Cirripedia having a carapace, consisting either of a capitulum on a peduncle, or of an operculated shell with a basis. Body formed of six thoracic segments, generally furnished with six pairs of cirri; abdomen rudimentary, but often bearing caudal appendages; mouth with the labrum not capable of independent movements; larva firstly uniocular, with three pairs of legs, lastly, binocular, with six pairs of thoracic legs.
In the sketch of the three Orders given in the Introduction, it will have been seen that the differences in their structure are so great, that it would have been hardly possible to have given a single blended account of the whole Class. But as all common Cirripedes are included in the present Order, here would have been the natural place for a full description of their external and internal structure. Having, however, been necessarily, yet perhaps unfortunately, led to give, in my former volume, a description of this kind of the Lepadidæ; and as it is necessary to give a similar account of the other great family of the Order, namely, the Balanidæ, I have found it more convenient to make this latter account comparative and supplemental to the former one on the Lepadidæ, and so serve for the Order, rather than attempt to give a separate description in full of it. For this latter plan would have involved much useless repetition, as, on account of the many exceptions and limitations necessary to almost every statement, there is little choice between a description of great length and a mere diagnostic character of the Order, such as I have given above.
The Thoracica may be divided into three very natural Families, of nearly equal value; firstly, the Balanidæ, or sessile Cirripedes, which may be subdivided into two sub-families, also very natural, the Balaninæ and Chthamalinæ; secondly, the Verrucidæ, containing only one genus; and thirdly, the Lepadidæ, or pedunculated Cirripedes. These three families differ from each other, besides in mere external appearance, almost exclusively in the relation of the different portions of their external covering or carapace, and of the muscles moving such portions. In the Balanidæ, the four opercular valves surrounding the orifice leading into the sack, are capable of other movements, besides being opened and shut; whereas all the other valves are immoveably united together. In the Lepadidæ, the valves answering to the opercular valves, are furnished with a muscle only for shutting them; whereas the peduncle answering to the basis in the Balanidæ is capable of various movements. In the Verrucidæ the shell is singularly asymmetrical; only half of the operculum (either the right or the left side, this varying even in the same species) being moveable; the other half being immoveably united to the remaining valves; and the whole shell has only one muscle serving to shut the moveable half of the operculum. All the internal parts and organs are very similar in the above three Families. If, however, the internal structure of one of the two sub-families, into which the Balanidæ may be divided, namely, of the Balaninæ, be compared with that of the Lepadidæ, several important differences may be detected; — on the one hand, in the Balaninæ, the presence of branchiæ, the extremely complicated cementing apparatus, the difference in structure between the third and succeeding pairs of cirri, the large palpi, the notched labrum, and the laterally double teeth of the mandibles; — and on the other hand, in the Lepadidæ, the presence of ovigerous fræna, caudal appendages, bullate labrum, and often prominent olfactory orifices. But if the Lepadidæ be compared in these several respects with the other sub-family, or Chthamalinæ, which cannot possibly be removed out of the family of Balanidæ, many of these differences break down and disappear, in some or all of the species.
The Lepadidæ include, as has previously been noticed, a much greater range of difference than the Balanidæ; and this is what might have been expected, for it is the most ancient family, and extinction has done its work, separating genera, which, in accordance to analogy, we may suppose were once more nearly connected by intermediate forms. The Lepadidæ, in one sense, may be taken as the type of their order; for they have undergone less “morphological differentiation;” that is, they differ the least from the last larval stage, and seem to give the most general idea of a Thoracic Cirripede. On the other hand, if we mean, as some authors do, by the word type, that form which, in the group in question, has been most modified, and illustrates every peculiarity of its class in the strongest manner, then we must look to the Balaninæ, and to its typical genus, Balanus, for the most Cirripedial form. In this genus the different portions of the carapace differ most, and subserve to a certain extent different ends, and in minute structure are most complicated; here the cementing apparatus, which offers the main characteristic of the whole sub-class, is most complex; here the several pairs of cirri differ most from each other in structure and action; here the peculiar branchiæ (organs apparently derived from the modification of another organ, itself confined to Cirripedes, viz., the ovigerous fræna) are best developed; here the nervous system is most highly concentrated; and, lastly, here we meet with the largest and most massive species of the whole group.
1. Family BALANIDÆ, (or Sessile Cirripedes).
Cirripedia without a peduncle; scuta and terga furnished with depressor muscles; other valves united immoveably together.
TABLE OF CONTENTS.
|
|
Page |
|
Structure of shell |
34 |
|
“of the individual compartments |
43 |
|
“of the radii |
45 |
|
“of the alæ |
47 |
|
“of the sheath |
48 |
|
“of the basis |
49 |
|
“of the opercular valves (scuta and terga) |
51 |
|
Growth of whole shell and microscopical structure |
54 |
|
Muscles of sack |
61 |
|
Branchiæ |
63 |
|
Thorax and body |
65 |
|
Muscular system |
68 |
|
Movements and muscles of the cirri |
71 |
|
Mouth |
74 |
|
Cirri |
81 |
|
Caudal appendages |
85 |
|
Alimentary canal |
85 |
|
Circulatory system |
87 |
|
Nervous system |
88 |
|
Eyes and vision |
93 |
|
Acoustic organs |
95 |
|
Olfactory sacks |
97 |
|
Male organs of generation |
97 |
|
Female organs of generation |
100 |
|
Metamorphoses and homologies |
102 |
|
Larva, first stage |
103 |
|
Larva, second stage |
109 |
|
Larva, last or pupal stage |
110 |
|
Act of metamorphosis |
126 |
|
On the homologies of the carapace |
131 |
|
Cementing apparatus |
133 |
|
Affinities, classification, variation |
152 |
|
Rate of growth, exuviation, &c. |
156 |
|
Geographical range and habitats |
159 |
|
Geological history |
172 |
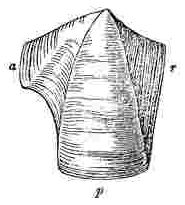
Almost every one who has walked over a rocky shore knows that a barnacle or acorn-shell is an irregular cone, formed generally of six compartments, with an orifice at the top, closed by a neatly-fitted, moveable lid, or operculum. Within this shell the animal’s body is lodged; and through a slit in the lid, it has the power of protruding six pairs of articulated cirri or legs, and of securing by their means any prey brought by the waters within their reach. The basis is firmly cemented to the surface of attachment. The whole shell, basis, and operculum consists, as we have already seen, of the first three segments of the head, modified into a singularly constructed carapace, which encloses the mouth and rest of the body. The anterior extremity of the shell is situated in the centre of the basis, where indeed, by due care, the antennæ of the pupa may be always detected; and the posterior extremity is directed vertically upwards.
The best published description of the structure of the shell of a sessile Cirripede, is given by Dr. Coldstream, in the ‘Encyclopædia of Anatomy and Physiology,’ article ‘Cirrhopoda.’
Structure of Shell.
When the shell of a sessile Cirripede or barnacle, for instance, of a Balanus, is first examined, the structure appears extremely complicated; but this can hardly be considered as really the case. The structure will, I think, be best understood by recalling to mind that of Pollicipes, — the oldest known genus, from which, in one sense, all ordinary Cirripedes, both sessile and pedunculated, seem to radiate. I must premise, and the fact in itself deserves early notice, that the homologies of the several parts in the pedunculated and sessile Cirripedes admits of no doubt, — that is, if amongst the pedunculated, the genus Pollicipes, or certain species of Scalpellum, be taken as a standard of comparison. The peduncle corresponds with the basis, as may be clearly seen, if a Pollicipes with a short peduncle, and a Balanus, with a deep cup-formed or cylindrical basis be compared, for the contained parts are similar, and both grow at their upper edges upwards and outwards. Secondly, the valves round the lower part of the capitulum of a Pollicipes, though generally much more numerous, and forming more than one whorl or circle, and not so closely packed together, answer to the compartments forming the shell of a sessile Cirripede; this is shown by their lateral and downward growth, by their upper ends generally projecting freely above the cavity in which the animal’s body is lodged; and in the case Pollicipes mitella, by an actual resemblance in outline, some being triangular, some broad at the upper end, and some sub-rhomboidal, and, lastly, in the manner in which they slightly overlap and indent each other: moreover further resemblances in the relative position and even in the size of the several valves, will hereafter be pointed out between certain sessile genera amongst the Chthamalinæ and certain genera of the Lepadidæ. Thirdly, the scuta and terga in Pollicipes, so strikingly resemble in manner of growth in position relatively to the animal’s body — in shape — and even in being articulated together, the valves which form the operculum or lid of sessile Cirripedes, that their identity is at once obvious.
Dr. J. E. Gray long ago observed these homologies. If Lepas be taken, the comparison is not quite so simple, owing to the growth of all the valves in that genus being upwards; but in several species of Scalpellum we may see the intermediate steps between the normal downward growth of the valves in Pollicipes, and the abnormal upward growth in Lepas.
It may be well here further to premise, that apparently none of the sutures in the shells of Cirripedes correspond with the articulations between the three archetype cephalic segments, of which the whole shell is formed; or with the eight elemental pieces, of which each separate segment in the archetype crustacean is known to consist. But, as I believe, the several valves in the shell of a Cirripede are homologous, or at least analogous, with the sclerodermic plates, of which the carapace of the Podophthalmia is formed; with this difference, that in the latter they become, after their first formation, united together into a single piece, and are thus moulted as a whole; whereas in Cirripedes, the valves or sclerodermic plates are not moulted, but continue to be added to throughout life.
Milne Edwards, ‘Annales des Sciences Naturelles,’ tom. xviii, (1852), .
In Pollicipes, there is no difficulty in understanding the growth of the lower valves of the capitulum, especially if a species be taken in which these valves stand a little way apart: at each period of growth, they are added to at their basal edges and a little way up both sides; at the same time, a new membrane connecting them together is formed, the old membrane disintegrating, or being left hanging in tatters to the last zone of growth. Now if we look at the shell of a sessile Cirripede, there is no essential difference in the growth of the compartments or valves; all grow downwards and laterally; but they overlap each other much more laterally than in Pollicipes, and the connecting membrane is in most parts reduced to a mere film jammed in between the valves; but, in the case of the opercular membrane, it still remains wide, and is periodically moulted.
In the annexed woodcut (fig. 1), of the rostrum of Balanus Hameri, the downward growth and the lateral growth on both sides is plain. The modified sides (rr) for convenience sake, have been called the radii; they invariably overlap the adjoining compartments. The middle part (p), has been called the wall, or parietal portion: in the specimen figured, the walls and radii are distinctly separated, but in some cases, especially amongst the Chthamalinæ, the lines of growth are absolutely continuous from one to the other. In fig. 2 of a Lateral compartment of the same Balanus, we have the same essential structure; but the left side (a) is more protuberant, and is hollowed out in its lower half; it is, also, more distinctly separated from the parietal portion: this side has for convenience been called the ala; it is invariably overlapped by the adjoining compartment: in some few cases, as in Pachylasma, the ala is not hollowed out in its lower part, and from being added to in a straight line along its whole edge, with the lines of growth continuous with those on the wall, it differs hardly at all in appearance from a radius. Lastly, in fig. 3 of the carina, or compartment facing the rostrum, we have alæ (aa) on both sides; these being, as in all cases, overlapped by the adjoining compartments.
Fig. 1.
Fig. 2.
Fig. 3.

p, p, Parietes; r, r, Radii; a, a, Alæ.
Fig. 1, Rostrum with two radii, serving in the Chthamalinæ for rostro-lateral compartments.
Fig. 2, always serving for lateral and carino-lateral compartments.
Fig. 3, Carina, serving in the Chthamalinæ, also, as a rostrum.
Now, the compartments in the shell of every sessile Cirripede, are without exception constructed on the above three simple patterns. In number, they are 8, 6 or 4, or all confluent together.
Considering this simplicity in growth and form of the separated compartments, it seems at first surprising that the construction and enlargement of the whole shell in Balanus, should long have been viewed as a difficulty. But the radii, from growing against rectangular indentations, or rather furrows, in the opposed compartments, come to be set a little inwards; and their external surfaces assume a very different appearance from the wall-portions of the compartments, which grow against the surface of attachment. In different species, the summits of the radii (and of the alæ) grow either very much more obliquely than in the species figured, or more squarely — that is, they extend from tip to tip of the adjoining compartments, parallel to the basis. In this latter case, and when the surfaces of the radii differ considerably in appearance from the walls, as in Balanus tintinnabulum (Plate 1), I am not at all surprised that the radii should have been described as separate elements, and called “areæ interjectæ,” or “compartments of the second order:” for the shell of this Balanus seems to be composed of six wedges with their points upwards, namely, the parietal portions of the compartments, and of six other narrower wedges, the radii, with their points downwards; and the fact that these latter wedges consist simply of the sides of the parietal portions, modified by growing against the adjoining compartments, is completely masked. I should add, that sometimes the radii are not developed, which simply means that the overlapping lateral edges of the compartments have not been added to during growth.
The alæ are originally developed at the period of the metamorphosis, as slight lateral protuberances in the upper part of the compartments; from being overlapped, and therefore not exposed to external influences, and from growing (as in the case of the radii) against rectangular indentations or furrows in the adjoining compartments, they generally assume an extremely different appearance from the parietes, and might naturally be thought to have a very different nature. But the alæ in all cases (as is obvious in Pachylasma) are nothing but the protuberant lateral edges of the compartments, rendered thin and modified during growth. In order that the margins of the alæ should be received in an indentation, the upper internal surfaces of the walls of the recipient compartments are thickened all round, excepting where they receive the alæ. This thickened, upper, internal portion of the walls or shell, together with the alæ themselves, form the part called the sheath. The sheath sometimes blends insensibly into the lower parts of the compartments, and then perhaps it would not be thought to be a distinct element; but often it is abruptly separated by an overhanging edge (see Pl. 9, fig. 5 b, 9 b; Pl. 20, fig. 1 b; Pl. 25, fig. 1, K′) from the lower part, and then the sheath greatly complicates the internal appearance, but not the essential structure of the shell. The sheath acts beautifully, like an internal hoop, in strengthening the shell round the orifice, where it is naturally weaker than at the lower or basal end, where it adheres to the surface of attachment: in the upper part of the shell, moreover, the sutures between the compartments do not go straight through, but owing to the alæ projecting and being overlapped, are extremely oblique; or the joints, in the language of carpenters, may be said to be broken.
There is one other point of structure in the shells of the Balanidæ, more especially of species like Balanus tintinnabulum, which adds to their apparent complexity, namely, that the rim or orifice of the shell formed by the upper ends of the compartments, projects considerably above the opercular valves. In a young Balanus, immediately after the metamorphosis, the operculum is attached by the opercular membrane all round to the summits of the compartments, and there cannot be said to be any orifice to the shell itself, but only an orifice or slit between the opercular valves; but during growth, as the compartments are added to at their basal edges, their upper ends are deserted, and cease to enclose the sack, within which lies the animal’s body. Hence the upper ends come to project freely, either quite separately as in some species of Pollicipes, where they cannot be said to form an orifice; or more or less united into a ring so as to form an orifice, as in the different species of Balanidæ. It follows, that to understand the real shape of a Balanus, or rather of the cavity enclosing the animal’s body, all that part of the shell which projects above the opercular membrane, may, in imagination, be removed as something extraneous, like so many spines; not that I mean to say that these points of shell are dead; on the contrary, they are often porose and penetrated by numerous threads of corium. This upper part of the shell, thus produced so as to form an orifice, no doubt serves to protect the less strong and moveable operculum.
Fig. 4.
Octomeris.
Fig. 5.
Chthamalus.
Fig. 6.
Chamæsipho.
Fig. 7.
Balanus.
Fig. 8.
Tetraclita.
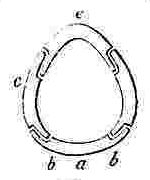
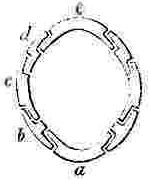
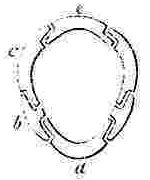
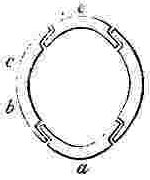
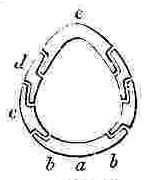
a, Rostrum; b, Rostro-lateral, c, Lateral, d, Carino-lateral compartment; e, Carina.
Horizontal sections through the Shells of the principal genera of Balanidæ, showing the arrangement of the Compartments. Genera 4, 5, and 6 belong to the Chthamalinæ; 7 and 8 to the Balaninæ.
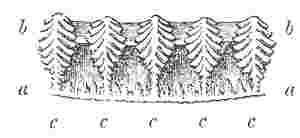
Number and Arrangement of the Compartments. — I have already stated that the shell, in every one of the Balanidæ, consists of eight, six, or four compartments, or of all fused together into a single piece; and that the compartments themselves are all constructed on the three simple patterns of which woodcuts (figs. 1, 2, 3) have been given. They are arranged in a certain definite order. The type arrangement is found amongst the Chthamalinæ, as might have been expected, inasmuch as this sub-family is so closely related to the ancient genera Pollicipes and Scalpellum, whence all the Thoracic Cirripedia may be said to radiate. In Octomeris (fig. 4) the type-arrangement of the compartments, eight in number, is well shown; the rostrum and carina resemble each other, and have alæ on both sides, and therefore are overlapped on both sides: the rostro-lateral compartments have radii on both sides, and therefore overlap the adjoining compartments on both sides; the lateral and carino-lateral compartments have radii on their carinal, and alæ on their rostral sides; and therefore overlap on one side, and are overlapped on the other side. Now the shell of every other sessile Cirripede differs, I believe, from that of Octomeris, only in the fusion together or abortion of some of the eight normal compartments: in one genus, however, Catophragmus, outer whorls of small compartments, arranged like the lower valves in the capitulum of Pollicipes, are superadded. The genus Chthamalus (fig. 5) differs from Octomeris only in the carino-lateral compartments being aborted, (as will presently be discussed), and hence has six compartments. Chamæsipho (fig. 6) differs from Chthamalus only in the rostro-lateral and lateral compartments being fused together; and hence has only four compartments. In Balanus (fig. 7) and the whole sub-family of the Balaninæ, the rostrum is compounded of the true rostrum, as seen in the type Octomeris, and of the two rostro-lateral compartments; hence this compounded rostrum has radii instead of alæ on both sides, and there are only six compartments. Tetraclita (fig. 8) and Elminius differ from Balanus only in having the carino-lateral compartments absent, and probably aborted; hence there are only four compartments. Lastly, in Pyrgoma, all the compartments are blended together into a single piece.
