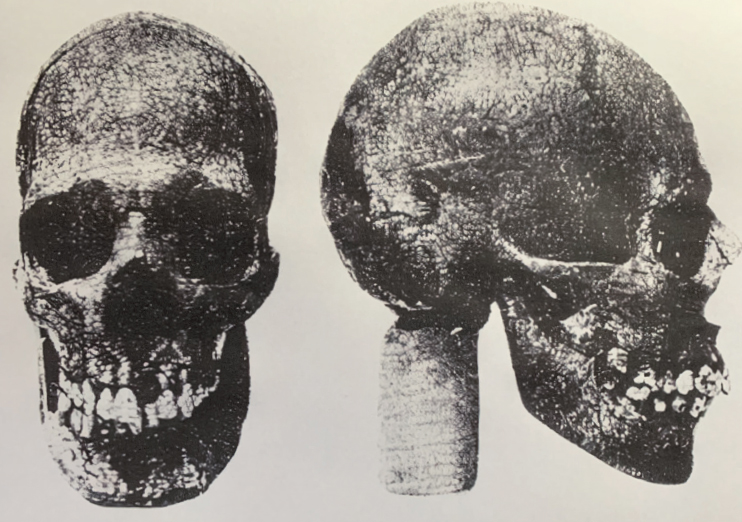
Microcephalic skull excavated at Kato Zakros, Crete, and dated to about 2000 years ago. Image reproduced courtesy of Dr AN Poulianos.7
‘It’s crystal clear—it’s a modern human, if one with many problems,’ claimed Dr Alan Thorne.
‘It was totally outside the range of modern human variation. There was no way it could be modern. Very, very clear-cut,’ said Professor Peter Brown.
Others exclaimed:
‘It just invites tremendous scepticism.’
‘I don’t believe it.’
‘There is only one skull and that’s not proof of anything at all.’
‘Entire careers are at stake.’
‘One group is going to be entirely wrong.’1
After news broke of the H. floresiensis discovery, it rapidly became clear that this species would be very controversial.
This would not have been wholly unexpected. In their announcement of the new species, Peter Brown and colleagues, having carefully examined all the H. floresiensis bones, came up with two possible explanations for them.2 One was that H. floresiensis was a dwarfed form of H. erectus. According to this scenario, it was assumed that H. erectus had somehow arrived on Flores from Java,3 and that an insufficient amount of food or other conditions on the island led to an evolutionary response to dwarf. Peter Brown and colleagues’ alternative idea was that H. floresiensis was a remnant population of some very early hominins, such as those that lived in what is now Africa a million or so years ago.
Science proceeds by researchers questioning and testing others’ hypotheses and conclusions; this is the normal and appropriate procedure. And so researchers would have begun asking questions such as ‘Did the team look at all possible explanations for the Liang Bua bones?’ or ‘Did they interpret their information correctly?’ or ‘Could the bones belong to Australopithecus or to some already known species of Homo?’
But then, suddenly, something came in from left field. On 31 October 2004, just three days after the announcement of H. floresiensis, an opinion piece appeared in Adelaide’s Sunday Mail newspaper. Referring to the discovery, but with a curious lack of supporting images, Professor Maciej Henneberg (University of Adelaide) claimed that: ‘Dimensions of the face, nose, and jaws were not significantly different from modern people … but the measurements of the brain fell a long way below the normal range.’ He added: ‘The bell rang in my head … the skull is of a microcephalic individual—there is not a single significant difference between Homo floresiensis and a microcephalic from ancient Crete.’4
Maciej Henneberg and colleagues were to expand on this, suggesting that LB1 had a brain size equal to the smallest of any human ancestor, while its teeth were similar to modern human teeth. They noted that LB1 had a normal-sized face but a brain size equal to early hominins from three to four million years ago, yet the species had the intelligence to make stone tools. Its body size was, ‘considering its dating, way off the scale’.5 They discounted the idea of island dwarfing for H. floresiensis, reasoning that had H. erectus voyaged to Flores once, it could have done so regularly, rather than become isolated and subsequently dwarfing. They also viewed Flores as not such a small island as to generate dwarfing as an evolutionary response to that environment. So island dwarfing of H. erectus, in their view, was unlikely to explain the existence of H. floresiensis.
Interestingly, Henneberg noted that many of the skeletal comparisons in Brown and colleagues’ descriptions of H. floresiensis were made with the australopithecines rather than with Homo. This led him to wonder whether the discovery indicated the presence of a hitherto unknown form of Australopithecus that had left Africa at least a million years ago and reached the Indonesian archipelago.6 Nevertheless, he and his colleagues did not test this thought-provoking idea.
Seeking an alternative explanation for the Liang Bua bones, Henneberg and Dr Alan Thorne proposed that the individual known as LB1 was a modern human with the medical condition microcephaly (literally ‘tiny brain’)—in essence, they were saying that this was not a new species at all. It was a contentious suggestion by well-known researchers.
We didn’t know it at the time, but the microcephalic modern human theory was the first of several arguments that would be put forward to oppose the acceptance of H. floresiensis as a new species. Over the next few years, we would read about the use of Laron syndrome, cretinism and Down syndrome to explain the characteristics of LB1, not to mention connections with small-statured modern human populations. The discovery of H. floresiensis had really started something.
