding trophic level, which explains why few ecosystems contain more than five trophic levels. Beyond five trophic levels, rarely is there enough energy to support much else.
Sometimes the fewer numbers of organisms in each succeeding trophic level are illustrated as a pyramid, with the earlier trophic levels at the base, building up to the narrow peak. Not only is a pyramid of numbers helpful, but also a pyramid of biomass is effective when demonstrating the total dry weight of living material in each succeeding trophic level. Depending on what is occurring in any particular ecosystem, occasionally the pyramid is inverted, with the primary producers representing either fewer numbers or less biomass and the larger base on top. Such situations are usually short-lived.
The energy accumulated by the plants in a given ecosystem is known as the primary production. The rate at which energy is stored as organic matter due to photosynthesis is called primary productivity. This stored energy is usually expressed as kcal/m2/year, or as biomass gained per unit area over a given unit of time. Since some of the energy is immediately metabolized to maintain the plant's respiratory activities, it is sometimes helpful to calculate the gross primary productivity, which is the total rate of photosynthesis. Therefore, the gross productivity minus the respiration rate (energy used for respiration) leaves the net productivity (see Figure 15.4). It should be remembered that productivities are rates, yields are weights (or expressed as volume), and the standing crop is the total amount of biomass per unit area at any given time.

Figure 15.4 Annual net primary productivity of natural vegetation.
Some people warn of vast collapse of ecosystems (ecosystem collapse). In the past, factors that caused something on the scale of what people are now warning about include large asteroids that collided with the earth, vast volcanic eruptions, and sudden changes in climate. Ecosystem collapses such as these in the past resulted in mass extinctions. A mass extinction is when there is a widespread and rapid decrease in biodiversity (variety of life) on earth. Today, humans are radically altering many ecosystems, while many other ecosystems are being affected, but nature is resilient and adaptable. What we are doing to the earth is important, but it is not the same as an asteroid or several supervolcanos erupting.
BIOMES
The major ecosystems, often called biomes, are regarded as distinct entities with distinct forms of life. These regional ecosystems generally grate into one another, forming a gradient or transitional zone. When the transition from one ecosystem to another is abrupt, leaving a significantly different environment along the barrier between the two, such as the beach or coastal zone that lies between the ocean and land, such a zone is sometimes called an ecotone. The major biomes of the world are depicted in Figure 15.5 in terms of their mean annul temperature and precipitation. Each of these ecosystems is described below.
Tropical Rain Forest
Characterized by an annual rainfall of more than 400 cm (over 150 in.) and warm temperatures during most of the year rather than just during a specific warm or rainy season, tropical rain forests are wet and green all year. The soils tend to be nutrient-poor because the warmth and moisture have oxidized most of the available nutrients from the soil, which are then leached out by all the rain. Because the conditions are warm and moist, decomposers consume the organic material practically as soon as it becomes available. There is usually only a very thin layer of rich organic material, on top of the forest floor, and very little topsoil.

Figure 15.5 Biomes are major ecosystems with distinctive forms of life. The biomes graphically represented here are presented in terms of temperature and precipitation.
In terms of the overall number of species, or species diversity, the tropical rain forest is unsurpassed. These forests are most extensive in the northern part of the Southern Hemisphere, where they are being logged rapidly. In many cases it is local governments and international corporations that favor logging these forests for the short-term profits despite the long-term losses, should most of these forests disappear. In addition to the extinction of hundreds of thousands of species, it has been predicted that the loss of these forests will contribute to what many call either global warming or climate change. Global warming is usually defined as a long-term increase in the average temperature of the earth's climate system. Climate change refers to changes in the earth's weather patterns that last for many years (some changes in the earth's weather patterns last for decades, others last for hundreds, or thousands, or even millions of years). The data collected that measure the climate are tested statistically to determine if such long-term changes are statistically significant.
Tropical Seasonal Forest
Unlike the tropic rain forests, which are largely evergreen since most of the trees retain leaves year-round, tropical seasonal forests are composed of more deciduous tree species, due to more prolonged dry seasons. The dry seasons are similar to winter in colder climates, except for the lack of cold. This is because during the dry periods, and during the cold periods, many species become relatively dormant, and the majority of tree species lose their leaves. When the rains return, the plants leaf out and the organisms become more active.
Tropical Savanna
Typical and extensive in Africa, tropical savannas occur where the rainfall is capable of supporting vast expanses of grass with some trees and shrubs, but never any forests. Because of limited rain and the prolonged dry season, woody plants do very poorly. The wet season supports grasses that rapidly dry out and remain dry the rest of the year. Some hardwoods persist in the lower areas with whatever runoff there is.
Tropical Thornwood
Where the rainy season is sufficiently wet to support some semidesert scrub, but conditions are too dry to support a forest, sometimes there are zones of tropical thornwood.
