4
The Spinal Cord
INTRODUCTION
The spinal cord is essential to the conscious and unconscious control of the body. It transmits and manages the flow of information continuously between the brain and the rest of the body, and facilitates homeostasis in response to changes within and outside the body. The significance the spinal cord is such that damage due to trauma or disease can result in catastrophic losses of function. For example, traumatic damage to the cervical spinal cord can result in the loss of voluntary movement and sensation in all limbs and the trunk; impaired respiratory function, sometimes requiring permanent mechanical ventilation; loss of normal bowel, bladder and sexual function and control; and the disruption of autonomic homeostatic mechanisms resulting in hypotension, spasticity and the risk of autonomic dysreflexia.
Knowledge of the complex anatomy and physiology of the spinal cord enables the health care professional to understand the outcomes of insults to this important part of the central nervous system, and to plan management and rehabilitation effectively.
GROSS ANATOMY
The spinal cord is a cylindrical structure forming a major part of the central nervous system. It is flattened posteriorly and anteriorly, and divided incompletely into right and left sides longitudinally by the anterior median fissure and posterior median sulcus (Figure 4.1).
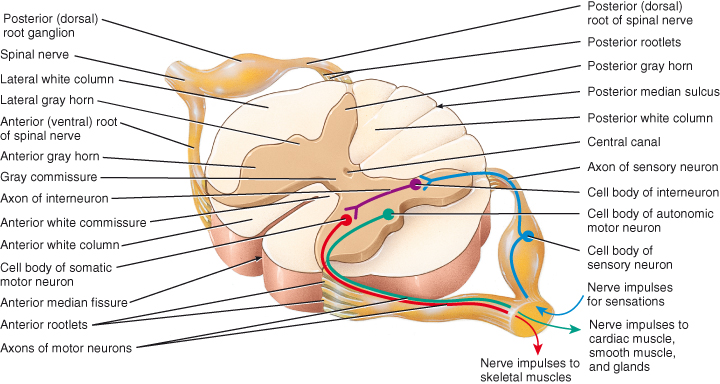
Figure 4.1 Internal anatomy of the spinal cord: the organisation of grey matter and white matter.
Reproduced from Principles of Anatomy and Physiology 12e by Gerard Tortora and Bryan Derrickson. Copyright (2009, John Wiley & Sons). Reprinted with permission of John Wiley & Sons Inc.
The cord begins as a continuation of the medulla oblongata and extends through the foramen magnum of the occipital bone usually to the upper border of the second lumbar vertebra, though it may terminate between the twelfth thoracic vertebra and the third lumbar vertebra (Figure 4.2). In adults it measures approximately 2.5 cm in circumference, and 42–45 cm in length. It lies within the vertebral column and is protected by it and its associated ligaments and muscles, and by the meninges and cerebrospinal fluid.
Figure 4.2 External anatomy of the spinal cord and the spinal nerves. Posterior view of entire spinal cord and portions of spinal nerves.
Reproduced from Principles of Anatomy and Physiology 12e by Gerard Tortora and Bryan Derrickson. Copyright (2009, John Wiley & Sons). Reprinted with permission of John Wiley & Sons Inc.
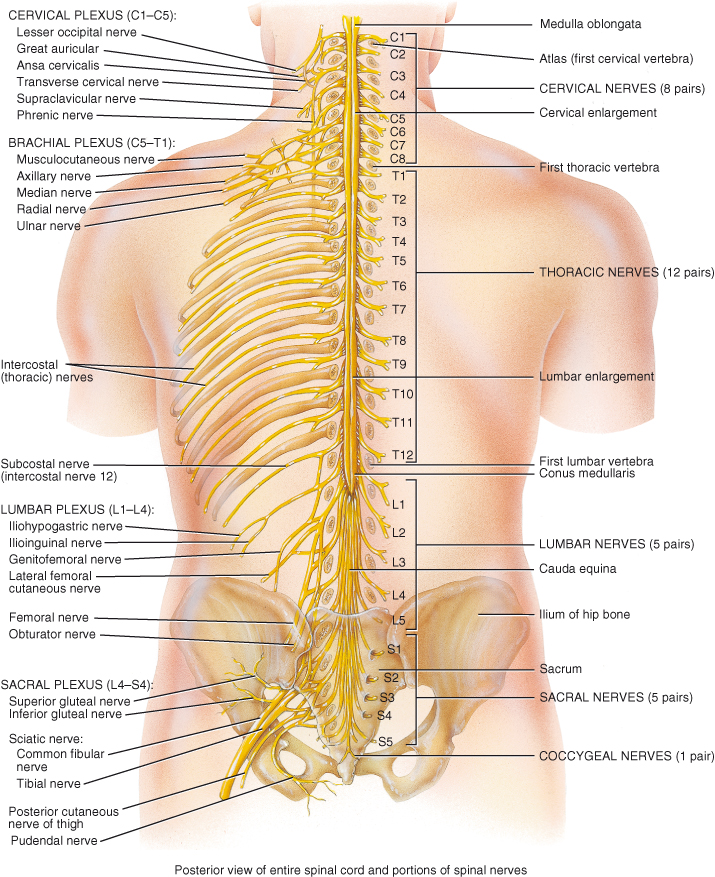
The cord has two enlargements: the cervical enlargement which extends from the fourth cervical to first thoracic vertebra from which arise nerves supplying the arms (brachial plexus), and the lumbar enlargement extending from ninth to twelfth thoracic vertebrae (spinal segments L1–S3) from which arise nerves supplying the legs (lumbosacral plexus).
