
In geologic time the first period of the Mesozoic Era is known as the Triassic Period. It began 251 million years ago, at the close of the Permian Period, and ended 199.6 million years ago, when it was succeeded by the Jurassic Period.
The Triassic Period marked the beginning of major changes that were to take place throughout the Mesozoic Era, particularly in the distribution of continents, the evolution of life, and the geographic distribution of living things. At the beginning of the Triassic, virtually all the major landmasses of the world were collected into the supercontinent of Pangea. Terrestrial climates were predominately warm and dry (though seasonal monsoons occurred over large areas), and the Earth’s crust was relatively quiescent. At the end of the Triassic, however, plate tectonic activity picked up, and a period of continental rifting began. On the margins of the continents, shallow seas, which had dwindled in area at the end of the Permian, became more extensive. As sea levels gradually rose, the waters of continental shelves were colonized for the first time by large marine reptiles and reef-building corals of modern aspect.
The Triassic followed on the heels of the largest mass extinction in the history of the Earth. This event occurred at the end of the Permian, when 85 to 95 percent of marine invertebrate species and 70 percent of terrestrial vertebrate genera died out. During the recovery of life in the Triassic Period, the relative importance of land animals grew. Reptiles increased in diversity and number, and the first dinosaurs appeared, heralding the great radiation that would characterize this group during the Jurassic and Cretaceous periods. Finally, the end of the Triassic saw the appearance of the first mammals—tiny, fur-bearing, shrewlike animals derived from reptiles.
Another episode of mass extinction occurred at the end of the Triassic. Though this event was less devastating than its counterpart at the end of the Permian, it did result in drastic reductions of some living populations—particularly of the ammonoids, primitive mollusks that have served as important index fossils for assigning relative ages to various strata in the Triassic System of rocks.
The name Trias (later modified to Triassic) was first proposed in 1834 by the German paleontologist Friedrich August von Alberti for a sequence of rock strata in central Germany that lay above Permian rocks and below Jurassic rocks. (The name Trias referred to the division of these strata into three units: the Bunter [or Buntsandstein], Muschelkalk, and Keuper.) Alberti’s rock sequence, which became known as the “Germanic facies,” had many drawbacks as a standard for assigning relative ages to Triassic rocks from other regions of the world, and so for much of the 19th and 20th centuries Triassic stages were based mainly on type sections from the “Alpine facies” in Austria, Switzerland, and northern Italy. Since the mid-20th century more complete sequences have been discovered in North America, and these now serve as the standard for Triassic time in general. Meanwhile, studies of seafloor spreading and plate tectonics have yielded important new information on the paleogeography and paleoclimatology of the Triassic, allowing for a better understanding of the evolution and extinction of life-forms and of the paleoecology and paleobiogeography of the period. In addition, paleontologists continue to be occupied with defining the lower and upper boundaries of the Triassic System on a worldwide basis and with understanding the reasons for the mass extinctions that took place at those boundaries.
The environment at the beginning of the Triassic Period had relatively little variation. All of the Earth’s continents were joined into a continuous swath of land that nearly stretched from one pole to the other, whereas the remainder of the Earth’s surface during this time was occupied by a large ocean. This arrangement simplified ocean circulation and thus the distribution of heat and moisture. In addition, since the temperature gradient between the poles and the Equator was relatively small, the Triassic world contained fewer distinct habitats than during other periods.
At the beginning of the Triassic Period, the land was grouped together into one large C-shaped supercontinent. Covering about one-quarter of the Earth’s surface, Pangea stretched from 85° N to 90° S in a narrow belt of about 60° of longitude. It consisted of a group of northern continents collectively referred to as Laurasia and a group of southern continents collectively referred to as Gondwana. The rest of the globe was covered by Panthalassa, an enormous world ocean that stretched from pole to pole and extended to about twice the width of the present-day Pacific Ocean at the Equator. Scattered across Panthalassa within 30° of the Triassic Equator were islands, seamounts, and volcanic archipelagoes, some associated with deposits of reef carbonates now found in western North America and other locations.
Projecting westward between Gondwana and Laurasia along an east-west axis approximately coincident with the present-day Mediterranean Sea was a deep embayment of Panthalassa known as the Tethys Sea (previously discussed in chapter 1). This ancient seaway was later to extend farther westward to Gibraltar as rifting between Laurasia and Gondwana began in the Late Triassic (about 229 million to 200 million years ago). Eventually, by Middle to Late Jurassic times, it would link up with the eastern side of Panthalassa, effectively separating the two halves of the Pangea supercontinent. Paleogeographers reconstruct these continental configurations using evidence from many sources, the most important of which are paleomagnetic data and correspondences between continental margins in shape, rock types, orogenic (mountain-building) events, and distribution of fossilized plants and land vertebrates that lived prior to the breakup of Pangea. In addition, the apparent polar-wandering curves (plots of the apparent movement of the Earth’s magnetic poles with respect to the continents through time) for modern-day Africa and North America converge between the Carboniferous and Triassic periods and then begin to diverge in the Late Triassic, which indicates the exact time when the two continents began to separate and the Tethys Sea began to open up.
Thick sequences of clastic sediments accumulated in marginal troughs bordering the present-day circum-Pacific region as well as the northern and southern margins of the Tethys, while shelf seas occupied parts of the Tethyan, circum-Pacific, and circum-Arctic regions but were otherwise restricted in distribution. Much of the circum-Pacific region and the northeastern part of Tethys were bordered by active (that is, convergent) plate margins, but the northwestern and southern margins of Tethys were passive (that is, divergent) during the Triassic. At the end of the Triassic, increased tectonic activity contributed to rising sea levels and an increase in the extent of shallow continental shelf seas.
Along the western margin of modern North America, a major subduction zone was present where the eastward-moving oceanic plate of eastern Panthalassa slid under the continental plate of Pangea. The Panthalassa plate carried fragments of island arcs and microcontinents that, because of their lesser density, could not be subducted along with the oceanic plate. As these fragments reached the subduction zone, they were sutured onto the Cordilleran belt of North America, forming what geologists refer to as allochthonous terranes (fragments of crust displaced from their site of origin). This process of “accretionary tectonics” (or obduction) created more than 50 terranes of various ages in the Cordilleran region, including the Sonomia and Golconda terranes of the northwestern United States, both of which were accreted in the Early Triassic (about 251 million to 246 million years ago). The former microcontinent of Sonomia occupies what is now southeastern Oregon and northern California and Nevada.