While challenges to the proposed ancestry of H. floresiensis were anticipated, it was quite unexpected that the loudest assertions were about how H. floresiensis was not a new species at all but a modern human. Those making such assertions had to explain away the obvious differences between H. floresiensis and us, the solution being to claim that H. floresiensis—or at least the individual LB1—was a modern human with a pathological condition.
But back to 2004. With so much buzz about the new species, and with the annual conference of the Australasian Society for Human Biology (ASHB) coming up, Dr David Cameron (University of Sydney) and I thought there was an opportunity to engage with a wide audience on the differing ideas about H. floresiensis. The conference program had already been finalised but we were able to add an extra session to focus on this new little species. David and I would present our work, as well as co-chair the gathering, while Professor Henneberg and Dr Denise Donlon (University of Sydney) would also take part.
For our presentation, I compared data from the skull shapes of different early human species and a couple of microcephalic skulls of modern humans described in archaeological excavation reports. In a nutshell, the results showed that H. floresiensis clustered with the earliest species in our genus, H. habilis. And there was something else: it was notably separate from any modern humans, including the microcephalic skulls. It seemed that H. floresiensis (LBI) was most similar to early Homo skulls. Essentially, I laid out an evidence-based position that H. floresiensis was not a modern human with microcephaly.
My conviction became even stronger when I saw the images presented by Maciej Henneberg in his talk. Realistically, practically anyone looking at the images of the Minoan microcephalic skull and that of LB1 could point out the differences fairly easily. Some of these differences are what distinguish H. floresiensis from modern humans, so for clarity I’ll run through these.
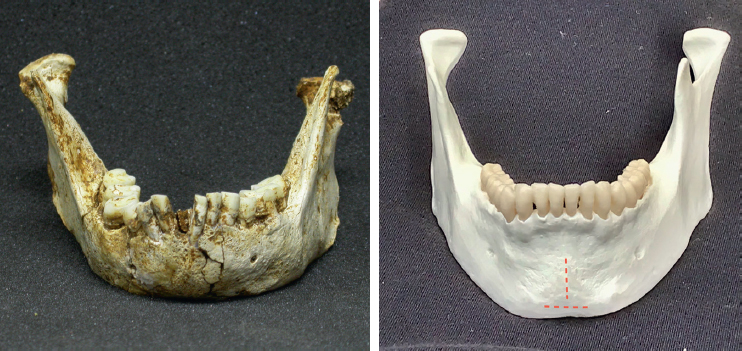
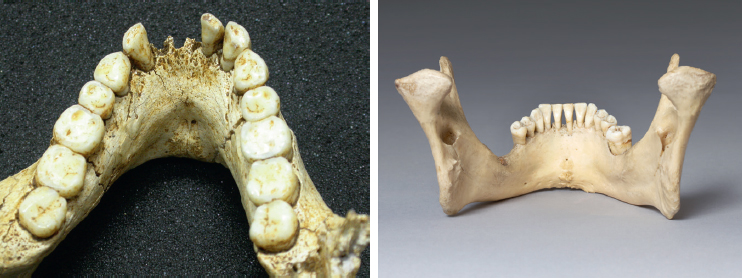
In comparing the two skulls, we can see that LB1’s forehead is low while the Minoan has the high, rounded forehead typical of modern humans. LB1 doesn’t have a chin; instead, the jaw slopes backwards. The Minoan, on the other hand, certainly has a chin. In side view, the LB1 face projects, while the Minoan’s is vertical, just like ours. Also note that LB1 has bone protruding around the upper rim of the eyes, and a column of bone extends from the eyeteeth up towards the edge of the nose—these two structures are not seen on the Minoan.
Frankly, sitting there at the ASHB conference, I was astounded. The two skulls were just so dissimilar; in fact, the only feature they had in common was a relatively small head in relation to the face. Yet Maciej’s presentation clearly had support from some people in the audience. Things got pretty lively as people tried to come to grips with the opposing views on H. floresiensis.
At one point, someone who was clearly frustrated or straight-up annoyed by the new species, called out, ‘When I was at school we all knew that in evolution one species evolved into another.’ In other words, their view was that human evolution follows a ‘straight line’ of ancestor and descendent species, with only a single human species existing at any one time, and so H. floresiensis could not have existed at the same time as H. sapiens. David Cameron jumped in to say that it was now generally agreed that human evolution is more like a tree, where multiple human species ‘branches’ can coexist at any point in time. As I recall, this was met by the comment that if Homo floresiensis was a new species, then we would have to unlearn everything we know about human evolution.