Desert
Because most deserts receive less than 20 cm of rain a year, their vegetation is sparse, and some deserts have little to no vegetation. Other deserts have many specialized plants that store water in their thick, fleshy tissues. Such plants are succulents.
Temperate Forest
Unlike the tropical areas, temperate climates have more moderate temperatures. Here, forests grow that are classified either as temperate forests or as temperate deciduous forests, having warm summers and cold winters. Like tropical seasonal forests, as well as other seasonally cold or arid ecosystems, most trees in this biome lose their leaves as the weather grows colder and grow new ones as the weather warms up. Soils are rich in organic matter. Despite richer soils, temperate forest trees rarely reach the heights typical of those in the tropical rain forest.
Temperate Evergreen Forest
Temperate evergreen forests grow on poor soils in areas where drought and fire frequently occur. The species that dominate these forests are usually needle-leaved conifers, such as pine, fir, and spruce, as well as some broad-leaved evergreens.
Temperate Rain Forest
Temperate rain forests prevail in cool areas near the ocean where there is considerable rainfall during the winter and much fog during the summer, such as the rain forests of the Pacific Northwest.
Temperate Woodland
In the American southwest, between the grassland and semidesert biomes at the lower elevations and the pine forests at higher elevations, are temperate woodlands that are too dry to support a forest but too moist for mere grassland. Instead, they support a range of communities from rather dense woodlands of small trees to more open woodlands with taller but more isolated trees.
Urban Ecosystems
Urban and suburban ecosystems are the newest, fastest-growing distinct ecological entities. And yet, until recently, they were the most overlooked habitats despite their far-reaching effects. New to much of the world, few urban environments have been in existence for more than 5,000 years. During the interim, vast areas have been transformed. Urban ecosystems are responsible for subsuming a range of other habitats into what now constitutes part of the meshwork of urban communities, including agricultural areas, which affect nearby rivers, estuaries, and even oceans. Because of gases released due to certain activities and machines that are controlled by humans, which are the dominant organisms inhabiting these ecosystems, the atmosphere is being altered in minor ways that may lead to long-term consequences.
As the world human population grows, we continue to destroy healthy, “natural” habitats, replacing them with urban habitats, which in some respects are also “natural.” In these habitats we find many species do quite well, not in spite of us, but because of us. We have created an environment where far more than rats, roaches, and pigeons excel. Often, we find such species, which we now associate with urban ecosystems, doing better than they did before we created cities, towns, and suburbs. The world is currently undergoing one of the major ecological changes of all geologic time, and the primary ecosystems to benefit, often at the expense of other ecosystems, are urban ecosystems. In the process, world species diversity is expected to decline, perhaps precipitously. With innovative wild nature friendly urban planning, and with targeted introductions, it is possible to create and improve green spaces that become important for the survival of many species.
Temperate Shrubland
The temperate shrubland communities are often called chaparral. These relatively dry areas are found near water, such as along parts of coastal California or the Mediterranean. These areas get little to no rain during the summer and are subject to frequent burning. After the fires, the shrubs grow back, regenerating from the roots and tissues left near the ground.
Temperate Grassland
Depending on the country, the temperate grasslands are known either as prairie, steppe, pampas, or veldt. Located on vast lands in the interiors of continents where there is enough moisture to support shrubs and grassland, but not enough rain for a forest, temperate grasslands usually develop rich deep soils over many hundreds of thousands of years, which are now used in many parts of the world to support productive agriculture. Much of America's Midwest and the Ukraine were temperate grasslands that are now farmed intensively.
Temperate Desert
True deserts are not very common in temperate regions. Such deserts are usually too dry for much life to exist. Rather, most temperate deserts are classified as semideserts, or as cool temperature semideserts. Too dry to support grasslands, they are often dominated by shrubs, such as sagebrush, with many perennial grasses throughout.
Boreal Forest
The boreal forest, often called the taiga, stretches through much of Canada and Siberia, where it's cold and windy and the soils tend to be rocky and poorly developed. To survive here, a species must be able to tolerate the harsh winters. Spruce trees, pines, and firs dominate this biome, because these trees do well under these conditions that most other species have difficulty surviving.
Tundra
Tundra conditions exist farther north than the boreal forest, as well as at high elevations, where it is too cold for most trees to grow. Vegetation above the treeline primarily consists of sedges, grasses, mosses, lichens, herbaceous flowering plants, and dwarf woody shrubs.
Alpine tundra, unlike the true tundra of the north, is similar. Except that these communities are found on mountaintops between the timberline and the highest areas, where little survives. Both environments share several characteristics, including shallow soils and a brief growing season. In some tundra regions the soils remain frozen all year round, a phenomenon called permafrost. Where this occurs, sometimes the surface thaws, but just below the surface, temperatures never rise above freezing. Tundra plants grow at a comparatively slow rate. In addition, because of the cold weather, dead material decomposes at a rate that is many times slower than in warmer environments.