In Pollicipes, the old type-form of the whole order, and in Scalpellum, we have four valves, (answering to the operculum), surrounding the aperture leading into the sack, and the valves below are arranged in successive whorls, with a strong tendency to alternation. For, the rostrum alternates with, that is faces the interval between, the two scuta; the carina alternates with the two terga; and the upper lateral valves alternate with the scutum and tergum on each side. These four valves, namely, the carina and rostrum, which resemble each other in structure, and the pair of upper latera, which are larger than the other lateral valves, together form the uppermost whorl, or that beneath the scuta and terga. In the next whorl we have the rostro- and carino-lateral valves, alternating with those above them; and beneath them there are generally other valves, which decrease in size and display the same tendency to alternation. The valves here just specified, namely, the rostrum, carina, and three pairs of lateral valves, in the Lepadidæ, are so much larger, and are so much more commonly present, than the other valves of the capitulum, that to them alone I affixed special names. Now if amongst sessile Cirripedes we look to that genus, viz., Catophragmus, which comes in its whole structure the nearest to Pollicipes, one of the Lepadidæ, we find (as in fig. 4), firstly, a rostrum and carina resembling each other, and a pair of lateral compartments, larger than the other lateral pairs; these four valves alternating with the opercular valves: and, secondly, we find, but forming part of the same whorl, a pair of rostro-lateral and a pair of carino-lateral compartments, which, just as in Pollicipes, are larger than the exterior and lower valves. These lower little valves, I may remark, decrease in size in the successive whorls, and tend to alternate in position, just as in Pollicipes. Observing these several striking points of correspondence in the valves, (and indeed in the whole structure), of Catophragmus and Pollicipes, one is strongly inclined to suspect that in Catophragmus, and therefore in Octomeris and other sessile Cirripedes, although the rostro- and carino-lateral compartments appear to lie in the same whorl with the rostrum, carina, and large lateral compartments, yet that they really belong, as in Pollicipes and Scalpellum, to a lower whorl. Now if a very young shell of Balanus, immediately after the metamorphosis, be examined, the carino-lateral compartments will be found not to have been developed; they first appear after two or three zones of growth have been added to the other compartments; bearing in mind that in Pollicipes and in Catophragmus the lower whorls are added successively during growth, we find in this fact strong confirmation of the view that the carino-lateral compartments normally belong to a whorl beneath that including the rostrum, carina, and lateral compartments. Whether the rostro-lateral, like the carino-lateral compartments, are developed subsequently to the others, I have had no opportunity of ascertaining, and therefore cannot confirm the above analogical conclusion, namely, that they, also, belong to a lower whorl.
In the sub-family Balaninæ, which includes Balanus (woodcut 7), and Tetraclita (woodcut 8), the shell is characterised by not having rostro-lateral compartments, and by the rostrum having radii: now in Pachylasma giganteum, which undoubtedly belongs to the sub-family Chthamalinæ, at a very early age the rostro-lateral compartments become blended with the true rostrum, making a compound rostrum, exactly like the rostrum in the Balaninæ; distinct evidence of a similar fusion is retained throughout life (Pl. 15, fig. 1) in all three species of Chelonobia, which is undoubtedly a member of the Balaninæ. Hence, I think, we may conclude that in all the genera of the Balaninæ the rostro-lateral compartments are probably not aborted, but are blended with a normal rostrum (resembling that in woodcuts 4, 5, 6), making together a compound rostrum furnished with radii: it must, however, be observed that I could not detect any actual evidence of this fusion in Balanus, even immediately after the metamorphosis. In Chamæsipho (woodcut 6), either the rostro-lateral compartments attain a most unusual breadth, or, as is more probable, they have become confluent with the lateral compartments, which in the Lepadidæ seem to be the most persistent of all the lateral valves. In such genera as Tetraclita and Chthamalus, in which the carino-lateral compartments are absent, they may be fused with the lateral compartments or with the carina; but seeing that they are normally developed later than the other valves, it appears to be the simplest theory to assume, until the contrary be proved, that they are aborted. Finally, the somewhat unexpected conclusion that the shell (not including the operculum) of sessile Cirripedes normally consists of eight valves, four belonging to an upper whorl, and four to a lower whorl, all forced into a single ring, and often more or less fused together, though not strictly proved, is rendered highly probable. I will only further add, that the Basis perhaps represents several whorls of the small valves or scales on the peduncle of Pollicipes, fused together; the comparison of the basis with the calcareous cup, forming the lowest portion of the peduncle in Lithotrya, which has been made by some authors, I do not think is very accurate, as the cup in Lithotrya seems to have a special relation to the boring habits of that genus.
Fig. 9.
Basal edge of wall of compartment in Balanus tintinnabulum; a, a, outer lamina; b, b, inner lamina; c, c, longitudinal septa uniting the inner and outer laminæ with their ends denticulated.
Structure of the Individual Compartments.
If the basal margin of a compartment, for instance, of Balanus tintinnabulum, be examined, it appears sufficiently complicated, being composed of an outer and inner lamina, separated by longitudinal septa, which are denticulated at their bases; and the tubes formed by these longitudinal septa are crossed by transverse septa. On the other hand, in some cases, as in the genera Chthamalus and Elminius, each compartment consists of a simple shelly layer. These two extreme states graduate into each other: we have, firstly, on the internal surface, quite irregular points and ridges; these become regular, causing the internal surface to be longitudinally ribbed; then these ribs themselves become finely furrowed on their sides and at their lower ends, producing sharp, minute ridges, the ends of which I have called the denticuli; and, lastly, some of the denticuli on the adjoining longitudinal septa become united into a solid layer, forming the internal lamina of the wall. These denticuli do not generally cover the whole surface of the longitudinal ribs, but leave a portion near the outer lamina of the compartment smooth. The denticulated ends of the longitudinal septa project beyond the basal edge of both the outer and inner laminæ, and enter the mouths of the tubes (where such occur) in the basis, and thus strengthen the shell. The whole of the internal lamina generally is more or less striated longitudinally, thus displaying its origin from the union of the inner edges of the longitudinal septa. I need only further remark that on the internal surface of the outer lamina, between the main longitudinal septa, there are generally (as in the woodcut) smaller longitudinal ridges, which do not reach the inner lamina, and on this account alone are not called septa.
Tubes are formed by the longitudinal septa, between the outer and inner laminæ. These tubes are almost square, and are occupied by threads of corium, which enter at pores left open between the edge of the compartment and that of the basis on which it rests. The tubes extend high up the compartments; but in the uppermost part they are generally cut off by thin, transverse, calcareous septa, deposited by the ends of the threads of corium; a cancellated structure being thus produced. Or the uppermost part of each tube becomes filled up solidly with compact shelly layers, which are always first thrown down on the side of the tube facing the outside, and thus greatly strengthen the shell: in several instances, as in Balanus perforatus and Tetraclita porosa, in which the disintegration of the upper part of the shell is a necessary element in its growth for the enlargement of the orifice, these filled up tubes become exposed. In Coronula and Tubicinella, the tubes in their upper parts are, I believe, crossed only by transverse membranous septa.
Anomalies and exceptions. — In Tetraclita (Pl. 10, fig. 1 g, 1 h) from the branching of the longitudinal septa, several irregular rows of tubes are formed. In certain varieties of Balanus balanoides (Pl. 7, fig. 2 b), and in B. cariosus (Pl. 7, fig. 3 b), slight branching ridges on the internal surface of the walls, seem to answer to the longitudinal septa, and produce, during the downward growth of the shell, extremely irregular cells, and short tubes. In Balanus vinaceus (Pl. 2, fig. 7 d), the internal lamina, instead of being solid, as in every other species, is left cancellated, and thus betrays, much more plainly than usual, its origin in the united denticuli of the adjoining longitudinal septa. In Balanus porcatus, between the main longitudinal septa, there are (Pl. 6, fig. 4 e) what may be called rudimentary and disconnected longitudinal septa. In Coronula and its allies (Pl. 16, fig. 6, and Pl. 17, fig. 4 c) it is the outer lamina of the compartment which is anomalous; for in the two or three lower zones of growth, it forms only a ledge on each side of the longitudinal septa; which ledges, higher up, become confluent, and so form an ordinary outer lamina. In Coronula, also, the wall of each compartment (see transverse section, Pl. 16, figs. 5, 7) is very remarkable from being deeply folded, the folds being on their internal faces firmly calcified together, and on their external faces closely pressed together (often with a neatly serrated suture), so that the whole nature of the shell might be, as has happened, easily quite misunderstood; and the walls be considered as very thick, instead of being, as is really the case, very thin. In Chelonobia (Pl. 15, fig. 1), however, the walls are truly of such great thickness, that the nature of the relative parts might likewise be misunderstood; in this genus the ovarian tubes enter the walls, extending up between the longitudinal septa, or, as they may here be more naturally called, the radiating septa. I will specify a few more peculiarities worthy of remark: — in some species of the sub-genus Acasta, clefts are left, covered only by membrane, on the lines of suture (Pl. 9, figs. 7 a, 8 a), between the compartments, just above the basis; and in other species the basis is perforated by numerous membrane-covered, minute orifices. In Platylepas, each compartment has one deep inward fold (Pl. 17, fig. 1), somewhat analogous to the three folds in Coronula; this fold is produced into an internal midrib, supporting and rendering convex the membranous basis; in this genus, also, the rostrum, owing to its midrib, is generally thrust a little on one side, and the shell thus rendered asymmetrical. In Chamæsipho scutelliformis the shell is symmetrically perforated (Pl. 19, fig. 4 a) by four apertures. Lastly, in Chthamalus Hembeli and intertextus, after a certain age, the basal edges of the walls become inflected, and continue to grow inwards till they entirely take the place of the true membranous basis.
Structure of the Radii.
Radius. — This term, as we have seen, is applied to that side of the compartment, the growth of which is modified, by abutting against and overlapping the adjoining compartment. Hence the structure of the radius is essentially the same with that of the parietal portion of the compartment. When best developed, as in Balanus tintinnabulum, the radius consists of an outer and inner lamina, separated by denticulated septa, extending in horizontal lines parallel to the basis, and is consequently perforated by minute tubes or pores. The tubes become filled up solidly much more commonly than do the parietal tubes; and the inner lamina, in such cases, is hardly distinct from the outer lamina. The denticuli often fail, or are present only on the lower sides of the septa; and very frequently the edge of the radius can only be said to be crenated. Notwithstanding these frequent anomalies, if a series of species and genera be taken, it is certain that there is, as might have been expected, a close relationship in internal structure, between the radii and the parietes. The edge of the radius is received in a slight furrow (generally marked like a seal, with the impression of the denticulated septa) in the opposed compartment: sometimes the outer edge or lid of the recipient furrow, is so broad as to give the false appearance of a radius having been developed, at least in the lower part of the shell, on both sides of the suture. A crest of corium runs into each suture between the edge of the radius and the furrow in the opposed compartment; and when the radius is permeated by pores (as in woodcut 10), threads of corium branch off this crest, and enter the pores. In the lower part of the shell, these crests of corium project from the corium forming and surrounding the sack; but in the upper part of the shell, above the opercular membrane, and therefore above the sack, the corium is produced up each line of suture as a separate ribbon. In proportion as these ribbons extend more or less near to the summit of the shell, so do the edges of the radii continue to be added to, to a greater or less height from the basis; and consequently their summits become less or more oblique.
Fig. 10.
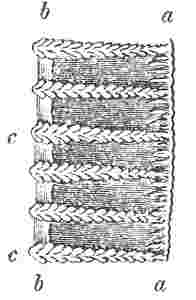
Edge of the radius of Balanus tintinnabulum. a, outer lamina; b, inner lamina; c, denticulated septa, uniting the two laminæ.
Peculiarities in the Structure of the Radii. — In some of the species of Tetraclita, in which genus the walls consist of several rows of tubes, the radii are likewise perforated by several rows; and in some of the other species (Pl. 10, fig. 1 h), the edge, or disarticulated surface of the radius, is marked by irregularly branching ridges; and these evidently correspond with the branching septa or ridges of the wall. In Chelonobia, the outer lamina of the radius, as well as of its recipient furrow, is of extraordinary thickness; and this lamina, in C. testudinaria (Pl. 14, fig. 1 a, 5, b, and Pl. 15, fig. 1, f), is modelled into sharp transverse ridges and valleys. In the Chthamalinæ, the radii, like the parietes, are simply solid; and apparently in consequence, for the sake of strengthening the sutures, the edges of the radii, and of the recipient furrow in Octomeris (Pl. 20, fig. 3 a) and in Chthamalus dentatus and Hembeli (Pl. 18, fig. 3 b, 5 a), are neatly dentated. In some other species of Chthamalus (Pl. 19, fig. 1 a), the radii present a slight modification of this structure, the sutures being formed by oblique interfolding laminæ. In the radii of Coronula and Tubicinella, there is a peculiarity, in apparent connection with the fact, that in these genera the parietal tubes are not crossed by transverse calcareous septa, namely, that the pores by which the radii are permeated keep unclosed throughout their length, and open into a special longitudinal tube (Pl. 16, fig. 7, d′), which runs along that margin of the wall, whence the radius arises. In Coronula the wall is of extreme thinness, and in conformity so is the true radius, but that the shell might not thus be rendered very weak, complementary or pseudo-radii are developed on their inner sides (Pl. 16, fig. 7, adjoining the true radii A d, C d, and shaded by distant convex lines). Even in the allied genus Xenobalanus, in which the whole shell tends to become rudimentary, traces of these pseudo-radii (Pl. 17, fig. 4 b, d) can be detected. In Coronula, though the radii (Pl. 16, fig. 7, A d, C d) are, by the above special means, rendered thick, and though the alæ also are thick (C a′, D a′), yet together they do not equal in thickness the folded walls; and consequently, there is left between the radii and alæ square chambers (v), occupied by the branching ovarian tubes.
Structure of the Alæ.
These project, generally abruptly, from the sides of the upper part of the compartments; they appear from the first growth of the shell; they are overlapped by the radius and by part of the wall of the adjoining compartment; they are thinner, and have, owing apparently to being overlapped, a very different aspect from the parietal portion; but they do not differ from it in essential nature. They are solid, that is, they are never permeated by pores; but their edges are generally crenated, and there is, in some cases, as in Chelonobia, sufficient evidence that these crenations answer to the horizontal septa on the edges of the radii (also often reduced to mere crenations), and consequently, likewise, to the longitudinal septa of the parietes. In Coronula the edge of each ala consists of a medial ridge, sending off denticulated septa on both sides, and is therefore anomalous as compared with the alæ in other genera, but corresponds in structure with the similarly anomalous radius of Coronula. In order to allow of the growth of the edge of the ala, a fine thread of corium runs up the narrow furrow in which the edge is lodged, proceeding from the corium of the sack. In proportion as this thread runs up higher or lower, so are the summits of the alæ rendered, during growth, less or more oblique.
Structure of the Sheath.
As the compartments overlap each other, the edges of the alæ would have projected, and the inner surface of the orifice of the shell would not have been smooth and rounded, had not that part of each wall, which does not overlie an ala, been thickened so as to allow of the formation of a shoulder or indentation, against which the edge of the ala fits and abuts. The thickened portions, and the alæ themselves, together form the sheath, of which the use seems to be to strengthen, like a broad internal hoop, the upper part of the shell round the orifice, where naturally it is weak. The sheath is composed of successive, fine, shelly layers, which extend, as the shell is added to at the basal margin, lower and lower down on the inner surface of the walls. The lower edge of the sheath either simply projects a little inwards, or more commonly is formed into a sharp depending ridge, as represented in fig. 1, K′, Pl. 25. In some species of Pyrgoma (Pl. 13, fig. 2 b), the sheath reaches nearly to the bases of the compartments; and in Chelonobia (Pl. 14, fig. 4 e c e), the inner layer of shell surrounding the sack, which seems to correspond more nearly to the sheath than to the inner lamina of the walls, actually rests on the basal membrane. The opercular membrane is generally, but not invariably, attached only a little way above the lower edge of the sheath: at each exuviation, a new opercular membrane is formed, and is attached to the next lower zone of the sheath; the old membrane being cast off, but a circular slip of it is left, investing the last zone. Hence the whole upper part of the sheath above the opercular membrane, comes to be thus invested; and is marked by circular lines, one above the other, caused by the successive exuviations. This investing membrane often supports rows of minute bristles, directed upwards. Generally, a film of shell is deposited, at the period of the formation of each new opercular membrane, on that part of the sheath which lies immediately beneath. This innermost film or thin layer of shell, on the lines of suture between the compartments, breaks joint, at least in some cases, with the underlying shelly layers, — that is, the suture in this last-formed film does not lie exactly over the suture in the subjacent layers of the sheath. In Tubicinella, the sheath extends down close to the basis; and what is unique in this one genus, the opercular membrane, gradually thinning out downwards, closely adheres to the whole inner surface of the shell. In Tubicinella and in Xenobalanus (Pl. 17, fig. 4 b), the sheath separates easily into separate successive rims of shell; and this structure evidently is for the sake of facilitating the breakage of the upper end of the shell, which, as we shall presently see, is necessary to allow of the increase in size of its orifice.
Structure of the Basis.
This, in several genera and species, is composed of simple membrane, and consists of successive, concentric, circular slips, added round the outside, at each period of growth. In some species of Tetraclita and Balanus the basis is calcareous, but diaphanous, very thin, smooth, or somewhat granulated. In other cases it consists of a single calcareous lamina, either smooth, or with ridges radiating from its centre; it is formed of two laminæ, (as is most usual in Balanus,) separated by radiating septa. These septa, as well as the radiating ridges in the case of the single lamina, are homologous with the longitudinal septa of the parietes. The denticulated ends of the latter enter the mouths of the tubes formed by the radiating septa of the basis: threads of corium pass between the denticuli of the parietal septa, and thus enter the basal tubes. The ends of these threads of corium generally deposit transverse calcareous septa, exactly as within the parietal tubes. When the basis is thick the septa themselves (ccc) between the proper basal tubes, become porose, (or rather cancellated,) and they sometimes expand into a very thick, cancellated layer, separating the outer lamina (a) of the basis from the proper basal tubes, which always lie close under the inner lamina (b). This structure differs only slightly from that seen in the parietes of Tetraclita, in which the branching of the longitudinal parietal septa, produces thick walls, formed of several rows of tubes or pores. With respect to peculiarities in structure of the basis, Balanus lævis offers the most remarkable case; for here, in specimens which have grown crowded together, the whole interior appears sometimes to have become too much elongated or too deep for the animal’s body, and consequently the lower part of the deeply-concave basis has been filled up (Pl. 4, fig. 2 a) by thin, irregular, calcareous diaphragms. In elongated specimens, also, of Balanus balanoides, the shell sometimes appears to have grown too long for the animal’s body; but in this case the membranous basis becomes extremely convex inwards; it still reaches the basal edges of the parietes all round, but in the middle it is raised high above the surface of attachment; yet sometimes threads of the cementing tissue depend from the middle part to the surface of attachment. In Balanus terebratus (Pl. 8, fig. 2 a, 2 b), and in some species of Acasta, the basis is riddled, as previously stated, by numerous, minute, membrane-covered orifices. In B. declivis the membranous basis is always extremely oblique, owing to the rostral end of the shell being twice as high as the carinal and opposite end.
Fig. 11.
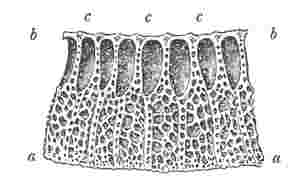
Portion of edge of basis of Balanus tintinnabulum, a, a, outer lamina; b, b, inner or upper lamina; c, c, c, porose or cancellated radiating septa.
Regarding the very remarkable means by which the basis of sessile Cirripedes is cemented to the surface of attachment, it will be convenient to defer for a little the description, on account of its necessary length.
Structure of the Opercular Valves (Scuta and Terga).
These are situated on each side of the slit or orifice leading into the sack; from their shape, their powers of movement, their separation by flexible membrane from the shell, to which they serve as a lid, they appear at first as if they constituted an element very distinct from the shell itself, but this is not the case. They are, together with the opercular membrane, as essentially as the whole of what is externally visible, a part of the modified carapace, of which they occupy the upper or posterior extremity: from tracing the metamorphoses, or even by comparison of a Balanus with Pollicipes, there can be no doubt of the truth of this conclusion. The opercular valves are four in number, — a pair of scuta and a pair of terga; but the latter in Coronula diadema and reginæ, are either aborted or represented by a mere rudiment; and in Xenobalanus both scuta and terga are quite absent. In several cases, more especially in the genus Pyrgoma (Pl. 13, fig. 1 b), the scutum and tergum on each side are calcified together, so that sometimes not even a trace of the line of junction can be discovered. In most cases the scutum is firmly united, being articulated in a manner presently to be described, to the tergum; but in Coronula, Tubicinella (Pl. 17, fig. 3 c), and Platylepas, the ends of these valves are simply approximated.