Microcephalic skull excavated at Kato Zakros, Crete, and dated to about 2000 years ago. Image reproduced courtesy of Dr AN Poulianos.7
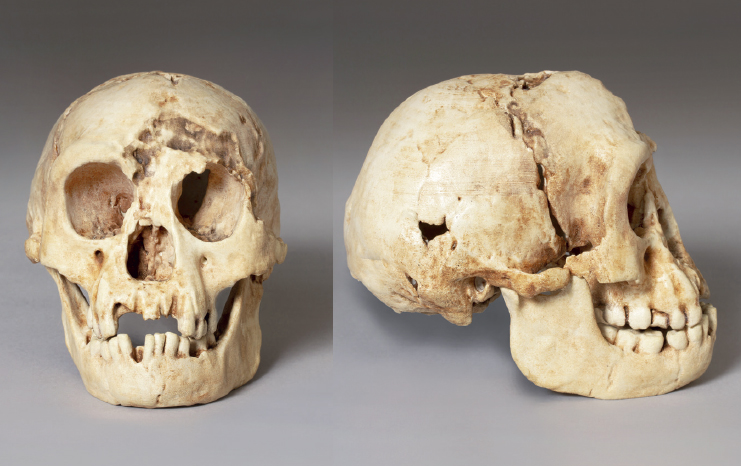
The skull of H. floresiensis (LB1; replica). Images provided by Maggie Otto.
This heated exchange revealed to me that underlying the microcephaly hypothesis was a gut reaction against H. floresiensis as a new species. The acceptance of H. floresiensis was a problem if people viewed human evolution as a straight line, because H. floresiensis lived at the same time as modern humans. The concept of a linear evolution developed after Charles Darwin, who proposed in his book The Descent of Man that we are descended from an unknown form of Old World primate.8 Ernst Haeckel, a German biologist, had come to the same conclusion. But there had to be some intermediary form between apes and us, and Haeckel even created a species name for this hypothetical being: Pithecanthropus. The ‘straight line’ idea of human evolution emerged as more primitive-looking hominin fossils were discovered and scientists began to think of these as evolving in a linear sequence from the most primitive hominins to modern humans.
Others saw some of these fossils as dead-ends on a more tree-like structure that nevertheless incorporated a dominant ‘trunk’ leading to modern humans.9 That is, A. afarensis was followed by A. africanus, which was followed by H. habilis, which evolved into H. erectus, and we evolved from that species. There were two acknowledged divergences from this straight line: the robust Paranthropus coexisted in Africa with the australopithecines and early Homo; and the famous Neanderthals, Homo neanderthalensis, which lived in Europe and the Middle East from at least 200 000 to 30 000 years ago, existed at the same time as earlier populations of H. sapiens.
Since the 1980s, though, more and more discoveries have indicated that, in human evolution, different species persisted alongside others, while some species remained unchanged for long periods (see the species timeline that appears prior to chapter 1). Just as for other mammals in the fossil record—elephants, pigs, antelopes and so on—human evolution now looks much more like a tree of many branches than a straight line.
At the end of our ASHB conference session, I met Professor Richard Wright, who said that he was impressed with my skull analyses. Richard has a prodigious reputation regarding that sort of data analysis, alongside an impressive list of academic achievements, so when he raised the idea of working together on a project about the microcephalic–early hominin debate, I was most enthusiastic. Over dinner that evening, Richard, Professor Colin Groves, Denise Donlon, David Cameron and I discussed the pros and cons of the idea. I had actually thought that any wider interest in this angle would fizzle out because it was so obvious that LB1 was not a modern human, but the others were much, much wiser than me. And Colin was already well ahead of the rest of us, brimming with ideas on how to get started. He asked Richard if he knew of any microcephalic skulls that we could examine. I assumed Colin would work on these, but to my surprise, he said, ‘Oh no, I thought you could.’
And so, as a group, we started planning the work ahead. We would test the microcephalic hypothesis and work out to which species H. floresiensis was most closely related. This meant assessing the microcephalic question and the two ideas originally proposed by Peter Brown and colleagues: that H. floresiensis was a remnant population of archaic two-million-year-old hominins from Africa, or that it was a dwarfed H. erectus.
We had an ambitious project ahead of us, but we were starting from a good position. This was partly because we had already laid some of the groundwork, and partly because of the skills and experience of our team members. Denise had studied up on the H. floresiensis arm, leg, knee and foot bones, as well as the pelvis on which Peter Brown and colleagues had just published. David had worked out the most likely place H. floresiensis fitted on the human evolutionary tree. I had found that the microcephalic modern skull clustered with that of normal modern humans, while LB1’s was most similar to early Homo skulls from Africa. And Colin was a powerhouse of knowledge on everything to do with evolution, taxonomy and hypothesis testing.
By early 2005, our team was hard at work. But within a few months, we had something else to add to the mix. Maciej Henneberg and Alan Thorne formally followed up Maciej’s earlier newspaper article and ASHB presentation by publishing in the online journal Before Farming.10 In their article, they argued that the LB1 skull and the Minoan microcephalic skull were both characterised by very small braincases, but their faces were ‘within three standard deviations of the normal human range’. This meant that both faces were within the range of modern human face sizes, albeit at its outer edge. They cited small braincases and normal-sized faces as characteristics of microcephaly.