Below the lumbar enlargement the cord tapers and becomes conical; this area is called the conus medullaris. The cord is anchored to the base of the vertebral column by the filum terminale, a continuation of the pia mater composed of non-nervous tissue which extends from the conus medullaris and attaches to the coccyx.
The spinal cord does not grow to the same length as the vertebral column; it fills only the upper two thirds of the vertebral space. As a result spinal nerves from the lower parts of the cord travel downwards within the vertebral space before exiting. The nerves of the lumbar and sacral area resemble a horse’s tail as they travel down through the vertebral space and are known as the cauda equina (Figure 4.2).
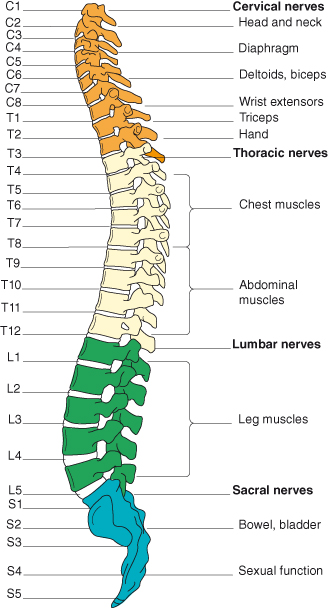
Nerves leave and enter the spinal cord in 31 pairs (Figure 4.2), one pair for each segment of the cord. The anterior or ventral roots of each nerve carry motor fibres away from the cord. The posterior or dorsal roots carry sensory fibres into the cord. There are 8 cervical segments and nerve pairs, 12 thoracic, 5 lumbar, 5 sacral and 1 coccygeal. The term ‘segment’ defines the area of cord from which a pair of nerves arises. However there is no structural segmentation within the cord; it is continuous throughout its length and its structure changes only gradually. The spinal nerves are named and numbered according to where they emerge from the vertebral column. Cervical nerves 1–7 emerge above their respective vertebrae. Cervical 8 emerges between the seventh cervical and first thoracic vertebrae. The remaining nerves are named for the vertebra below which they emerge. Each muscle or group of muscles innervated from a spinal segment is called a myotome (see Figure 4.3).
Figure 4.3 The spinal nerves and their areas of innervation. Reproduced from Peate I, Nair M, eds (2011) Fundamentals of Anatomy and Physiology for Student Nurses. Oxford: Blackwell Publishing Ltd., with permission.
Structure
The meninges which cover and protect the spinal cord, and the spinal blood supply are described in Chapter 2.
The spinal cord consists of grey and white matter (Figure 4.1). White matter is composed of sensory, motor and interneurone fibres and surrounds the grey matter. The grey matter forms a central H or butterfly shaped mass and is composed of the cell bodies of sensory neurones, lower motor neurones and interneurones, unmyelinated fibres and dendrites of interneurones and motor neurones lying at right angles to the long axis of the cord. The grey matter is rich in blood vessels and glial cells and is pierced by a central canal. This small channel extends from the fourth ventricle of the brain through the length of the cord and contains cerebrospinal fluid.
The grey matter
The grey matter is divided into regions called horns, linked together by the grey commisure. The horns of grey matter are named for their location:
- Anterior (ventral) grey horns – contain cell bodies of somatic motor nerves which carry impulses to skeletal muscles.
- Posterior (dorsal) grey horns – contain the cell bodies of somatic and autonomic sensory neurones.
- Lateral grey horns –contain cell bodies of autonomic motor neurones that regulate activity of smooth muscle, cardiac muscle and glands.
The anterior and posterior horns run the entire length of the cord. The small lateral horns project from the cross bar portion of the H from the second thoracic segment to the first lumbar segment only. All neurones with their cell bodies lying in the grey matter are multipolar.
