20
Plant Kingdom: Plantae
In most classification systems, the plant kingdom Plantae includes several groups of simple photosynthetic organisms, sometimes known as eukaryotic algae, a reference to when people still grouped the prokaryotic blue-green bacteria with the algae. Euglenophyta, Chrysophyta, and Pyrrophyta, three phyla of unicellular eukaryotic algae often called plants, were described in Chapter 18. The term “true algae” is no longer a technical phrase since there are so many algal groups that are not directly related to one another. Nevertheless, the common terminology is presented in conjunction with the formal classification because it continues to be used in most texts. Of the following eukaryotic algae, Chlorophyta (green algae) are thought to be the ancestors of most modern plants because they contain the photosynthetic pigments chlorophyll a and b, as well as beta-carotene; they store their food reserves as starch, and their cell walls are composed of cellulose. They are mostly freshwater organisms, though some are marine.
Phaeophyta (brown algae) are almost all marine and are common in cooler oceanic regions. Rhodophyta (red algae) are mostly warm-water, marine species, though some are freshwater. Most of the common species of seaweed are members of the brown and red algal groups.
Each of the three preceding phyla varies according to: (i) types of photosynthetic they contain, (ii) type of food reserves stored internally, and (iii) components found in their cell walls.
These algae are not included in the plant kingdom merely because they photosynthesize, since many protists and monerans also have that capacity. Nor is tissue differentiation always a key factor in determining where to draw the line. Multicellular algae have no true roots, stems, or leaves. Their simple body form is termed a thallus, which is why they are sometimes called thallophytes. Many biologists call algae plants, but only lower plants.
The reason such matters are complex is that these organisms represent many different lineages and many different steps in a continuum connecting the most primitive forms of life, such as bacteria, to the most complex multicellular organisms. It isn't always clear just where one should draw the somewhat arbitrary lines that artificially separate each of the six described kingdoms – Archaebacteria, Eubacteria, Protista, Fungi, Plantae, and Animalia.
A reason the algae have been kept from the plant kingdom in some classifications is their reproductive structures. Higher plants have reproductive structures encased inside a protective wall of sterile cells that protect the developing zygotes before they are released from the female reproductive organs, where they were produced. Algae lack a protective wall of nondividing (sterile) cells, and their zygotes do not develop into embryos until after having been released from the female reproductive organs.
When lower plants and higher plants are placed in separate kingdoms, then the higher plants are sometimes called the Embryophyta since the female reproductive organs retain the zygotes until after they have developed into embryos. Embryophytes include most trees, shrubs, herbs, grasses, ferns, and mosses. In addition to the lower plants, the major groupings in the plant kingdom include the Bryophyta (mosses, liverworts, and hornworts) and the Tracheophyta, which include all the vascular plants (psilopsids, club mosses, horsetails, and ferns) and seed plants (gymnosperms and angiosperms). However, the name for vascular plants, which was Tracheophyta, meaning tube plant, has since been eliminated from the classification. (See Glossary for more on the above terms.)
MOVE TO LAND
True algae are mostly restricted to aquatic environments. The move to land was accompanied by many adaptations to what would otherwise have been a hostile environment. Out of water, plants were met with dry conditions, ultraviolet light, and nutritional problems. In addition, in the air these plants no longer benefited from the surrounding water's buoyancy, so some structural support became necessary.
Terrestrial plants had to obtain water by a new means, since they were no longer bathed in it. The water had to be both procured and then moved from its point of uptake to the other structures. In addition, the photosynthetic products had to be transported from their specialized photosynthetic structures to the other parts of the plant. Vascular tissues evolved that solved all these problems.
Excessive water loss from evaporation had to be curtailed, while the moist tissues necessary to allow gaseous exchange for metabolic and photosynthetic purposes had to be maintained. This led to highly evolved mechanisms that controlled overall water loss, while enabling structures to remain moist.
Reproductive needs also had to be modified in terrestrial environments, where flagellated sperm cells no longer had the surrounding water in which to swim. And special structures evolved that protected the early stages of embryonic development from desiccation.
The modifications that differentiate many terrestrial plants from their aquatic counterparts are understood only in terms of the factors affecting plants living in terrestrial versus aquatic environments. The following mechanisms have helped make it possible for embryophyte plants to inhabit terrestrial habitats:
- A waxy cuticle usually covers the aerial parts of the embryophyte plants, acting as waterproofing and preventing excessive water loss.
- All embryophytes are oogamous. That is, they have two types of gametes, one of which, the female, is typically the large, nonmotile egg cell, the oogamete.
- All embryophytes have multicellular sex organs covered with a layer of protective cells that are sterile. The male sex organs are known as antheridia, and the female are archegonia. Within the sex organs the gametes are protected from desiccation.
- All embryophytes have egg cells (oogametes) that are fertilized within the archegonia.
- While inside the archegonium, the zygotes develop into multicellular diploid embryos that obtain some of their water and nutrients from the parent plant.