Henneberg and Thorne also disputed Peter Brown and colleagues’ estimation of LB1’s stature of 1 metre. Instead, they estimated that LB1 would have been 151–162 centimetres tall ‘depending on method of reconstruction’, although they did not detail how they worked this out. They pointed out that a modern human skeleton of similar stature had been excavated from another cave on Flores, Liang Toge, and dated to 3500 years ago.11
More theories about the origins of LB1 were to come, but before delving into those, let’s take a closer look at the claims around microcephaly, particularly the impact this condition has on bones, which is the main evidence my team was dealing with. When I started reading up on microcephaly, I expected it to be a singular thing, with one set of characteristics that could be compared to the features of the H. floresiensis bones. But it wasn’t that clear-cut. For example, I found that it’s a very rare condition. It occurs in one in two million births in Scotland and one in 250 000 births in the Netherlands,12 while in Sweden the incidence is predicted to be between one in 25 000 and one in 50 000.13 Moreover, there are over 400 forms of microcephaly, not just one, as I had assumed. The condition is seen more commonly in regions with high rates of consanguinity—when close family members, such as cousins, have children—such as Turkey, Pakistan and some countries in the Middle East.14 All microcephalics have a small brain that might be accompanied by other abnormalities such as short arms in relation to legs, short stature or joint defects. There may be cognitive impairment, where a person has trouble concentrating, remembering, learning new things, or making decisions that affect everyday life. The condition can range from mild to severe.15
The microcephalic skull on which Henneberg and Thorne had based their idea was excavated by Professor N Platon in 1962 from a grave in Malakari cave on eastern Crete. The find was dated to around 2000 BCE (that is, 4000 years ago), from the Minoan period.16 Professor Poulianos, who reported the find, estimated that the skull belonged to a person of about twenty years of age.17 The brain size was 350 cubic centimetres. Professor Poulianos concluded that the skull was that of a microcephalic. Fortunately for us, he had also published the measurements of the skull and I could now incorporate these into my analyses.18
I should also say that the discovery of the microcephalic skull on Crete was an exceptional find. It is rare to uncover bones in archaeological excavations that show individuals who had microcephaly. My research uncovered only five cases. Apart from the Minoan, there was a microcephalic from excavations in Sano cave, Japan, which I had used in my presentation at the ASHB conference. There was also one from ancient Egypt; one from a prehistoric burial ground in Peru; and one, a four-year-old child, from a cave on the Spanish island of Mallorca. Only the Minoan and Sano cave individuals were published with skull measurements that were necessary for our kind of analysis.
In 2006, Professor Teuku Jacob (Gadjah Mada University, Yogyakarta) and colleagues, including Maciej Henneberg and Alan Thorne, presented another explanation for H. floresiensis. They proposed that LB1, as well as other individuals found in the Liang Bua excavations, comprised an earlier population of small-statured modern humans. LB1, they claimed, showed signs of developmental abnormality, including microcephaly. They believed they had found 140 features on the LB1 skull—combined with those on two Liang Bua jaws—that were within the range of what we see in modern human skulls and jaws. In their view, some characteristics, such as the form of the eye socket and features at the back of the skull of LB1, are found in modern-day populations in South-East Asia, and in particular some people on Flores; here, they mentioned the Rampasasa villagers living near Liang Bua cave.
To test their idea, we needed to include the Rampasasa villagers in our study, but we had no luck in finding any information about them (note the postscript to this chapter). The next best thing we could do, then, was to use skull measurements of small-statured populations from elsewhere in the region. The closest to Flores were the people who live on the Andaman Islands in the Bay of Bengal, to the north-west of Sumatra.
In terms of what we had for comparison, we had amassed skull measurements of the Minoan and Sano microcephalic individuals, modern humans, a small-stature modern human population, the Liang Toge individual (mentioned in Professor Jacob’s publication) and key early hominins. Now we had to find out how similar these were to LB1 in skull shape and thus obtain a better idea of the ‘family tree’ relationships between them.
Professor Richard Wright analysed the skull data. The results showed that the Minoan skull clustered with modern humans, albeit at the outer edge of the range; that is, its skull, although smaller than that of modern humans, was overall very similar to them. On the other hand, the shape of LB1’s skull was quite different from the Minoan skull and all modern human skulls in our study. In fact, LB1 was close in shape to two 1.7–1.8-millionyear-old hominin skulls from Africa: a H. habilis skull from Tanzania, and a skull from the Koobi Fora research area in Kenya whose species is still being debated.19 That said, we were acutely aware that analysing skulls was not the end of the story. Other skeletal characteristics were vital when considering the origin of H. floresiensis, one of which was the relative length of the arm to the length of the leg (body proportions). Dr Denise Donlon therefore compared the arm-to-leg proportions of A. afarensis, Australopithecus garhi, H. habilis, H. ergaster, H. floresiensis and modern humans. She found that H. floresiensis limb proportions were similar to 2.5-million-year-old species A. garhi and dissimilar to any other species in the analysis.20
These outcomes led us to propose two possible scenarios for H. floresiensis. It could have evolved from a previously unknown early hominin that shares skull similarities with the Koobi Fora hominins and limb proportions with A. garhi, even though H. floresiensis is much younger than these species and was discovered in Indonesia, half a world away from Africa. Alternatively, it might have been in the process of evolving from Australopithecus to Homo when it wandered out of Africa.21 These hypotheses were exciting because they potentially opened up a different way of thinking about what happened during human evolution, and they were also thought-provoking because they implied a movement of hominins out of Africa earlier than anyone had thought possible.