The anterior horns contain some interneurones but are mostly composed of the cell bodies of somatic motor neurones. The axons of these cells leave the spinal cord via the anterior roots to travel to their effector organs, the skeletal muscles. The varying size of the ventral horns at different levels of the spinal cord reflects the varying amounts of grey matter present; the cervical and lumbar enlargements are due to the greater number of motor axons needed to innervate the upper and lower limbs.
The lateral horn neurones are sympathetic autonomic motor neurones serving the visceral organs. Their axons leave the cord via the ventral roots. The ventral roots carry both somatic and autonomic motor neurones.
The dorsal horns are composed entirely of afferent interneurone fibres from peripheral visceral and somatic sensory receptors whose cell bodies are found in the dorsal root or spinal ganglion. After entering the spinal cord via the dorsal root these axons may follow a number of paths. Some immediately enter the posterior white matter of the cord and travel upwards to synapse at a higher cord level or in the brain. Others synapse with interneurones in the dorsal horn grey matter at their entry level.
Within the grey matter neurone cell bodies form functional groups called nuclei. Motor nuclei provide output to effector tissues via motor neurones. Sensory nuclei receive input from sensory receptors via sensory neurones.
White matter
The white matter is composed of myelinated and unmyelinated nerve fibres and is organised as three columns on each side of the spinal cord:
- The ventral column lies between the ventral grey horn and the anterior median fissure.
- The dorsal column lies between the dorsal grey horn and posterior median sulcus.
- The lateral column lies between the anterior and posterior columns.
The anterior white commissure lies in front of grey commissure, connecting the white matter on left and right sides of cord. Each column is divided into tracts. A tract, or funiculus, is a distinct bundles of nerve fibres with a common origin or destination carrying the same type of information (Figure 4.4). The fibres within each tract are uniform in their diameter, myelination and conduction speeds. Sensory ascending and motor descending tracts are continuous with motor and sensory tracts in the brain. Tracts are named for their location, origin and destination, and direction of transmission, e.g. the spinothalamic tract carrying impulses from the spinal cord to the thalamus, or the corticospinal tract carrying impulses from the cortex to the spinal cord.
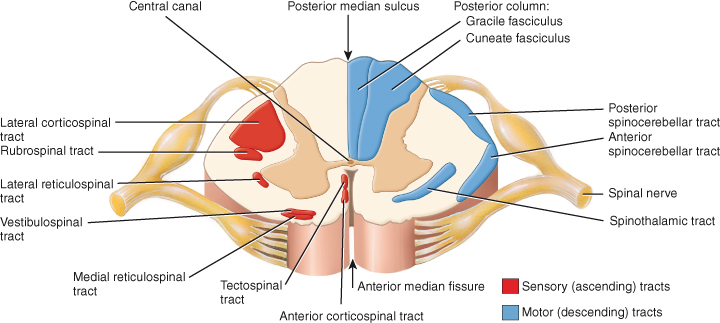
Figure 4.4 Locations of major sensory and motor tracts, shown in a transverse section of the spinal cord. Sensory tracts are indicated on one half, and motor tracts on the other half of the cord, but actually all tracts are present on both sides.
Reproduced from Principles of Anatomy and Physiology 12e by Gerard Tortora and Bryan Derrickson. Copyright (2009, John Wiley & Sons). Reprinted with permission of John Wiley & Sons Inc.
Most tracts will cross from one side of the central nervous system to the other at some point in their journey; this may occur in the brain or the spinal cord. Most tracts include two or three neurones along their length. All tracts are paired, with one half of the pair on each side of the spinal cord or brain. The centres and tracts that link the brain and the rest of the body are called pathways.
Sensory ascending pathways carry impulses from the periphery to the brain. There are three main ascending pathways:
1. The nonspecific or anterolateral pathway carries impulses from many different sensory receptors to the brain stem. It is formed mostly of the lateral and anterior spinothalamic tracts. Decussation occurs in the spinal cord (Figure 4.5). It transmits temperature, pain and coarse touch which are not clearly localised.