Scuta. — These valves are important, inasmuch as the animal’s body is attached to them; in Pl. 25, fig. 1, the broken line, surrounding a, b, shows where the body has been cut, in removing the scutum on the near side, the other scutum, S, being left articulated to the tergum, T. In shape the scuta are generally sub-triangular; but in some species of Pyrgoma and in Chelonobia, &c. they are much elongated. The lines of growth are usually prominent; and along the occludent margins the alternate, or sometimes every third or fourth line, is developed into a knob, which produces a serrated edge, serving to lock the two opposed valves together; there is, however, no trace of this structure in Coronula and Tubicinella. In some species of Pyrgoma, a ledge of considerable breadth (Pl. 13, fig. 3 e, &c.) is developed along the occludent margins of the two scuta, as well as of the two terga, giving them an anomalous structure. The Terga differ considerably in outline in the different genera and species: their shape approaches more nearly to a triangle than to any other regular form; but there is generally a projection or spur on the basal margin, on the side towards the scutum. In some species of Pyrgoma, the tergum is of so irregular a shape as to defy description. In most cases, a longitudinal depression or furrow runs down the valve, from the apex to the extremity of the spur; and it not rarely happens that the sides of this furrow become folded inwards and almost closed. The spur probably answers to the basal point of the usually sub-rhomboidal tergum in Pollicipes and Scalpellum. The tips of the terga in some species of Balanus, &c., are specially modified into sharp points or beaks (Pl. 2, fig. 3 b, 3 d), bowed a little inwards, and projecting considerably above the tips of the scuta; this is effected by the medial, uppermost part of the valve being internally thickened and hardened, and then, by the disintegration of the two margins and the external surface, the internal modified portion becomes exposed. The whole valve, also, at least in such cases as in Balanus psittacus, appears to be forced slowly upwards in the articular furrow of the scutum. I am assured, by a competent observer, that the beaks of the terga in B. porcatus can give an object placed within the orifice of the shell a sharp tap.
In comparing the Tergum of one of the Balanidæ with that of a typical member of the Lepadidæ, for instance, that of Balanus with that of Pollicipes, apex corresponds with apex: the extremity of the spur in Balanus corresponds with the basal point of the whole valve in Pollicipes: the scutal margin, (which in Balanus homologically extends down to the extremity of the spur), corresponds with the scutal margin of Pollicipes: the carinal margin in Balanus corresponds with the upper carinal margin in Pollicipes: the basal margin of Balanus on the carinal side of the spur, corresponds with the lower carinal margin in Pollicipes: lastly, (and this is the chief difference), in Balanus there is no appreciable occludent margin, the apex of the valve being brought close to the upper angle of the scutal margin; in Chthamalus, however, there is yet left some remnant of an occludent margin, — which margin in Pollicipes is conspicuous.
The scutum and tergum, with the few exceptions above stated, are articulated together at a large or open angle. The articulation (see Pl. 11, fig. 5 b, c, d, and fig. 6 b, c) is effected by the margin of the tergum being a little inflected, and lodged in a furrow in the margin of the scutum. This furrow in the scutum has its further border generally prominent and often reflexed or curved over; I have called it the articular ridge; it, also, is lodged in a furrow in the upper part of the tergum, which again is bordered by a ridge, viz., the articular tergal ridge. So that in both scutum and tergum there is an articular furrow, bordered in each case, on one side by the margin of the valve, and on the other side by the so-called articular ridge. In Chelonobia (Pl. 14, fig. 1 b) the articular ridge of the scutum is horny. When, as often happens, the scuta and terga have been much worn, the manner of their articulation (Pl. 18, fig. 1 a) is pretty well shown even from the outside; in this case their external appearance is very different from what it is in those individuals (fig. 1 c) of the same species, which have not suffered disintegration. This articulation of the scuta and terga is prefigured amongst the Lepadidæ, in Pollicipes mitella, and in Lithotrya.
The scuta are brought together by a short, strong, straight, adductor muscle (Pl. 25, fig. 1, a); its attachment leaves (with very few exceptions, as in Tubicinella) a rounded impression, or even pit, on the under side of the valve in its upper part. This pit is frequently bounded, on its lower side, by a sharp ridge, which, though not in actual connexion with the adductor muscle, I have, for convenience sake, called the adductor ridge; it serves apparently to give support to the animal’s body; in some few cases (as in B. psittacus, Pl. 2, fig. 3 c) it is confluent at its upper end with the articular ridge, and converts the whole basi-tergal corner of the valve into a deep cavity. In some of the species of Pyrgoma (Pl. 12, fig. 5 c, 7 b), and in some varieties of Creusia, this adductor ridge is enormously developed, so as to depend far beneath the true basal margin, or that to which the opercular membrane is attached. At the basi-tergal corner of the valve, there is generally a small pit or impression, and sometimes distinct crests, for the attachment of the lateral depressor muscle. At the rostral end there is, also, a small cavity formed by the overfolding of the occludent margin (rarely furnished with crests) for the attachment of the rostral depressor muscle. In the Terga, at the basi-carinal corner, there are usually crests, though sometimes feebly developed, for the attachment of the tergal depressor muscle. But in Chelonobia, Coronula, Tubicinella, Platylepas, and in some other cases, there are no crests. The crests, when well developed, are furnished with rectangular sub-crests or denticuli on both sides; in fact they resemble, and are probably homologous with, the denticulated ribs or septa in the parietes, radii, and basis. Altogether the scuta and terga are attached, as far as muscles are concerned, to the shell and sack, by three longitudinal pairs.
Growth of the Whole Shell, and Its Microscopical Structure.
The opercular valves are added to along their basal margins alone; the animal’s body, together with the several muscles, becoming attached at each period of growth lower and lower down to the valves; this no doubt is effected by the absorption of the upper surfaces of the muscles, and the formation of new fasciæ on their lower surfaces. The opercular membrane, which, though thin and flexible, forms part of the general outer surface of the animal as much as does any portion of the rigid shell, with which indeed it is strictly homologous, is periodically moulted, together with the integuments of the whole included animal. The new opercular membrane is of course each time formed a little larger than the old one. In Coronula and Tubicinella, however, several successive opercular membranes are preserved one over the other, and the outside membrane gradually disintegrates; in these cases the undermost opercular membrane is formed wrinkled and considerably too large, so as to allow of being stretched, before it is finally cast off. In Tubicinella, the opercular membrane runs down, adhering to the inner surface of the shell, to nearly the basis, and hence during the diametric growth of the shell, it is longitudinally split, and is repaired by slips of new membrane, which resemble the radii in form and in direction of the lines of growth.
In some species of Pyrgoma, the ledge (limbus occludens) which is added along the occludent margin of both scuta and terga, and in some species of Balanus a narrow rim, or slight protuberance which is added along the carinal margin of the terga, offer unimportant exceptions to the rule, that the opercular valves grow only at their basal margins.
The basis is added to only round the circumference, and hence increases in diameter, and, when concave, in depth. The compartments grow at their basal margins, where they are in contact with the basis; hence the shell is added to in height, and, owing to the outward inclination of the compartments, also, in basal diameter; but the compartments likewise, in most cases, grow along both lateral margins, that is, on the edges of the radii and alæ; and hence the upper part of the shell, also, increases in diameter. The orifice of the shell, moreover, thus becomes enlarged. In some cases the shell is destitute of radii, only sutures being present, that is, the compartments do not grow laterally; and sometimes, as in the whole genus Pyrgoma, there are not even sutures, the compartments having been fused together: in both these cases, the shell can increase in diameter only at the base; and the orifice, it might have been thought, would necessarily have remained, to the destruction of the animal, of the same minute size, as when first formed after the metamorphosis: this certainly would have been the case had not the upper ends of the compartments, surrounding and forming the orifice, been nicely adapted always to yield, in a certain limited degree, to the disintegrating influences to which every shell is exposed, but which most Cirripedes can resist; and the disintegration of the narrow end of a conical tube, of course increases the diameter of its orifice. In Tubicinella, in which the shell is furnished with narrow radii, and does increase in diameter from top to bottom, the increase is not sufficient in proportion to the continued elongation of the shell; to compensate for this, the orifice is enlarged at short intervals by the breakage of the upper end of the shell, for which purpose (as explained under the genus) it is evidently constructed. Hence we see that, in certain Cirripedes, decay or disintegration, and breakage, are necessary elements in their growth! It is a remarkable fact, which I cannot explain, that in some species in which the orifice of the shell is usually increased by disintegration, if individuals are so situated that they are not exposed to sufficiently energetic disintegrating influences, as may be inferred from the well-preserved condition of the whole surface of the shell, then the radii become developed, and the orifice is increased in size by the diametric growth of the upper part of the shell: I have seen instances of this in Tetraclita porosa, and purpurascens, and in Balanus perforatus: it appeared, but of course erroneously, as if the lateral growth of the compartments had been subjected to the will of the animal.
Considering the strength of the shell of sessile Cirripedes, the separation of their compartments one from another and from the basis, during growth, has justly been thought a surprising circumstance. In most Chthamalinæ and in some species of Balanus, however, if the shell be boiled in caustic potash, the compartments fall apart with a touch; this shows that their union is due to animal and probably to organised matter, and the growth of such matter between the opposed edges of the compartments, and their consequent gradual separation, offers no particular difficulty. But in many Balani, boiling in potash for hours does not seem even to weaken, in the least degree, the sutures, which are wonderfully strong — the shell often breaking rather than yield on these lines; if, however, the shell be dissolved in acid, the animalised tissue which is left easily separates on the lines of suture, and if this tissue be boiled in potash, the remnants of the compartments fall quite separate. These facts seem to me to show, that the compartments in such cases are joined along the lines of suture by tissue, which must be in a calcified state, but which, nevertheless, continues to grow by intersusception; in other words, I believe that the tips of the complicated ridges and points interlocking on the lines of suture, are not separated from each other by films of corium or simple animal matter, but are actually united by corium in a calcified, yet still growing condition.
In ordinary Crustaceans, the growth is periodical and sudden; a new and larger carapace, for instance, is formed under the old one, and after the exuviation of the latter, the new one soon hardens, and does not subsequently increase in size; so it is in the case of Cirripedes, with the membranes of the body, and even with certain parts, as the opercular membrane, of the external covering. But a Cirripede cannot, like a crab, crawl into some crevice and remain protected till its shell becomes hardened; hence, probably, it is that the shell is never wholly moulted. Even if the margins of the opposed compartments and of the basis were to grow rapidly, the shell would necessarily be much weakened on the lines of suture, and unable to withstand the heavy breakers, to which so many species of sessile Cirripedes are exposed. On the other hand, although the margins are thus compelled to grow slowly, they do not grow continuously, as may be seen in the zones of increment on all the valves, corresponding, I believe, with the periods of exuviation of the membranes of the body. A layer of shell, often very thin, seems to be generally deposited over the whole internal surface of the several valves, at the same time that the marginal zones are added; so that the only essential difference in the growth of the external covering, in Cirripedes as compared with ordinary Crustaceans, is that the old shell is not cast off, but adheres to the outside of the new shell, and that the margins are added to (in certain definite directions) slowly yet not continuously, instead of the whole being formed at a single period.
In the genus Alcippe, and in Cryptophialus, the whole of the external membranes are moulted, excepting the surface of attachment; but then these Cirripedes live in cavities which they form for themselves, and are thus protected. In Lithotrya the membrane of the peduncle, with its little valves or scales, is moulted, but here, again, this very part is protected by the tubular cavity, which the animal forms and inhabits. Neither of these three genera belong to the Balanidæ, or sessile Cirripedes, which we are now more especially describing.
If, now, a section of one of the shelly zones of growth be carefully examined, it can in some cases be distinctly seen to be formed of successive, excessively fine laminæ; but the animalised tissue (which differs much in amount in different Cirripedes) left after the shell has been dissolved in acid, exhibits, in most cases, neither laminæ nor any other structure whatever. The shell seems to be the actual pulpy corium, or true skin, in a calcified condition, but generally with its cellular structure modified and much reduced: I have taken a bit of recently-formed shell of Tetraclita and of Coronula, with the corium still adherent to its under surface, and after dissolution in acid, I could not distinguish the part, which had just before existed as shell, from the corium itself. In the case of Coronula, immediately prior to the period of moulting and growth, I found the unaltered corium so charged, as to effervesce, with carbonate of lime, either in a state of dissolution, or in granules too minute to be visible under the highest powers.
The sutures between the several compartments and the basis are covered by thin membrane, which is continually splitting during the growth of the opposed edges of the underlying shell; but previously to each splitting, a new slip of membrane is, I believe, already formed under the old one; so that the corium is not even momentarily exposed. Owing to this manner of growth, the slips of membrane consist of successive rims united together; in most cases, these soon become abraded from the older parts of the shell, but are sometimes preserved. The last-formed slip of membrane over a suture is homologous with the opercular membrane; and both are strictly analogous with the ring of flexible membrane, forming the joint of the leg of a crab. In the latter case, the flexible membrane and hardened crust are both moulted together: in the opercular membrane, there is a double line of splitting, one close round the opercular valves, and the other at the basal edge of the sheath, and the intermediate portion is moulted, but with a zone of membrane left adherent to the non-moulted valves and sheath: lastly, in the slips of membrane covering the sutures, there is only a single line of splitting, and no portion, I believe, is moulted; the rims of membrane on each side remaining adherent on the compartments and basis, until worn away.
The opercular membrane, when closely examined, exhibits no structure, except that it can sometimes be plainly seen to be composed of successive, numerous, excessively thin laminæ. Occasionally, however, it presents the false appearance of being permeated by parallel and anastomosing vessels: this appearance is clue to one or more of the component laminæ having been wrinkled before a succeeding lamina was thrown down and attached to its under side. If a small piece of an opercular valve of Tubicinella, with the opercular membrane adhering to it, and with the corium under both, be dissolved in acid, it may be clearly made out that the corium under the valve has gone on being converted into shell, whereas under the opercular membrane it has been converted and condensed into fine constituent laminæ of chitine. Inasmuch as the successive layers of shell, during each period of growth, go on encroaching on those of the membrane, the line of junction between the shell and chitine becomes oblique or bevelled. The membrane on this bevelled line of junction assumes a slightly different aspect to what it has elsewhere; it becomes yellowish or brown, thicker and very much tougher. In many genera it is also furnished with a row of small bristles. At the period of exuviation the opercular membrane separates just outside this modified portion, leaving the latter adherent, as a rim or slip, on the valves. If, however, the opercular membrane be rudely torn off before its proper period of exuviation, it carries with it the as yet continuous, but already modified, slip. A slightly indented line may sometimes be traced before the period of exuviation, showing where the separation will take place: what produces this line I know not. The coloured, thickened, and modified slips of opercular membrane, which are thus retained adhering to the valves, and which together form an investing membrane, have been considered by most authors as the epidermis; but they have no more right to be thus called than has any other part of the opercular membrane. Exactly similar slips of membrane are left investing the sheath. So, again, the membrane which, when well preserved, invests the walls of the shell, is made up, as already stated, of successively adherent slips, which originally covered the lines of suture.
In the case of Coronula there is a peculiarity, described in the last section of this Introduction, (under the head of Cementing Apparatus), namely, that the two or three last-formed, exterior zones of the Basal membrane continue for a period to increase in width; being, as I believe, dragged one from over the other, with fresh laminæ of membrane continually thrown down. In this same genus, and in Tubicinella, the walls of the shell are invested by membrane, which is doubled inwards under their basal edges; and as the latter grow, the investing parietal membrane splits and separates from the basal membrane, and is pulled outwards and downwards. This inflected, often broad border of membrane, seems to me more strictly comparable with the opercular membrane, than with those narrow, thickened rims of yellowish membrane which in other Cirripedes cover the suture between the basal edges of the walls and the basis.
The little bristles above alluded to, which arise from the slips of membrane left adherent on the opercular valves, sheath, and walls, stand in rows; a row corresponding to each period of exuviation of the opercular membrane. The bristles are generally largest on the opercular valves and sheath; in Balanus tintinnabulum, they are from 1 to 2/1000ths of an inch in length, but they are longer in some other species. I may here mention, as showing the connexion of these bristles with the opercular membrane, that similar bristles occur in B. perforatus, scattered over the surface of that membrane, and are necessarily moulted with it. In the imbedded genera Coronula and Tubicinella, none of these bristles exist. When a portion of valve or shell, furnished with bristles, is dissolved in acid, tough, sinuous, and apparently hollow, threads are seen to run from their bulb-like bases, into and up the corresponding layer, which, before dissolution, existed as shell; and they terminate internally in very fine points, which I believe are united to the underlying corium. These threads, or tubuli, as I have called them in my volume on the Lepadidæ, are, in Tetraclita porosa, about 1/5000ths of an inch in diameter, but only half that size in B. tintinnabulum. On parts of the shell where there are no bristles, similar tubuli penetrate the shelly layers, and come to the surface. The tubuli running to the lowest and last-formed row of bristles, just after a period of exuviation, are so delicate as hardly, or not at all, to be distinguished; in the row above, they are plain and longer, and for the next two or three upper rows they are, in some cases, as in Tetraclita porosa, longer and longer, having been added to during each successive thickening of the valve. These tubuli consist of chitine, and no doubt first existed as threads of corium; they are so tough that they must serve to strengthen the successive layers of shell, but I imagine their chief function is to keep up the vitality of the newly-formed layers of shell. May we not, also, venture to suppose that by their means, some degree of sensibility is given to the bristles? I need only further remark, that in some species of Balanus and of Chthamalus, the under side of the shell is penetrated by irregular pores, large enough to be visible to the naked eye, into which threads of corium penetrate; but these can hardly be said to appertain to the microscopical structure; and are more nearly related to those pores and furrows, formed by the greater or less development of the longitudinal septa, and in which the threads of corium deposit, or rather become changed into, transverse septa, or solid shelly matter, as previously described.
I regret that I have used this term “tubuli”; for the threads thus designated, I believe, are not the same with the tubuli of Dr. Carpenter, which are not left after dissolution in acid. I have seen tubuli, as called by me, in the shell from the leg of a crab, after having been placed in acid.
Muscles of Sack.
In the pupa, the thorax, as we shall hereafter more fully see, is continuous with, and opens into the large anterior end or front part of the head; but during the metamorphosis (Pl. 30, fig. 2), the thorax of the Cirripede becomes, owing to the almost transverse position occupied by the young animal within the pupa, to a great extent internally separated from the anterior end, — which anterior end forms, as we know, either the peduncle or the basis. Hence it comes to pass that the body or Thorax (Pl. 25, fig. 1) is lodged within a sack (f) within the shell. The chitine membrane lining this sack is excessively thin and transparent, but less so in Xenobalanus and Tubicinella; it is obviously continuous with that investing the body of the animal; it is also essentially continuous with the opercular valves and membrane, and consequently with the whole shell. It is periodically moulted. It is lined by corium, as is likewise the surrounding shell; hence the corium is double round the sack, as indeed might have been expected from the shell and opercular valves (at least their upper parts) being formed by the prolongation, as is obvious in the pupa, of the posterior edges of the carapace. Between the two folds of corium, which are united together by transverse ligamentous fibres, branching out at both extremities, like the roots and branches of a tree, we have the longitudinal muscles, which go to the opercular valves; and likewise a layer-like mass of branching ovarian tubes (Pl. 25, fig. 1, g): the ovarian tubes, however, are often confined to the base of the sack. In Xenobalanus, the two folds of corium are united by longitudinal membranous septa, making a series of quite peculiar, square tubes.