Streams and Rivers
These systems typically involve both fresh water and a moving current. But beyond that, streams and rivers vary considerably. They range all the way from a roaring torrent pounding down a snowcapped mountainside, to the trickle of a dried-up summer brook, or an immense, meandering river such as the Amazon, working its way through the vast region of tropical rain forest in South America.
Ponds and Lakes
As with streams and rivers, most ponds and lakes have fresh water in common (though some are saline). They are associated with a wide range of typical species and associated habitats, and they occur within a wide range of temperatures and nutrients. There are different categories of ponds and lakes, but most limnologists (specialists who study freshwater environments such as lakes and ponds) find it helpful to group these freshwater bodies together. The experts have a vocabulary that enables them to describe particular freshwater habitats with precision.
Freshwater Wetlands
Freshwater wetlands also vary considerably. The domain of most wetland biologists ranges from open water to wet meadows (which may be wet only during the rainy season). Specific vegetational patterns are associated with each type of wetland.
MARINE COMMUNITIES
Although the major biomes presented in most texts are usually restricted to terrestrial communities, there are many marine communities, those aquatic communities that contain more salt than occurs in fresh water. Like many of the communities presented above, marine communities also tend to merge into one another, but it is possible to identify distinct zones. Some of these are presented below.
Estuaries
The region where fresh water from streams and rivers drains into salt water is called an estuary. These coastal areas vary in salinity depending on the geography and the tides. The constant mixing and influx of nutrients, combined with the many protected habitats that typify these estuarine environments, make them one of the most productive ecosystems worldwide.
Estuaries often have tidal marshes. These are coastal areas with typical vegetational patterns. The dominant plant of salt marshes is Spartina, which is a genus, or group, of closely related grasses. Spartina, as well as many aquatic species inhabiting estuaries, can tolerate a range of saline conditions from several times the salinity of the ocean, which is 34 parts per thousand, to fresh, which has no salt at all. Pools on top of the salt marsh, due to evaporation, can become several times more saline than the open ocean. It should come as no surprise that of those organisms that inhabit the estuarine environment, most are well adapted.
These rich, protected environments are the breeding grounds for a wide range of invertebrates and fish, as well as species such as colonial water birds that feed on both of these. Many migrating birds and fish arrive in schools, sometimes just to feed. Others remain long enough to mate and lay eggs, and birds often spend an entire summer.
Littoral Zone
The intervening area between where the water reaches during high and low tide is known as the littoral zone. Many of the organisms inhabiting this zone can tolerate major swings in temperature and salinity, as well as the periods of desiccation when left out of water during low tide. Depending on the local conditions, littoral zones may be either rocky, sandy, or muddy. Each supports very different communities.
Coral Reefs
Coral reefs, along with estuaries and tropical rain forests, are among the most productive ecosystems in the world. Coral reefs occur in warm, shallow marine waters around the world. Usually confined to the tropics, coral reefs share several characteristics. They are all built by coral polyps (phylum Cnidaria) that secrete the hard, limy, calcareous material that becomes the infrastructure for all the other associated organisms essential to a healthy reef. Reefs may be exposed at low tide, but the majority of the coral lies under water. Coral reefs grow in clear, oxygenated waters where waves break. Coral species grow in water less than 100 m (328 ft) deep because they can only exist where sufficient light reaches the associate, photosynthesizing organisms, such as the symbiotic species of algae.
Open Ocean
Of the other marine communities, the largest is found in the open ocean. Past the sublittoral zone that occupies the continental shelves (the edges of the continents where the underwater terrain gradually drops off), the continental masses drop off rapidly and, in some places, the ocean becomes very deep. In the clearest water, light can penetrate the first 100 m (328 ft). Beyond that, little to no photosynthesis is possible. Organisms, however, continue to live where there is no light, even as far as the deepest marine canyons that reach several kilometers in depth. The deepest recorded part of the Pacific Ocean is over 7 miles deep (36,200 ft deep). The small creatures that make up the plankton are those drifting about in the water. Such species include algae and other organisms that are so small they are supported by the water's buoyancy. The larger animals, such as mature fish, make up the nekton. Such species usually have more directed, controlled movements.
MINERAL NUTRIENT CYCLES
Within each of the ecosystems described above are specific biologically active minerals that cycle through the plants and animals in that environment. These cycles are known as biogeochemical cycles. Unlike solar energy, the elements that function as nutrients in ecosystems can be recycled and are used over and over again in living organisms. The major biological nutrients are carbon, hydrogen, oxygen, nitrogen, and phosphorus. Other elements also prove important in lesser amounts; these include boron, calcium, chlorine, cobalt, iodine, iron, magnesium, manganese, potassium, and sodium.
Many biologically active elements emanate from rocks as the rocks and minerals erode, or from the soils they help form. They are taken up through roots or are absorbed from water or through the air. The minerals may be incorporated into an organism's cellular structure, where they are used and then released back into the system, or they may be incorporated into the organism's tissues, to be released upon death and decomposition. Either way, the minerals eventually become available again where they can be absorbed and used by other organisms.