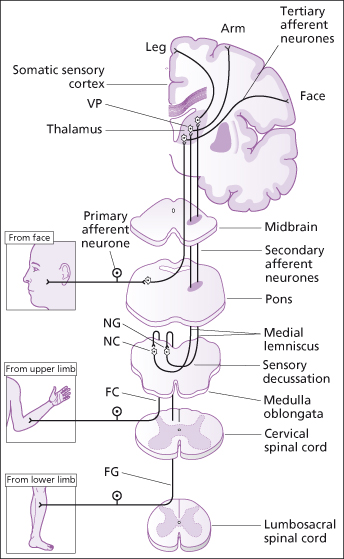
2. The specific ascending pathway or the medial lemniscal system carries precise transmission of impulses from a single type of sensory receptor or a small number of related sensory receptors which can be precisely located on the body surface i.e. discriminative touch, vibration and proprioception. It is formed mostly by the dorsal column (fasciculus cuneatus and the fasciculus gracilis) in the spinal cord and the medial lemniscus tracts which arise in the medulla and terminate in the thalamus, from where impulses are forwarded to the somatosensory cortex (Figure 4.6).
3. Anterior and posterior spinocerebellar tracts transmit impulses from muscle and tendon stretch to the cerebellum, where they are used to coordinate skeletal muscle activity. The pathways either do not decussate or do so twice so that information arises from and terminates on the same side.
Figure 4.5 Sensation. The spinothalamic pathway. ASTT, anterior spinothalamic tract; LSTT, lateral spinothalamic tract; VP, ventral posterior nucleus of thalamus.
Reproduced from C. Clarke et al., Neurology: A Queen Square Textbook, Wiley-Blackwell, with permission.
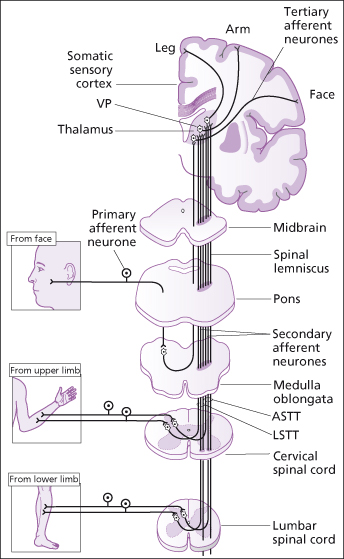
Figure 4.6 Sensation. The posterior column → medial lemniscal pathway. FC – fasciculus cuneatus; FG – fasciculus gracilis; NC – nucleus cuneatus; NG – nucleus gracilis; VP – ventral posterior lateral and ventral posterior medial nuclei of thalamus.
Reproduced from C. Clarke et al., Neurology: A Queen Square Textbook, Wiley-Blackwell, with permission.
Motor descending neurones carry impulses from the brain to peripheral effectors. There are two major descending pathways, the direct system and the indirect system:
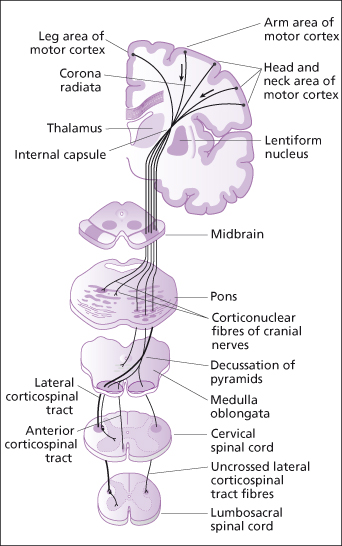
- Direct. The pyramidal or corticospinal tract, arising mainly in the pyramidal neurones of the precentral gyrus, carries impulses through the brain without synapsing until they reach the spinal cord (Figure 4.7). Here they synapse with interneurones or ventral horn motor neurones. This direct pathway carries impulses for fast or skilled movement.
- Indirect. The extrapyramidal tract includes all pathways outside the pyramidal tract (see Chapter 3). The term extrapyramidal has been superceded as it is now known that pyramidal tract neurones project into and influence the activity of much of the extrapyramidal tract. The complex, multisynaptic pathway is now referred to as the indirect or multi-neuronal pathway, or by the individual tracts of which it is comprised. The reticulospinal and vestibulospinal tracts maintain balance by varying the tone of postural muscles. The rubrospinal tracts controls flexor muscles and the tectospinal tracts mediate head and eye movements in response to visual and auditory stimuli. Table 4.1 provides more detail of each of the motor tracts.
Table 4.1 Descending motor tracts of the spinal cord.