- In addition to producing gametes, embryophytes produce spores, reproductive cells that develop directly into full-grown plants without first having to undergo fertilization by joining with another cell. The embryophyte sporangia produce spores that are covered with a protective jacket of sterile cells.
BRYOPHYTA: MOSSES, LIVERWORTS, AND HORNWORTS
The bryophytes represent about 25,000 species of mosses, liverworts, and hornworts. On the basis of any one characteristic it is difficult to distinguish them from the thallophytes, or from what are sometimes referred to as the true algae. In contrast to algae, which tend to be composed of either single cells or filaments (sheets of cells), which can intertwine to create more complex body structures, bryophytes are rarely filamentous, except during one stage in the life history of the mosses. Instead, bryophytes are composed of cells that form tissues called parenchyma; they are characterized by loosely fitting cells that have thin walls of cellulose. In between the cells are intercellular spaces incorporated within the cellular network.
In bryophytes, the principal photosynthetic pigments are chlorophyll a and b. These are biochemically similar to the pigments of the chlorophytes, from which bryophytes probably arose. Their energy reserves are stored in the form of starch, and their cell walls contain cellulose.
Bryophytes are usually terrestrial. But they remain somewhat dependent on their ancestral aquatic environment. This has kept their distribution limited to moist environments or to environments that are moist during a critical period each year. These small plants need water for their flagellated sperm cells to swim from the antheridia to the egg cells in the archegonia. Without any vascular tissue, their ability to move fluids internally across long distances is limited. And since xylem, the vascular tissue in higher plants, is necessary for support, the upper size limit of these plants without such supportive tissue is kept at a minimum.
All the bryophytes have an alternation of generations, with a sporophyte generation (diploid) and a gametophyte generation (haploid). Among the larger, more complex algae, most of which have an alternation of generations, there is some tendency toward a reduction of the gametophyte, multicellular stage, and an emphasis on the sporophyte, multicellular stage. In both the brown and red algae, the sporophyte generation is prominent, as is the case in the vascular plants. This tendency is not apparent in the bryophytes, in which the haploid gametophyte stage is clearly dominant. It is the leafy green gametophytic stage of the bryophytes that produces the gametes. These swim through moisture, present as a film of either rain or dew, from the antheridia to the archegonia. Here the sperm fertilize the eggs, producing zygotes that make diploid sporophytes by mitotic division. The sporophyte plant is attached to the gametophyte and grows directly from it. The sporophyte produces the sporangia, organs that contain and release the asexual spores. Spores are asexual because, unlike gametes, they never meet in a sexual union. (See page 73, Figure 4.5, Life cycle of a moss.)
The moss gametophyte generation possesses what appear to be, but aren't, true roots, leaves, and stems (see Figure 20.1). Bryophytes have no vascular tissue, a critical component in such structures. Root-like organs in plants without vascular tissues are called rhizoids; they function like roots, by anchoring the plant and absorbing water and nutrients. The stem is “stemlike” and sometimes is referred to as the stalk. The “leafy” parts are sometimes referred to as “leaves” because of a lack of better terminology.
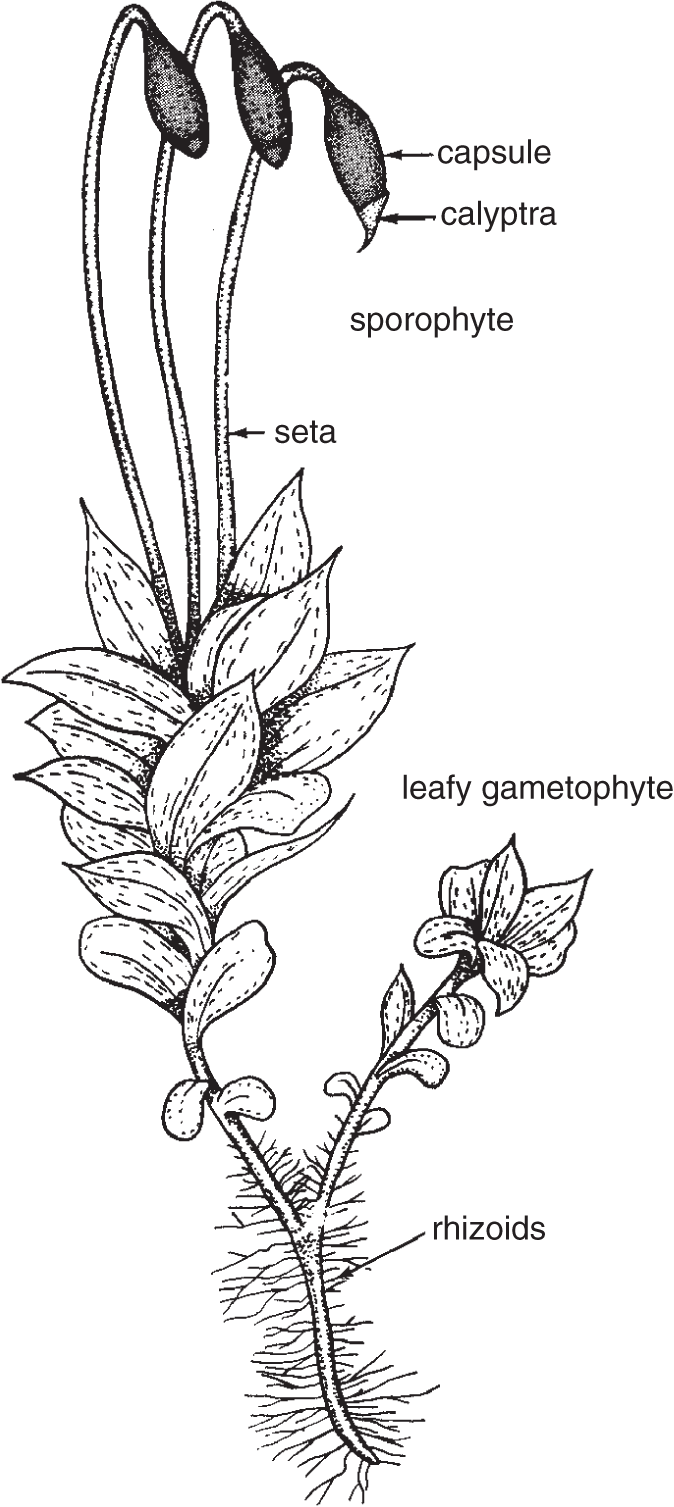
Figure 20.1 Moss, illustrating rhizoids, leafy gametophytes, and attached sporophytes. The same plant has sporophyte and gametophyte generation (sporophyte grows from the gametophyte).
VASCULAR PLANTS