Meanwhile, Professor Dean Falk and colleagues were also testing the microcephalic idea for LB1 (as mentioned in chapter 1). Professor Falk is a specialist in the evolution of the brain, specifically how it led to the emergence of language, music, analytical thinking and even warfare; as part of her research, she has even studied Albert Einstein’s brain!22 Surprisingly but happily, imprints of LB1’s brain remained on the inside of the skull, and Falk and colleagues were therefore able to make a 3D model of the brain’s form, with all its lumps and bumps—these replications are called endocasts. When she and her team compared the LB1 endocast with that from a microcephalic, they found that the two were quite different. Not only did they differ significantly in shape, they had other dissimilarities. For example, where the lumps of the brain just behind the forehead (called frontal lobes) were wide and flat on LB1, the microcephalic’s frontal lobes were narrow and pointed. In fact, Dean and colleagues found nothing in LB1’s endocast to suggest microcephaly.23
Neither the work of Dean and her colleagues nor ours dampened the microcephalic idea, however. Professor Robert Martin (Field Museum of Natural History, Chicago) and colleagues, experts in how human bodies grow and change in their proportions, did their own investigation into LB1.24 They argued that if LB1was a dwarfed descendant of H. erectus, as initially proposed by Peter Brown and colleagues, then its brain would be much bigger than it was. Dismissing H. erectus as an ancestor of H. floresiensis, Martin and colleagues declared that LB1 had to be pathological. Developing this idea, they noted that LB1’s short height, small and receding jaw, and some dental characteristics overlapped with those of microcephalics. The most likely explanation, in their view, was that LB1 could well be a microcephalic modern human.
Oddly, Martin and colleagues did not address one of the original and potentially key explanations for H. floresiensis: that it could be an archaic hominin species. They excused themselves for not testing this hypothesis with the stated reason that ‘LB1 may derive from a more primitive (preerectus) population that cannot be addressed by consideration of modern human developmental abnormalities’.25 I did not know what this meant. For more than a century, palaeoanthropologists have managed to work out the species of the fossils they discover. This process is not new. But such a position also supported my earlier thoughts about rocking the boat: that H. floresiensis as a new species was way beyond the comfort zone of some researchers.
It wasn’t the last we heard of H. floresiensis being a modern human population with some sort of abnormality, or at least that LB1 was a modern human with an abnormality. Rather, 2006–07 turned out to be a pretty busy period regarding the H. floresiensis debate. New ideas and theories kept rolling in. Gary D Richards (University of California) suggested that the Liang Bua cave individuals were modern humans who developed a mutation in a particular gene, causing a growth hormone deficiency.26 The population therefore became dwarfed in that island environment. Meanwhile, Professor Israel Hershkovitz (Tel Aviv University) and colleagues, noting the extremely short stature of H. floresiensis and its small brain, looked at whether these individuals were modern humans who had a condition called Laron syndrome.27
I had never heard of Laron syndrome. When I did an online search, I found that Dr Zvi Laron and colleagues had identified the syndrome in 1966. By 2015, it was estimated there were at least 500 people with Laron syndrome worldwide, although it is likely many cases remain undiagnosed.28 The cases occur in the Middle East, mid and South Asia, and the Mediterranean, with the largest single group of affected individuals (about 100 people) living in southern Ecuador.29 The genetic disorder occurs in some children of members of closely related families. It affects growth, resulting in short stature, among other complications. If the condition is not treated, adult males typically reach a maximum height of between 116 and 142 centimetres, and females grow to between 108 and 136 centimetres.30 Keep in mind that LB1 is 106 centimetres tall, close to the smallest among Laron syndrome females.
I needed to know more and eventually tracked down a published photo of the head of a very young, unidentified child with Laron syndrome.31 Taking a close look, I saw that the child had the high rounded forehead of a modern human infant, quite different from LB1’s backwards-sloping forehead—the face did not project but was vertical like ours. This image of a child with Laron syndrome seemed very different from what I could see in LB1’s skull. Emeritus Professor Zvi Laron describes this protruding forehead as a characteristic of Laron syndrome. Sufferers also have short arms and legs, small hands and feet, defective teeth that break easily and have many caries (holes), and a head that is disproportionally wide compared to the base of the skull.32 H. floresiensis, however, has long arms but relatively short upper legs, very long feet, teeth which are in fairly good shape, and a skull that is widest towards its base. Laron syndrome was not shaping up well as a description of H. floresiensis.
Dean Falk and colleagues’ interest was piqued too, and they searched the medical and clinical literature for further information. Having scoured the relevant publications, they listed the characteristics of people with Laron syndrome: short stature, protruding forehead, saddle nose, short face, broken and discoloured deciduous teeth with irregular growth and crowding of the permanent teeth, delicate arms and legs, small head circumference, and short limbs relative to trunk.33 They, too, determined that the facial characteristics of Laron syndrome were quite at odds with those of LB1.34
Still, the ‘pathology hypotheses’ kept on coming. Professor Bill Jungers would wryly observe, ‘Another day, another pathology.’