Worldwide climatic conditions during the Triassic seem to have been much more homogeneous than at present. No polar ice existed. Temperature differences between the Equator and the poles would have been less extreme than they are today, which would have resulted in less diversity in biological habitats.
Beginning in the Late Permian and continuing into the Early Triassic, the emergence of the supercontinent Pangea and the associated reduction in the total area covered by continental shelf seas led to widespread aridity over most land areas. Judging from modern conditions, a single large landmass such as Pangea would be expected to experience an extreme, strongly seasonal continental climate with hot summers and cold winters. Yet the paleoclimatic evidence is conflicting. There are several indicators of an arid climate, including the following: red sandstones and shales that contain few fossils, lithified dune deposits with cross-bedding, salt pseudomorphs in marls, mudcracks, and evaporites. On the other hand, there is evidence for strong seasonal precipitation, including braided fluvial (riverine) sediments, clay-rich deltaic deposits, and red beds of alluvial and fluvial origin. This dilemma is best resolved by postulating a monsoonal climate, particularly during the Middle and Late Triassic, over wide areas of Pangea. Under these conditions, cross-equatorial monsoonal winds would have brought strong seasonal precipitation to some areas, especially where these winds crossed large expanses of open water.
Another indication of temperate and tropical climates is coal deposits. Their presence invariably indicates humid conditions with relatively high rainfall responsible for both lush vegetational growth and poor drainage. The resultant large swamps would act as depositional basins wherein the decomposing plant material would be transformed gradually into peat. Such humid conditions must have existed in high latitudes during the later stages of the Triassic Period, on the basis of the occurrence of coals in Triassic formations in Arctic Canada, Russia, Ukraine, China, Japan, South America, South Africa, Australia, and Antarctica.
It has been postulated that, because of the large size of Panthalassa, oceanic circulation patterns during the Triassic would have been relatively simple, consisting of enormous single gyres in each hemisphere. East-west temperature extremes would have been great, with the western margin of Panthalassa being much warmer than the eastern. A permanent westerly equatorial current would have provided warm waters to Tethys, enabling reefs to develop there wherever substrates and depths were favourable.
Additional important evidence regarding paleoclimate is provided by the nature of Triassic fossils and their latitudinal distribution. The biotas of the period are fairly modern in aspect, and so their life habits and environmental requirements can be reconstructed with relative confidence from comparisons to living relatives. For example, the presence of colonial stony corals as framework builders in Tethyan reefs of Late Triassic age suggests an environment of warm shelf seas at low latitudes. These seas must have been sufficiently shallow and clear to allow penetration of adequate light for photosynthesis by zooxanthellae, a type of protozoa inferred to be, perhaps for the first time in geologic history, symbiotically associated with reef-building corals and aiding in their calcification.
The geographic distribution of modern-day animals indicates, with few exceptions, that faunal diversity decreases steadily in both hemispheres as one approaches the poles. For example, ectothermic (cold-blooded) amphibians and reptiles show a much higher diversity in the warmer low latitudes, reflecting the strong influence of ambient air temperatures on these animals, which are unable to regulate their internal temperature. The evidence from Triassic fossils, however, is equivocal: the distribution of Triassic amphibians and reptiles shows only a slight change with latitude, although the distribution of ammonoids from the upper part of the Lower Triassic shows a much stronger geographic gradient. It may be that Triassic marine invertebrates were more sensitive to differences in ambient temperature than land vertebrates or that ambient temperature differences were greater in the ocean than on land. There is also the possibility that both of these conditions existed.
The boundary between the Paleozoic and Mesozoic eras was marked by the Earth’s third and largest mass extinction episode, which occurred immediately prior to the Triassic. As a result, Early Triassic biotas were impoverished, though diversity and abundance progressively increased during Middle and Late Triassic times. The fossils of many Early Triassic life-forms tend to be Paleozoic in aspect, whereas those of the Middle and Late Triassic are decidedly Mesozoic in appearance and are clearly the precursors of things to come. New land vertebrates appeared throughout the Triassic. By the end of the period, both the first true mammals and the earliest dinosaurs had appeared.
Periodic large-scale mass extinctions have occurred throughout the history of life. Indeed, it is on this basis that the geologic eras were first established. Of the five major mass extinction events, the one best known is the last, which took place at the end of the Cretaceous Period and killed the dinosaurs. However, the largest of all extinction events occurred between the Permian and Triassic periods at the end of the Paleozoic Era, and it is this third mass extinction that profoundly affected life during the Triassic. The fourth episode of mass extinction occurred at the end of the Triassic, drastically reducing some marine and terrestrial groups, such as ammonoids, mammal-like reptiles, and primitive amphibians, but not affecting others.
Though the Permian-Triassic mass extinction was the most extensive in the history of life on Earth, it should be noted that many groups were showing evidence of a gradual decline long before the end of the Paleozoic. Nevertheless, 85 to 95 percent of marine invertebrate species became extinct at the end of the Permian. On land, four-legged vertebrates and plants suffered significant reductions in diversity across the Permian-Triassic boundary. Only 30 percent of terrestrial vertebrate genera survived into the Triassic.
Many possible causes have been advanced to account for these extinctions. Some researchers believe that there is a periodicity to mass extinctions, which suggests a common, perhaps astronomical, cause. Others maintain that each extinction event is unique in itself. Cataclysmic events, such as intense volcanic activity and the impact of a celestial body, or more gradual trends, such as changes in sea levels, oceanic temperature, salinity, or nutrients, fluctuations in oxygen and carbon dioxide levels, climatic cooling, and cosmic radiation, have been proposed to explain the Permian-Triassic crisis. Unlike the end-Cretaceous event, there is no consistent evidence in rocks at the Permian-Triassic boundary to support an asteroid impact hypothesis, such as an anomalous presence of iridium and associated shocked quartz (quartz grains that have experienced high temperatures and pressures from impact shock). A more plausible theory is suggested by finely laminated pyritic shales, rich in organic carbon, that are commonly found at the Permian-Triassic transition in many areas. These shales may reflect oceanic anoxia (lack of dissolved oxygen) in both low and high latitudes over a wide range of shelf depths, perhaps caused by weakening of oceanic circulation. Such anoxia could devastate marine life, particularly the bottom-dwellers (benthos). Any theory, however, must take into account that not all groups were affected to the same extent by the extinctions.