The above-mentioned muscles are attached at their upper ends to the opercular valves, and at their lower ends to the basis. There are, in fact, three pairs, but the pair attached to the basi-carinal angles of the two terga (Pl. 25, fig. 1, i), are almost invariably confluent, forming one great bundle; the second pair is attached to the lateral or basi-tergal corners of the two scuta, and are hidden in the figure; the third pair (h) is attached also to the scuta, to their rostral angles. These muscles can only act as depressores; they are often extremely powerful; they belong to the voluntary class, for they are transversely striped. By their action, the opercular valves are capable of varied slight movements, within the limit allowed by the width of the flexible opercular membrane. By the action of the lateral scutal depressores, the orifice leading into the sack is opened, the movement being generally aided by the protrusion of the cirri. By the sudden contraction of the rostral scutal depressores, the blows which are sometimes given by the beaked terga at the opposite end of the operculum, are probably effected. By the contraction of all three pairs of muscles, the opercular valves are held down with quite surprising force. The valves can be raised only by the action of the animal’s body against the basis.
In Coronula these muscles are more spread out, and do not extend down to the basis; their lower portions, as is likewise the case in Tubicinella, do not exhibit transverse striæ, and hence tend to pass either into the involuntary class, or into ligament. This condition of the muscles, in the above two genera, accords with the little-developed state of their opercular valves. In Xenobalanus, there is no longer any evidence of the muscles being collected into five or six bundles, for they are thinly and almost uniformly spread out, and show in no part transverse striæ. I may add that in much elongated specimens of Balanus balanoides, these muscles become in their lower part ligamentous, and destitute of striæ.
Branchiæ.
In the Balaninæ, a pair of Branchiæ is always present: they lie on each side, in a somewhat curved position, in the angle between the sides of the shell and the basis. In Pl. 25, fig. 1, they are exactly covered, on the further side, by the body of the animal. They are attached near each other at the carinal end of the sack in a vertical line, and likewise on each side in a transverse line, extending from close beneath the spur of the tergum towards the point of attachment of the body to the scutum. In Balanus, as in the figure (Pl. 25, fig. 3) of B. tintinnabulum, each branchia consists of a medial fold of skin, a little curved conformably with the sack, and slightly tapering towards its rostral and free extremity; but this fold is almost hidden by the vertical sub-folds or membranous ridges, themselves plicated and sub-plicated, which project on both sides: these vertical folds are free at their tips: at their lower attached ends, they are thickest. On the side nearest the wall of the shell, the whole branchia has a bilobed appearance, owing to a very deep indentation caused by the projection of the scutal lateral depressor muscle; the sub-folds on this side are also more plicated. The branchia essentially is an inward plicated fold of the membranes of the sack; for its outer, very thin tunic is continuous with and moulted with that lining the sack; and within it we have two layers of delicate, pulpy, transparent corium, united together (as is best seen in Coronula) by ligamentous fibres, branched at their two ends, all exactly as in the corium surrounding the sack. There are here no distinct vessels, any more than in other parts of the body, but a fluid could easily circulate in the interspaces of the corium. From the large size of this organ, and its simplicity of internal structure, being adapted exclusively to expose a great surface of skin to the water, I do not doubt that it has been correctly considered as a respiratory organ. By the voluntary movements of the opercular valves (i. e. part of the carapace) the water is constantly being pumped in and out of the sack; the movement, indeed, may be almost compared to the heaving of a man’s chest. Moreover, the branchiæ on each side are attached so closely to the spur of the tergum, that each time the latter is moved, the whole branchia must, I think, be agitated, and the folds opened, as by the action of a lever.
In our two commonest, tidal, sessile Cirripedes, viz. Balanus balanoides and Chthamalus stellatus, I have observed that, when left uncovered by water, they kept the orifice of their operculums a little open, with a bubble of air within their sacks, so that the orifice was in fact closed by a thin septum of water, with air beneath; when disturbed, they closed their operculums with force, and expelled the bubble of air with a clicking noise, which has been noticed by Dr. Coldstream, and has been thought to be made by the movement of the operculum itself. Bal. crenatus, a deep-water species, when out of water, keeps its operculum closed.
‘Encyclopædia of Anatomy and Physiology;’ article Cirrhopoda.
In Coronula, Platylepas, Tubicinella, and Xenobalanus, each branchia consists of two unequal folds, both plicated on both sides: in the two latter genera, they extend far down the deep and elongated sacks, and hence the area of surface altogether gained is extremely great. In most of the species of Chthamalus, the branchia consist of a small fillet barely plicated: in the allied Chamæsipho columna, they are rudimentary, forming a smooth little pouch only 1/100th of an inch in length: in Chthamalus scabrosus they are quite aborted, being perhaps represented by a slight hairy ridge; but in Chthamalus dentatus, and therefore within the limits of the same genus, the branchiæ (and this seems to me a singular fact) are large, each being composed of two plicated folds, as in Coronula. Tapering filaments situated near the bases of the cirri, such as those occurring in several species of the Lepadidæ, are not found in any sessile Cirripede; but I have observed nearly similar filaments, projecting upwards and inwards at the base of the sack, in several species of Balanus and in Coronula; those which I examined were simply occupied by delicate corium, and no doubt must aid in exposing a greater surface of corium to the circumambient water.
Burmeister has given a good figure (Tab. 2, fig. 10) of the branchiæ of Coronula, (but the two folds are shaded too unequally), in his ‘Beiträge zur Naturgeschichte der Rankenfüsser.’
In my former volume on the Lepadidæ, I have described the ovigerous fræna occurring on the two sides of the sack, to which the ovigerous lamellæ are attached by a peculiar glandular secretion: in the Balanidæ there are no ovigerous fræna, but the branchiæ just described are identical with the fræna in essential structure and in position; differing only in being placed a little nearer to the carinal end of the sack, and in being generally (but not always) larger and more plicated: seeing this, and that in Alcippe lampas, and in some species of Pollicipes, — the genus which comes nearest to the Balanidæ, — the ovigerous fræna are large and are destitute of glands, and have therefore lost their normal function of supporting the ovigerous lamellæ, I can hardly doubt that the branchiæ in the Balanidæ are the ovigerous fræna of the Lepadidæ in a modified condition; a transformation of function not greater than that of the swimming bladder of a fish into the lungs of the higher Vertebrata.
There is, I conceive, no foundation for the belief of some authors that the branchiæ of the Balanidæ are in any way connected with the ovaria.
Thorax and Body.
Parts of the body included within the shell or carapace. — These parts (Pl. 25, fig. 1) consist of the prominent mouth, and of the thorax (c′), with its largely developed portion, called the prosoma (c), and with its appendages. The abdomen is quite rudimentary, being represented merely by a small portion of membrane surrounding the anus, and sometimes inserted like a wedge between the inwardly inflected posterior thoracic segments; in only two genera (Catophragmus and Pachylasma), its nature is rendered somewhat plainer by supporting caudal appendages. The probosciformed penis lies folded under the thorax; and I believe (from what is seen in the anomalous genus Proteolepas), that it normally arises from the ventral surface of the terminal point of the rudimentary abdomen. The thorax is laterally compressed, the ventral surface being very narrow, with the bases of the cirri placed closely together. It consists, in appearance, of two very different portions; one a soft, more or less rounded bag, which I have called the prosoma; and the other, which supports the five posterior pairs of cirri, is narrower, invested with stiffer membrane, and is more or less distinctly composed of five segments. These segments (Pl. 26, fig. 8) on their dorsal and dorso-lateral surfaces, are generally driven like wedges one into the other, with their points directed anteriorly: on the ventral surface the articulations are transverse. The prolongation (e) of the thin membrane (a) surrounding the anus (b), that is, the rudiment of the abdomen, which sometimes carries caudal appendages, almost divides (in appearance, whether really I know not) the hindermost thoracic segment along the medio-dorsal line, into two parts. I have given the above drawing of these segments, but with the dorsal surface much flattened, in Coronula diadema; in most species of Balanus, however, the wedges formed by one segment being driven into another, are much sharper; on the other hand, in Xenobalanus they are nearly straight and transverse. The three posterior segments are always the most distinct; the two next segments are also distinct laterally, but along the dorsal surface they become, excepting in Xenobalanus and some few other cases, completely confluent. The greater distinctness of the posterior segments is conformable to what takes place in the higher Crustacea. The articulations between the segments are folded inwards, and are formed of thin membrane, which in some cases, as in Coronula diadema, forms a marked contrast with the much thicker, stiffer, and yellowish membrane of the segments themselves; in Balanus tintinnabulum, however, the whole membrane of the five thoracic segments is very thin, excepting small wedge-shaped portions along the medio-dorsal line. The infolded articulations between the segments supporting the three anterior pairs of cirri (at least in the Balaninæ), are much wider than those between the three posterior segments; the former segments, with their cirri, being consequently capable of being moved further apart from each other. Could there have been any doubt as to the distinctness and reality of the five thoracic segments, it would have been set aside by the arrangement of the muscles attached to them, as will presently be described. I need only add, that in many genera there are shield-like swellings at the exterior bases of the pedicels of the posterior cirri, which I for some time thought were the epimeral elements of the thoracic segments; but I now believe them to be parts of the pedicels of the cirri. The basi-exterior margin, moreover, of the pedicel of the third pair of cirri, in many species of the Balaninæ (Pl. 25, fig. 1), is produced as a plate, thickly fringed with fine hairs, half across the dorsal surface of the thorax; serving, apparently, as a brush to clean the sack, or to prevent the ingress of any intruding substance.
Von Siebold and Stannius, in their ‘Anatomie Comparée,’ tom. i, , and , (foot-note), consider the articulated probosciformed penis as an elongated abdomen; a view which, at the commencement of my examination, I was tempted to admit; but the position of the caudal appendages on the dorsal basis of the penis, suffice, I think, to show that this view is not correct; for these caudal appendages evidently correspond with those borne on the very extremity of the abdomen in the pupa. Nor, indeed, does the position of the anus accord well with such a view.
The soft, rounded, bag-like portion of the body, which I have called the prosoma, is usually separated by a notch from the five posterior thoracic segments; at its upper end it may be said to carry the mouth and first pair of cirri. The prosoma includes the main part of the stomach and the broad ends of the vesiculæ seminales. It is always clothed by very thin membrane, which in Chthamalus dentatus, is hairy. In Tubicinella and Xenobalanus, the prosoma is much elongated, being produced far down the deep sack. That the prosoma is mainly formed by a great development of that segment (homologically the second thoracic segment) which carries the first pair of cirri, is certain, and I should not have hesitated to have said that it was exclusively so formed, had not the first thoracic segment in the anomalous genus Cryptophialus been developed as a distinct and free segment, not attached to the carapace; showing that possibly in other Cirripedes, the dorsal half of this first thoracic segment may be concerned in the formation of the free prosoma.
Muscular System.
Attachment of the Body to the Shell. — The prosoma which carries the posterior thoracic segments, and in appearance the mouth, is the only part of the body which is attached to the general covering (Pl. 25, fig. 1), namely, to the opercular valves. Except through the continuity of the lining membranes of the sack, the body lies free within the walls of the shell. The area of attachment (shown by a sinuous broken line round a and b) extends from about the middle of the two scuta down to their basal margins. As these valves lie obliquely across the orifice of the shell, the animal’s body comes to be suspended almost in the middle of the sack. The two scuta, as we have seen, have the power of opening and shutting a little; and are brought together by the adductor scutorum muscle (a), which is generally very powerful. The body is attached to these valves, round and beneath the adductor, so as to hide it until one of the valves be removed. The attachment is chiefly effected by three pairs of widely expanded, superficial muscles, two pairs of which are spread over the flanks of the prosoma, and the third pair over its rounded (properly dorsal) surface, which lies close to the rostral compartment (A, fig. 1) of the shell. I should have stated, that my chief examination of the attachment of the body to the scutal valves, has been made on Coronula balænaris, and less closely on Balanus tintinnabulum. Within these three pairs of superficial muscles, there are (besides the adductor) no less than five other pairs; of these one long pair is attached at one end to the basal margin of the labrum (e), and at the other end, to the under side, near to the basal margin of the scuta: two other, shorter, parallel pairs of muscles are attached at one end to the interspace of membrane between the basal edge of the labrum and the adductor scutorum muscle, and at the other end, to the under side of the scuta, above the attachment of the first pair: the fourth and shortest pair curls close under the adductor, and is there attached at both ends beneath it. The action of these four pairs of muscles must be to draw back, from the orifice of the shell, the mouth, and that interspace of body between the basal margin of the labrum and the adductor muscle. This movement I saw in living specimens. The last and fifth pair of muscles is small, but of considerable length; it is a diverging pair, attached at the converging end, above and exteriorly to the adductor muscle; and at the diverging end, low down on the under side of the scuta; I am very doubtful regarding the function of this pair. Altogether we have seen that round and within the fleshy pedicel, by which the body is attached to the scuta, there are no less than eight pairs of muscles. The central space between these muscles is hollow, and here many lacunal channels seem to converge. These muscles receive nerves from the supra-œsophageal ganglions. The interspace above alluded to, between the basal edge of the labrum and the adductor scutorum muscle, occupies a very different position according as the animal’s body is protruded as far as it can be, or is retracted. It is homologically part of the third cephalic segment; and consequently the mouth ought to have stood posteriorly (i. e. above, in the position figured in Pl. 25, fig. 1) to this interspace; yet, in fact, when the animal is retracted within its shell, the mouth usually lies almost directly beneath this interspace and the adductor scutorum muscle.
Besides these muscles of attachment, the prosoma is furnished with several other muscles. There are superficial muscles running up towards the basal margin of the sides of the mouth; and other deeper muscles, to which, I presume, the movements of the mouth, as a whole, are due. The muscles moving the gnathites do not, as far as I could make out, extend beneath the basal edge of the mouth. There are, also, powerful muscles giving movement to the basal segments of the pedicel of the first pair of cirri. Again, there are superficial muscles running to the next succeeding thoracic segment; the anterior ends of which are separated by a clear interspace from the lower ends of the above-described superficial muscles, by which the prosoma is attached to the scuta. On each flank, moreover, but more deeply imbedded, are the long flexor and extensor muscles, presently to be described, running to the five posterior thoracic segments. The last muscle which I need here mention, is a deep-seated diverging pair, attached near the upper end of the stomach, on its ventral surface, and diverging from this point to the sides of the prosoma high up beneath the mouth. The probable action of this pair, as well as of the three superficial pairs of muscles by which the body is attached to the scuta, is to draw up the whole prosoma towards or from the orifice; and likewise to contract it firmly, so as to serve as a fulcrum for the movements of the five posterior thoracic segments, together with the cirri, which they carry.
The muscles of these five thoracic segments are numerous and powerful; they are also complicated, chiefly owing to the segments on their dorsal and dorso-lateral surfaces being driven, like wedges, one into the other. As far as I could make out, there are on each side three, superficial, dorso-lateral and lateral muscles (generally, if not always, destitute of striæ), which do not cross the articulations, but extend merely from articulation to articulation; and of which the function can be only to contract each separate segment, and consequently to open out the intermediate infolded articulations; the effect of this would be to separate slightly the cirri from each other, — more especially those borne on the two or three anterior segments, between which the infolded articulations are deeper or broader. There are other more deeply imbedded, powerful, long, dorso-lateral extensor, and ventri-lateral flexor muscles, attached at one end within the flanks of the prosoma, and at the other end to the successive segments of the thorax. The action of the former is to straighten and stretch out the thorax; of the latter, or ventri-lateral muscles, to retract it. In tracing these muscles, a fascia could be seen to become attached to a segment, and then this same fascia would run on to the next succeeding segment: the effect of this must be, that each segment can be retracted and protracted either from the prosoma as a fulcrum, or from the antecedent segment as a fulcrum: we have, also, seen that each segment can, by the agency of the superficial, non-striated muscles, contract itself. Hence these thoracic segments are capable of diverse movements, as was very evident when the shell of a living specimen was opened. By one movement in common, the whole five posterior segments could be drawn back, so as to become even partly imbedded in the prosoma: lateral, twisting or wriggling movements were also quite distinct: the three posterior segments seemed to be capable of less independent movements than the anterior segments; and I observed that the more powerful flexor and extensor muscles did not run into these three posterior segments. The cirri, of course, partake of the movements of the thorax; and in watching, in an uninjured specimen, the alternate, protruding, gracefully sweeping and retracting movements of the posterior pairs of cirri, it was evident that the thorax was the chief agent in their movement. Besides the muscles now mentioned, there are some immediately to be noticed, which extend from within the thoracic segments to within the pedicels of the cirri.
Movements and Muscles of the Cirri.
Although the cirri have not been described, it will be most convenient here to treat shortly of their muscles. Each cirrus consists of a pedicel, having a long basal and a short upper segment, supporting two multiarticulate rami. The lower segment of the pedicel can be drawn forward by an adductor muscle, attached low down within this segment, and crossing at right angles (at least in the case of the anterior cirri) the corresponding muscle of the opposed cirrus, on the central, ventral surface of the thorax. This segment can also be drawn back by a muscle springing from the dorso-lateral surface of the thorax, and running only a little way within the segment: I am far from sure that the lower segment does not possess other muscles. The short upper segment of the pedicel can be moved backwards and forwards, as I saw in living specimens, independently of the lower segment; this movement being best seen in the anterior cirri, which are much more often moved independently of each other than are the posterior cirri. The rami are capable, I believe, of being moved backwards and forwards as a whole, by the movement of the few lower segments, which are generally more or less confluent. They can, also, be curled up and uncurled by the combined movement of each separate segment. The uncurling seems to separate the two rami a little laterally. Each ramus, at least in the two or three anterior pairs, can be moved to a certain extent, independently of the other ramus of the same cirrus; and the few terminal segments, either of both rami or of one ramus, are often a little moved and curled (and this is especially the case with the long anterior ramus of the first pair), without the lower segments or the pedicel being moved.
The flexor and extensor muscles, which, as I believe, move the upper segment of the pedicel (a and b, Pl. 29, fig. 1), are attached at their upper ends to its basal margin, and are thus enabled to draw it a little way down within the lower segment, and so move it. The short flexor muscle (c), which is attached at its lower end within the upper segment of the pedicel, and the longer extensor (d), also, attached within this same lower segment, serve, I believe, to move the lower, partially confluent segments of each ramus as a whole. In the case of these muscles, and of those last mentioned, I am surprised that the extensors (b) and (d) are not attached nearer to the exterior and dorsal surface. Other muscles (e, f) attached at their lower ends within the upper segment of the pedicel, run up each of the two rami to their tips, with some of the fasciæ terminating within each segment: of these muscles, the outer one (f, f) appears to be the extensor, and the inner one (e, e) the flexor. But besides these, there are other short flexor muscles (g, g) which run on the anterior face, from segment to segment, serving to pull the front edge of one segment within the edge of the next lower segment. These muscles differ much in plainness in the several genera: they are very distinct in Coronula. In some specimens of this genus, a few of the articulations between the basal segments of the rami having been obliterated, the short muscles (g, g) running from articulation to articulation were absent, and their presence and nature in the upper segments thus rendered the plainer. The muscular system in the several pairs of cirri seems to be the same, with the exception of the first pair, in which the muscle answering, as I suppose, to (a), namely, the flexor of the upper segment of the pedicel, is much spread out at its lower end, and is there attached to the exterior surface of the lower segment.
For a considerable time I thought that there were muscles going to the spines, especially to those which arise from the upper dorsal edge of each segment; but I have since ascertained that these are the cases within which new spines, with their lower ends doubled like the fingers of a glove hastily pulled off, are in process of formation.
The backward and forward movements of the segments, both in the rami and in the pedicels of the cirri, are apparently effected, as already noticed, by the outer or inner (as the movement may be) basal edge of one segment being drawn a little way down within the next succeeding lower segment. If, at the same time, both the inner and outer margins of all the segments were drawn one within the other, the whole limb would necessarily be shortened; and I distinctly saw a shortening action, with very slight movement in any other direction, in the first and second pairs of cirri; and I think it almost certain that this movement might be performed by the other cirri. If I correctly understand a statement of Milne Edwards, this is an important fact, as he asserts that only the higher Crustaceans possess the power of shortening their limbs.