Some of the earliest known vascular plants had roots that functioned as holdfasts and absorbed water. They also had vascular tissue for water and nutrient movement. This tissue also provided strength and helped hold the plant up in the air. In addition, these early vascular plants had a waxy cuticular layer covering the leaves for water retention. The fossil record indicates there was a trend toward the reduction of the gametophyte generation in favor of the more dominant sporophyte generation, which contained the sporangia. The earliest vascular plants probably produced only one kind of spore from one kind of sporangium, a process called homospory. After germination, these spores developed into gametophytes with antheridia and archegonia. They produced the sperm and eggs, respectively. However, the trend toward heterospory (the production of two different kinds of spores) is also evident in the overall evolution of the vascular plants.
Following the evolution of the aquatic vertebrates, which took place about 500 million years ago, the first vascular plants to colonize the land appear in the fossil record; at about the same time, some arthropods also began colonizing terrestrial habitats.
A significant innovation unique to the vascular plants is the seed, which consists of an embryo and some stored food enclosed within a protective coat. The earliest known fossilized seeds date back 350 million years.
Five major groups of tracheophytes, or vascular plants, are discussed below. To date, more than 260,000 species of vascular plants have been described.
PSILOPHYTA
Psilophytes are the only vascular plants that do not have roots or leaves. This is the most primitive phylum (many botanists prefer to call it a division) of vascular plants is Psilophyta. The psilophytes resemble some of the branching filamentous green algae (Chlorophyta) from which they probably arose. The psilophytes have true stems, with simple vascular tissue, branching from slender rhizomes, which are elongated, underground, horizontal stems; they are not true roots. The rhizomes have unicellular rhizoids − thin, root-like structures that are similar to root hairs. No true leaves are present, although the aerial stems are green and perform photosynthesis.
Sporangia develop at the ends of the stems and produce haploid spores.
These fall to the ground and give rise to the subterranean gametophytes. The gametophytes bear both archegonia and antheridia. Each gametophyte produces both eggs and sperm. The sperm travel to the eggs, where fertilization occurs. Then the diploid zygote begins developing into a sporophyte.
The psilophytes evolved during the Silurian period and thrived more than 300 million years ago. However, with the exception of three surviving species, the entire group is extinct. Many botanists believe some psilophytes evolved into the ferns.
LYCOPHYTA: CLUB MOSSES
Lycophyta, or the club mosses, also appeared during the Silurian, about 400 million years ago. They were among the largest and most dominant plants during the Devonian and Carboniferous periods. Toward the end of the Permian, they were superseded by more advanced vascular plants. About 1,000 species are still found throughout much of the world, although all are quite small and amount to little more than ground cover.
Club mosses have true leaves, stems, and roots. They may have evolved from algae that penetrated the ground, occasionally sending branches above the surface. Some of their leaves are specialized. Called sporophylls, these leaves have sporangia that bear spores. Many species have club-shaped structures called strobili at the ends of their stems. The strobili, formed from clusters of sporophylls, are the source of the group's common name. It should be pointed out, however, that the club mosses are not related to the true mosses, or bryophytes.
Some club mosses are heterosporous, having two types of sporangia. The larger spores, known as megaspores, develop into the archegonia-bearing female gametophytes. The other sporangia produce smaller spores, the microspores, which develop into antheridia-bearing male gametophytes.