The trilobites, a group of arthropods long past their zenith, made their last appearance in the Permian, as did the closely related eurypterids. Rugose and tabulate corals became extinct at the end of the Paleozoic. Several superfamilies of Paleozoic brachiopods, such as the productaceans, chonetaceans, and richthofeniaceans, also disappeared at the end of the Permian. Fusulinid foraminiferans, useful as late Paleozoic index fossils, did not survive the crisis, nor did the cryptostomate and fenestrate bryozoans, which inhabited many Carboniferous and Permian reefs. Gone also were the blastoids, a group of echinoderms that persisted in what is now Indonesia until the end of the Permian, although their decline had begun much earlier in other regions. However, some groups, such as the conodonts (a type of tiny marine invertebrate), were little affected by this crisis in the history of life, although they were destined to disappear at the end of the Triassic.
The end-Triassic mass extinction was less devastating than its counterpart at the end of the Permian. Nevertheless, in the marine realm some groups such as the conodonts became extinct, while many Triassic ceratitid ammonoids disappeared. Only the phylloceratid ammonoids were able to survive, and they gave rise to the explosive radiation of cephalopods later in the Jurassic. Many families of brachiopods, gastropods, bivalves, and marine reptiles also became extinct. On land a great part of the vertebrate fauna disappeared at the end of the Triassic, although the dinosaurs, pterosaurs, crocodiles, turtles, mammals, and fishes were little affected by the transition. Plant fossils and palynomorphs (spores and pollen of plants) show no significant changes in diversity across the Triassic-Jurassic boundary. Sea-level changes and associated anoxia, coupled with climatic change, were the most likely causes for the end-Triassic extinction.
The difference between Permian and Triassic faunas is most noticeable among the marine invertebrates. At the Permian-Triassic boundary the number of families was reduced by half, with an estimated 85 to 95 percent of all species disappearing.
Ammonoids were common in the Permian but suffered drastic reduction at the end of that period. Only a few genera belonging to the prolecanitid group survived the crisis, but their descendants, the ceratitids, provided the rootstock for an explosive adaptive radiation in the Middle and Late Triassic. Ammonoid shells have a complex suture line where internal partitions join the outer shell wall. Ceratitids have varying external ornamentation, but all share the distinctive ceratitic internal suture line of rounded saddles and denticulate lobes, as shown by such Early Triassic genera as Otoceras and Ophiceras. The group first reached its acme and then declined dramatically in the Late Triassic. In the Carnian Stage (the first stage of the Late Triassic) there were more than 150 ceratitid genera. In the next stage, the Norian, there were fewer than 100, and finally in the Rhaetian Stage there were fewer than 10. In the Late Triassic evolved bizarre heteromorphs with loosely coiled body chambers, such as Choristoceras, or with helically coiled whorls, such as Cochloceras. These aberrant forms were short-lived, however. A small group of smooth-shelled forms with more complex suture lines, the phylloceratids, also arose in the Early Triassic. They are regarded as the earliest true ammonites and gave rise to all post-Triassic ammonites, even though Triassic ammonoids as a whole almost became extinct at the end of the period.

Fossilized echinoids (sea urchins). Shutterstock.com
Other marine invertebrate fossils found in Triassic rocks, albeit much reduced in diversity compared with those of the Permian, include gastropods, bivalves, brachiopods, bryozoans, corals, foraminiferans, and echinoderms. These groups are either poorly represented or absent in Lower Triassic rocks but increase in importance later in the period. Most are bottom-dwellers (benthos), but the bivalve genera Claraia, Posidonia, Daonella, Halobia, and Monotis, often used as Triassic index fossils, were planktonic and may have achieved widespread distribution by being attached to floating seaweed. Colonial stony corals became important reef-builders in the Middle and Late Triassic. For example, the Rhaetian Dachstein reefs from Austria were colonized by a diverse fauna of colonial corals and calcareous sponges, with subsidiary calcareous algae, echinoids, foraminiferans, and other colonial invertebrates. Many successful Paleozoic articulate brachiopod superfamilies (those having valves characterized by teeth and sockets) became extinct at the end of the Permian, which left only the spiriferaceans, rhynchonellaceans, terebratulaceans, terebratellaceans, thecideaceans, and some other less important groups to continue into the Mesozoic. The brachiopods, however, never again achieved the dominance they held among the benthos of the Paleozoic, and they may have suffered competitively from the adaptive radiation of the bivalves in the Mesozoic.
Fossil echinoderms are represented in the Triassic by crinoid columnals and the echinoid Miocidaris, a holdover from the Permian. The crinoids had begun to decline long before the end of the Permian, by which time they were almost entirely decimated, with both the flexible and camerate varieties dying out. The inadunates survived the crisis. They did not become extinct until the end of the Triassic and gave rise to the articulates, which still exist today.
Vertebrate animals appear to have been less affected by the Permian-Triassic crisis than were invertebrates. The fishes show some decline in diversity and abundance at the end of the Paleozoic, with acanthodians (spiny sharks) becoming extinct and elasmobranchs (primitive sharks and rays) much reduced in diversity. Actinopterygians (ray-finned fishes), however, continued to flourish during the Triassic, gradually moving from freshwater to marine environments, which were already inhabited by subholostean ray-finned fishes (genera intermediate between palaeoniscoids and holosteans). The shellfish-eating hybodont sharks, already diversified by the end of the Permian, continued into the Triassic.
Fossils of marine reptiles such as the shell-crushing placodonts (which superficially resembled turtles) and the fish-eating nothosaurs occur in the Muschelkalk, a rock formation of Triassic marine sediments in central Germany. The nothosaurs, members of the sauropterygian order, did not survive the Triassic, but they were ancestors of the large predatory plesiosaurs of the Jurassic. The largest inhabitants of Triassic seas were the early ichthyosaurs, superficially like dolphins in profile and streamlined for rapid swimming. These efficient hunters, which were equipped with powerful fins, paddlelike limbs, a long-toothed jaw, and large eyes, may have preyed upon some of the early squidlike cephalopods known as belemnites. There also is evidence that these unusual reptiles gave birth to live young.
On land the vertebrates are represented in the Triassic by labyrinthodont amphibians and reptiles, the latter consisting of cotylosaurs, therapsids, eosuchians, thecodontians, and protorosaurs. All these tetrapod groups suffered a sharp reduction in diversity at the close of the Permian. In fact, 75 percent of the early amphibian families and 80 percent of the early reptilian families disappeared at or near the Permian-Triassic boundary. Whereas Early Triassic forms were still Paleozoic in aspect, new forms appeared throughout the period, and by Late Triassic times the tetrapod fauna was distinctly Mesozoic in aspect. Modern groups whose ancestral forms appeared for the first time in the Middle and Late Triassic include lizards, turtles, rhynchocephalians (lizardlike animals), and crocodilians.