‘Annales des Sciences Naturelles,’ tom. xviii, 1852, .
When a Cirripede is alive, the action of the cirri is really beautiful: from the position of the thoracic segments, the posterior cirri (three pairs in the Balaninæ and four pairs in the Chthamalinæ) form a sort of semicircle facing the mouth: the anterior cirri stand further apart, and are opposed in pairs to each other, with the first pair pointing beyond the mouth. Together the cirri form a hollow cone, not circular but elongated, with the mouth situated at the lower anterior end. The posterior cirri are protruded, by the movement of the whole thorax, curled up, close along the carinal end of the orifice; as they are protruded, they diverge, both by the movement of their pedicels, and, as I believe, by the separation of the thoracic segments. As the two rami of each separate cirrus are uncurled, they also diverge a little; as do the double rows of spines on the segments in each ramus, by their elasticity. By the movement of the thorax, the cirri are then swept towards the rostrum; and, lastly, they are brought perpendicularly down towards the mouth with a rapid movement, which would be beautifully adapted to catch any object floating or swimming in the water; hence I have called the action of the cirri, captorial. When the shell of a Balanus is broken open, the second and third pairs of cirri are repeatedly clasped over the mouth with a convulsive movement, in a manner indicating, I think, that their chief function is to seize and carry to the mouth any object entangled by the sweeping movement of the three posterior pairs. The first pair is also well adapted to aid in this seizing action; but I suspect that the long anterior ramus likewise acts as an organ of touch, warning the animal of danger. The mouth being itself moveable as a whole, — the outer maxillæ being capable of a backward and forward sweeping action, and being furnished with orifices apparently olfactory, — the inner maxillæ having more diversified movements, — the toothed mandibles overhanging the œsophagus, — and the œsophagus itself possessing a powerful swallowing movement, are all admirably adapted to secure any prey, when once entangled by the cirri.
Mouth.
The mouth, in the sub-family Chthamalinæ, cannot be distinguished from that of the Lepadidæ, which has been pretty fully described in my former volume. In the Balaninæ, however, the labrum differs considerably in not being swollen; that is, in its outer and inner fold of membrane being close together, and in having a central notch: the palpi are also larger, and the lower teeth on the mandibles, are laterally (Pl. 26, fig. 5) double, as will be more particularly stated under these two sub-families. I have given a drawing (Pl. 26, fig. 1) of the mouth, seen from above, of Balanus perforatus, with the right-hand palpus (d′) and outer maxilla (a′) cut off, in order that the labrum (e), mandibles (c), and inner maxilla (b) might be better shown; the cut-off bases (x, x) of the first cirrus on each side are also shown. In fig. 2 we have the deep supra-œsophageal cavity in Bal. improvisus torn open and laid flat, with the inner surfaces of the labrum (c) and outer maxillæ (a) exhibited, the palpi, mandibles, and inner maxillæ having been removed. Figs. 3 and 4 will presently be referred to; they are parts of the mouth, with the muscles, &c. removed, of Coronula. The mouth differs extremely little in the different genera and species of the Balanidæ, much less than amongst the Lepadidæ. In the Balaninæ, the crest of the labrum is sometimes hairy, instead of having, as is usual, from two to six teeth on each side of the central notch: in Balanus improvisus (Pl. 26, fig. 2) and eburneus, and in Chelonobia, the crest on each side of the central notch (e′) is furnished with a row of finely graduated teeth. A sub-triangular portion of the inner fold of membrane of the labrum, which overhangs the œsophagus, is always thickened and yellowish; it is also often punctured in patterns (Pl. 26, fig. 2, f), which, I believe, give attachment to little muscles that serve to open the upper end of the œsophagus. Opposite to this thickened, sub-triangular portion of membrane, the thin membrane forming the supra-œsophageal cavity (or the cavity surrounded by the gnathites) is strengthened by a pair of curved ribs (h, fig. 2) of thickened yellowish membrane, running down from the inner bases (a′′) of the bilobed outer maxillæ to the opening of the œsophagus (g): a broad branch from each of these ribs supports the sides of the orifice of the œsophagus; and this branch almost joins on to a slightly thickened rim or bar (f′), which branches off from the upper part of the sub-triangular (f) inner fold of the labrum. This structure, in Bal. improvisus, is represented in Pl. 26, fig. 2, as well as it could be, considering that the deep supra-œsophageal cavity has to be torn open; and then laid flat.
The Palpi (Pl. 26, fig. 6) differ little, except in size, in the different genera, being squarish, more or less elongated, or even approaching to club-shaped: in most of the Balaninæ they are larger even than the mandibles, of which they normally form a part. Their upper margins, especially towards their free extremities, are always thickly clothed with spines; and there is generally a single row, either short (r) or long, of spines of greater length, which arise from a little above, and stand almost in a parallel line to, the basal margin. On the internal surface, there is sometimes a row (t) of very short little spines, which overhang the crest of the labrum. The Mandibles (Pl. 26, fig. 5) have from three to five teeth; the lower point or angle is generally pectinated. In Coronula and its close allies, there are some small teeth intermediate between the four or five main teeth; and in these genera, though members of the sub-family Balaninæ, the lower teeth exhibit only rudiments of being laterally double. The Maxillæ sometimes have a notch under the upper large pair of spines, and in Octomeris brunnea there is a double notch: in many species of Balanus, the inferior corner stands up like a step (Pl. 26, fig. 7, a): in many other genera and species, the whole edge is straight. In all, or almost all cases, the row of spines on the middle portion is double. The Outer Maxillæ are always bilobed on their inner faces, and are clothed with bristles. On all the gnathites, the bristles are often doubly serrated.
M. Martin St. Ange describes, in his ‘Mémoire sur l’Organisation des Cirripèdes,’ p and 32, “une petite langue” in the mouth of Lepas; but I may venture to assert that such does not exist; it is merely the point of union between the outer maxillæ. M. St. Ange, in his comparison of the mouth of Lepas with that of Phyllosoma, compares the mandible of the latter with the palpus of Lepas; the first maxilla of Phyllosoma with the mandible of Lepas; and so on with the other gnathites.
Muscles and functions of the Gnathites, and their confluence. — The outer maxillæ appear at first like a deeply-lobed lower lip, for they reach over almost to the labrum (Pl. 26, fig. 1), and thus partially cover the other organs; they are separately capable of a strong and rapid, to and fro movement, by which no doubt they sweep any prey, entangled by the cirri, towards the other gnathites. Each outer maxilla is furnished with a pair of muscles, apparently a flexor and extensor; there is also a little muscle between the two maxillæ, I presume for the purpose of bringing them together. The outer and inner maxillæ generally stand close together, and in several genera a little way apart, from the mandibles; but there is no trace of any labrum or true lower lip, bounding the mandibles and orifice of the œsophagus. The outer and inner maxillæ and mandibles are not opposed in pairs to each other, but against the thickened inner fold of the labrum; almost in the same manner as the posterior pairs of cirri are not opposed one to the other, but to the mouth.
I have described pretty accurately the muscles of the mandibles in my former volume, and there given a drawing (Pl. 10, fig. 1) of them. There are four muscles: first, the depressor muscle, which is the largest, and is attached, at its upper end, to ligamentous apodemes under the free toothed portion of the jaw; and at its lower end, spreading considerably out, is attached to a concavity close above the basal margin of the labrum; to understand the action of these muscles, it should be borne in mind that the mandible almost faces the labrum. In some genera, as in Coronula, the swelling near the basal margin of the labrum (Pl. 26, fig. 3, k), caused by the internal concavity for the above muscle, is conspicuous. The depressor muscle is opposed by a small elevator, attached to the mandible close by the depressor; thence it runs upwards, and is united at its upper end to the base of the palpus, at the point where the latter adheres to the labrum: I have ventured to call this muscle the elevator, from being apparently so well fitted for this purpose; but I feel some little doubt, from having observed an apparent slight movement in the palpi of living Balani; and this is the only muscle entering those organs. The free part of the mandible is articulated on a square, thickened piece of membrane, forming part of the side of the mouth (Pl. 26, figs. 3, 4, c; and Pl. 10, fig. 1, a, b, in my volume on the Lepadidæ); to this square piece of membrane, two short muscles are attached, one above the other, and which ought, in the Plate in my former volume, to have been represented crossing the depressor muscle at nearly right angles; at their further ends they are attached to about the middle of the labrum, where, at least in Coronula (Pl. 26, fig. 3, i), a slight concavity can be detected. The action of these two muscles must be to draw the whole mandible against the labrum; and the depressor muscle might, at the same time, draw the toothed edge downwards, and thus force any prey into the œsophagus.
This is figured by Burmeister in his ‘Beiträge zur Naturgeschichte der Rankenfüsser,’ Tab. 2, fig. 6.
The inner maxillæ are likewise furnished with four muscles, very nearly as figured in my former volume (Pl. 10, fig. 10); namely, two muscles, one inside and the other outside the curious apodeme, which in the Balanidæ (Pl. 26, fig. 7, b′) is as invariably present as in the Lepadidæ: these two muscles are attached at their lower ends to the outer membrane of the mouth, close to its basal articulation: the outer one of these two muscles would, I presume, act as an elevator, and the inner one as a depressor; the free part of the organ working on the top of the apodeme, like an axe, on a hinge, on the top of a pole. But there is also a larger depressor muscle, in an analogous position with that (i. e. the first-mentioned muscle) of the mandibles; and a fourth muscle, crossing the latter depressor at nearly right angles, and attached (as far as I could make out) on the side of the orifice of the œsophagus, close under the mandibles: the action of this latter muscle would be to draw the whole organ towards the labrum.
I must not conclude my description of the mouth, without drawing particular attention to its peculiar compounded nature. It is prominent, and is capable, as a whole, of movement; it is separated from the body by a fold or articulation, which can be traced all round. It is, as we have seen, composed of a broad labrum and three pairs of gnathites, which latter have only their terminal segments free; and these surround a conical hollow, at the bottom of which lies the opening of the œsophagus. The prominence of the whole mouth appears to result from the lateral fusion of the two basal segments of the three pairs of gnathites. I have examined the mouth of ordinary Crustaceans, and can see no trace of a structure like this. That there has been some union of the parts is indisputable; for the palpi, which in ordinary Crustaceans are quite free, are here firmly united to the upper and outer corners of the labrum; and indeed, at first appear to be more intimately connected with the labrum than with the mandibles. The palpus on its upper and exterior surface, is in direct continuity with the square thickened piece of membrane, on which the mandible is articulated, and likewise with that side of the upper or free portion of the mandible which faces the labrum. This face of the mandible, beneath the toothed edge, is hollowed out or arched (Pl. 26, fig. 5, p), owing to the above-mentioned continuity of its membrane with that of the palpus. On the lower surface, the palpus is firmly united to the lateral corners of the labrum; or indeed the corners of the labrum may be almost said to be formed by the soft, swollen bases of the palpi: the point of union, when viewed from the outside, is seen to form a knob on the shoulder of the labrum, beneath the level of its crest, and at this knob (Pl. 26, fig. 3, close to d′) several thickened bands in the surrounding membrane unite. The free portion of the palpus stands out transversely behind (i. e. anteriorly to, in a homological sense) the labrum. I suspect that the palpus possibly may consist of two segments, of which the terminal one is free, and the lower one confluent with the labrum.
Before proceeding any further, I should observe that figs. 3 and 4, in Pl. 26, represent the membranes of the mouth of Coronula diadema, perfectly cleaned. In fig. 3, all the front part of the mouth has been removed, the mandible on one side, the labrum with the two palpi, and the œsophagus being alone left, and these are viewed from the inner side; the front part, however, of the supra-œsophageal cavity has been cut away. In fig. 4, the labrum, with the œsophagus, has been removed, whilst the two outer maxillæ, the right-hand inner maxilla and mandible (with the exterior and basal portions, d, d′′, of one palpus) are seen from the outside; but in order that these parts should all be shown, the whole of the right-hand side of the mouth has been spread out, for the teeth of the mandible should have stood in a vertical line between the two outer maxillæ. In the mandibles, the free upper part is separated, by a distinct articulation, from the square piece of thickened membrane (fig. 3, c) on which it is supported; and this latter is separated by a second articulation from a portion of thickened membrane (c), the basal edge of which forms the third and lowest articulation, separating the mouth from the body. This basal, thickened portion of membrane curls round and inwards, towards the outer maxillæ or front of the mouth, and its terminal points sometimes even penetrate a little way within the muscles, like apodemes: it is not distinctly separated by any line or suture from the membrane, which forms the whole broad labrum; so that I at first concluded that the labrum dipped under the mandibles, and thus afforded a support on which they were articulated; but this appears so opposed to all analogy, that it is more probable that the above basal thickened portion of membrane is truly the basal segment of the mandibles, completely confluent with the labrum; and it is, I think, not very improbable that even a large portion of what in appearance belongs to the labrum, namely, those concavities to which the muscles of the mandibles are attached, may, also, be part of the basal segment of the mandibles. Whether or no there really are two segments beneath the upper free portion of the mandibles, which have become laterally confluent with other parts, I must think that the square thickened piece of membrane (c) represents at least one segment. I may here observe, that Prof. Milne Edwards seems to consider the mandible of the higher Crustaceans as answering homologically to the haunch of the leg; but, according to M. Brullé, there ought to be two basal segments (sous-maxillaire and maxillaire) bearing the proper mandible, and giving rise, on the outer side, to the palpus, — a structure which perfectly corresponds with my view of the mandible and palpus in Cirripedes.
‘Annales des Scienc. Nat.,’ 3d series, Zoolog., tom. ii, .
Maxillæ: the point whence the long apodeme (b′, Pl. 26, fig. 4 and fig. 7) arises, according to Audouin’s views, must mark an articulation, and this would separate the upper free segment from the lower segments, which I believe to be laterally confluent with the organs on each side. The thickened membrane, of which the upper free part is formed, extends a little distance beyond the insertion of the apodeme; and this small portion beneath the point of insertion may possibly answer to the square, thickened piece of membrane, or second segment, supporting the mandibles. Beneath it, a rather wide expanse of thin, flexible membrane reaches down to the basal fold surrounding the mouth, and may thus form the third segment.
Outer Maxillæ: the upper free segment has a spinose lobe (a′′, Pl. 26, figs. 2 and 4), on its inner face, which may indicate a lower and second, almost free segment. Passing over this, we have, on the outside of the mouth, beneath the free, upper segments, an expanse of membrane, which, on the side, close to the inner maxillæ, is perforated (Pl. 26, fig. 4, n) by orifices which I believe are olfactory. In some species, as in Bal. eburneus and improvisus, there is a longitudinal medial suture in this expanse of membrane, which I suppose indicates the lateral confluence of the middle segments of the two outer maxillæ. A short, transverse articulation or fold separates this middle segment (fig. 4, a) of each maxilla from the third or basal segment; and this latter (a) is separated from the body by a very distinct fold, which (at least amongst the Lepadidæ) sends inwards a short, medial, tongue-formed apodeme. Here, then, we apparently have, as in the mandibles, two segments under the upper free segment of each outer maxilla, laterally confluent with the adjoining organs. But I must state that, in old specimens, and only in old specimens of Coronula diadema, I have found under the outer maxilla an additional transverse ridge and fold, which plainly shows how easily a mere thickening of the membrane might be mistaken for an articulation. I can, however, hardly persuade myself that the articulated membrane, under the free part of the mandibles, which has now been figured and described, has no homological signification; and the fusion of the palpus and labrum seems too plain to be mistaken. Hence I must conclude that the mouth, in the Cirripedia, does truly exhibit a compounded structure of a very peculiar nature.
Cirri.
There are always six pairs; each biramous and multiarticulated, supported on a pedicel formed of two segments. A shield-like swelling at the exterior bases of these pedicels often appears like another segment; but such, I believe, is not its nature. The five posterior pairs answer to the five pairs of ambulatory legs in the higher Crustaceans; and as in the case of the latter, the three, or the four hindermost pairs almost invariably resemble each other. The first pair, which is homologous with the outer maxillipeds of ordinary Crustaceans, is separated by an interval from the second pair; — though this is not the case with the legs of the pupa, from which the cirri are metamorphosed. These anterior cirri are attached to the lateral edges of the mouth, namely to the thickened rim of membrane, forming the supposed basal segment of the mandibles. They are capable of more diversified movements than the other cirri: the anterior ramus is always elongated, with the terminal segments more or less tapering, and is directed beyond (or anteriorly to) the mouth: the shorter ramus closely resembles in structure the rami of the second pair. In the Chthamalinæ the second pair, and in the Balaninæ the second and third pairs (as will be more particularly described under these sub-families) differ in structure from the posterior pairs, from which they are separated by a slight interval. The number of segments on the posterior cirri is often great, amounting in Chelonobia even to fifty. Each segment normally is furnished on its inner face, which is usually somewhat protuberant, with from two to rarely eight or ten pairs of long spines or bristles, placed in a double row; the two spines in the lower pairs stand nearer to each other, and are shorter than the spines in the upper pairs. Between each pair of spines there is either a single, very thin bristle, or often a tuft of such. The pairs are directed somewhat upwards, and they diverge when the cirri are uncurled; their function is obviously to entangle the prey. On the dorsal or exterior surface of each segment, close to its upper margin, there is a tuft of spines, often composed of thicker and thinner spines; these, I believe, serve to prevent any creature intruding within the sack. On both sides of the upper margin of each segment, there is generally a row of short, blunt, excessively minute spines, which only deserve notice, inasmuch as it is by their increase in number and size, and by the spreading out of the dorsal tufts, and, lastly, by the increase of the little tuft intermediate between the pairs of spines situated in front, that the segments on the two or three anterior pairs of cirri become covered, like brushes, with bristles. The bristles or spines on the second and third cirri are often, especially in Tetraclita, doubly and coarsely pectinated. The bristles on the pedicels follow the same arrangement as on the rami; namely, being in regular pairs on the posterior cirri, and crowded thickly, like a brush, on the anterior cirri. The segments in the shorter ramus of the first cirrus, and in both rami of the second, and often of the third cirrus, are broader than the segments of the posterior cirri; they are, also, especially in the genus Balanus, frequently produced in their upper, ante-lateral corners, into remarkable prolongations (see Pl. 29, fig. 4, of the third cirrus of Bal. perforatus), clothed on their inner surfaces, and at their extremities, by numerous bristles. The number of the segments in each cirrus is in some degree variable, and increases with age; this is likewise the case, to a certain extent, with the number of the spines borne on each segment.
As compared with ordinary Crustaceans, I presume the two rami answer to the “tige” and “palpe” of Milne Edwards; and the pedicel (as I have called it) to the two basal segments of the leg. The “fouet” or flabellum does not appear to be developed in any Cirripede; for though the filamentary appendages in certain genera of Lepadidæ, might at first be thought to be of this nature, yet their usual position beneath the basal articulation of the first pair of cirri, and the occasional presence of more than one, proves, I think, that such is not the case.
According to this author’s new nomenclature, the pedicel would consist of the coxopodite and basipodite; the tige would be the ischiopodite and following segments; and the palpe would be the exopodite; the epipodite or flabellum being absent. (‘Annales des Sciences Naturelles,’ tom. xviii, 1852.)