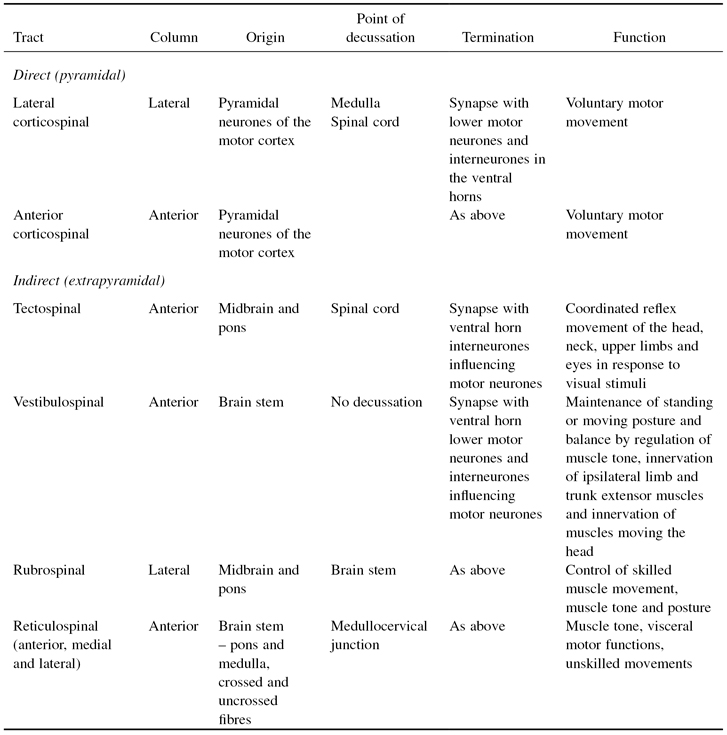
Figure 4.7 Corticospinal (pyramidal) motor pathways.
Reproduced from C. Clarke et al., Neurology: A Queen Square Textbook, Wiley-Blackwell, with permission.
PHYSIOLOGY
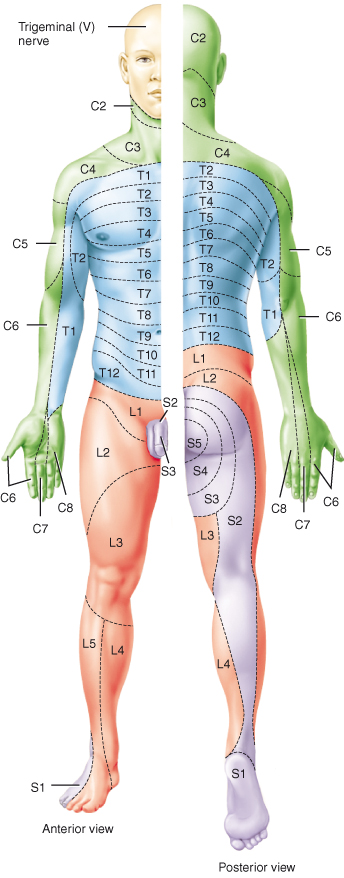
Unipolar sensory nerves enter the spinal cord through the posterior roots; their cell bodies form the posterior root ganglia. Sensory nerve tracts in the posterior horns carry afferent, ascending impulses generated by cutaneous receptors in the skin in response to pain, temperature, light touch and pressure, to the brain. A series of three neurones carry these impulses to the opposite cerebral hemisphere, where the sensation is perceived (Figure 4.5). Pain and temperature sensations travel to the cerebral hemispheres within the lateral spinothalamic tracts and cross over or decussate on entry to the cord. The anterior spinothalamic pathways carry information about pressure/coarse touch and itching and also decussate within a few segments of their entry to the spinal cord. The area of skin monitored for sensory information by a specific pair of spinal nerves is called a dermatome (Figure 4.8).
Figure 4.8 Distribution of dermatomes.
Reproduced from Principles of Anatomy and Physiology 12e by Gerard Tortora and Bryan Derrickson. Copyright (2009, John Wiley & Sons). Reprinted with permission of John Wiley & Sons Inc.
Sensory nerve tracts also carry vibration and light touch (discriminative, e.g. stereognosis) information from the skin, and ‘conscious proprioception’ from proprioceptors in tendons, joints and muscles. Proprioceptors provide information about balance and posture and the position of the body in space (see Chapter 11). It is referred to as conscious proprioception as this is information about position of which we are aware. These impulses may be carried by a three neurone pathway to the opposite cerebral hemisphere or by a two neurone pathway to the cerebral hemisphere on the same side. The impulses ascend within the dorsal or posterior columns (fasciculus cuneatus and the fasciculus gracilis) and decussate at the medulla (see Figure 4.6).
Motor nerve tracts carry efferent, descending impulses from the brain to the periphery resulting in contraction of voluntary, involuntary and cardiac muscle, and secretion by glands controlled by the autonomic system. Motor pathways use two neurones, upper motor neurones and lower motor neurones. The cell bodies of upper motor neurones supplying voluntary muscle lie in the motor cortex. The nerve fibres decussate in the medulla oblongata and, in the spinal cord, form the lateral corticospinal or pyramidal tracts (Figure 4.7).