Cretinism was the condition that Peter Obendorf (Royal Melbourne Institute of Technology) and Emeritus Professor Charles Oxnard (University of Western Australia) proposed to explain H. floresiensis. Their view was that the Liang Bua group was part of a long-term modern human population that suffered from an iodine deficiency resulting in thyroid and growth problems.35 They decided that the characteristic hips, arms, legs, shoulder bones and foot size of those suffering from cretinism were in the main similar to the features of the two H. floresiensis individuals, LB1 and LB6. Obendorf and Oxnard were very much aware that LB1’s brain size was tiny (426 cubic centimetres), much smaller than that of any modern human. By suggesting that cretins of small-statured South-East Asian populations could have brain sizes of between 400 and 500 cubic centimetres, it seemed as if they were proposing that LB1 and LB6 were cretins from an unidentified, modern human, small-statured population.
Colin Groves was particularly curious about the claim that H. floresiensis individuals were cretins. While in Europe to undertake research, he took the opportunity to examine and measure ten cretin skulls and seven cretin skeletons at the Museum of Natural History in Basel. Along with Catherine Fitzgerald (ANU), he presented his findings at the 2009 ASHB annual conference, informing the audience that cretins’ leg bones are far longer than those of H. floresiensis, that cretins’ upper arms and feet are also quite different from those of H. floresiensis, that cretins do not have the small brain of LB1, and that cretins’ teeth are in very poor condition (or missing) in comparison to the teeth of H. floresiensis. In total, Colin displayed nine PowerPoint slides documenting the skeletal differences between cretins and H. floresiensis. He also found that Obendorf and Oxnard had misinterpreted some of the skull characteristics.36
The skull of a cretin differs to that of LB1 in having a high forehead, no mounds of bone above the eyes or the valley-like structure behind these, and the face does not project from under the eyes as LB1’s does, although it projects just below the nose. Also, there is a chin, and the jaw is small in relation to the face.
Peter Brown roundly and comprehensively rejected cretinism as an explanation for LB1 and LB6. From the medical literature, he compiled the characteristics of cretinism across the whole body, forty-nine in all. Of these, thirty-three would have shown up on the bones we have for the two Liang Bua individuals, but not one of those cretinism characteristics was present. A quick look down his list said it all, really—the most prevalent word was ‘No’.37
Karen Baab and colleagues then took up the mantle. I’d met Karen when we were working with other volunteers at the early hominin site of Dmanisi. Karen and colleagues used a computerised 3D skull-shape-analysis technique to test whether cretinism or Laron syndrome could explain the form of LB1’s skull, or whether its skull was more similar to extinct species of Homo.38 They found that LB1’s skull shape was outside the range of variation in cretinism and was very different from those of humans with Laron syndrome. Instead, it was most similar to some fossil hominins.
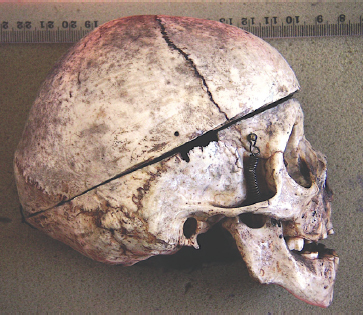
Skull of a person with cretinism. The cretin’s skull would have been cut at some time for demonstration or research purposes. Image provided by D Bulbeck and M Oxenham.39

LB1 skull (replica). Image taken by Maggie Otto. (Photos not to scale.)
The issue of whether H. floresiensis was a modern human with pathology or a new hominin species continued to engage researchers’ interest. It fuelled debate at international conferences in the United States, Indonesia and Europe, with multiple speakers presenting each view. At the same time, we were also getting to know more about the strange mix of features in H. floresiensis. As I related in chapter 1, at first we were confronted with a human-like creature that had some ape-like, australopithecine, and even early and later Homo characteristics. Now we were finding out that this little being had hunched shoulders and that its arms faced slightly forward compared to ours. Its feet were extremely long in proportion to its legs, and they were flat where ours are arched. There were also some odd curvatures in some of the toes. Ape-like wrist bones completed the picture.
Equipped with this clearer picture of what H. floresiensis would have looked like, and in light of our research, along with that of Dean Falk and Karen Baab and their colleagues, I entertained the idea that the ‘pathology’ papers would cease. But this was a short-lived thought. Microcephaly continued to be advocated as an explanation for H. floresiensis. RC Vannucci and colleagues, for example, performed some analyses on the skulls and endocasts of microcephalic individuals, comparing them to those of LB1. They concluded that LB1 was ‘a brain-damaged individual, who suffered secondary microcephaly with associated severe motor disability (cerebral palsy) and likely mental deficiency’.40
Down syndrome was the next—and final—condition used in an attempt to explain LB1 as something other than a new hominin species. In 2014, Maciej Henneberg and colleagues proposed that LB1 was a small modern human with Down syndrome.41 I decided to investigate this assertion, aiming to present my findings at the 2014 Australian Archaeological Association annual conference. I first looked up the skeletal characteristics of people with Down syndrome in the Cambridge Encyclopedia of Paleopathology. Then I listed the characteristics of LB1 and set about ticking any that also appeared in people with Down syndrome—nothing on that list got a tick. I also did the opposite, listing all the characteristics of people with Down syndrome and looking to tick those that also appeared in LB1, but again there were none.