SPHENOPHYTA: HORSETAILS
Another group of vascular plants is Sphenophyta, commonly known as the horsetails because that is what they look like. They appeared during the Devonian, around 360 million years ago, and dominated forests during the late Paleozoic era. About 250 million years ago, they began to decline. Today there are some 25 species left, most of which are relatively small.
They all have true roots, stems, and leaves. Living horsetails lack cambium, though many of the larger extinct forms had a vascular cambium. This is the layer of dividing cells in the trunk that produces the secondary tissues that allow plants to grow more tissues as they increase in height, width, and weight. As certain plants grow older, it is the continuing cell divisions in the vascular cambium that account for a plant's increase in girth. (In this case, girth is the measurement around the trunk. For comparison, you might measure the girth around a man's waist.)
The most common group of living horsetails, Equisetum, is homosporous; their spores give rise to gametophytes that bear both archegonia and antheridia.
PTEROPHYTA: FERNS
Many botanists feel that the pterophytes, or ferns, evolved from the psilophyte stock. Ferns have true roots, stems, and leaves. They first appeared during the Devonian, around 400 million years ago, and became important during the Carboniferous, about 300 million years ago. They then declined, along with the psilophytes, lycophytes, and sphenophytes. However, the decline wasn't nearly so drastic for the ferns as it was for their contemporaneous vascular plants; today there are about 11,000 fern species living throughout the world.
Most forms are quite small, though one group, the tree ferns, is the exception. They have a woody trunk with leaves (fronds) and sometimes attain heights of nearly 25 m (82 ft). This is possible because ferns have well-developed vascular systems containing xylem (water-transporting cells) and phloem (sugar-transporting cells). But, unlike the conifers and broad-leaved trees, ferns do not have any secondary growth, so it is not possible for the fern trunk to increase in girth. This creates an upper height limit for most ferns and is the reason why their stems often lie on the ground, with only the leaves growing upward.
The dominant stage in the fern's life cycle is the leafy diploid sporophyte (see Figure 20.2). The sporangia are often located on the underside of the frond. Those fronds containing sporangia are called sporophylls. The location of the sporangia on each fern depends on the species.

Figure 20.2 Mature fern sporophyte, illustrating leafy diploid stage.
Most ferns have just one type of spore. Under the proper conditions, all the spores develop into gametophytes that bear both archegonia and antheridia. The gametophytes never develop vascular tissue, and they rarely grow very large. Because their free sperm cells are flagellated and require moisture to reach the egg cells inside the archegonia, ferns tend to be restricted to moist environments. When transplanted, however, they have little trouble growing in drier environments, although fertilization will not occur. Where fertilization does occur, the zygote develops into the sporophyte without passing through a protected seed-like stage.
GYMNOSPERMS
In some classifications, the remaining vascular plants are placed in the group Spermatophyta. This group includes all seed-bearing plants, which includes the gymnosperms and angiosperms. Gymnosperm means naked seed, whereas angiosperm plants have seeds that are enclosed in an ovary. Gymnosperms include cycads, the ginkgoes (just one living member, the rest are now extinct), conifers, and others. The angiosperms include all the grasses, sedges, rushes, and other flowering plants.
It has been found that most of the groups called gymnosperms aren't closely related. Thus, there has been a tendency to list each category separately without placing them under the gymnosperm heading. Here, each of the groups that used to be classified as gymnosperms has been moved up to a taxonomic level that, among botanists, is often called the division. However, the term “phylum” is more useful.
A distinctive characteristic of the peculiar gymnosperm phylum known as seed ferns is not that its species don't produce spores, as true ferns do, but that instead they produce seeds. It is thought that the cycads and angiosperms (flowering plants) evolved from this group.
Another unusual group, the Gnetophytes, includes about 70 desert and montane (living on mountains) species with flower-like reproductive structures. These plants are heterosporous. The dominant stage in their life cycle is the sporophyte. They are dioecious (have separate male and female plants), produce pollen, and bear small, naked seeds. The Gnetophytes appear to be relatives of the ginkgo and the conifers. Both the seed ferns and the Gnetophytes were important components of Carboniferous forests.
CYCADOPHYTA
The earliest fossils of the ancient division of plants called Cycadophyta, or the cycads, date to about 240 million years ago. They reached their climax during the Triassic and Jurassic, about 200 million years ago, and then declined steadily. Now only about 100 species survive.
The unbranched, erect trunk of the cycad bears a crown of leaves that resembles a tree fern or a palm tree, except for its large, upright cones. The tallest cycads reach about 20 m (65 ft), but most are less than 2 m (6.5 ft). Their leaves are pinnately compound (one vein bearing many leaflets). All the cycads are confined to the tropics, except those grown ornamentally.
Each species has separate male and female plants. The males bear staminate cones (with pollen) and the females have seed-bearing, or ovulate cones. There is one exception; instead of bearing cones, the female cycads in the genus Cycas have seed-bearing megasporophylls that resemble leaves.