Carbon Cycle
The carbon cycle passes carbon from the atmosphere to organisms and back through respiration and photosynthetic processes. Most carbon is released into the atmosphere in the form of carbon dioxide (CO2). In water, carbon may exist either as dissolved carbon dioxide or as bicarbonate (HCO3−). Gradually, over hundreds of millions of years, considerable amounts of carbon have accumulated in some systems in the form of peat, coal, oil, or natural gas. Such carbon reservoirs can remain locked up for millions of years before rejoining the carbon cycling system again.
Since the beginning of the industrial revolution, the burning of fossil fuels has released much of this previously locked-up carbon into the atmosphere in molecules of carbon dioxide. Many believe all this carbon dioxide has caused or contributed to a greenhouse effect, that they say is warming up the atmosphere, because atmospheric carbon dioxide prevents some of the earth's heat from radiating back into space. Although the carbon dioxide concentration in the atmosphere has increased significantly over the years that humans have been burning forests, wood, coal, oil, and natural gas, the average global temperatures have not increased nearly as much as mathematical models predicted. The concentration of carbon dioxide in the earth's atmosphere is now 415 parts per million (ppm). The last time the earth's atmosphere had this much carbon dioxide in it was three million years ago. 500 million years ago the earth's atmosphere had 7,000 parts per million of carbon dioxide.
Many scientists say an average rise of a few degrees temperature globally can alter weather patterns and melt portions of the polar icecaps, which might lead to a rise in sea level, submerging today's beaches, salt marshes, deltas, and urban centers situated at sea level. In addition, the damage caused from hurricanes, which regularly hit coastal cities and towns, could affect millions of people. The U.S. government has already begun to buy out people who live in such areas, enabling them to move to higher ground.
Even without human interference, the earth's climate continually changes. For instance, the earth is overdue for our next ice age, due to cyclical periods of global cooling, that lead to mile deep glaciers over many parts of earth that are currently densely populated. As snow accumulates, water that evaporates from the oceans gets tied up in ice, which causes the sea level to drop (or lower), which exposes vast areas that are currently underwater. The increase in the atmosphere of carbon dioxide and other greenhouse gases that is being caused by humans may buy us time before the next global cooling.
Nitrogen Cycle
Amino acid and protein molecules contain nitrogen. However, even although there is far more nitrogen than carbon dioxide in the atmosphere (carbon dioxide = 0.03%; molecular nitrogen = 78%), most of this nitrogen (N2) is in a form that can be used by only a very small group of specialized organisms.
Nitrogen gas (N2) does not react with water, though it does dissolve in water. Nitrates and ammonia easily dissolve in water. Some bacteria and blue-green bacteria (cyanobacteria) can convert, or fix, gaseous (molecular) nitrogen into an aqueous form, ammonia (NH3), by reducing it. This involves adding electrons, a process that is usually accomplished by combining the nitrogen with hydrogen, which carries an electron with it. It is primarily from these nitrogen-fixing microorganisms that most gaseous nitrogen is converted to a form capable of being used by other organisms. However, some nitrogen exists as ammonium ions (NH4+) or nitrate ions (NO3−) which, having been eroded from rocks and leased from soils, are dissolved in the waters around the world.
Certain bacteria and fungi use the nitrogenous materials found as proteins and amino acids in dead organisms for their metabolic purposes. They then release ammonia and ammonium ions back into the soil. Other bacteria in the soil that are known as nitrifying bacteria oxidize ammonia to nitrate ions (NO2−) by removing electrons (oxidation), usually by removing hydrogens, which carry the electrons with them.
Lightning also produces nitrate. All these sources of fixing nitrogen are necessary to the rest of the organisms found around the globe, because without fixed nitrogen there would be far less plant life and without plants little else could survive.
Because fixed nitrogen is so important to plants, it is a key ingredient in many fertilizers. Since it is expensive, farmers often choose to rotate their crops rather than fertilize, occasionally growing alfalfa or any of several other members of the pea family (Fabaceae, previously called Leguminosae, commonly called legumes), all of which have symbiotic nitrogen-fixing bacteria living in nodules attached to the roots. By growing such a crop, not only is the alfalfa harvested for horse and cattle feed, but fixed nitrogen is manufactured by the symbiotic bacteria and released into the soil, free of charge. The symbiotic nitro-fixing bacteria live in close proximity to legumes (plants in the pea family) so that they may receive nutrients from the plants. In exchange, the host plants benefit from the fixed nitrogen fertilizer. Denitrifying bacteria live under anaerobic conditions (no oxygen), such as exist deep in the soil or in mud. They complete the nitrogen cycle by returning nitrogen back to the air.