The mammal-like reptiles, or therapsids, suffered pulses of extinctions in the Late Permian. The group survived the boundary crisis but became virtually extinct by the end of the Triassic, possibly because of competition from more efficient predators, such as the thecodonts. The first true mammals, which were very small, appeared in the Late Triassic (the shrewlike Morganucodon, for example). Although their fossilized remains have been collected from a bone bed in Great Britain dating from the Rhaetian Stage at the end of the Triassic, the evolutionary transition from therapsid reptiles to mammals at the close of the Triassic is nowhere clearly demonstrated by well-preserved fossils.
First encountered in the Early Triassic, the thecodonts became common during the Middle Triassic (about 246 million to 229 million years ago) but disappeared before the beginning of the Jurassic some 176 million years ago. Typical of this group of archosaurs (or “ruling reptiles”) in the Triassic were small bipedal forms belonging to the pseudosuchians. Forms such as Lagosuchus were swift-running predators that had erect limbs directly under the body, which made them more mobile and agile. This group presumably gave rise to primitive dinosaurs belonging to the saurischian and ornithischian orders during the Late Triassic to Early Jurassic. The early dinosaurs were bipedal, swift-moving, and relatively small compared with later Mesozoic forms, but some, such as Plateosaurus, reached lengths of 8 metres (26 feet). Coelophysis was a Late Triassic carnivorous dinosaur about 2 metres (6 to 8 feet) long. Its fossils have been found in the Chinle Formation in the Petrified Forest National Park of northeastern Arizona in the United States. The dinosaur group was to achieve much greater importance later in the Mesozoic, resulting in the era being informally called the “Age of Reptiles.”
Some of the earliest lizards may have been the first vertebrates to take to the air. Gliding lizards, such as the small Late Triassic Icarosaurus, are thought to have developed an airfoil from skin stretched between extended ribs, which would have allowed short glides similar to those made by present-day flying squirrels. Similarly, Longisquama had long scales that could have been employed as primitive wings, while the Late Triassic Sharovipteryx was an active flyer and may have been the first true pterosaur (flying reptile). All these forms became extinct at the end of the Triassic, their role as fliers being taken over by the later pterosaurs of the Jurassic and Cretaceous.
Land plants were affected by the Permian-Triassic crisis, but less so than were the animals, since the demise of late Paleozoic floras had begun much earlier. The dominant understory plants in the Triassic were the ferns, while most middle-story plants were gymnosperms (plants having exposed seeds)—the cycadeoids (an extinct order) and the still-extant cycads and ginkgoes. The upper story of Triassic forests consisted of conifers; their best-known fossil remains are preserved in the Upper Triassic Chinle Formation.
While extensive forests did exist during the Triassic, widespread aridity on the northern continents in the Early and Middle Triassic limited their areal extent, which resulted in generally poor development of floras during this period. However, in the Late Triassic the occurrence of water-loving plants, such as lycopods (vascular plants now represented only by the club mosses), horsetails, and ferns, suggests that the arid climate changed to a more moist monsoonal one and that this climatic belt extended as high as latitude 60° N. Subtropical to warm-temperate Eurasian flora lay in a belt between about 15° and 60° N, while north of this belt were the temperate Siberian (Angaran) flora, extending to within 10° of the Triassic North Pole. In the southern continents the Permian Glossopteris and Gangamopteris seed fern flora, adapted to cool, moist conditions, were replaced by a Triassic flora dominated by Dicroidium, a seed fern that preferred warm, dry conditions—which indicates major climatic changes at the Permian-Triassic boundary. Dicroidium, a genus of the pteridosperm order, was part of an extensive Gondwanan paleoflora that was discovered in the Late Triassic Molteno Formation of southern Africa and elsewhere. This paleoflora extended from 30° to well below 60° S. Few fossil remains exist from the Triassic for the equatorial zone between 15° N and 30° S.
In the oceans the coccolithophores, an important group of still-living marine pelagic algae, made their first appearance during the Late Triassic, while dinoflagellates underwent rapid diversification during the Late Triassic and Early Jurassic. Dasycladacean marine green algae and cyanobacteria were abundant throughout the Triassic.
Most of the dinosaurs of this period were smaller than those that appeared later in the era. Coelophysis and Herrerasaurus were small agile dinosaurs that possessed several morphological features adapted to hunting prey. Some Triassic dinosaurs, such as Plateosaurus, did grow to relatively large sizes, however.
This is the name of a genus of small carnivorous dinosaurs found as fossils from the Late Triassic Period of North America.
Coelophysis was a primitive theropod dinosaur. Usually growing to a length of about 2 metres (6.6 feet), it was very light, weighing only about 18–23 kg (40–50 pounds), and had a long, slender neck, tail, and hind leg. The head was long and narrow, and the jaws were equipped with many sharp teeth.
Coelophysis, like other predatory dinosaurs, was an agile, lightly built predator that possibly fed on other small reptiles and early relatives of mammals. It is representative of the basal stock from which later, more derived theropod dinosaurs evolved. Coelophysis is known from a massive death assemblage of hundreds of skeletons found at Ghost Ranch, near Abuquiu, New Mexico, and first excavated in 1947.
Herrerasaurus, a genus of primitive carnivorous dinosaur or close relative of the dinosaurs, was found as a fossil in Argentine deposits from the Late Triassic Period. It had long, powerful hind legs for running and short forelimbs equipped with three recurved claws for grasping and raking. The lower jaw possessed large inward-curving teeth and was flexible for holding prey. Herrerasaurus reached a length of about 3 metres (10 feet) and weighed about 180 kg (400 pounds).
Herrerasaurus flourished at a time just before dinosaurs became the dominant land animals. Its remains help clarify the sequence of anatomic changes that occurred during early dinosaur evolution. It closely resembled the common ancestor of all dinosaurs, and it retained the carnivorous habits and features of predatory animals that were ancestral to dinosaurs and their relatives. Although some features, such as their three-toed feet, resemble those of true theropod dinosaurs, they lack some features that distinguish theropods from saurischians, such as overlapping wrist bones and an opposable thumb.

The skull of the Herrerasaurus shows its lower jaw and inner-curving teeth. © www.istockphoto.com/breckeni
Fragmentary fossil remains of Herrerasaurus were first discovered in the early 1960s, but it was not until 1988, when several skeletons were discovered in the Ischigualasto Formation of northwestern Argentina, that researchers could complete the first picture of the animal.