Though the structure of the cirri is very uniform, yet we meet with some peculiarities. In Chelonobia, the segments of the posterior cirri bear only two pairs of main spines; whereas in some varieties of Balanus balanoides, they carry as many as ten pairs in a longitudinal row; but in this latter species, the number of these spines varies, in a singular manner, from six to ten pairs. In Tubicinella, the pairs of spines on the segments of the posterior cirri are arranged so closely one under the other, that they appear almost like a single transverse row. Considering the whole family, the third pair of cirri differs most in structure in the different genera. Thus, in Chthamalus antennatus, the anterior (or outer) ramus (Pl. 29, fig. 3) is thicker and much longer than the posterior (or inner) ramus; the number of the segments in one instance being, in the two rami, 53 and 18; in the longer ramus, the spines are arranged abnormally, tending to form a little circle round each segment; and the whole ramus may be said to be antenniformed, and I believe acts as an organ of touch: the relative number of the segments, I may add, in the two rami and the arrangement of their spines varies greatly in this species. In two other species of the same genus Chthamalus, we have occasionally the anterior ramus in some degree antenniformed, so that this whole structure is variable. In the allied Chamæsipho columna, it is the posterior or inner ramus which is antenniformed, but this peculiar development is more plainly marked in the case of the second pair of cirri than in that of the third pair. In Tetraclita porosa it is, also, the posterior ramus of the third pair which is antenniformed; in this third pair, and indeed in the other cirri, the relative numbers of the segments vary extremely. A similar structure in the third pair, but in a lesser and variable degree, may be observed in some of the other species of Tetraclita. In Balanus vestitus, also, we have, in the third pair, an analogous structure. It is scarcely possible to believe that the circumstance of the second pair of legs, which answer to the third pair of cirri, being antenniformed in certain decapod Crustaceans, is an accidental coincidence; it must be owing to some special affinity in the two groups.
In Chelonobia, the third pair of cirri is of unusual length compared with the second pair, but does not otherwise differ from the type of its sub-family: in Coronula and its allies, on the other hand, the third pair is very short and broad, as may be seen (Pl. 29, fig. 5) in Xenobalanus: in this latter genus, the front surfaces of the segments of the pedicels (fig. 6) of the posterior cirri, are extremely protuberant, almost as in Scalpellum vulgare.
The last peculiarity in the cirri at all worth mentioning, is in the sub-genus Acasta, in which, differently from in all other known Cirripedes, the anterior ramus of the fourth pair does not absolutely resemble the rami of the fifth and sixth pairs; in most of the species, the spines on this anterior ramus are more crowded together, are larger, and are mingled with some short thick points; and the spines in the dorsal tufts are also longer than in the two posterior pairs of cirri; but in A. sulcata (Pl. 29, fig. 2), and in a lesser degree in A. cyathus and A. purpurata, the front margins of the lower segments of this anterior ramus, and of the upper segment of the pedicel, are developed into strong, downwardly curved teeth: it is very remarkable that so beautiful a structure should be extremely variable, as it certainly is in Acasta sulcata.
Caudal Appendages.
With extremely few exceptions, these are present in all the Lepadidæ and Verrucidæ; whereas amongst the Balanidæ they occur only in the two species of Pachylasma, and in one species of Catophragmus; these being the genera most closely allied to the Lepadidæ, and where, consequently, their presence might have been anticipated. These appendages are seated close together over the anus; they are multiarticulate, each segment being sub-cylindrical, with a few small bristles round its upper edge.
Alimentary Canal.
I have not much on this head to add to what I have said under the Lepadidæ. As in that family, the strong internal membrane of the œsophagus terminates in a remarkable, bell-shaped expansion (Pl. 26, fig. 3, g′), which, as observed by M. St. Ange, serves to keep the upper broad end of the stomach expanded. The œsophagus is well furnished with constrictor and radiating muscles for closing and opening it; and it is thus capable of a strong swallowing movement. The stomach runs down to the lower end of the prosoma, and then doubling back on itself extends to the anus. As the prosoma is much elongated in Tubicinella and Xenobalanus, so is the stomach of unusual length in these genera. In several species of Balanus, the upper edge of the stomach is surrounded by from six to eight cæca; these cæca I ascertained, in Balanus perforatus, are branched, and penetrate a considerable way into the body; and some of them at least expand a little at their extremities. Each cæcum, from the manner in which it retained fluid, must, I think, be furnished, at the point where it enters the stomach, with a sphincter muscle. In Tetraclita, Chthamalus, Tubicinella, Coronula, and Xenobalanus, there are no cæca; but in Xenobalanus and Coronula balænaris, there are longitudinal, approximate folds in the upper, broad end of the stomach, which would serve to expose the food to a greater extent of digesting surface.
The presence and absence of these cæca in genera so closely allied as Balanus and Tetraclita, shows, I think, that these cavities are not of high importance; and I must doubt whether Von Siebold’s view (‘Anatomie Comparée,’ tom. i, ), that these cæca form a passage to a true or isolated liver, such as exists in the higher Crustacea, can be admitted. Cæca are said by Von Siebold to occur in some of the Entomostraca, as Daphnia, Argulus, &c.
As in the case of the Lepadidæ, a transparent, structureless, epithelial tube, composed of chitine, containing more or less digested food, is found, in specimens preserved in spirits, occupying the whole length of the stomach, and where there are cæca, sending branched prolongations into them. It does not extend into the œsophagus or into the rectum. This epithelial tube or model of the stomach, filled with excrement, is expelled by the rectum, whole, that is in a single piece, as I observed in some living specimens of Balanus balanoides: in some specimens, however, of Chthamalus stellatus, the excrement was ejected, perhaps from the animal being confined, in fragments, and the sack thus became befouled. Beneath the epithelial layer, the stomach is lined by a delicate, pulpy and cellular mucus layer, which easily peels off in flakes: this is surrounded by a muscular layer with the fibres closely approximated and transverse; and this by a layer of stronger, longitudinal muscles, but more distant from each other. Lastly, outside this double muscular layer, there is a rather thick, somewhat laminated, pulpy layer, abounding with cells, often nucleated, and frequently containing much oily matter. This structure agrees closely with Dr. C. H. Jones’s account of the external covering of the stomach in Daphnia, and which he believes to be hepatic: as in Daphnia, there does not seem to be any ducts. I may here observe, that within the upper part of the prosoma, but not immediately connected with the stomach, I have often observed much white pulpy substance, permeated by lacunal passages, and exhibiting no structure except some excessively minute cells.
‘Philosophical Transactions,’ 1849, . Karsten (‘Nov. Actorum Acad. Nat. Cur.,’ 1845, tab. xx) has excellently figured the testes, as the hepatic glands; and has indicated the ovaria as salivary glands; it is singular that this anatomist overlooked the ducts which lead from his supposed hepatic glands, into the vesiculæ seminales, within which he observed spermatozoa.
The rectum, lined by membrane continuous with that investing the thorax (and seen through it, in Pl. 26, fig. 8, c), extends inwards to about opposite the bases of the third or fourth pairs of cirri. It is longitudinally plaited; the ends of the folds forming a sort of valve where joined on to the stomach. It is coated by circular, transverse muscular fibres: judging from the movements, the anus itself is surrounded by a strong sphincter muscle. The anus opens on the dorsal surface of the thorax (fig. 8, b); but as in the genera, in which caudal appendages occur, it opens under them, the orifice, I believe, is homologically terminal, and owes its dorsal aspect to the aborted state of the whole abdomen, and to the great development of the probosciformed penis; for the anus may be said to be situated on the dorsal base of this organ.
Altogether we see that the alimentary canal is of a very simple structure. The food, judging from the contents of the stomach, seems generally to be composed of infusoria and minute animals: but in the case of Tetraclita, I have been surprised at the size and number of the included amphipod, isopod, and entomostracan Crustaceans, in one case, together with an annelid. I have, also, sometimes seen some confervoid matter within the stomach.
Circulatory System.
On this subject I can add nothing, except to express my conviction that there is no heart, or true vessels; the circulation being strictly lacunal. A passage has often been quoted from Poli, in which he states that he saw a pulsating organ, close above the anus; but I have seen this movement, which appeared to me to be a convulsive twitching of the sphincter muscle of this orifice. The largest lacunal channel extends down the middle of the rostral compartment of the shell: and this answers to the rostral channel down the peduncle in the Lepadidæ. Large nerves and the main pair of unbranched ovarian tubes (Pl. 25, fig. 1, leading into g) extend along this channel. At the basis (at least in Coronula) this channel joins on to a large circular one, running all round the sack, and sending off branches into the mass of ovarian tubes and cæca.
Nervous System.
It has been shown in my former volume, that in Lepas and in some other genera of the Lepadidæ, there are six main ganglions; one supra-œsophageal, and five infra-œsophageal, or thoracic. In Pollicipes, however, there are only four thoracic ganglions, the last ganglion supplying the three posterior pairs of cirri with nerves, whereas in the other genera, the last ganglion supplies only the fifth and sixth pairs of cirri. In this genus, moreover, the lateral fusion of the ganglions has been so complete, that there is no evidence of their having been formed by the union of two. Amongst sessile cirripedes, we discover evidence of much higher concentration even than in Pollicipes. My chief examination has been confined to Coronula diadema, and to Balanus tintinnabulum: and in these genera we find (and the fact appears to me highly remarkable) as high a degree of concentration in the infra-œsophageal ganglion as in any decapod Crustacean, for instance, as in Maia, judging from the figure given by Milne Edwards; for all the nerves, with the exception of those connected with the supra-œsophageal ganglions, radiate from a single great ganglion. The nervous system is, moreover, otherwise complicated.
It must, however, be observed that, according to Mr. Dana, there is in certain suctorial Entomostracans, as in Caligus, only one infra-œsophageal ganglion. Mr. Dana speaks of this as resulting from reduction. In Cirripedes, from the gradation which may be observed from Lepas through Pollicipes into Balanus, the ganglions are certainly not reduced but concentrated. In Van de Hoeven’s figure of the nervous system in Limulus, there is seen to be no chain of thoracic ganglions; all the nerves rising from the circa-œsophageal collar; but this, on the other hand, seems hardly developed into a ganglion.
To begin with Coronula diadema the great infra-œsophageal ganglion (Pl. 27, fig. 1, A) is seated nearly opposite to the anterior margin of the second pair of cirri, which are homologous with the first pair of legs in the decapod Crustaceans. This ganglion shows no trace of any longitudinal medial suture; its shape is hardly discoverable, for it is formed by the union of eleven principal pairs of nerves, besides several arising from its under surface; in outline, however, it may be said to be divided into a posterior and anterior half; the latter being somewhat heart-shaped, and the posterior half elongated. The nerves going to the five posterior pairs of cirri rise from the posterior margin of the ganglion, and run for some distance in a sheet, parallel and close together; the pair, however, going to the second pair of cirri soon branches off from the others. Each of these nerves enters at the inner and posterior margin of the cirrus to which it belongs, and, at least in the case of the first pair, divides into two branches as it enters. The nerves (Pl. 27, fig. 1, r, r) going to the fifth and sixth pairs of cirri are more closely united together than are the others, and appear, till they branch off, like a single large nerve. That which belongs to the sixth cirrus gives off, opposite to the fifth cirrus, a branch (s) nearly as large as itself, which enters the probosciformed penis. I may remark, that homologically this is the only abdominal nerve in any cirripede of the Order. From the under side of the nerves which run to the five posterior pairs of cirri, small branches are given off, extending dorsally into the thorax.
The anterior end of the great infra-œsophageal ganglion is formed by the union of a set of nerves, extending parallel in a bundle in a directly opposite direction to those running to the five posterior pairs of cirri. These nerves consist of an outer larger pair (r) entering the first pair of cirri; and within these, and rather dorsally to their roots, we have the circa-œsophageal chord (c, c), or collar nerve; between the roots of the latter, and on the ventral surface (or near side of the figure), there are three closely united, small pairs, running to the gnathites, and, as I believe, to the olfactory sacks. From the under (or dorsal) surface of the anterior end of the ganglion, two nerves, larger even than the circa-œsophageal chord, and which I shall call the splanchnic pair (d, d) arise; and the singular course of these nerves will presently be described; between this great pair, there is a single (b) medial nerve, which runs down and branches into that large diverging muscle, which is attached to the upper ventral surface of the stomach. Posteriorly to these three nerves, we have two pairs of much smaller nerves (not figured), running dorsally into the body, so that we have seven nerves rising from the under surface of the infra-œsophageal ganglion. I need only further add, that on each side of this ganglion, between the nerves going to the first and second pairs of cirri, there is a moderately sized nerve (k), which appeared to run into the muscles of the thorax: a nerve in a similar position is figured by Milne Edwards in Maia.
The circa-œsophageal chord (c, c) nearly equals in length the whole distance from the centre of the main ganglion to the posterior end of the thorax. This collar bows out on each side, where passing the œsophagus (œ), which is seated at its anterior end. From the collar a branch is given off on each side, which I traced as far as between the mandibles and maxillæ; from analogy with other Crustaceans, it perhaps runs to the mandibles. The collar has not a transverse commissure, such as described by Milne Edwards in the Podophthalmia, and as figured by Van de Hoeven in Limulus.
The supra-œsophageal ganglions (B) present a singular contrast with the infra-œsophageal ganglion in their little development, size, or degree of confluence. They lie directly under the basal edge of the labrum. They are laterally quite distinct, and consist merely of a slight enlargement of the circa-œsophageal chord. From the anterior edge of each ganglion, a broad nerve (f) extends for some distance in a straight line, and, on close examination, can be seen to be formed of two nerves closely united, of which the inner and smaller one, after a space, appears to cross over the larger nerve: both become at this point tortuous, and, giving off branches (m, m), form a plexus. The two nerves (f) then bend inwards, and almost touching each other, run down, together with the two ovarian simple ducts, along the rostral compartment of the shell. No doubt, if the smaller branches from these nerves could be traced, they would be seen to form a network over the whole sack; and would therefore enclose, as in a cage, the rest of the nervous system. These nerves correspond, I believe, to the two pair of antennular nerves of ordinary Crustaceans, and hence I will call them by this name. Just in front, at the outside corners of the two supra-œsophageal ganglions (B), a branch (e′) arises, which I traced to the ends of the adductor scutorum muscle, and to those several muscles which serve to retract the interspace of membrane between the mouth and the adductor.
The pair of great splanchnic nerves above alluded to, which arise from the anterior and dorsal surface of the infra-œsophageal ganglion, are in Pl. 27, fig. 1, d d, (and in fig. 2), laid flat; but in nature they first bow outwards, and then, penetrating deeper into the body, approach each other, and running nearly parallel, pass round the lower end of the œsophagus: their course consequently is nearly similar to that of the circa-œsophageal chord, with this difference, that the outwardly bowed portion is situated near the infra-œsophageal, instead of near the supra-œsophageal ganglion. The splanchnic nerve, a little beyond the supra-œsophageal ganglion, joins a plexus (d′); and into this plexus another large nerve (e) which I will call the supra-splanchnic nerve, sends branches; this nerve takes an almost semicircular bend over the ovarian glands. The supra-splanchnic nerves (e, e), though appearing to spring from the supra-œsophageal ganglions, do really arise, as may be seen by tracing the constituent fibres, from the circa-œsophageal chord. The plexus (d′) lies close to the coats of the upper end of the stomach: several branches, proceeding from it, run further on, but I was able to trace only a few of them: one went (at least in the case of Balanus perforatus), to the adductor scutorum muscle: another branch spread out on the flanks of the prosoma: I strongly suspect that one branch goes to the acoustic sack: it appeared, also, as if some of the small branches entered the second plexus (m), where the inner antennular nerve and ophthalmic nerve cross over the outer antennular nerve.
I have called the nerves (dd, ee) splanchnic and supra-splanchnic, from their course and apparent function in supplying the viscera. In the descriptions of the nervous system of other Crustaceans I can find nothing analogous to my great splanchnic nerve (dd); the so-called supra-splanchnic nerves (ee), which arise from the circa-œsophageal chord, seem to be the analogues of the ordinary splanchnic nerves, though these latter are always described as uniting into a single medial branch. The plexus (d′) is the cervical ganglion of M. Martin St. Ange, who has likewise indicated the course of my splanchnic and supra-splanchnic nerves; but the plexus, when viewed as a transparent object, hardly appears to me to be ganglionic in its nature. In my former volume on the Lepadidæ, I quite misunderstood the course of these splanchnic nerves.
‘Mémoire sur l’Organisation des Cirripèdes,’ .
From the commissure between the two supra-œsophageal ganglions, a straight chord (Pl. 27, fig. 1, g) arises, which terminates in a small ganglion (C), barely exhibiting traces of being formed of two laterally confluent ganglions. This is the ophthalmic ganglion. The chord connecting it with the two supra-œsophageal ganglions is accompanied by a small nerve (h) which runs on to the muscles round the adductor scutorum muscle; the chord is encased by much fibrous tissue, and its dissection is thereby rendered difficult. From the ophthalmic ganglion, on each side, a nerve (i) goes forth and crosses the antennular nerve; these, if I could have traced them, would have been found to run, as may be safely inferred from what is known in Balanus tintinnabulum, to a pair (D, D) of eyes.
In Balanus tintinnabulum, the structure of the great infra-œsophageal ganglion (Pl. 27, fig. 2, A) is essentially the same as described under Coronula. The great pair of splanchnic nerves springing from its under side, are here actually twice as large as the circa-œsophageal chord. The plexus (d′) formed by the splanchnic nerve (d), on each side, with the supra-splanchnic nerve (e), which arises close posteriorly to the supra-œsophageal ganglion, is here much less complicated, but is perfectly distinct; and there was no appearance of the cervical ganglion of M. Martin St. Ange. The chord (g) running from between the two supra-œsophageal ganglions to the ophthalmic ganglion, is nearly as large as the double antennular nerve (f) on each side of it. The ophthalmic chord (which is accompanied in its whole course by a small branch running to the adductor scutorum muscle) terminates in a small ophthalmic ganglion (C), which seems to be formed by the almost complete fusion of two ganglions. This ganglion is hardly larger than the chord which it terminates: it appeared to me to give rise to more than one pair of nerves, and a single nerve (to my surprise) joined the branch just mentioned, which goes to the adductor scutorum muscle.
From each supra-œsophageal ganglion, two closely united antennular nerves (f) extend, of which the inner one crosses over the main or exterior nerve, nearly opposite to the ophthalmic ganglion, and here forms (m) a plexus. The structure of this plexus I was not able, any more than in Coronula, to make out thoroughly, but I traced quite distinctly a long nerve (i) running from it into what must be considered as the eye. As in the case of Coronula, I traced a nerve on each side from the ophthalmic ganglion into the plexus, where I lost it; and as here in Balanus, I saw on each side of the ophthalmic ganglion a cut off nerve, of about the size of that which runs from the plexus on each side into the eye, I think we may safely conclude that the latter or optic nerve does really arise from the ganglion here called ophthalmic. I may add that the analogy of the nervous system in the Lepadidæ most strongly confirms the view of this latter being the ophthalmic ganglion.
Eyes and Vision.
The optic nerve (i), running from the plexus to the eye, is of considerable size; it runs nearly parallel to the main antennular nerve, diverging from it a little. It retains nearly the same diameter throughout; and gives off only one single, small, inner branch. It can be traced beyond the basal edges of the scuta, to just under the upper edge of the transparent opercular membrane, which unites the scuta to the sheath of the rostrum. The nerve itself, at a little distance from its further end, was, in a full-sized specimen, 5/1000ths of an inch in diameter; widening a little, it expands slightly, and abruptly terminates in a circular disc, about 8/1000ths of an inch in diameter, (see Pl. 27, fig. 5). The nerve just beneath this slight expansion, is coated all round by pellets of dark purple pigment-cells, but not actually united into a continuous layer. These pigment-cells are the more conspicuous from the surrounding parts being colourless. I could not make out distinctly any cornea; and I suppose the external transparent membrane, to which the above slight circular expansion is attached, acts as such. This description very closely agrees with that given of these organs in Bal. rugosus of Gould, (B. crenatus?) by Dr. Leidy, who first discovered the eye in the adult cirripede, but he did not observe the ophthalmic ganglion. These eyes differ from those in some of the genera of the Lepadidæ, only in the greater length of the optic nerve, and by standing laterally further apart from each other.