The cell bodies of upper motor neurones supplying involuntary muscle lie in the mid-brain, brain stem, cerebellum or spinal cord. They form the extrapyramidal rubro-, reticulo-, tecto- and vestibulospinal tracts, some of which decussate while others do not. These tracts influence muscle activity that maintains posture and balance, coordinates voluntary muscle movement and maintains muscle tone. Table 4.1 provides a summary of the main motor spinal tracts.
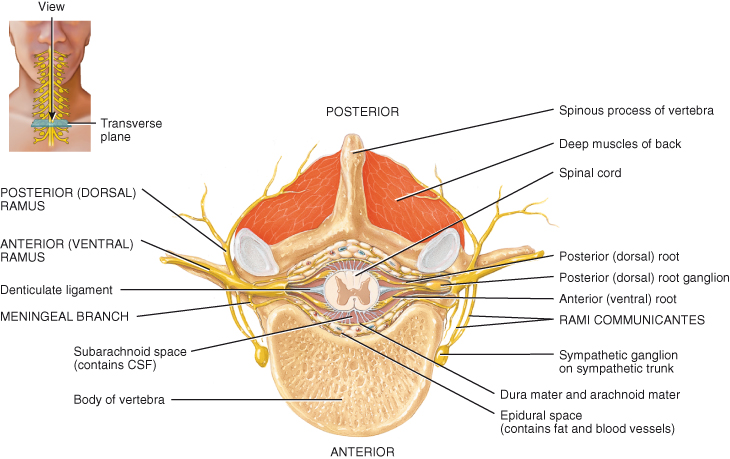
All upper motor fibres synapse with lower motor neurones in the anterior horns of grey matter. The lower motor nerve fibre leaves the spinal cord in the anterior root, joining with afferent sensory fibres to form a spinal nerve (Figure 4.9) which passes through an intervertebral foramen. The cell bodies of the lower motor neurones are stimulated and inhibited by various upper motor neurones and interneurones within the spinal cord which work together to produce smooth coordinated movement. The lower motor neurone forms the final common pathway for conduction of impulses to voluntary, skeletal muscle.
Figure 4.9 Branches of a typical spinal nerve, shown in transverse section through the thoracic portion of the spinal cord.
Reproduced from Principles of Anatomy and Physiology 12e by Gerard Tortora and Bryan Derrickson. Copyright (2009, John Wiley & Sons). Reprinted with permission of John Wiley & Sons Inc.
Spinal nerves
The spinal nerves form part of the peripheral nervous system, carrying communication between the periphery of the body and the central nervous system. After leaving the cord, spinal nerves branch extensively to innervate the trunk and limbs. As previously stated, each spinal nerve has two points of attachment to the spinal cord, a posterior or dorsal root containing sensory nerve fibres and an anterior or ventral root containing motor fibres. The two roots combine into a single mixed nerve. Immediately after exiting the intervertebral foramina the spinal nerves branch into three strands or rami (Figure 4.9):
- The ventral rami supply the anterior and lateral aspects of the body. In some areas the anterior rami unite almost immediately to form plexuses before going on to supply skin, bone, muscles and joints.
- The posterior rami pass backwards and supply the skin and muscles of a small area of the back of the head, neck and trunk.
- The rami communicantes arise from the base of the ventral rami in the thoracic and upper part of the lumbar spine and carry sympathetic fibres. The rami communicantes have two branches, white and grey (see Chapter 5).
A plexus is a complex interwoven network of nerves serving a particular area of the body, i.e. the cervical plexus serving the neck, the brachial plexus serving the upper limbs and the lumbosacral plexus serving the lower limbs (Figure 4.2). During the developmental process small skeletal muscles innervated by different ventral rami fuse to become larger muscles but retain their original sensory and motor nerve innervation. This results in convergence of the ventral rami of adjacent spinal nerves; their fibres blend to produce a series of compound nerve trunks which make up a plexus.
The cervical plexus is composed of the first five cervical spinal nerves. It is found at the level of the first to fourth cervical vertebrae, covered by muscle. This plexus includes the phrenic nerve which originates from C3, 4 and 5 and passes through the chest to supply the diaphragm, a key respiratory muscle. Other component nerves innervate the neck muscles and the skin of the neck and upper chest.
The brachial plexus is composed of rami from spinal nerves C5–8 and T1. This plexus supplies the skin and muscles of the arms, shoulders and some chest muscles. It lies in the neck, shoulder and axilla. It gives rise to five large nerves:
- The axillary nerve supplies the deltoid muscle, shoulder joint and overlying skin
- The radial nerve, the largest branch, supplies the triceps, wrist extensors, and the skin of the thumb, first two fingers and the lateral half of the third finger
- The musculocutaneous nerve supplies the muscles of the upper arm and the skin of the forearm.