Karen Baab and colleagues were also on the case. Examining the clinical literature on Down syndrome, they found that people with this condition have skulls often described as somewhat flattened at the back and base. The midface is hypoplastic, which means that the cheekbones and eye sockets have not grown as much as the rest of the face. The jaw is normal in size, but some individuals might have an unusually small chin. Down syndrome individuals also tend to have a short stature and a higher foot-to-thigh ratio than those who do not have the syndrome.42
Down syndrome skulls are of course modern in form, while LB1’s is not. LB1’s short stature of just over 1 metre and its disproportionally short lower limbs are primitive features also not seen in modern humans. All in all, Baab and colleagues found no support for LB1 being interpreted as a modern human with Down syndrome. They considered that LB1, and the other bones assigned to H. floresiensis, were better interpreted as a distinct species whose affinities lay with early Homo.
Dr Michael Westaway and colleagues also responded to the claim that H. floresiensis had Down syndrome.43 They focused on jaw differences, and especially on one of the key things Peter Brown and colleagues had pointed out. It has long been established that early hominin species lacked chins. This is just one of several characteristics that show H. floresiensis is an archaic species, not a modern human.
I want to digress just for a moment on the subject of the chin, particularly as Michael Westaway and colleagues used the non-chinned jaws of H. floresiensis in their analysis. By ‘chin’, we are not just referring to the shape of the front of the jaw. Colin Groves alerted his students and conference audiences to an important feature of modern human jaws that had been described by Jeffrey Schwartz and Ian Tattersall.44 This distinction is critical to the question of whether H. floresiensis is a modern human. The centre of the front of the modern human jaw is slightly raised, in the form of an upside-down ‘T’; this bone reinforces the jaw. This specialised form of the jaw develops well before birth and is unaffected by the type of food that might be eaten or how vigorously we chew. But the Liang Bua jaws do not have this formation. We do not see this on H. erectus jaws or on other archaic hominin jaws.
Left, the front of the H. floresiensis jaw, showing its smooth surface; and right, a modern human jaw (replica) showing the upside-down ‘T’ bone formation extending from under the incisors and across the base of the jaw. Images provided by Mike Morwood (LB1 jaw) and Debbie Argue (modern human jaw). (Photos not to scale.)
The reinforcement on the H. floresiensis jaws, however, is on the inside. Immediately below the front teeth is a ledge that slopes outwards and downwards. Below this is a bit of a depression before there is another ledge which stretches across the base of the jaw. Together, these structures reinforce the H. floresiensis jaw. Modern humans do not have these formations—the inside of our jaw descends straight down from behind our front teeth.
Based on the form of the H. floresiensis jaws, Michael Westaway and colleagues determined that neither LB1 nor LB6 could have been modern individuals, let alone have Down syndrome.
Even more evidence was to emerge that showed H. floresiensis was not a modern human with pathology. Antoine Balzeau (Muséum National d’Histoire, Paris) and Phillippe Charlier (Paris Descartes University) studied a computerised tomography (CT) scan of the skull of H. floresiensis.45 As most people know, CT scans can show hidden features inside bones that can’t otherwise be seen, even on close inspection. Scanning the skull of LB1, Balzeau and Charlier were able to see a variation in thickness across it, as well as the structure inside the bone. They also scanned thirteen microcephalic skulls from a number of museums, and some normal modern human skulls. Furthermore, from previous work they already had scans of twenty-six early hominin skulls, including A. africanus, H. habilis, H. erectus from Java, and some Neanderthals. This was an impressive range of CT scans with which to test whether the LB1 skull was a microcephalic or if it fitted better with any of the skulls of the australopithecines or early Homo.
Left, inside view of a H. floresiensis jaw; and right, a modern human jaw (replica). The H floresiensis jaw is much smaller than modern human jaws. Images provided by Mike Morwood (LB1 jaw) and Debbie Argue (modern human jaw).
Balzeau and Charlier discovered much about LB1’s skull structure. Most of this information was fairly complex, but the most relevant thing regarding our story was that all of the microcephalic skulls shared the internal skull bone conditions of modern humans, while LB1 did not. Balzeau and Charlier concluded that LB1 was not a modern human, with or without microcephaly. Disappointingly, however, they also found that the features they observed were not helpful for working out just which species H. floresiensis was closely related to.
So many hypotheses and so much controversy about this small being. Where does this leave us in the ‘pathology’ debate?
As I pointed out earlier in this chapter, it is normal procedure to test new scientific hypotheses. Most people would know that science works by examining all the facts concerning the question that is being investigated, and then determining which hypothesis best explains that information. Other researchers may then choose to test the hypothesis using additional data. Scientific progress is achieved in this way—nothing new here.