‘Proceedings of the Acad. Nat. Sciences of Philadelphia,’ vol. iv, 1848, . I may add that I have, also, observed the supra-œsophageal and ophthalmic ganglions in Bal. perforatus.
I may here mention that I tried a few simple experiments on the senses of Balanus balanoides, B. crenatus, and Chthamalus stellatus. I found these three species very sensitive to shadows, that is, to an object like my hand passing even quickly, and at the distance of about a foot, between them and the source of the light. They were indifferent to a gradual change from bright to obscure light; but instantly perceived and drew in their cirri, when my hand was passed between the basin in which they were kept and the window, even when this was tried rather late on a dusky evening; and likewise when my hand was passed between them and a single candle. I took, of course, the precaution of passing my hand in other directions, but this never produced any effect. These species are moderately sensible to any vibration in the vessel in which they were kept, but they were indifferent to noises made in the air, or in the water. I found it impossible to touch, under water, an individual shell ever so lightly with a needle, without all the immediately surrounding individuals, when several adhered together, perceiving it, and retracting their cirri: it made no difference whether the one touched had already withdrawn its cirri and was motionless: from this fact, and from seeing that a similar but slighter effect was produced by touching the rock on which the specimens adhered, I infer that the perception by the others of the one being touched, is communicated by vibration. When an individual was touched under water, not by a needle, but by a pointed camel-hair brush, it generally withdrew its cirri, but the neighbouring specimens took no notice: when touched by a single hair of the brush, no notice was taken, unless the skin of the orifice leading into the sack was so touched. In these trials, it is of course necessary carefully to avoid intercepting the light. I could not make out that cirripedes perceived odours diffused in the water.
I find that this fact was long ago observed by Von Siebold, ‘Anatomie Comparée,’ tom. i, .
Acoustic Organs.
These are situated in the same position as in the Lepadidæ, namely, in a slight swelling on the sides of the thorax (Pl. 25, fig. 1, d′) just beneath the basal articulation of the first pair of cirri. The orifice in Tubicinella and Xenobalanus is slightly produced, or is tubular; the free part in the former genus projecting 5/100ths of an inch. The structure of all the parts is essentially the same as in the Lepadidæ, but I think all are proportionally larger. The external membrane of the body is turned inwards at the orifice, as a short flattened tube, which widens considerably (being, in a middle-sized specimen of Coronula, 4/100ths of an inch in width) before it abruptly terminates. The meatus, as I have called the sack-like cavity which encloses the true acoustic sack or vesicle, is formed of pulpy membrane, and is apparently continuous with the corium of the whole body, but by dissection it can be separated entire. The acoustic vesicle is of various shapes, as we shall immediately see; but in all essential respects it is identical with the same part in the Lepadidæ; it is formed of the same peculiar, soft, elastic, brownish, transparent tissue, which seems to be composed of fine, transverse pillars, becoming towards the outside fibrous, and at their inner ends appearing when viewed vertically from above, like hyaline points. In Coronula diadema, I observed on the outside of the acoustic vesicle, some excessively minute bristles, only 1/3000ths of an inch in length, seated on little eminences. I examined carefully the contents of the vesicle in this species, in specimens well preserved in spirits, and there was nothing within but a very little, thin, pulpy fluid, and a few yellowish nucleated cells, here and there aggregated into small groups. In Coronula, the flattened acoustic vesicle is elongated, with a somewhat sinuous, but not very irregular margin (Pl. 27, fig. 4), and is without any ridges on the surface; its neck or orifice projects at right angles to the elongated portion, which stands obliquely to the tubular orifice of the meatus. In a moderately-sized specimen of Coronula diadema, the elongated portion of the acoustic vesicle was, 6/100ths of an inch in length. In Tubicinella, the acoustic vesicle is heart-shaped, with the neck attached to its broader end; and the surface is covered by zig-zag ridges. In Balanus tintinnabulum (fig. 3), the acoustic vesicle is almost square at the lower end, with the neck placed at one of the upper corners; on the external surface, there is an oblique prominent ridge or fold, which sends off downwards another ridge; its length, in a large individual, was 5/100ths of an inch.
In all these cases, the acoustic vesicle is mainly attached by its neck, to the upper end of the sack-like meatus; but there is likewise a layer of soft, pulpy, cellular matter, slightly connecting that side of the vesicle which is opposite to the neck, with the walls of the meatus or outer sack. The mouth or orifice of the vesicle is closed by a delicate lid or diaphragm, which can easily be separated; and this diaphragm is formed by the expansion of a large nerve, which here abruptly terminates. In a very large specimen of Coronula diadema I clearly made out the existence of this nerve, and traced its course for some distance from the point where the summit of the meatus and the neck of the vesicle are joined together; the nerve first runs posteriorly, and then turns inwards and doubles back or anteriorly; and I clearly followed it to the antero-lateral sides of the uppermost end of the stomach, where it seemed to enter a ganglion, so that I unfortunately cut it off, but found only a slight plexus, with the cut off nerve apparently running onwards with nearly the same diameter. The diameter is great, fully equalling, in its widest part, that of the circa-œsophageal chord; but it is very much flattened, and so has not nearly so much bulk as that nerve. Before it reached the stomach, it gave off one branch, which ran towards the mouth. The only nerves which, from their size, could, I think, be continuous with this from the acoustic sack, are the main branches proceeding from that plexus (d′) formed by the interbranching of the splanchnic and supra-splanchnic nerves.
I have always feared that anatomists would reject my view of these organs being acoustic, owing to the absence of otolithes; but I observe that so high an authority as Von Siebold (‘Anatomie Comparée,’ tom. i, ) does not believe that otolithes occur in the acoustic organs even of the highest Crustacea. He considers an “ampoule volumineuse, a parois mince, remplie d’un liquide transparent,” and a “membrane tympanique,” though having a fissure in the centre, as sufficient. I may here remark, that the nerve proceeding from the acoustic vesicle in Cirripedes, and apparently running to the splanchnic nerve, may easily be placed in connexion with the antennular nerves, by the second plexus (m) in figs. 1 and 2, pl. 27. I should infer from Von Siebold’s remarks on his ampoule volumineuse in the higher Crustacea, that my acoustic vesicle answered to the labyrinth in higher animals.
Olfactory Sacks.
I can add nothing to the account given of these organs under the Lepadidæ: I saw them in all the genera which I examined for this object. In Coronula diadema the orifices are large; they are seated in the usual position (Pl. 26, fig. 4, n), in the confluent segments, beneath the free part of the outer maxillæ, and somewhat exteriorly, or as near as possible to the inner maxillæ. In no sessile cirripede are the orifices produced or tubular, as is the case with several genera amongst the Lepadidæ. I failed, as heretofore, in tracing with certainty the nerve, which appears to enter the base of the sack, to its ganglion.
Male Organs of Generation.
All the Cirripedes of the family we are now describing, are bisexual or hermaphrodite; and no instance has been observed of the presence of males or complemental males. I have very little to add to the observations made by M. Martin St. Ange and R. Wagner, and to those given in my former volume. The testes seem always to be confined to within the thorax, including the prosoma. With their ducts, they resemble club-moss or stag’s horns, with the extremities a little enlarged: a figure of a small portion from Balanus perforatus is given in Pl. 25, fig. 2. It is quite surprising how like in structure and appearance the branching ovarian tubes often are to the testes with their ducts; but the latter are of smaller diameter. Two main ducts generally unite just before entering the broad, often reflexed, end of the vesicula seminalis: in Coronula balænaris, however, I observed four ducts entering this receptacle. The two vesiculæ seminales, lying within the thorax and prosoma, are usually very long and tortuous: they are formed of a thin inner tunic, which is strengthened by thicker reticulated lines, and of an outer layer of transverse fibres, which are either elastic, or probably muscular, as they serve to expel the contents with force when the end is cut off. The inner tunic is prolonged up the probosciformed penis, at the base of which the two vesiculæ unite. The contents of the vesiculæ are commonly pulpy and cellular; and from the cells the spermatozoa are developed; soon after their development, they are, as it appears, expelled.
The ‘Report’ on M. Martin St. Ange’s memoir was laid before the Academy of Sciences, July 14, 1834, so that I suppose it was read previously to this date. R. Wagner’s paper was published in ‘Müller’s Archiv,’ 1834, . Burmeister’s ‘Beiträge zur Naturgeschichte der Rankenfüsser,’ was published this same year, 1834; so that these three authors published almost contemporaneously.
A far better figure is given by Karsten (‘Nov. Act. Acad. Cæs. Nat. Cur.,’ 1845, Pl. 20, figs. 2, 3, 4), but under the erroneous supposition that these organs were hepatic.
In Conchoderma aurita, the ducts, as shown by Burmeister (‘Beiträge,’ &c. tab. ii, fig. 17), unite half way up the probosciformed penis.
I have seen the spermatozoa in Balanus crenatus, perforatus, and balanoides, and in Chthamalus stellatus. The cells, from which the spermatozoa are developed, and which are often found in vast numbers within the vesiculæ, are on an average about 1/5000th of an inch in diameter. The spermatozoa differ remarkably within the vesicula of the same individual, according to their state of development. I have observed in B. perforatus and in the Chthamalus, that the shortest, and therefore, I presume, the youngest (Pl. 29, fig. 7, a), had a globular head with no projection in front: as they increased in size, this head became less in diameter, and a short tapering filament, (a, b,) like the tail, projected out of it. This anterior filament does not lie in exactly the same line with the posterior filament, which is straight as an arrow. In Bal. crenatus, the anterior filament was 1/2000th of an inch in length, and the posterior filament 4/2000th, giving a total length of 5/2000th: in the longest and best developed specimens of Chthamalus stellatus, the nodular enlargement was much elongated and spindle-shaped, and not above half the diameter it had in the earliest stage; the posterior filament (measured from the front of the enlargement, this consequently being included) was 5/2000th in length, and the front part only 1/4000th, giving a total length of 11/4000ths of an inch. These observations agree pretty well with Kölliker’s; but this author states, that perfectly developed spermatozoa are absolutely without any nodular enlargement: if this be the case, I have never chanced to see the spermatozoa in their perfect condition. Mr. Bate, also, figures some (Pl. 29, fig. 7, c) in this state, without any enlargement.
‘Annales des Sciences Naturelles,’ (2d series), tom. xix, . Kölliker refers to Wagner’s paper on the same subject, in Wiegmann’s ‘Archiv,’ 1835, part ii, pl. iii, fig. 9. He also refers to Von Siebold’s observations. Mr. C. Spence Bate has figured, in the ‘Annals and Magazine of Natural History’ (vol. viii, 2d series, 1851), the spermatozoa of Balanus balanoides, perforatus, and of Verruca (Clitia) Strömia, and of these I have given copies, Pl. 29, fig. 7.
The probosciformed penis lies adpressed on the under side of the thorax, with its apex generally projecting between the first and second pairs of cirri. It presents the same ringed or articulated structure as in the Lepadidæ: it arises from an unarticulated projection or support, which also forms the posterior border to the anus. This support often terminates, as first observed by Poli, in a very sharp point; but this point cannot be of much functional importance, for though present in Balanus balanoides, it is absent in the closely allied B. crenatus; in Tubicinella there is only a rudiment of this point; I have not observed it in any member of the Chthamalinæ. The strong, transverse and longitudinal muscles with which the penis is furnished, are attached to this support. The apex or orifice of the penis is, I believe, invariably surrounded by some bristles. Its length varies much, according to its state of contraction or relaxation; and this again, I believe, is dependent on the condition of the male secreting organs. In a small specimen of Elminius modestus, the penis was actually thrice as long as the whole thorax, including the prosoma: in Pachylasma and in Octomeris angulosa, the penis is very short, being equal only to once and a half the length of the pedicel of the sixth cirrus: in Octomeris brunnea, the unarticulated support is much elongated, being as long as the pedicel of the sixth cirrus, in which respect this organ resembles that of Ibla quadrivalvis, and of no other Cirripede. From the attachment of the penis at the posterior end and on the under side of the anus — from the position of the caudal appendages (where such occur) over the anus — from the position of these same appendages in the pupa — and lastly, from the position of the papilla-like penis in the abnormal Proteolepas, I infer that, homologically, the penis is situated at the apex of the abdomen, on its ventral surface; and that, consequently, this organ cannot be considered as the abdomen itself in a modified condition.
Female Organs of Generation.
I have scarcely anything to add to the statements in my former volume. These organs consist of the true ovaria, or glandular bodies seated on each side, not far from the basal edge of the labrum; of the main or unbranched ovarian ducts; and of the (Pl. 25, fig. 1, g) ovarian branching tubes and cæca. I traced distinctly in Balanus, Tetraclita, and Coronula, the two main ovarian ducts, running from within the prosoma to the layer of inosculating, branching, ovarian cæca which overlie the basis. In Coronula diadema one of these main ducts was 1/100th of an inch in diameter. Though I traced these ducts near to the grape-like, glandular masses, which I cannot doubt are the true ovaria, I did not succeed in tracing them into actual connection. As in the Lepadidæ, these ovarian glands lie on the sides, near the basal margin of the labrum, and almost under, but rather to the outside of the antennular nerves. The branching and inosculating ovarian cæca form a layer, which corresponds with the mass filling up the peduncle in the Lepadidæ. In Tetraclita they do not cover the whole basis, but are confined to the circumference; they, however, likewise extend up between the two layers of corium round the walls of the shell, and chiefly in the interspaces between the depressor muscles of the opercular valves. In Chelonobia, they enter between the radiating septa in the thickness of the walls: in Coronula diadema, they extend from over the basal membrane into the six large square chambers (Pl. 16, fig. 7, v) separating the radii and alæ: in Tubicinella they are confined to the basis: in Xenobalanus, they form a layer over the basis and likewise round the upper part of the peduncle-like body, which answers to the shell of other sessile cirripedes.
These are well described in Lepas, by R. Wagner, in ‘Müller’s Archiv,’ 1834, . Von Siebold, I observe, refers to Burmeister as the first author who discovered the ovarian cæca within the peduncle; I had thought that M. Martin St. Ange had a prior claim.
These are obscurely figured by Karsten (‘Nov. Act. Acad. Cæs. Nat. Cur.,’ 1845, Pl. 20, fig. 1d) as salivary glands; they were so considered by Cuvier and M. Martin St. Ange: I may observe that salivary glands have not been positively recognised in any Crustacean.
As after the most careful and repeated examinations of various Lepadidæ, I was convinced that there were no oviducts, so I have come to a similar conclusion in regard to the Balanidæ; the ova being brought to the surface, by the formation of a new membrane round the sack underneath them, and by the subsequent exuviation of the old membrane. The ova are united together by a most delicate tunic investing each egg; the ovigerous lamellæ being thus formed, as in the Lepadidæ. In the cases of Chthamalus stellatus, Balanus balanoides, and Platylepas decorata, I saw a pair of very distinct but fragile lamellæ. In Xenobalanus, the two ovigerous lamellæ form two sub-cylindrical packets, pointed at their lower ends and often cohering. There are no ovigerous fræna, for the attachment of the lamellæ; the ova being sufficiently well retained, as it would appear, by the well-closed shell. I have elsewhere stated my full belief that it is the ovigerous fræna which have been metamorphosed into the branchiæ of the Balanidæ. Most sessile cirripedes breed when very young; and I have every reason to believe that they breed several times in the year. The ova are ovate, and vary in length from 14/2000th of an inch in Chthamalus, to 19/2000th in some species of Balanus, in which this greater length was owing to a more elongated shape, — up to 25/2000th in some other species of Balanus. The ova are wonderfully numerous, especially in the genus Coronula.
I may here mention the singular case of some elongated specimens of Balanus balanoides, from Tenby, in South Wales: some of these presented nothing abnormal; but in no less than seven specimens, the two, three, or four posterior pairs of cirri, either on one or both sides, were in an almost rudimentary condition, being of small size and having a shrunk and wasted appearance. In six out of these seven specimens, the probosciformed penis was quite short and abruptly truncated, as if from abortion. By cutting off the truncated apex, and cleaning the external tissue, I ascertained that it was imperforate, apparently in all the cases, and I am certain of this fact in several of the cases. In three of the specimens, I examined the vesiculæ seminales; in one, I found some spermatozoa, but cohering together in a peculiar manner; in the second, there were no spermatozoa; and in the third, the vesiculæ were shrunk, empty, and quite rudimentary in size. So that these three individuals certainly could not have impregnated their own eggs; nevertheless, within the shell of these very three, there were perfectly developed larvæ: I am led to conclude from this fact, that adjoining specimens in a perfect condition had, by means of their long probosciformed penis, effected the fecundation of their imperfect neighbours. I need only further add, that some out of the above six specimens, with more or less aborted cirri and imperforate male organs, were infested by a peculiar parasite, allied to Bopyrus, and that these specimens did not contain ova.
I have given a short notice on this parasite, in my former volume on the Lepadidæ, in a foot-note to .
Metamorphoses and Homologies, throughout the Order of Thoracica.
In my former volume, the metamorphoses were described under three principal stages or heads; but whether these three included all the main changes, I was then hardly able to conjecture. But now I have reason to believe that such is the case, for in the genus Cryptophialus, belonging to the Abdominalia, the whole course of the metamorphosis, from the egg to the pupa, takes place within the sack of the parent; and I found, when having, on the coast of South America, numerous specimens to examine, that the egg-like larvæ (Pl. 24, fig. 15-18) could be naturally grouped into two main stages, but with many transitional intermediate grades (answering to the successive moults in the first stage of ordinary larvæ), before they passed into the third or pupal stage. And the first two stages in these egg-like larvæ of Cryptophialus, clearly seem to correspond with the first two stages in ordinary larvæ; for in both the chief changes are, the shortening of the terminal projection — the increase in size and approximation on the ventral surface of the anterior horns or cases for the antennæ — and the compression of the whole body. In all members of the Thoracica, the metamorphosis seems to run a remarkably uniform course. The larva in the first stage undergoes several moults and slighter changes — how many is not known — before arriving at its second main stage, which has been observed only in one single instance; and judging from Cryptophialus, this second stage passes abruptly by one moult into the pupal stage; and this, certainly, passes abruptly into the Cirripedial or mature stage.
Larva, First Stage.
The larvæ in this stage are known, amongst the Balanidæ, in Balanus, Pyrgoma, Coronula, Platylepas, and Chthamalus; and these genera include all the principal forms. Amongst the Verrucidæ they are known in its one genus, Verruca. Amongst the Lepadidæ, in Scalpellum, Ibla, Alcippe, Lepas, Conchoderma, &c.; and in all these genera the larvæ present no important difference — hardly any difference which could be viewed as generic, were these larvæ independent animals, — as may be inferred, chiefly, from Mr. C. S. Bate’s descriptions. The abstract given in my former volume was not accompanied by any illustrations, and I have consequently here given (Pl. 29, fig. 8), a view of the larva, in the first stage before moulting, of Scalpellum vulgare: the natatory legs are not drawn with accuracy, only the relative position of the several organs having been carefully attended to. I have also had copied from Mr. Bate’s memoir, a figure of the larva (Pl. 29, fig. 9) of Balanus balanoides, in its first stage, before moulting, with its ventral surface exhibited; and another figure (with a few trifling alterations made after examining specimens most kindly sent me by Mr. Bate) of the larva of Chthamalus stellatus (fig. 10), in its first stage, but after moulting once. It should be observed that Mr. Bate has given a drawing of the larva of this latter cirripede, in the first stage, before moulting; and it does not differ essentially from that just referred to (fig. 9), of B. balanoides, but is rather more fully developed. These drawings suffice to show the character of the larvæ in the first stage, both before and after the first moult, and even after the second moult, throughout the Order of Thoracica. The larvæ sometimes undergo their first moult within the sack of their parent, as I have been informed by Mr. Bate, and as I have observed in Coronula.
‘Annals and Magazine of Natural History,’ vol. viii (2d series), 1851, Plates 6, 7, and 8.