Cycad pollen is carried by the wind to the naked ovules. Upon landing, the pollen grain develops a tubular extension called the pollen tube. This produces flagellated sperm cells that swim a short distance to reach the egg cells.
GINKGOPHYTA
Before most members of the plant division Ginkgophyta became extinct, the group that now consists of only the ginkgo was extremely diverse. It was an important constituent of the Mesozoic forests around the world, especially during the Jurassic period, about 150 million years ago. The genus Ginkgo is thought to extend all the way back to this period, which is why ginkgoes are often called “living fossils.” The group began to decline toward the end of the Mesozoic and continued its decline through the Tertiary until reaching its current situation in which there is just one surviving member, Ginkgo biloba. The ginkgo, as it is commonly called, is native to western China, where it has been in cultivation for hundreds of years. During the past hundred years, the ginkgo has become a very popular street tree throughout much of the rest of the world.
The fan-shaped leaves are borne on a long stalk (the petiole). The species is dioecious. Male trees have catkin-like pollen-bearing cones that, although not very conspicuous, are similar to the male cones found among the conifers. Being heterosporous, the pollen-bearing cones produce microspores that are carried by air. Some land on the female tree's mature ovules. The female gametophytes are buried in the ovule, where the small archegonia are formed. The entrance to the ovule is made through a small opening, the micropyle. The microspores are drawn into the micropyle. They lie next to the ovule and develop into male gametophytes that grow into the ovule. The male gametophyte releases motile sperm cells, one of which fuses with the egg nucleus to form a zygote, from which the embryo develops. Tissues grow around the embryo and form a true seed with a soft outer fleshy layer.
CONIFEROPHYTA: CONIFERS
The phylum Coniferophyta includes plants usually referred to as conifers, such as species of pine, spruce, fir, hemlock, cypress, redwood, larch, juniper, and yew, as well as others that aren't as well known. The conifers include about 50 genera, comprising over 600 species. Their fossil history dates back to the late Carboniferous, some 300 million years ago.
Although the cone is a conspicuous feature distinguishing many members of this group, it does not appear on all conifers. For example, juniper berries are actually small cones with fleshy scales. And yews have seeds surrounded at the base by a fleshy, berry-like pulp. In biological terms, the distinction between cones and more berry-like forms of reproduction is significant in terms of the mode of dispersal used by the conifers involved. Cones are adapted for wind dispersal of their seeds. The others are adapted to animal dispersal.
Trees are the diploid sporophyte stage of the life cycle. The cones are actually tight clusters of modified leaves known as the sporophylls, which, in the case of the pines, are also known as pine cone scales. Each sporophyll contains two sporangia in which haploid spores are produced through meiosis. These trees are heterosporous. The large female cones contain the sporangia that produce megaspores; the small male cones contain sporangia that produce microspores. All seed plants, both the gymnosperms and angiosperms, are heterosporous.
Each pine scale contains two sporangia; each sporangium has a small opening, the micropyle. Meiosis occurs inside the sporangium, producing four haploid megaspores, three of which disintegrate. The remaining megaspore, through repeated mitotic divisions, becomes the female gametophyte, which, unlike that in the ferns, is considerably reduced in size. It is located within the cone. The gametophyte is not free-living, nor does it contain chlorophyll. Each female gametophyte produces several archegonia in which egg cells develop. Together the entire structure consisting of the integument, the sporangium, and the female gametophyte is called an ovule.
Within the male cones are sporangia that produce microspores. These become pollen grains, which develop a thick coating, resistant to desiccation. They have small wing-like structures that help them along when carried aloft by the wind. Inside the pollen grains, the haploid nucleus divides mitotically and the pollen grains become four-celled. Two of these cells degenerate. When the sporangia burst, millions of mature pollen grains are released. This is the male gametophyte stage cycle.
Most of the pollen grains that land on the female cones fall between the scales. Some of these land near the opening of the micropyle. When a pollen grain lands, touching the end of the sporangium inside the micropyle, it grows a pollen tube. The germinated pollen grain, which is now the pollen tube, grows down through the sporangium and penetrates one of the archegonia of the female gametophyte. There the tube releases its nuclei, which, in this case, are sperm that developed when the cells in the pollen tube were dividing. The sperm fertilize the egg and the resulting zygote produces an embryo sporophyte. This is still contained within the female gametophyte inside the pine cone. Finally, the cone sheds its ovules, more commonly known as seeds, which grow into adult sporophyte trees.
ANTHOPHYTA/ANGIOSPERMS: FLOWERING PLANTS
The phylum Anthophyta, more commonly known as the angiosperms, or flowering plants, also called anthophytes, are by far the most successful group of living plants, totaling over 350,000 described species. It is their flowers and fruit that differentiate them from all other plants. The earliest known angiosperm-like plants first appear in the fossil record during the Jurassic, about 150 million years ago. But there is some speculation that angiosperms may have existed as long ago as the Permian, about 250 million years ago. It was not until the Cretaceous, 100 million years ago, that there was a rapid decline in the dominance of the gymnosperms. Then, suddenly, and from that time on, the fossil record reflects the diverse spectrum of angiosperms dominating most flora throughout the world.