This dinosaur genus is known from extensive fossil material found in Europe dating to the Late Triassic Period. The fossils were representative of the prosauropods, an early group that might have been ancestral to the giant sauropod dinosaurs of later time periods.
Plateosaurus was among the earliest dinosaurs to attain a relatively large size, growing to about 8 metres (26 feet) long. It was more massive than earlier dinosaurs and had bones that were stocky and thick. Although Plateosaurus could rise up on its two very strong hind legs, its forelimbs also were relatively well developed and strong, and it may have walked on two or four legs for various purposes. The small skull was perched atop a long, flexible neck and contained flat teeth serrated on the front and back edges.
Plateosaurs were the first known large herbivores among the dinosaurs. Dinosaurs related to Plateosaurus have been found in South Africa, North America, and China. The prosauropod group of dinosaurs is not found after the Early Jurassic Period (about 200 million to 176 million years ago), which is when the first of the large “true” sauropod dinosaurs appeared. This fact, along with the increasing trend to large size among prosauropods, supports the idea that sauropods evolved directly from prosauropods, although some authorities regard the two as separate groups.
In addition to the dinosaurs that evolved during the period, the Triassic was the time of several notable mammal-like reptiles. The genera Bauria, Cynognathus, Thrinaxodon, and Tritylodon give important clues to paleontologists studying the emergence of mammals. The Triassic also saw the arrival of the pterosaurs, ichthyosaurs, as well as the possible precursors to teleost fishes and the ancestors of the present-day sturgeon (Acipenser).
This genus of mammal-like reptiles that inhabited parts of present-day South Africa during the Early Triassic Period. The skull of Bauria had several mammal-like features. A secondary palate separates air and food passages. The teeth show specialization and are differentiated into a set of incisor-like, caninelike, and molarlike cheek teeth. A single bone, the dentary, dwarfs the other lower jawbones, a trend toward the mammalian condition of only one bone, the dentary. Bauria and its relatives did not survive the Early Triassic.
The Chondrosteiformes were an extinct order of ray-finned saltwater fishes (class Actinopterygii) comprising a single family Chondrosteidae. These fishes were prominent in seas during the Early Triassic to Late Jurassic (from 251 million to 145.5 million years ago). Some species were suctorial feeders that probably gave rise to present-day sturgeon.
Members of this genus of extinct advanced therapsids (mammals and their relatives) were found as fossils in Lower Triassic deposits in South Africa and South America. Cynognathus is representative of the Theriodontia, a group of cynodont therapsids that gave rise to the earliest mammals.
Cynognathus was approximately as large as a modern wolf and, like the wolf, was an active predator. The body of Cynognathus was not massively constructed. The tail was short, and the limbs were tucked well under and close to the body, providing the potential for rapid and efficient locomotion. The skull was long and had openings for the attachment of strong muscles used in opening and closing the jaws. The lower jaw was dominated by the dentary bone. The other lower-jaw elements, characteristic of reptiles, were relatively reduced, as in mammals and their near relatives. The teeth were regionally specialized on the jaw into different forms, as in mammals. Incisors adapted to nipping were followed by strongly developed canines, important features in predatory animals. Separated from the canines by a gap, or diastema, was a series of cheek teeth that sliced the animal’s food into smaller, more easily swallowed particles. A well-developed secondary palate separated food passages from breathing passages. The vertebral column was well differentiated.
This genus of extinct pelecypods (clams) serves as a guide, or index, fossil in Triassic rocks. The shell is characterized by a wide dorsal region and by fine, radiating, riblike lineations. The shell is circular in outline and may show fine growth lines.
This genus of extinct reptiles is very closely related to the ancestral archosaurs (a group containing present-day crocodiles and birds and ancestral dinosaurs and pterosaurs). Specimens are found as fossils in Middle Triassic rocks of South Africa (245 million to 240 million years ago). Euparkeria was about 1 metre (3 feet) long and lightly built. It probably progressed on all four limbs or on only two back limbs. Like other archosaurs, Euparkeria had an opening in the skull between its eyes and nasal breach (the antorbital opening) and two additional apertures in the skull behind one eye (the upper and lower temporal openings). Its teeth were set in sockets, rather than being attached to the side of the jawbone or perched atop it. These teeth were long, sharp, and recurved, which attested to the carnivorous habit that seems to have been common among the first archosaurs. Euparkeria also possessed teeth on its palate, which was also common among earlier reptiles and amphibians.
Most members of this group of extinct aquatic reptiles were very similar to porpoises in appearance and habits. Ichthyosaurs were distant relatives of lizards and snakes (lepidosaurs) and were the most highly specialized aquatic reptiles, but ichthyosaurs were not dinosaurs.
Ichthyosaurs had a very wide geographic distribution, and their fossil remains span almost the entire Mesozoic Era. However, they were most abundant and diverse during the Triassic and Jurassic periods (251 million to 145 5 million years ago). Excellent fossil specimens occur in the fine-grained Early Jurassic shales of southern Germany. In one specimen, the entire outline of the body is preserved, including the outline of a well-developed, fleshy dorsal fin. Several specimens are known in which the skeletal remains of small, immature ichthyosaurs are fossilized within the bodies of larger individuals, even within the birth canal.

The cast of a Lower Jurassic ichtyosaur depicts the skeletal structure, while the overlaid shadow approximates the original body shape. Shutterstock.com
Ichthyosaurus, a representative genus from which the larger group takes its name, was about 3 metres (10 feet) long and was probably able to move through the water at high speeds. Very fishlike in appearance, it is especially well known from Early Jurassic deposits in England. The body was streamlined. No distinct neck was present, and the head blended smoothly into the body. The limbs were modified into paddlelike appendages used to steer the animal. It propelled itself by using a well-developed fishlike tail and by undulating the body. The vertebral column, which was formed from disklike structures, bent downward into the lower lobe of the caudal, or tail, fin. The upper lobe was unsupported by bone. Early reconstructions of ichthyosaurs showed them with the spinal column straightened, and it was not until well-preserved evidence was found that the bent condition of the backbone became apparent. The skull and jaws of Ichthyosaurus were long and contained numerous sharp teeth. The eyes were very large, and the nostrils were positioned far back on the top of the skull (another specialized adaptation to an aquatic existence). They probably fed largely upon fish as well as other marine animals. It is unlikely that they ventured onto land, and they certainly reproduced in the water. If stranded ashore, they would have been as helpless as beached whales.