Phosphorus Cycle
Phosphorus usually occurs in molecular form as a solid, or as a component of a liquid, though it almost never occurs as a gas. It is indispensable to life since it is an important component of adenosine diphosphate (ADP), atom transfer radical polymerization (ATRP), ribonucleic acid (RNA), and deoxyribonucleic acid (DNA). It is also found in cell membranes, as well as in shell, bone, and teeth. The phosphorus cycle is termed a sedimentary cycle because unlike the previous two cycles, carbon and nitrogen, which are termed atmospheric cycles (because carbon and nitrogen occur as gases during part of their cycles), phosphorus is a sedimentary cycle because its cycling involves the erosion, weathering, and the breakdown of rocks, minerals, and soils in ways that release minerals containing phosphorus. Through erosion, phosphate (PO43−) is formed. Plants then absorb this phosphate. When animals eat the plants, or when animals eat other animals that eat plants, the phosphorus is in a form that is usable to them. Any excess phosphorus excreted by the animals is in the form of phosphate, so it may be reabsorbed by the plants.
Although much of the phosphorus stays in terrestrial ecosystems, a certain amount that originates from rocks and is absorbed by plants, which were eaten by animals, eventually gets washed downstream and out into the ocean. Eventually, it settles to the ocean floor, where it remains for what may be millions of years before being exposed and eroded out, becoming part of the cycle again.
Much phosphorus used to be mined in the form of bat and bird droppings, the guano which accumulates in bat caves or on some oceanic islands where thousands of sea birds have nested for generations. Other sources exist, such as from bones, certain minerals, and from local deposits in Florida (formed from mineralized ocean creatures that died millions of years ago), but these are limited, and therefore phosphorus fertilizers are expensive.
Water Cycle
Water is essential to all life, so the cycling of water keeps life on earth going. The water cycle, also called the hydrologic cycle, or hydrological cycle, incorporates the evaporation of water from liquid into gas (water vapor). Water vapor is the most important and the least often mentioned greenhouse gas. As the largest contributor to the greenhouse effect, it is essential to the process that controls and moderates the temperatures on earth. At the correct humidity, and temperature of the air, and atmospheric pressure (barometric pressure), water will change its state from a gas to a liquid. It condenses much as it does from the air onto the outside of a cold glass of soda on a hot summer day. This moisture in the air is constantly forming clouds, and droplets are continually forming. The smallest droplets are light enough to float in the air. When they bump into each other and finally become large enough, they fall back to the earth as either rain or snow, or hail, or sleet, depending on the temperature and the specific wind, humidity, and barometric pressure. The snow eventually melts. The rain eventually flows into streams and lakes and into the ocean. And it continually evaporates and condenses, and evaporates and condenses. And so the water cycle continues supporting the cycle of life.
BEHAVIORAL BIOLOGY
Another integral part of ecology is the study of animal behavior, sometimes called ethology (or biopsychology). These and related words are combined in many ways to describe the research of certain specialists. Behavioral ecology primarily concerns what animals do in natural environments, particularly with regard to adaptation, natural selection, and evolution. More recently, we see fields emerging such as environmental psychology and ecological psychology. Both of these involve the relationship people have with their surroundings. Humans have always been very human centric, that is, we think of nearly everything in terms of ourselves. When we think of why ecology is important, we think more of preserving natural ecosystems for our own benefit, and less of preserving nature just because that is the right thing to do. More people are appreciating the importance of natural systems. We are appreciating animals in new ways; even understanding animals have feelings and emotions and deserve some rights too. Plants are also beginning to be viewed in terms of their behavior.
Each organism's behavior is often related to its physiological features such as size, weight, and color, as well as its structure and the function of its parts. The selective pressures that help shape each species' characteristics include the environment, food, predation, and reproduction. There are adaptations found among each species that have to do with all of the above. Understanding these adaptations helps one to understand ecosystem complexity.
Ecology and behavior, as well as evolution, are all closely related. The study of one invariably includes the others. For instance, in the discussion of the competitive exclusion principle in this chapter, the concepts of niche, habitat, predation, dominance, species diversity, and community dynamics were all invoked. Symbiosis, commensalism, parasitism, and mutualism, all of which have to do with behavior, are discussed below, with brief definitions of each of the major terms.
Most animals have an area in which they spend the majority of their time. This includes where they may interact with other members of the same species, look for food, rest, and sleep. Such an area is called a home range. This differs from an animal's territory, which is a subset of its home range. A territory is just the area that is defended, usually from the other members of the same species.
Not all organisms, but many, are social, and their social behaviors vary from species to species, and often from population to population. Some animals have social hierarchies, relationships in which the status, or position, of each animal with regard to the other members of the group is well established, either by some aggressive behavior, or just by relative size, age, sex, or developments over time as animals grow, age, mature, or die, leaving new openings in the hierarchy.
Some social aggregations amount to more than just simple “pecking orders” of dominance–subordinance relationships. While certain species have very little, if any, division of labor within their social structure, other species have populations that function as a society. A society is a long-lasting arrangement of animals in which individuals are constantly changing, but the continuity of the social structure remains intact.