- The median nerve supplies the muscles of the front of the forearm, and the small muscles and skin of the thumb, first two fingers and lateral half of the third finger
- The ulnar nerve supplies the muscles of the ulnar aspect of the forearm and the palm of the hand, and the skin of the little finger and medial aspect of the third finger
The lumbar plexus is comprised of the ventral rami of the first three and part of the fourth lumbar nerves. It lies in front of the transverse processes and behind the psoas muscle. The main branches are:
- The iliohypogastric, ilioinguinal and genitofemoral nerves supplies the muscles and skin of the lower abdomen, upper and medial aspects of the thigh and the inguinal region
- The lateral cutaneous nerve of the thigh supplies the skin of the lateral aspect of the thigh
- The femoral nerve divides into cutaneous and muscular branches to supply the skin and muscles of the front of the thigh. The saphenous nerve is another branch of the femoral nerve and it supplies the medial aspect of the leg, ankle and foot
- The obturator nerve supplies the adductor muscles of the thigh and skin of the medial aspect of the thigh down to the knee
- The lumbosacral trunk travels to the pelvis and combines with the sacral plexus
The sacral plexus is comprised of the ventral rami of L4 to S3. It lies in the posterior wall of the pelvic cavity. The main branches are:
- The sciatic nerve, the largest in the body, supplies the hamstring muscles and then divides into:
- the tibial nerve, serving the muscles and skin of the posterior lower leg, sole of foot and toes, the heel, the lateral aspect of the ankle and some of the dorsum of the foot
- and the common peroneal nerve, serving the skin and muscles of the anterior aspect of the lower leg and dorsum of the foot and toes.
- The pudendal nerve (S2–4) serves the external anal and urethral sphincters and surrounding skin
The coccygeal plexus is small, comprised of part of the fourth and fifth sacral and coccygeal nerves, and serves the skin around the coccyx and anal area.
The spinal nerves T2 to T11 do not form plexuses. They are called the intercostal nerves and pass directly to the structures they innervate. These include the intercostal muscles between the ribs, abdominal muscles and the skin of the chest and back.
Reflex activity in the spinal cord
A reflex is an immediate, rapid, involuntary, predictable sequence of motor actions that occur in response to a particular sensory stimulus. Reflexes may be inborn, such as the protective withdrawal reflexes seen in response to pain. These reflexes result from connections made between neurones during development. The reflex response of limb withdrawal may be so rapid that perception of pain in the cerebrum and withdrawal of affected part may be simultaneous. Reflexes may also be acquired, i.e. a driver responds to a danger on the road ahead by putting a foot on the break. These complex motor patterns are rapid and automatic, but learned rather than innate and are strengthened by repetition. Most reflexes, acquired or innate can be modified or suppressed.
A reflex arc is the pathway followed by the nerve impulse that produces a reflex action. The building blocks of a reflex arc are:
1. Sensory receptor – a specialised cell or the distal end of a sensory nerve
2. Sensory neurone along which the impulse is conducted into the central nervous system grey matter. In some reflex pathways branches of the neurone also relay impulses to the brain, bringing conscious awareness of the reflex action
3. Interneurone (association neurone) in the spinal cord to carry the impulse from sensory to motor neurones
4. Motor neurone to carry impulse to the effector organ that will respond
5. Effector organ – this may be a muscle or gland
If the effector is skeletal muscle, then the reflex is a somatic reflex. If the effector is smooth muscle, cardiac muscle or a gland it is a visceral (autonomic) reflex (autonomic reflexes are presented in Chapter 5).
Reflexes may be monosynaptic or polysynaptic. In monosynaptic reflex arcs, such as the stretch reflex, the response is very rapid. Polysynaptic reflexes produce more complex responses; the interneurones of the arc may synapse with nerves serving several different muscle groups.
The patellar stretch reflex is an example of a monosynaptic reflex arc (Figure 4.10). Stretch reflexes cause muscles to contract in response to stretching of the muscle (increase in muscle length) providing automatic regulation of muscle length. The patellar stretch reflex is triggered by the stretching of sensory nerve endings in the patellar tendon and the thigh muscle spindles by a hammer strike to the tendon. An impulse travels along the sensory nerve into the spinal cord to the cell body of the lower motor neurone in the anterior grey horn. The ensuing motor impulse results in sudden contraction of the thigh muscle and an extension of the limb with a forward kick of the foot (Figure 4.10). Loss of this reflex indicates damage somewhere in the reflex pathway and, because it is easy to provoke, this reflex is commonly used as a diagnostic tool.