Among all angiosperms, the diploid sporophyte, retaining and nourishing the gametophyte, dominates the life cycle. And it is the small gametophyte that retains and nourishes the immature eggs and nonmotile sperm during development. Flowering plants are heterosporous, having two different-sized spores, and oogamous, having sperm and eggs. Their zygotes develop into seeds and fruit that are highly evolved for protection and dispersal. One of their most distinguishing characteristics is the unique reproductive system involving flowers. The floral structures coevolved with the plant's pollinating vectors, which are modes of transferring pollen from one flower to another, such as the wind, insects, and other animals. These vectors appear to have helped angiosperm dispersal into habitats where other plants might not have reached so readily.
Early angiosperms were probably pollinated by the wind. The ovule of modern gymnosperms exudes a sticky substance that traps windborne pollen grains. Similarly, the first angiosperms probably had such a pollinating mechanism.
Some insects, such as certain beetles, may have become dependent on readily accessible sticky, sugary droplets produced by ovules. Insects traveling from one ovule to another may have inadvertently carried pollen with them, conferring a reproductive advantage to some plants, helping to pass on the genes of plants that had larger nectar-secreting organs (nectaries) and other structures that lured insects. (Nectar is defined in the Glossary.) Plants with certain scents and brightly colored flower parts, arranged in ways that signaled potential pollinators, had selective advantages. These are all insects that specialize in nectar consumption. Because certain insects carried pollen from flower to flower, many became vital to their host plants. Both the plants and the insects benefited from this relationship, and they coevolved. By the early Cenozoic, about 65 million years ago, many modern groups of flowers, as well as the bees, wasps, moths, and butterflies that pollinated them, had already evolved.
TYPICAL FLOWER
Plants have reproductive structures (often called flowers) with cells that divide meiotically, much as humans have cells in our testes and ovaries that divide meiotically, and in the process they produce our sperm and eggs. What makes plants and humans different, besides everything that is obvious and you already know, is that plants often have many flowers, and because of this, they can produce many seeds. Each seed is formed after two cells that were formed from meiotic divisions fuse, much as a human sperm and eggs fuse, forming a zygote. Each zygote grows into a genetically unique seed. Because plants produce so many seeds each year, they can adapt rapidly to new viral, fungal, and bacterial-borne disease, and they can adapt to attacks from insects and other animals. For these reasons, flowers are remarkable structures that have helped plants survive and thrive and continually change in ways we are just beginning to appreciate.
The typical flower consists of four concentric circles or whorls of modified leaves. Each has a different name: sepals, petals, stamens, and carpels, each of which is attached to the same receptacle, which is the part of the modified stem that the flower parts grow out of. Such flowers are said to be complete (see Figure 20.3). Plants with flowers that lack any of the first four elements are said to have incomplete flowers.
The sepals, which enclose the other parts of the flower inside the bud, are usually green because they photosynthesize. Together, the sepals constitute the calyx. After the sepals, the next structures that grow from the receptacle are the petals. Together, all the petals are called the corolla. The calyx and the corolla constitute the perianth.
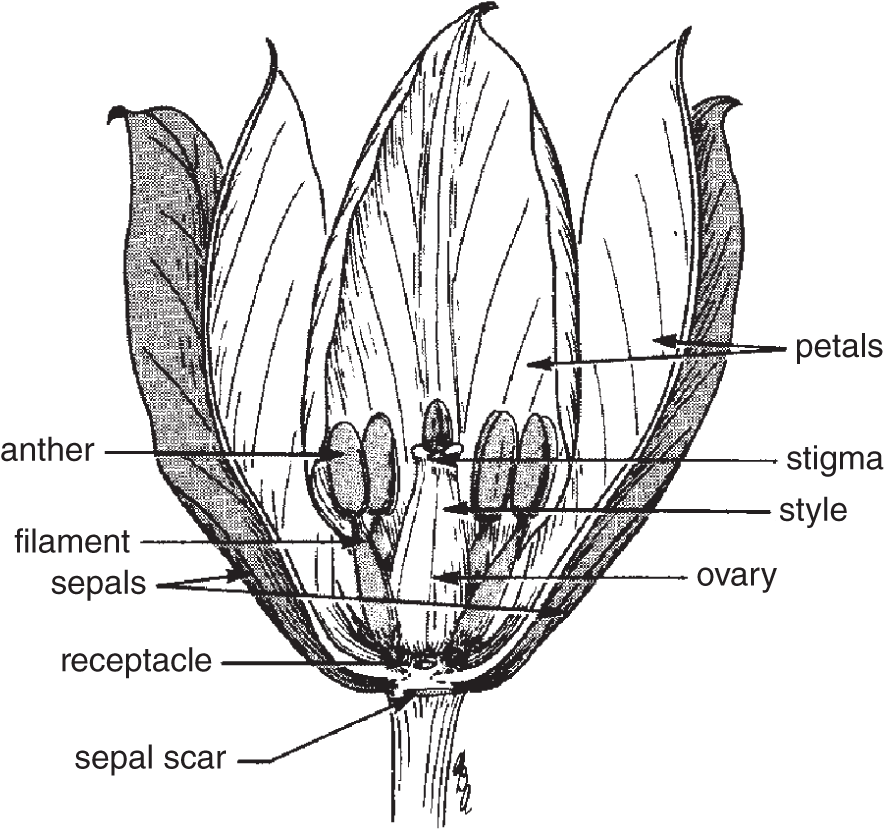
Figure 20.3 Diagram of a tulip flower, illustrating different floral parts.