Ichthyosaurs are first known from the Triassic Period of Asia, where they began as long-bodied, undulating swimmers without many of the specializations seen in later species. By the Late Triassic some lineages had achieved great size. Fossils from the western United States and Canada indicate that some ichthyosaurs could exceed 13 metres (43 feet) in length. Deep-bodied and with long fins, these appear to have been ambush predators that fed on fishes. The typical ichthyosaur form was fully realized by the Early Jurassic, when the tunalike body plan suggestive of high-speed pursuit and great mobility asserted itself. By this time, however, the other lineages of ichthyosaurs had become extinct. Ichthyosaurs persisted into Late Cretaceous times and may have been well adapted for deep diving as well as near-shore predation, but all species became extinct well before the end of the Cretaceous Period.
Scientists might be able to trace the origins of the teleosts, the dominant group of fishes in the world today, back to Leptolepis, genus of marine fishes very closely related to the first teleosts. Leptolepis was abundant in the world’s Mesozoic seas and was herringlike in size and appearance. Fragmentary remains from earlier and later rocks may indicate an earlier origin and longer persistence for the genus than the Jurassic period dates indicate. In many anatomical details, Leptolepis is intermediate between the more primitive holosteans and the more advanced teleost fish.
Marasuchus, a genus of archosaurian reptiles, inhabited part of present-day South America during the Ladinian Stage (some 237 million to 229 million years ago) of the Middle Triassic Epoch. Marasuchus fossils were discovered in the Los Chañares Formation of the Ischigualasto–Villa Union Basin in northwestern Argentina. Marasuchus was not a dinosaur. Members of this genus and others (such as Silesaurus and Eucoelophysis) are classified as basal dinosauromorphs, or direct precursors to the dinosaurs. Together the basal dinosauromorphs and the dinosaurs make up the Dinosauromorpha, a group containing all reptiles more closely related to dinosaurs than to pterosaurs.
Marasuchus was lightly built and small, growing to 30–40 cm (about 12–16 inches). It was bipedal, walking with an upright (parasagittal) gait, like that of modern mammals and birds. All parts of the reptile’s skeletal anatomy are known from fossils except for the skull and lower jaw. One of the diagnostic features of dinosaurs, a hole in the hip socket (acetabulum) of the pelvis for the femur (thigh bone), is absent in Marasuchus; however, it possessed characteristics, such as an elongate pubis and the presence of an anterior trochanter on the femur, similar to those found in dinosaurs. The presence of these characteristics in Marasuchus shows that some of the features limited to dinosaurs and their close relatives had begun to evolve in the Middle Triassic prior to the formal origin of dinosaurs in the Late Triassic.
Until 2003, Marasuchus was thought to be one of the closest relatives of the dinosaurs. At present, it has been supplanted by other dinosauromorphs such as Silesaurus. Nonetheless, Marasuchus remains an important animal for understanding the origin and evolution of dinosaur characteristics.
Myophoria, a genus of extinct clams found as fossils in Triassic rocks, is readily identified by its distinctive shell form and ornamentation. As a result, it is a useful guide, or index, fossil for the Triassic Period. The shell in Myophoria is angular, with prominent ribs that radiate from its apex. Fine growth lines encircle the shell at right angles to the ribs.
Members of this genus of marine reptiles have been found as fossils from the Triassic Period in southwestern and eastern Asia, North Africa, and especially Europe.
Nothosaurus was characterized by a slender body, long neck and tail, and long limbs. Although the animal was aquatic, the limbs were less specialized for swimming than they were in more advanced sauropterygians such as pistosaurs, pliosaurids, and plesiosaurids. The palate in the nothosaurs was closed, the air passages being separated from the food passages—an adaptation that aided feeding while in the water. The skull was long and flat with large openings. Numerous pointed teeth were present along the margins of the jaws. Nothosaurus moved through the water by undulating its body and by swimming with its limbs. As did the other sauropterygians, Nothosaurus evolved from terrestrial reptiles distantly related to lizards and snakes.
Phytosaurs were a group of heavily armoured semiaquatic reptiles. These animals were found as fossils from the Late Triassic Period. Phytosaurs were not dinosaurs. Both groups, rather, were archosaurs.
Phytosaurs were able to move about easily on land, and, although they were not ancestral to the crocodiles, they were distantly related and resembled crocodiles in appearance and probably in habits as well. The long, pointed jaws were armed with numerous sharp teeth, and it is probable that the phytosaurs preyed largely upon fishes. Like crocodiles, they had several rows of bony armour embedded in the skin along the back. The nostrils in the phytosaurs were set on a crest high on the skull in front of the eyes. This adaptation allowed them to float just underneath the water’s surface, with only the nostrils protruding above it.
Phytosaur fossils occur in North America, Europe, and India, but their remains have not been found in the southern continents. Familiar genera include Phytosaurus, Belodon, and Rutiodon, which was more than 3 metres (10 feet) long and whose skull alone measured about 1 metre.
Pleuromeia, a genus of extinct lycopsid plants from the Triassic Period, is characterized by an unbranched trunk up to 2 metres (6.6 feet) tall. Unlike other arborescent lycopsids of the Carboniferous Period (about 360 million to 300 million years ago), such as Lepidodendron and Sigillaria, Pleuromeia had a four-lobed bulblike base rather than a branching underground rhizome. A crown of long, thin leaves persisted near the growing tip of the trunk. Leaves and leaf bases were lost from lower portions of the plant. Like its relatives, Pleuromeia reproduced by spores. Some species produced a single large cone at the trunk apex, and others may have produced many smaller cones. Nonetheless, the details of how Pleuromeia reproduced remain unclear. The genus was widely distributed, and specimens are known from Russia, Europe, China, and Australia.
Pterosaurs were a group of flying reptiles that flourished during all periods (Triassic, Jurassic, and Cretaceous) of the Mesozoic Era. Although pterosaurs are not dinosaurs, both are archosaurs, or “ruling reptiles,” a group to which birds and crocodiles also belong.
Ancestors of pterosaurs tended toward a bipedal gait, which thus freed the forelimbs for other uses. These limbs evolved into wings in birds and pterosaurs, but, instead of feathers, pterosaurs developed a wing surface formed by a membrane of skin similar to that of bats. In bats, however, all of the fingers except the thumb support the membrane. In pterosaurs, the membrane was attached solely to the elongated fourth finger (there was no fifth finger). The first three fingers were slender, clawed, clutching structures. When the pterosaur was not in flight, the finger and membrane were extended rearward along the flanks. In addition to the main flight membrane, an accessory membrane stretching between the shoulder and wrist reduced turbulence on the wing. The pterosaur wing appears to have been well adapted to flight. Embedded within it was a system of fine, long, keratinous fibers that ran parallel to one another like the feather shafts of birds. This arrangement enhanced strength and maneuverability in flight.