Symbiosis is another arrangement whereby species interact in a much more intimate way. It sometimes means living together, but usually the term is used in reference to an interaction from which both species derive a mutual benefit. Normally, symbiosis is broken into three categories:
- Mutualism – an arrangement between species in which each benefits from another and neither is harmed by the relationship.
- Commensalism – one species benefits while the other is neither harmed nor benefited by the relationship.
- Parasitism – one species benefits and the other is harmed. More often than investigators realize, the individuals we believe are being harmed may derive some benefit from this relationship too.
Generally, the only difference between parasitism and a predatory−prey relationship has to do with size. Often parasites are thought of as small animals that live on or in their host. Those that live on a host are ectoparasites and those that live inside their host are endoparasites.
Predation, the feeding of free-living organisms on other free-living organisms, is one of the main forces affecting species adaptation and change over time (both behavioral change during a lifetime and genetic change over time). Organisms that reduce their chance of being eaten may accomplish this through self-defense, escape, or hiding. Generally, the net positive effect is that they increase their chances of survival and, therefore, of passing on offspring to the next generation.
The different types of defense are physical defense, such as that employed by thorns or spines, and chemical defense, such as chemicals that render the individual bad tasting or poisonous. With plants, if a chemical defense is effective, perhaps only one leaf is eaten. Many animals also have chemical defenses, which may be advertised with bright warning coloration: aposematic coloration warns potential predators to avoid these bad-tasting or poisonous organisms. There are even aposematic sounds and odors.
CAMOUFLAGE
Many organisms blend in with their background. Camouflage is a means of disguise that conceals an organism. As humans, we often see camouflage in visual terms (animals colored or shaped to blend in with their background), largely because we're visually oriented. However, there are other means of disguise, such as those that make an animal difficult to hear, smell, or feel. Both prey and predators may be well camouflaged. Not only animals use camouflage, but many plants also do too. However, only a limited number of biologists have recorded such instances. The basic categories of camouflage are:
- Concealing or cryptic – a plant or animal hides by blending in with its background, by being the same color or by having a similar pattern. For instance, an animal that lives on sand might be sand-colored, or one that lives in the trees might be green to blend in with the foliage.
- Countershading – instead of being the same color all over, a plant or animal may be darker above and lighter below, enabling it to blend in better with its background as viewed from below or above. Such is the case with many sea birds, and with many fish, so they blend in with the sky when viewed from below, and they blend in with the water when viewed from above.
- Disguise – when plants or animals look like something else. An insect might look like a leaf, stick, or a thorn.
- Disruptive – when animals have patterns, spots, bands, or stripes that break up their outline (silhouette) or make them blend in, so they don't stand out. Rather than being a uniform color, an animal may have blotches or patches that make it look like part of the vegetation, or whatever the organism is blending in with. A type of disruptive camouflage may be called snakeruptive. This is what we often see with snake bands or stripes. When a predator focuses on a snake's bands or stripes when the snake is not moving, the predator gets ready to attack. Then when the snake begins to move in an attempt to escape, the brain of the predator may have trouble switching fast enough to what the blend of moving bands looks like, or may have trouble finding the snake when the stripe disappears because the snake quickly moved on, and then into a hole or under some leaves, or into the water.
- Mimicry – when plants or animals look like another bad-tasting or dangerous species.
MIMICRY
Different species sometimes resemble one another for reasons of defense. For instance, should coloration, pattern, or design be aposematic, many of several different species may evolve a similar aposematic coloration because predators learn to avoid anything looking like that. Such a phenomenon, where both the model and the mimic are unpalatable, is known as Mullerian mimicry. Interestingly, there are some species that are not advertising an unpalatable chemical or physical defense, even though they still resemble other aposematically colored species. These animals are called mimics because they mimic a model, and the phenomenon is termed Batesian mimicry.
WATER TREATMENT, RECYCLING, AND COMPOSTING
The sections provided above are covered in most biology texts when discussing ecology; however, it's important to remember that when people think of ecology, they often conjure up thoughts related to pollution abatement; issues such as water treatment (sewage treatment), mining, energy production (wind energy, solar energy, burning wood, burning coal, oil, gasoline, natural gas), recycling, and composting. These practices help with producing energy in a clean way, as well as with the disposal of materials that humans have used and maybe conserved, reused, or discarding of materials in a cleaner and more sustainable manner. In my ecology courses, I teach sections on these topics, and when possible I take classes on field trips to landfills, water treatment plants, nuclear power plants, solar energy plants, wind turbines, and all the other options available. I wish people thought more about preserving natural habitats, with all the species living in healthy communities, instead of merely waiting until a species is endangered and then trying at the last minute to save it. I wish we were more sensitive to how we develop areas, so we could plan for the future when there will be more people, and fewer wide-open wild places. I present the pros and cons of each option; we discuss the costs, and the history and implementation of environmental laws, rules, and regulations involved with issues involving environmental issues such as conservation, wastes, air pollution, water pollution, recycling, and resource recovery.