Figure 4.10 Stretch reflex. This monosynaptic reflex arc has only one synapse in the CNS – between a single sensory neurone and a single motor neurone. A polysynaptic reflex arc to antagonistic muscles that includes two synapses in the CNS and one interneurone is also illustrated. Plus signs (+) indicate excitatory synapses; the minus sign (−) indicates an inhibitory synapse.
Reproduced from Principles of Anatomy and Physiology 12e by Gerard Tortora and Bryan Derrickson. Copyright (2009, John Wiley & Sons). Reprinted with permission of John Wiley & Sons Inc.
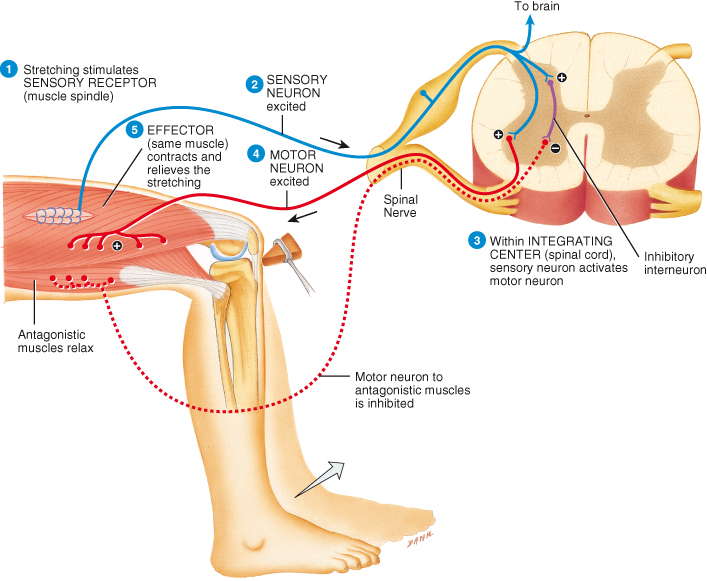
Although this simple stretch reflex is monosynaptic, a polysynaptic reflex arc is triggered simultaneously to relax the antagonistic muscles in an example of reciprocal inhibition (Figure 4.10). Reciprocal inhibition serves to stop the antagonistic muscles resisting the shortening of the stretched muscle which is essential for smooth coordinated movements. While the stretch reflex occurs automatically, higher centres in the brain are informed through ascending tracts of the degree of muscle stretch (length) and can subsequently modify the muscle activity through descending fibres.
Superficial reflexes are elicited by stroking of the skin with an object. The plantar response or Babinski response is the most clinically important. Stroking the sole of the foot should produce a plantar reflex (curling of the toes), but if there is damage to the motor cortex or the corticospinal tracts an abnormal response will occur causing the big toe to extend and the other toes to spread out laterally (Babinski response or sign). This response is present in infants but disappears as inhibitory descending motor pathways develop, if the higher centres or the descending pathways are damaged, the Babinski sign re-emerges. Superficial reflexes depend both on functional reflex arcs and on corticospinal tracts. The Babinski sign indicates an upper motor lesion.
Voluntary and reflex activities are often combined in stereotyped motor patterns embedded in the spinal cord. While walking, running or jumping are voluntary movements, assistance with coordination and control of muscle tone is maintained by spinal reflexes under the control of higher centres via the rubro-, reticulo-, vestibulo-, and tectospinal tracts.
Removal of descending input from the brain, as in spinal cord injury or stroke results in increased tone or spasticity of muscles below the level of injury. Damage to the reflex pathway, as in cauda equina injury, results in loss of muscle tone or flaccidity of muscles below the level of injury.
SUMMARY
This chapter has described the complex structure of the spinal cord and shown how its functions are both integral and essential to the somatic and autonomic functions of the body. Even a rudimentary understanding of the structure and functions of this intricate part of the central nervous system will aid the understanding of conditions such as traumatic spinal cord injury, cauda equina syndrome, poliomyelitis and motor neurone disease, facilitating and promoting patient education and care.
SUGGESTED READING
Marieb EM (2004) Human Anatomy and Physiology. (6th edition). San Francisco, Ca: Pearson Benjamin Cummings.
Martini FH (2006) Fundamentals of Anatomy and Physiology. (7th edition). San Francisco, Ca: Pearson Benjamin Cummings.
Tortora GJ, Derrickson SR (2006) Principles of Anatomy and Physiology. (11th edition). Hoboken, NJ: John Wiley and Sons.