The body was compact, and the hind legs were long and slender, like those of birds, and were easily able to support the animal on land. Despite the considerable size of the forelimbs, the bones were hollow and thin-walled, which kept weight low. The skull, with its long, slender beak, was delicate but strong, with most of the component bones being fused. The eyes were large, and the eyeball was reinforced by a series of bony plates (sclerotic ring).
The brain was large and apparently comparable in structure to that of birds, and, as in that group, sight rather than smell appears to have been the dominant sense. Most pterosaur remains are found in sediments close to what were bodies of water (fossils are well preserved in such places), so little is known about the diversity of forest or plains pterosaurs.
Traditionally, two major groups of pterosaurs have been recognized. Rhamphorhynchoidea is a term that has included all the basal forms up to the Late Jurassic Period (about 161 million to 145 5 million years ago). These are typified by relatively long tails, long fifth toes, sharply pointed teeth, and only slightly elongated wing metacarpals (palm bones). Rhamphorhynchoids were the first pterosaurs, and they are found in deposits from the Late Triassic Period Genera of this group include Eudimorphodon and Peteinosaurus, both found in Italian Triassic deposits; these had wingspans of less than 1 metre (3.3 feet) Dimorphodon, from the Early Jurassic of England, was about 1.5 metres (5 feet) from wingtip to wingtip Rhamphorhynchus was a late form from the Late Jurassic Period and had a wingspan of about 1 metre (3.3 feet). It has long been realized, however, that Rhamphorhynchoidea is an artificial grouping of primitive forms, as some members are actually more closely related to the other major group of pterosaurs, the Pterodactyloidea. Pterodactyloids appeared in the Late Jurassic and survived into the Cretaceous, when the earlier forms of pterosaurs had become extinct. The earliest Late Jurassic pterodactyloid is Pterodactylus, of which numerous individuals are known from Solnhofen Limestone of Bavaria, Germany. Pteranodon, which grew to 7 metres (23 foot), was also a Pterodactyloid. Lacusovagus (family Chaoyangopteridae, a group of toothless pterodactyloids) is known from a single fossilized skull discovered in Cretaceous rocks in Brazil. It possessed a 5-metre (16.4-feet) wingspan and is the only member of Chaoyangopteridae found outside China. No pterosaur remains are more recent than the Cretaceous; their ecological roles were eventually taken over by birds.
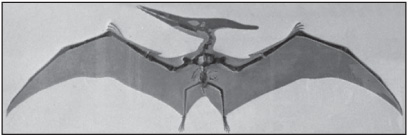
Pteranodon skeleton and restoration of wings. Courtesy of the American Museum of Natural History, New York
Remarkable for its distinctive shell, Tetractinella is a genus of extinct brachiopods (lamp shells) found as fossils in Triassic marine rocks. Its shell has prominent ribs and intervening troughs radiating from its apex and margins extending in a weblike fashion between the ribs; the shell is compressed in profile. One species of Tetractinella is an excellent example of a phenomenon known as homeomorphy, in which an organism simulates an unrelated organism in form and function. Tetractinella trigonella, a Middle Triassic species from Italy, is remarkably similar to the unrelated Cheirothyris fleuriausa, from the Late Jurassic (about 150 million years ago) marine rocks of Germany. The two forms are separated by a great geographic distance and by a large span of time.
Thrinaxodon is an extinct genus of cynodont, a close mammal relative. Members of this genus have been found as fossils in continental deposits formed during the Early Triassic Period in southern Africa. Thrinaxodon was a lightly built animal about 0.5 metre (2 feet) long.
Tritylodon, a genus of extinct cynodont therapsids (mammal relatives), have been found as fossils in Late Triassic and Early Jurassic rocks in southern Africa and North America. These fossils have been dated to between 208 million and 200 million years ago. Tritylodonts are characterized by a distinctive dentition: the anterior incisors are separated from the complicated cheek teeth by a pronounced gap. The cheek teeth possess two to four rows of cusps arranged longitudinally. In features of skull construction and general overall skeletal construction the tritylodonts closely approached true mammals, though they were too specialized to have given rise to the mammals and may have been contemporary with some of the earliest of them. In jaw construction and articulation tritylodonts were not mammalian. The lower jaw retained components from earlier amniotes rather than the single bone, the dentary, that is characteristic of the mammals. It is probable that the habits of Tritylodon were similar to those of the later rodents and multituberculates.
One example of a notable mollusk appearing during the period is Tropites, a genus of extinct cephalopods (animals similar to the modern squid and octopus but with an external shell) found as fossils in marine rocks of the Late Triassic Period. Because of its narrow time range, Tropites is a good index fossil (useful for stratigraphic correlations). Tropites is characterized by a distinctive, easily recognizable, globular shell within a central keel.
One link in the evolution of cone-bearing plants is exemplified by the genus Voltzia, a group dating to the Early Triassic epoch. It belongs to the family Voltziaceae, order Coniferales (sometimes Voltziales). The genus showed interesting modifications of the seed-cone complex of earlier forms. The pollen-bearing cone was an axis with spirally arranged pollen cases. The seed-bearing cone had three ovules on five flattened and fused scales, a trend of fusion and simplification that continued in later coniferous genera.
The Triassic Period is characterized by few geologic events of major significance, in contrast to the subsequent periods of the Mesozoic Era (the Jurassic and Cretaceous periods), when the supercontinent Pangea fragmented and the new Atlantic and Indian oceans opened up. This does not mean, however, that the period was geologically silent.
The beginning of continental rifting in the Late Triassic caused stretching of the crust in eastern North America along the Appalachian Mountain belt from the Carolinas to Nova Scotia, resulting in normal faulting in this region. There, grabens (fault-bounded basins) received thick clastic (rock fragment) sequences from the erosion of the nearby Appalachians, which were later intruded by igneous dikes and sills. In similar fault-controlled basins between Africa and Laurasia, evaporite deposits were formed in arid or semiarid environments as seawater from the Tethys Sea periodically spilled into these newly formed troughs and then evaporated, leaving behind its salts. Evaporites of Late Triassic and Early Jurassic age in Morocco and off eastern Canada were apparently deposited in such tectonically formed basins.