KEY TERMS
| abiotic | community | extirpation |
| aliens | competition | food chain |
| alpine tundra | competitive exclusion | food web |
| aposematic coloration | principle | freshwater |
| atmospheric cycles | contest competition | wetlands |
| autotroph | continental shelf | fundamental niche |
| bacteria | coral reef | fungi |
| Batesian mimicry | countershading | generalists |
| behavior | cryptic coloration | global cooling |
| behavioral ecology | cyanobacteria | global warming |
| biodiversity | (blue-green bacteria) | gradient zone |
| biogeochemical | death rate | greenhouse |
| cycles | deciduous | effect |
| biomass | decomposer | gross primary |
| biome | defense | productivity |
| biopsychology | denitrifying bacteria | habitat |
| biotic potential | density | herbivore |
| birth rate | density-dependent | heterotroph |
| blue-green bacteria | factors | home range |
| (cyanobacteria) | density-independent | humidity |
| boreal forest | factors | humidity |
| (taiga) | desert | interspecific |
| camouflage | dispersion | intraspecific |
| carbon cycle | disruptive coloration | life tables |
| carnivore | dominance | limnologists |
| carrying capacity | ecological | littoral zone |
| chaparral (temperate | homeostasis | logistic curve |
| shrubland) | ecology | longevity |
| chemical defense | ecosystem | marine |
| chemosynthesis | ecosystem | communities |
| climate | collapse | mass extinction |
| climate change | ectoparasite | microclimate |
| climax community | endoparasite | mimicry |
| clumped spacing | estuary | mimic |
| commensalism | ethology | model |
| Mullerian mimicry | producer | taiga (borial forest) |
| mutualism | quaternary consumers | temperate deciduous forest |
| natural selection | rainfall | temperate desert |
| nekton | random spacing | temperate evergreen forest |
| net productivity | realized niche | temperate forest |
| niche | recruitment | temperate grassland |
| nitrifying | reproductive strategy | temperate rain forest |
| bacteria | rivers | temperate shrubland (chaparral) |
| nitrogen cycle | scramble competition | temperate woodland |
| nitrogen-fixers | secondary consumer | temperature |
| omnivore | secondary productivity | territory |
| open ocean | sedimentary cycle | tertiary consumers |
| parasitism | social hierarchy | tidal marsh |
| pecking order | society | timberland |
| permafrost | specialists | transitional zone |
| phosphorus cycle | species diversity | trophic level |
| physical defense | species turnover | tropical rain forest |
| plankton | standing crop | tropical savanna |
| ponds and lakes | streams | tropical seasonal forest |
| population | sublittoral zone | tropical thornwood |
| population growth | suburban ecosystems | tundra |
| predation | succession | uniform spacing |
| predator | succulent | urban ecosystems |
| primary consumer | survivorship curve | weather |
| primary productivity | symbiosis |
SELF-TEST
Multiple-Choice Questions
- Ultimately, the __________ is the energy source in most ecosystems.
- sun
- moon
- stars
- gravity
- air
- Light energy is used by plants to convert, during photosynthesis, __________ and __________ into carbohydrates.
- water, carbon dioxide
- bread, water
- oxygen, water
- fats, proteins
- oxygen, carbon dioxide
- Organisms such as plants, that manufacture their own organic food molecules from inorganic substances, are the __________, and are often called __________.
- autotrophs, decomposers
- chemosynthetics, herbivores
- secondary consumers, tertiary consumers
- predators, carnivores
- producers, autotrophs
- Organisms that acquire food from dead plants and animals are called __________.
- herbivores
- heterotrophs
- decomposers
- chemosynthetics
- predators
- Organisms that eat plants, especially those that eat only plants, are called __________.
- predators
- carnivores
- autotrophs
- herbivores
- producers
- Animals that eat both plants and animals are called __________.
- pyramid builders
- food webbers
- omnivores
- detrivores
- omnitrophs
- The major terrestrial communities, those areas with distinct categories and types of organisms, are often called __________.
- rain forests
- biomes
- ecoclines
- forests
- prairies
- Characterized by more than 400 cm (over 150 in.) of rainfall and warm temperatures during most of the year, rather than just during a specific warm or rainy season, __________ are wet and green all year round.
- arctic tundra
- tropical oceans
- boreal forests
- tropical rainforests
- tropical seasonal forests
- Which of the following has more species of deciduous trees than species of trees that keep their leaves or needles all year long?
- tropical rain forests
- tropical seasonal forests
- temperate zone forests
- a and b
- b and c
- Communities that have less than 20 cm (8 in.) of rain a year are known as __________.
- deserts
- rain forests
- grasslands
- all of the above
- none of the above
- The newest, fastest-growing ecosystems being developed by people where they move in large numbers are known as __________.
- tropical rainforests
- salt marshes
- tundra
- streams and rivers
- urban and suburban ecosystems
- Much of America's Midwest and the Ukraine, which now support productive agriculture, were originally __________.