The pathology hypotheses generally focused on just the one set of H. floresiensis remains—LB1—with the implicit assumption that this was an abnormal individual in an otherwise normal population. But LB1 was not the odd person out here. The skeletal parts of the other individuals from the Liang Bua excavations represented the same (or even smaller) statured hominins as LB1. The (less complete) material we have of other individuals is all very similar to LB1.
Now, as we have seen, microcephaly and Laron syndrome are rare conditions. It would be reasonable to expect that, even if an archaeological excavation did reveal such a rare skeleton showing evidence of these syndromes, most of the other skeletal remains would be of a normal, non-pathological modern human population. But no bones of modern human stature or morphology have been recovered from the H. floresiensis levels in the excavations. Those who proposed these sorts of pathology for LB1 did not explain the absence of modern-looking, typical-statured human skeletal material in the deposit.
Finally, the proponents of the ‘modern humans with pathology’ explanation for the H. floresiensis population seemed not to realise something very significant about their proposals. At the time of this debate, the earliest evidence for modern humans in the region was 60 000 years ago, in Australia.46 And yet, evidence for H. floresiensis went back at least 95 000 years. The hypotheses which implied that the entire H. floresiensis population consisted of modern humans, with or without pathology, unconsciously pushed back the date of modern humans’ arrival in the region by 30 000 years. That would have been headline news indeed. But somehow, either the ‘pathology’ advocates did not realise this amazing implication of their work or they chose to ignore it, as it would have required a lot of explaining.
It is also worth pointing out that, since 2014, no new pathology hypotheses for H. floresiensis have been presented.
Time for some reflection. Clearly, the views on H. floresiensis were widely divided. This happens regularly in science, and it is expected that competing hypotheses will be tested. What was surprising was how strongly some researchers held onto the view that H. floresiensis was a modern human, even as the evidence for H. floresiensis being a new species became increasingly solid.
Professor Michael Lee (Flinders University; South Australia Museum), whom we meet again in chapter 3, shared the following with me on the polarisation of people’s views in the H. floresiensis debate:
It can happen in science. How the debate about birds and dinosaurs played out is similar to what happened in the case for H. floresiensis. Before the 1990s there was strong but not overwhelming evidence that birds descended from dinosaurs. A small but vociferous bunch of people argued that birds did not descend from dinosaurs, that they derived from much more primitive crocodile-like reptiles that date back to 240 million years ago. But from the 1990s, all these feathered dinosaur fossils from China appeared, which absolutely, convincingly demonstrated that dinosaurs gave rise to birds. Anyone coming into this research field after the 1990s has no problem accepting that birds descended from dinosaurs. But some people made up their mind very early in the debate when the evidence wasn’t conclusive. Once the evidence about the feathered fossil dinosaurs came in, these people still stuck to that initial opinion. They tried to reconcile the new evidence with their previous beliefs, rather than evaluating it objectively … [and they did this] in one of two ways.
The first was that the newly discovered feathered fossils were true dinosaurs that had nothing to do with birds. These dinosaurs evolved feathers separate to birds evolving feathers. The other explanation touted was that these feathered dinosaurs were true birds and had nothing to do with dinosaurs. So they were birds that had evolved all these dinosaur traits. Neither explanation is very plausible. So I think what happens is that people in general, not just scientists, can form very strong opinions early on in the debate when the evidence is ambiguous, and when new evidence comes in they are very reluctant to shift their view. They try to mould and shape new evidence to fit their long-held beliefs, rather than the other way round …
I think that’s what happened with H. floresiensis. Before all the detailed anatomical evidence came to light, the idea that the bones were actually some sort of pathological modern human probably couldn’t be completely discounted; a few people were attracted to that idea, making up their minds early on. When more and more evidence showed that H. floresiensis was a primitive species, those people had to try and shoehorn that evidence to fit the pathology idea and their explanations got more and more implausible … and none of them fitted very well. Moulding the evidence to fit your ingrained ideas is something scientists should always be guarding against.47
Fortunately, while the pathology debates were in full swing, other researchers were testing the ideas presented by Brown and colleagues when they first announced H. floresiensis. We will now take a closer look at these ideas.

As mentioned earlier in this chapter, my colleagues and I were unable to find data for the Rampasasa villagers whom Professor Teuku Jacob and colleagues suggested were descended from H. floresiensis. This was a problem that beset a number of researchers who wanted to test this hypothesis. However, late in 2018, all was revealed—not through analyses of skull measurements, nor by looking at the bones of this group, but by that marvel of modern science known as deoxyribonucleic acid, or DNA. Serena Tucci (Princeton University) and colleagues sampled the DNA of thirty-two adult individuals from Rampasasa village, searching for gene sequences that did not match known DNA from modern humans.48 Unknown DNA might have suggested that an unknown hominin could have contributed genes to the Rampasasa villagers’ ancestors, but no such DNA was found. The Rampasasa smallstatured group, Tucci and colleagues concluded, was not related to any unknown early hominin. Therefore, these villagers could not be descended from H. floresiensis (or any other hominin), and H. floresiensis, then, was not the ancestor of the Rampasasa village population, as proposed by Jacob and colleagues. This knowledge frees us to move on to the two supported hypotheses for the origin of H. floresiensis that are still being debated.