Mountain building was restricted during the Triassic, with relatively minor orogenic activity taking place along the Pacific coastal margin of North America and in China and Japan. The unmetamorphosed nature of the Triassic rocks of the Newark Group, a rock sequence in eastern North America known for its dinosaur tracks and fossils of freshwater organisms, indicates that its sediments were deposited after the main phase of the Appalachian orogeny in the late Paleozoic.
The three rock series of the Triassic Period are made up of seven stages. The Lower and Middle Triassic Series contain two stages each, whereas the Upper Triassic Series is divided into three stages. More specifically, the Induan and Olenekian stages make up the Lower Triassic Series, and the Anisian and Ladinian stages make up the Middle Triassic Series. In contrast, the Upper Triassic Series is divided into the Carnian, Norian, and Rhaetian stages.
The lowermost of two divisions of the Lower Triassic Series are those of the Induan Stage, representing those rocks deposited worldwide during Induan time (from 251 million to 249.5 million years ago). The stage name is derived from the Indus River in the Salt Range of Pakistan. The stratotype for the Induan, as originally defined, is the strata above the Chhideru beds and below the Upper Ceratite Limestone of the Salt Range. The Induan stage is subdivided into two substages, which in ascending order are the Griesbachian and Dienerian. Induan marine strata are correlated worldwide by six biozones containing ammonoid cephalopod index fossils. Five of these biozones have designated type localities in North America. These zones cannot be used for non-marine rocks, however the Induan Stage underlies the Olenekian Stage of the Lower Triassic Series and overlies the Changhsingian Stage of the Permian Series.
The uppermost of two divisions of the Lower Triassic Series is the Olenekian Stage, representing those rocks deposited worldwide during Olenekian time (249.5 million to 245.9 million years ago). The stage name is derived from the Olenyok, or Olenek, River of Siberia. The stratotype for the Olenekian, which was defined in 1956, is the strata in the lower course of the Olenyok River that rest upon deposits of the Induan Stage and that are overlain by those of the Anisian Stage. The Olenekian Stage is subdivided into two substages, which in ascending order are the Smithian and Spathian. Olenekian marine strata are correlated worldwide by five ammonoid cephalopod biozones, four of which have designated type localities in North America. These zones cannot be used for nonmarine rocks, however. The Olenekian Stage underlies the Anisian Stage of the Middle Triassic Series and overlies the Induan Stage of the Lower Triassic Series.
The lowermost of two divisions of the Middle Triassic Series is the Anisian Stage, representing those rocks deposited worldwide during Anisian time (245.9 million to 237 million years ago). The stage name is derived from an area of limestone formations along the Anisus River at Grossreifling in the Austrian Alps. The Anisian Stage is subdivided, in ascending order, into the Aegean, Bithynian, Pelsonian, and Illyrian substages. Anisian marine strata are correlated worldwide by seven biozones containing ammonoid cephalopod index fossils. All these biozones have designated type localities in North America. These zones cannot be used for nonmarine strata, however. The Anisian Stage underlies the Ladinian Stage of the Middle Triassic Series and overlies the Olenekian Stage of the Lower Triassic Series.
The Ladinian Stage is the uppermost of two divisions of the Middle Triassic Series. It represents those rocks deposited worldwide during Ladinian time (237 million to 228.7 million years ago). The stage name is derived from the Ladini people of the Dolomites in northern Italy. The stratotypes for the Ladinian are the Buchenstein and Wengen beds of the Dolomites. The Ladinian is subdivided into two substages, which in ascending order are the Fassanian and Longobardian. Ladinian marine strata are correlated worldwide by five distinct ammonoid cephalopod biozones, all of which have designated type localities in North America. These zones cannot be used for non-marine strata, however. The Ladinian Stage underlies the Carnian Stage of the Upper Triassic Series and overlies the Anisian Stage of the Middle Triassic Series.
The lowermost of three divisions of the Upper Triassic Series is the Carnian Stage, representing those rocks deposited worldwide during Carnian time (228.7 million to 216.5 million years ago). The stage name is probably derived from the Austrian state of Kärnten (Carinthia), where the stratotype is located. The Carnian Stage is subdivided into two substages, which in ascending order are the Julian and Tuvalian. Carnian marine strata are correlated worldwide by six ammonoid cephalopod biozones, all of which have designated type localities in North America. These zones cannot be used for nonmarine strata, however. The Carnian Stage underlies the Norian Stage of the Upper Triassic Series and overlies the Ladinian Stage of the Middle Triassic Series.
The Norian Stage is second of three divisions in the Upper Triassic Series. It represents those rocks deposited worldwide during Norian time (216.5 million to 203.6 million years ago). The stage was named after an ancient Roman province south of the Danube River in present-day Austria. The stratotype for the Norian is a formation known as the beds with Cyrtopleurites bicrenatus (an ammonoid index fossil) at Sommeraukogel, Hallstatt, Austria. The Norian Stage is subdivided into three substages, which in ascending order are the Lacian, Alaunian, and Sevatian. Norian marine strata are correlated worldwide by six ammonoid cephalopod biozones, all of which have designated type localities in North America. These zones cannot be used for nonmarine strata, however. The Norian Stage underlies the Rhaetian Stage of the Upper Triassic Series and overlies the Carnian Stage of the Upper Triassic Series.

The Lower Norian rock formations of the Petrified Forest National Park, Arizona, U.S. Shutterstock.com
The uppermost of three divisions in the Upper Triassic Series is the Rhaetian Stage, representing those rocks deposited worldwide during Rhaetian time (203.6 million to 199.6 million years ago). The stage name is derived from the Rhaetian Alps of Italy, Switzerland, and Austria; the stratotype is the Kössen beds at Kendelbachgraben, Sankt Wolfgang, Austria. Rhaetian rocks are transitional in age and sometimes placed in the Lower Jurassic. In Great Britain the Rhaetian (or Rhaetic) consists of lagoonal deposits, such as limestones, shales, and marls, as well as bone beds that contain fragments of amphibians and reptiles. Elsewhere Rhaetian marine strata are correlated worldwide by two distinct ammonoid cephalopod biozones, both of which have designated type localities in North America. Rhaetian rocks may also consist of shelf limestones with characteristic brachiopods, conodonts, and other shelly forms. The Rhaetian Stage underlies the Hettangian Stage of the Lower Jurassic Series and overlies the Norian Stage of the Upper Triassic Series.