- tropical rainforests
- boreal forests
- deserts
- tundra
- temperate grasslands
- The __________ forest, often called the taiga, stretches through much of Canada and Siberia. This biome is dominated by spruce trees, pines, and firs that do well under what would be harsh conditions for most other species.
- tundra
- boreal
- temperate
- tropical
- hardwood
- Vegetation above the tree line primarily consists of __________.
- sedges and grasses
- mosses and lichens
- herbaceous flowering plants
- dwarf woody shrubs
- all of the above
- The area between the high and low tide marks is known as the __________.
- littoral zone
- coral reef
- shore
- rocky intertidal zone
- beach
- Along with estuaries and tropical rain forests, __________ are among the most productive ecosystems in the world.
- deserts
- tundras
- beaches
- coral reefs
- temperate shrublands
- Specific biologically active minerals cycle through plants and animals and the environment; these cycles are known as __________.
- biogeochemical cycles
- pyramid of numbers
- pyramid of biomass
- biomes
- none of the above
- The increase in the average worldwide atmospheric temperatures, attributed to additional CO2 in the atmosphere, is known as the __________.
- bicarbonate cycle
- carbon cycle
- nitrogen cycle
- greenhouse effect and climate change
- acid rain effect
- Because amino acids and proteins need nitrogen, and because gaseous nitrogen is not the form that can be readily used by most organisms, it is through the conversion of gaseous nitrogen, by certain bacteria and blue-green bacteria, into an aqueous form, that this nitrogen becomes available to other organisms. This conversion is known as __________.
- nitrogen-fixing
- nitrogen-breaking
- nitrogen-mending
- alchemy
- composting
- Because fixed nitrogen is so important to plants, it is a key ingredient in many fertilizers. However, because this fertilizer is so expensive, farmers often choose to rotate their crops, occasionally growing plants with symbiotic nitrogen-fixing bacteria living in nodules attached to the roots. Such plants are members of the __________ family.
- rose
- buttercup
- lily
- heath
- pea
- The bacteria that live under anaerobic conditions (no oxygen), such as those living deep in the soil or in the mud, complete the nitrogen cycle by returning nitrogen back to the air. Such bacteria are known as __________.
- sedimentary bacteria
- atmospheric bacteria
- phosphorus bacteria
- nitrifying bacteria
- denitrifying bacteria
- Plotting the fate of the young and their chances of survival at each age enables researchers to construct __________.
- survivorship curves
- increase curves
- biotic potential curves
- carrying capacity curves
- niche breadth curves
- Since the biotic potential is higher for fruit flies than it is for people, flies have a higher __________.
- a
- d
- K
- r
- q
- The number of individuals of a particular species that a specific environment can support indefinitely, often referred to by the letter K, is known as the __________.
- carrying capacity
- specific environment
- species diversity
- population size
- ecological homeostasis
- Many species are being introduced to areas where they are not native; such exotic species are often referred to as __________.
- Greeks
- out-of-towners
- Martians
- aliens
- Batesians
- When individuals directly compete for a resource without regard to other individuals that may also be using that resource, it is known as __________.
- contest competition
- scramble competition
- realized competition
- unrealized competition
- lack of competition
- Most animals have an area where they spend the majority of their time. This includes where they may interact with other members of the same species, look for food, rest, and sleep. Such an area is called a __________.
- territory
- niche
- home range
- symbiosis
- habitat
- Long-lasting arrangements of animals where individuals change over time due to births and deaths, but the continuity of the social structure remains intact, are known as __________.
- pecking orders
- dominance–subordinance relationships
- mutualistic relationships
- symbiotic relationships
- societies
- An arrangement between species where each benefits from one another and neither is harmed by the relationship is known as __________.
- parasitism
- mutualism
- commensalism
- monogamy
- polygamy
- Instead of standing out, many organisms blend in with their background. Such a means of disguise that conceals an organism is known as __________.
- camouflage
- disruptive coloration
- chemical defense
- physical defense
- aposematic coloration
ANSWERS
- a
- a
- e
- c
- d
- c
- b
- d
- e
- a
- e
- e
- b
- e
- a
- d
- a
- d
- a
- e
- e
- a
- d
- a
- d
- b
- c
- e
- b
- a
Questions to Think About
- Compare and contrast the following terms: species, population, community, ecosystem.
- What are the differences between r- and K-selected species? Give two examples of each, and provide two examples of intermediary species that are neither, but have characteristics of both.
- Why does it work for some species to be generalists, while others are specialists? Compare the risks and benefits of these different life history strategies.
- Write a brief paragraph explaining four distinctly different scenarios in which four introduced species (imagined or real) have affected community dynamics in entirely different manners.
- Describe the conditions that would render a forest a climax community. What might change this climax community so it is no longer a stable assemblage of species?
- List 10 different ecosystems and briefly describe them.