Next, continuing distally (outward) along the modified stem, are the stamens, each of which consists of a long, slender stalk, and a filament, which bears pollen on the specialized portion at the tip, known as the anther.
The central part of the flower, which is the part farthest along the receptacle, is the carpel. This is composed of modified floral leaves that have become folded over to protect the ovule from otherwise predaceous pollinators. The pistil consists of one or more carpels in the center of the flower. Generally, the pistil has an expanded base, the ovary, in which are one or more sporangia. These sporangia are called the ovules. Each one divides meiotically, creating four haploid megaspores, three of which usually disintegrate. The remaining megaspore divides mitotically to produce the female gametophyte. One of the cells near the micropylar end becomes the egg cell, and some polar bodies remain.
Each anther consists of four sporangia. These divide meiotically to produce haploid microspores that mature into pollen grains, each with two nuclei. These pollen grains are the male gametophytes. When the pollen reaches the stigma, the receptive portion of the style extending above the ovary, a pollen tube grows through the stigma and the style into the ovary. When the tip of the pollen tube reaches the ovule, it enters the micropyle and releases both haploid nuclei into the female gametophyte (embryo sac). One of the haploid nuclei, which are known as sperm nuclei, although they are not motile, fertilizes the egg, and the zygote develops into the sporophytic embryo. Two polar nuclei from the female gametophyte combine, forming a diploid fusion nucleus. This in turn combines with the second sperm to form a triploid nucleus that divides mitotically and creates triploid tissue known as the endosperm. This tissue surrounds the embryo and acts as the stored food upon which the developing sporophytic seed feeds. Endosperm constitutes the nutritious part of most grains and many seeds that are eaten by much of the world's human population.
After fertilization, the angiosperm ovary, which surrounds the embryo, develops into the fruit. This protects the embryo from desiccation during the early stages of development. Later, the fruit acts as an agent of dispersal, either by being blown by the wind or falling to the ground and rolling. An animal may also aid in the dispersal process by carrying the fruit in its fur or ingesting the fruit and depositing it in feces.
The angiosperms are composed of two subgroups, the Monocotyledonae and the Dicotyledonae. Monocots, as they are often called, have embryos with one cotyledon, the leaf-like structure composing much of the seed with its endosperm, which is composed of nutrients used during germination. Monocots usually have parallel leaf venation. The flower parts are normally in multiples of three. Most forms lack a vascular cambium. The monocots include grasses, lilies, iris, and orchids. The flowers of grasses have greatly reduced petals, stamens, and carpels, and are thought to have evolved from a lily-like ancestor.
In contrast, the dicots have embryos with two cotyledons, their leaf venation is generally netted, and the lower parts are typically in groups of four or five. In addition, those dicots with secondary growth have a vascular cambium. All the flowering plants not included in the monocots are dicots; they are the most numerous and, in many respects, the most successful plants alive.
KEY TERMS
| algae | bryophytes |
| alternation of generations | calyx |
| angiosperms | carpels |
| anther | Chlorophyta |
| antheridia | club mosses |
| Anthophyta | complete flowers |
| anthophytes | cone scales |
| archegonia | cones |
| brown algae | Coniferophyta |
| Bryophyta | conifers |
| corolla | Monocotyledonae |
| cotyledon | mosses |
| Cycadophyta | nectaries |
| cycads | oogamete |
| Dicotyledonae | oogamous |
| dioecious | ovary |
| Embryophyta | ovulate cones |
| embryophyte | ovules |
| endosperm | perianth |
| ferns | petals |
| filament | Phaeophyta |
| flower | phloem |
| flowering plants | pistil |
| Gametophyte generation | Plantae |
| Ginkgo | pollen |
| Ginkgo biloba | pollen-bearing cones |
| Ginkgophyta | pollen grains |
| Gnetophytes | pollen tube |
| green algae | Psilophyta |
| gymnosperms | psilophytes |
| heterospory | psilopsids |
| homospory | pterophytes |
| hornworts | receptacle |
| horsetails | red algae |
| incomplete flowers | rhizoids |
| leaves | rhizomes |
| liverworts | Rhodophyta |
| Lycophyta | roots |
| megaspores | seed |
| megasporophylls | seed ferns |
| micropyle | seed plants |
| Microspores | sepals |
| Sphenophyta | thallophytes |
| sporangia | thallus |
| spores | Tracheophyta |
| sporophylls | triploid |
| sporophyte generation | true stems |
| stalk | vascular cambium |
| stamens | vascular plants |
| staminate cones | vascular tissue |
| stigma | waxy cuticlar layer |
| strobili | xylem |
| style |
SELF-TEST
Multiple-Choice Questions
Algae, Bryophyta, Vascular Plants, and Psilophyta
- The following is thought to be the group that is the direct common ancestor of most modern plants:
- Pyrrophyta (dinoflagellates)
- Chlorophyta (green algae)
- Phaeophyta (brown algae)
- Rhodophyta (red algae)
- Euglenophyta (euglenoids)
- Multicellular algae have __________.
- true roots
- true stems
- true leaves
- considerable tissue differentiation
- none of the above
- The simple body form of multicellular algae is called a __________.
- thallus
- sporangium
- micropyle
- cone
- angiosperm
- Because the simple body form of multicellular algae is called a thallus, as a group these organisms are sometimes called __________.
- sporantiophytes
- conifers
- angiosperms
- algophytes
- thallophytes
- Algal reproductive structures lack __________.
- a protective wall of nondividing cells
- zygotes that develop into embryos after having been released from the female reproductive organs
- a protective wall of sterile cells
- all of the above
- none of the above
- Bryophytes include __________.
- mosses
- liverworts
- hornworts
- all of the above
- none of the above
- The vascular plants include __________.
- club mosses
- horsetails
- ferns
- seed plants
- all of the above
- Many embryophytes can inhabit terrestrial habitats because __________.
- they have multicellular sex organs covered with a layer of protective, sterile cells
- their sporangia produce spores that are covered with protective sterile cells
- they have a waxy cuticle that usually covers the aerial parts of the plants
- all of the above
- none of the above
- Bryophytes differ from thallophytes (true algae) in that __________.
- bryophytes are rarely filamentous, except during one stage in the life history of mosses
- bryophytes are composed of cells that form tissues
- bryophytes are usually terrestrial, remaining somewhat dependent on their ancestral aquatic environment
- all of the above
- none of the above
- Bryophytes __________.
- need water for their flagellated sperm cells to swim from the antheridia to the egg cells in the archegonia
- lack the ability to move fluids internally across long distances
- lack the support of xylem
- all of the above
- none of the above
- All bryophytes have a(n) __________.
- alternation of generations
- sporophyte generation
- gametophyte generation
- all of the above
- none of the above
- A significant innovation unique to the vascular plants is the seed, which __________.
- consists of an embryo
- has some stored food
- is enclosed within a protective coat
- all of the above
- none of the above
Lycophyta, Sphenophyta, and Pterophyta
- Lycophytes have club-shaped structures at the ends of their stems known as strobili, which form clusters of sporophylls. It is from these structures that the group's common name, __________, has been derived.
- strobili mosses
- club-shaped mosses
- club mosses
- sporophyll mosses
- cluster mosses
- Club mosses have __________.
- true leaves
- true stems
- true roots
- all of the above
- none of the above
- Horsetails (Sphenophyta) have __________.
- true leaves
- true stems
- true roots
- all of the above
- none of the above
- Ferns (Pterophyta) have __________.
- true leaves
- true stems
- true roots
- all of the above
- none of the above
Gymnosperms, Angiosperms, and Flowers
- Gymnosperms include __________.
- grasses
- sedges
- rushes
- all of the above
- none of the above
- Angiosperms include __________.
- cycads
- ginkgoes
- conifers
- all of the above
- none of the above
- The conifers include __________.
- pines
- spruce
- fir
- all of the above
- none of the above
- The conifers include __________.
- cypress
- redwood
- larch
- all of the above
- none of the above
- The conifers include __________.
- hemlock
- junipers
- yews
- all of the above
- none of the above
- The __________ are, in terms of numbers of species, the most successful group of plants.
- angiosperms
- gymnosperms
- ginkgoes
- club mosses
- ferns
ANSWERS
- b
- e
- a
- e
- d
- d
- e
- d
- d
- d
- d
- d
- c
- d
- d
- d
- e
- e
- d
- d
- d
- a
Questions to Think About
- Compare and contrast land plants with those that live in water (give examples).
- Describe the types of waterproofing used by plants, and what their functions are.
- What are the differences between algae and higher plants?
- What role did vascular tissue play in the evolution of plants?
- What is a flower? Describe its structure and function.
- How does alternation of generations apply to different plants?
- What are the characteristics that differentiate major groups of plants?
- How do plants differ from all the other groups of organisms discussed so far?
- What are seeds? And what is their significance?
- Describe an angiosperm and give examples of different types of angiosperms.