I will now make a few remarks on these larvæ in the first stage, before and after the first moult, supplemental to those in my former volume. Their shape is oval, and the whole dorsal surface is evidently covered by a carapace. It is remarkable that the body exhibits no distinct articulations; those given by Goodsir being certainly erroneous. Commencing at the anterior extremity, the eye varies considerably in the state of its development; in Platylepas decorata it is nearly circular, and in most of the specimens very distinct; whereas in the allied Coronula balænaris, before the first moult, it is very imperfect, but afterwards square and of considerable size. In Balanus galeatus, in the immature larvæ dissected out of the egg, the cellular matter which was in process of conversion into the eye, formed a transverse band, obscurely divided into two portions, and this seems to indicate that the single eye is in fact formed by the confluence of two eyes. In Scalpellum vulgare, this heart-shaped eye lies between a V-shaped muscle, the nature of which I cannot understand, and which has not been represented in (Pl. 29, fig. 8, a). I need only further add, that in Chthamalus stellatus, after the first moult, the eye exhibits, in specimens sent me by Mr. Bate, some appearance of tending to become double.
‘Edinburgh New Philosophical Journal,’ July, 1843, Pl. 3, 4.
Arising posteriorly to the eye, we see, in Scalpellum vulgare, a pair of minute curved horns (b′), directed backwards; and within these horns I distinctly saw an articulated organ. These horns are difficult to be distinguished, and probably could not be made out previously to the first moult, in any larva of less size than that of Scalpellum vulgare. But after the first moult, Mr. Bate has seen, in two species of Balanus, in Verruca and in Chthamalus (fig. 10, b), a pair of articulated organs, in this same position, evidently now forming antennæ, and directed anteriorly, and free from any envelope. It is somewhat important, as we shall presently see, to bear in mind that these antennæ first appear within an envelope or horn; and that I detected that they included an articulated organ, before I had heard of Mr. Bate’s observations. These antennæ, from their small size, from being seated internally with respect to the horns containing the other pair of antennæ, and from the position which the latter assume in the later stages of the larva, I believe to be the first or anterior pair. Their position in appearance posteriorly to the large lateral horns, containing the second pair of antennæ, is probably due to the anterior cephalic segments having been driven inwards, the truncated outline of the front of the head, and likewise, probably, the position of the mouth between the bases of the natatory legs being thus caused.
In this same larva of Scalpellum vulgare, within the great lateral horns just alluded to (fig. 8, c), filiform organs, supporting rows of spines, could be distinguished; and these appeared to me to be antennæ. These horns or cases resemble in structure the smaller pair just described; they arise from the ventral surface, and can hardly, therefore, be considered as prolongations of the carapace. After the first moult (fig. 10, c) they are seen to have increased much in length: in some cases they are of considerable length before the first moult, as in Lepas: in the Balanidæ they seem to be generally shorter than in the Lepadidæ; but in Balanus galeatus I found them one third of the entire length of the animal. Whilst within the egg, these horns are adpressed laterally to the body, and so point posteriorly; afterwards they project rectangularly from the sides, or, as in Scalpellum vulgare, are directed somewhat anteriorly. As in the larvæ of all ordinary Crustaceans, as yet known, the antennæ are amongst the earliest developed organs; and as the first pair of natatory legs (Pl. 29, figs. 8-10, e) in these Cirripedial larvæ, might so very naturally be thought to be antennæ (as has been remarked to me by Mr. Dana), both from their structure and from their position a little anteriorly to the mouth, I am well aware that to prove my view correct, namely, that these horns are the second pair of antennæ in process of formation, it is not sufficient merely to have seen organs resembling antennæ within them; nor is it sufficient to advance the strictly analogical fact of the first-mentioned pair of antennæ, which in Scalpellum indisputably appear in their earliest condition within an envelope or horn. Further evidence is required, and this is presented in Cryptophialus, in which the lateral horns of the egg-like larva, in its first stage (Pl. 24, fig. 16), can be actually followed step by step until, in the second stage (fig. 17), just before passing into the pupa, the horns are seen to have become larger and more nearly approximated to each other on the ventral surface; and whilst in this condition, I several times dissected out the prehensile antennæ of the future pupa with every character perfectly recognisable. Hence I cannot doubt that in the larvæ of Cirripedes the law of development is, that in their very earliest condition, the small first pair of antennæ are enclosed in cases; and that the large second pair remains thus enveloped until the pupal stage. This conclusion, we shall immediately see, is in harmony with the late development of the succeeding appendages or organs of the mouth, which certainly do not appear in the first larval stage, and are not known to appear even till after the final metamorphosis.
According to M. Joly, (‘Annales des Sciences Naturelles,’ 2d series, tom. xix, ) in the larva of the macrourous Caridina, the natatory legs appear before the gnathites or parts of the mouth; so that in ordinary Crustaceans there is no invariable order of development from the anterior towards the posterior end of the body, as has sometimes been supposed.
The mouth is more or less probosciformed (Pl. 29, figs. 8-10, d), differing considerably in this respect in different species of the Lepadidæ; and this, probably, is due to the larva being born in a more or less mature condition. Its exact position likewise varies, for it arises either between the first or second pairs of natatory legs. It is known, from Mr. Bate’s observations, to have the power of movement. It is directed posteriorly, the œsophagus extending anteriorly; both these directions being the same as in the mature cirripede. Certainly during these early stages there are no jaws or gnathites; but the margin, answering to the labrum, is furnished with some short, thick, sharp spines, and with hairs. In Scalpellum vulgare the orifice of the œsophagus seems to lie rather beneath the upper prominent spinose edge, which, as just remarked, probably answers to the labrum; but this is one of the species in which the probosciformed mouth, at least before the first moult, is not much developed.
We come, now, to the three pairs of natatory legs: the first (Pl. 29, figs. 8-10, e) has throughout the order only one ramus, whereas the two succeeding pairs (f, g) are biramous. I must here remark that the straight and strong, and the curved plumose spines, with which these limbs, after the first moult, become furnished, now appear to me as more probably prehensile, rather than masticatory as I imagined in my former volume. That these spines are important organs to the larvæ I do not doubt. With regard to the homologies of these three pairs of limbs, my first impression was that they were the mandibles and the two pairs of maxillæ in their earliest condition; but I consider this view as quite untenable, for several reasons; viz., the wide interval between their bases and the mouth itself, — the somewhat variable position of the mouth with respect to the legs, — and the position which the latter occupy in the second larval stage. A far more tenable view is that these three pairs of legs are the three pairs of maxillipeds, in their earliest condition, in accordance with the view of M. Joly on the nature of the three very similar pairs of natatory legs in the larva of Caridina, a macrourous Crustacean. But, in Cirripedes, the three pairs of natatory legs, in the larva in the first stage, are apparently the very same as the first three pairs, in the larva in the second stage, and in the pupa. And in the pupa the first three pairs, which certainly correspond with the first three pairs of cirri in the mature animal, seem to me, for reasons presently to be assigned, to be the second, third, and fourth thoracic limbs. Hence I am led to the conclusion that the first pair of legs in the larva in the first stage, are homologically the second thoracic (answering to the third pair of maxillipeds in the higher Crustaceans), and that the two succeeding pairs are the third and fourth thoracic limbs; to be succeeded, in the pupal stage, by the fifth, sixth, and seventh thoracic appendages.
Mr. Dana, moreover, has remarked, (‘Crustacea: United States Exploring Expedition,’ ), “that he knows of no instance of a mandible becoming so completely a leg, as to lose wholly the mandibular function even of its basal portion.”
‘Annales des Sciences Naturelles,’ 2d series, tom. xix, 1843, . M. Joly’s observations were made on the Caridina. I owe to the great kindness of Mr. C. Spence Bate, an examination of some larvæ of the allied genus Hippolyte varians, and I found, on dissection, the view of M. Joly, that the three pairs of natatory legs are the maxillipeds, so far strongly confirmed, that they followed closely, with equal intervals, the mandibles and two pairs of maxillæ. The first pair of natatory legs in Caradina, in its earliest condition within the egg, is uniramous, like the first pair in the larvæ of Cirripedes. There is one fact which seems rather strongly opposed to the view of these three pairs of legs in the larvæ of the macrourous Crustaceans being the maxillipeds, which is that Capt. Du Cane (‘Annals of Nat. Hist.,’ 1838, vol. ii, pl. 6, and 7) observed only three pairs of limbs in process of formation posteriorly to the first three pairs, whereas there should be found, in accordance with M. Joly’s view, five pairs, i. e. all five pairs of ambulatory legs. This one fact countenances the view, which I hold on the nature of the legs in the larvæ of Cirripedes during their early stages, namely, that they are the second, third, and fourth thoracic limbs, to be succeeded by only three additional pairs.
Lastly, behind the natatory legs, on the ventral surface, (Pl. 29, figs. 8, 9, i), the body is much produced, and terminates in a horny fork, which, after the first moult (fig. 10, i), becomes much elongated. Anteriorly to this fork, on the ventral surface, there is another fork (l), and again above this I could distinguish, in Chthamalus stellatus, after the first moult, another fork (m), or at least a pair of short thick spines. From the structure of the forked abdomen in the known larvæ of the Podophthalmia, I presume that this portion of the body is the abdomen of the young Cirripede, but it is not at all plainly articulated. After the first moult, the posterior end of the carapace (h), which is always pointed, becomes much elongated and serrated on both sides; reminding one of the structure of the carapace of the so-called Zoea, or larva of certain Podophthalmia. Situated under this posterior prolongation of the carapace, there is a swelling (n, with long hairs on both sides), which apparently lies on the dorsal surface of the spinose and forked abdomen; here, when the larva is compressed, the cellular and oily contents of the body burst forth; and I suspect that this swelling is the anus, for it is known from the researches of Rathke, that the anus in the higher Crustaceans opens during the earliest periods dorsally.
I suspect that the account given by Goodsir (‘Edinburgh New Phil. Journal,’ 1848) of the posterior points of the carapace and abdomen in the larva of a Balanus, is not quite accurate.
‘Annales des Scienc. Nat.,’ tom. xx, .
Larva, Second Stage.
I have given, from Burmeister, a lateral view (Pl. 30, fig. 1) of the one single specimen, ever observed of a larva in this stage, belonging, as is supposed, to the genus Lepas. The carapace has now greatly altered its character. The two fleshy projections, as so called by Burmeister, by which the larva adhered to the sea-weed, were supposed by this author to include the great prehensile antennæ of the pupa; from my observations, already alluded to, on the two projections (Pl. 24, fig. 17) in the closely analogous egg-like larva, in the second stage, of Cryptophialus, by which it also adheres, I have not the least doubt that this is the case. The small, internal, and anterior pairs of antennæ, are, as it would appear, now aborted. The eye, according to Burmeister, has commenced becoming double; but the two approximate eyes are not as yet compound. The mouth is probosciformed (m), and does not differ much from its condition in the first stage; no gnathites were observed by Burmeister, and they could not be expected to be present, for they are not found even in the pupa. The mouth, which in the larva in the first stage differs in different genera, in being more or less advanced forward, here stands some way anteriorly to the natatory legs, as in the pupal condition. The first pair of legs is uniramous, and the two other pairs biramous; this fact, together with the number of the legs in this second stage being still three, and their structure being not very different, leaves little doubt on my mind that we here have the same three pairs as during the first stage. The abdomen has become much shortened, but still space is left for the development, in the pupa, of the three posterior pairs of legs. I may here remark that in the pupa the anterior natatory legs have become, like the others, biramous; but yet, as it were for the purpose of showing their metamorphosis from the uniramous legs of the earlier stages, they have their bristles arranged rather differently from those on the succeeding five pairs of legs.
‘Beiträge zur Naturgeschichte der Rankenfüsser,’ tab. 1, figs. 3, 4.
Larva in the Last or Pupal Stage.
I have given a lateral view of the pupa of Lepas australis (Pl. 30, fig. 2), illustrative of the description in my former volume: the specimen is drawn as if transparent, and it was to a certain extent thus rendered by boiling in caustic potash. A sketch of the position of the young Cirripede within the pupa, was made by the camera. At first the drawing will perhaps hardly be comprehended: the darker shaded portion to the left of the letter (b) shows the extent of the sack, with the included thorax and natatory legs of the pupa: to the right of the same letter, if we do not consider the young included Cirripede, the only organs distinguishable in the mass of cellular and oily matter, are the alimentary canal, the cement-glands (t), i. e. the incipient ovaria, and the cement-ducts (t′) which enter the antennæ. A view is also given (fig. 4) of the ventral surface of the pupa; and a transverse section (fig. 7) of the carapace, taken close to the eye-apodemes. On comparison with the larva in the second stage, the changes in external appearance and structure are not very great; the prehensile antennæ are freed from their cases; the two eyes stand further apart; the three posterior pairs of legs have been developed, and a small abdomen has become distinctly separated from the thorax. Before proceeding to make a few additional remarks and corrections to my former description of the pupa, it will be advisable, on account of the importance of the subject, to discuss the homologies of the limbs.
From the presence of eyes and of two pairs of antennæ in the larva, during its earlier stages, the front of the head consists, in accordance with all analogy, of three segments; the mouth, likewise, from being formed of three gnathites (which can be detected by dissection in the pupal state), consists, also in accordance with all analogy, of three segments, making altogether six segments — on the nature of which I apprehend no objection will be raised. In two out of the three orders into which Cirripedes may be divided, the mouth is succeeded, in the adult animal, by eleven most distinct segments; of which the first (i. e. the seventh cephalic) differs from the succeeding seven thoracic segments; and these seven again differ from the three abdominal and terminal segments. Hence it must be admitted that, as far as the cephalo-thorax of the archetype Cirripede is concerned, it consists, like that of the archetype Crustacean, of fourteen segments, of which eight succeed the first-named six that form the mouth and front of the head; and that, with the three abdominal segments, there are altogether seventeen segments. In the order Thoracica, however, which includes all common Cirripedes, both in the pupa and in the mature animal, only six thoracic segments with their appendages, succeed the mouth, two having been lost; and the question arises which are these two, whether the seventh and eighth, or the thirteenth and fourteenth (i. e. the two terminal thoracic) segments; for there is no reason to suspect any other segments of having disappeared. In my former volume, I inferred, without sufficiently entering into my reasons, that it was the seventh and eighth, i. e. the last cephalic and first thoracic segments, which had disappeared; but I now find that Mr. Dana believes that, in ordinary Crustaceans, the abortion of the segments with their appendages almost always takes place at the posterior end of the cephalo-thorax. Nevertheless, after due deliberation and fresh examination of the pupa, I must retain my former opinion, that it is the last cephalic and first thoracic segments which have either coalesced with the others, or wholly disappeared. In the pupa, the mouth, although functionless, has its place most plainly marked by being slightly prominent, and by the presence of a sort of labrum and of a shrivelled œsophagus, round which latter the gnathites and the new œsophagus of the future young cirripede are in process of formation. Now between the mouth of the pupa and the first pair of natatory legs, there is a space of membrane, equalling, when stretched out, the three succeeding thoracic segments in length and breadth: this interspace, I conceive, must have some homological signification; here then we have at least an appearance of the abortion of appendages; whereas, at the posterior end of the cephalo-thorax, no such appearance is presented. Moreover this interspace of membrane is divided nearly in the middle by a most conspicuous fold, which, on the view here adopted, would mark the separation of the seventh (cephalic) from the eighth (thoracic) segment; and the interspace and fold are thus simply explained. Lastly, I have shown, in the Introduction (), that the first and five succeeding pairs of cirri of the mature Cirripede present certain small, but significant, resemblances in structure and in the origin of their nerves, with the outer pair of maxillipeds and with the five pairs of ambulatory legs in the Podophthalmia; which resemblances are all futile, if the cirri belong to the 7th, 8th, 9th, 10th, 11th, and 12th segments of the cephalo-thorax, or those immediately succeeding the mouth; but are full of meaning, if the six pairs of cirri belong, as I believe, to the 9th, 10th, 11th, 12th, 13th, and 14th segments, or the six posterior segments of the cephalo-thorax.
‘Crustacea: United States Exploring Expedition,’ .
Before commencing on details, I may premise that I have examined the pupa of Lepas australis, pectinata, fascicularis, and anatifera, of Conchoderma virgata, partially of Dichelaspis Warwickii, of Ibla quadrivalvis, and of Alcippe lampas; and in the Balanidæ, of Balanus balanoides and Hameri. In the pupæ of all these genera there is a most close general agreement in structure, excepting in minute details: I was surprised to find exactly the same slight differences in the spines on the first pair of natatory legs, as compared with the succeeding pairs, in Balanus Hameri, as in Lepas. The abdomen and caudal appendages of the pupa in the abnormal Alcippe, as we shall presently see, offer the only marked exception to this uniformity of character throughout the Thoracica. The outline of the carapace or shell is usually not so blunt at the anterior end, as in the pupa of Lepas australis (Pl. 30, fig. 2); more commonly the shape is that of the pupa of Alcippe (Pl. 23, fig. 16). In Lepas pectinata the two posterior points of the carapace are produced into two short spines. The surface of the carapace in L. australis is lined, as represented in fig. 4: the colour of this species when alive was blue: in L. fascicularis the surface is punctured: in L. pectinata it is marked with curious points of various shapes, often star-shaped, in parts reticulated, and confluent along the dorsal margin, and in parts lined: in B. balanoides it is very obscurely punctured, and in B. Hameri the punctures pass into lines. The whole of what is externally visible consists of the carapace, for this is produced not only backwards, so as to enclose the thorax and abdomen with their appendages, but also forwards, so as to overhang the whole front of the animal; and the prehensile antennæ, in Lepas, Ibla, Balanus, and probably in all the genera, can be retracted within its lower edge. The protection afforded by the carapace to the antennæ is aided by two crests (Pl. 30, fig. 7, c) parallel to this lower edge. The whole sternal surface is very narrow (fig. 4), and is likewise protected by the carapace; that is, when the two sides are drawn together by the adductor muscle. The shell, however, when thus drawn together, gapes a little at the two ends, at least in the case of Lepas australis. The adductor muscle, if introduced in fig. 4, would have crossed close anteriorly to the basal margin of the mouth; and in fig. 2, its end on the near side would have been attached under the dark cæca, which enter the upper end of the stomach. The adductor is shaped almost like an hour-glass, and so differs from this muscle in the mature Lepas, in which it is of the same thickness throughout. I may here add that the pupa of Lepas australis could swim very rapidly, and often on one side in a circle; it could walk by the aid of its antennæ, but often fell over; being thus locomotive, and, as we shall immediately see, well provided with senses, it cannot be considered as very lowly organised.
I took this species alive in the Southern Atlantic Ocean; and, mistaking it for an independent Crustacean, was much perplexed where to class it. I had overlooked these specimens when publishing my former volume.
Acoustic Organs. — Commencing at the anterior end, two small elongated orifices, 10/6000th of an inch in diameter, (e, fig. 4, Pl. 30), may be seen; these lead, as described in my former volume, into a sack, with a bag suspended in it, which is provided with a large nerve, and which I believe to be the acoustic vesicle. These orifices occur in the carapace, either in the same position, or a little more posteriorly, in the pupæ of all Cirripedes. In Balanus balanoides they are minute, being only 2/6000th in diameter, but are surrounded with a border: in Conchoderma virgata they are also surrounded by a border: in Lepas pectinata, the orifices are 3/6000th of an inch in diameter, and are very singular from being seated on rounded prominences, causing the carapace to have two short, blunt horns in front. In Lepas australis, and I believe in the other species, the corium round the acoustic orifices is darkly coloured; and these coloured marks can be distinguished for some little time on the peduncle of the young Cirripede, after the metamorphosis, and after the entire organ, together with the whole pupal carapace and eyes, has been moulted. Knowing the connection in the higher Crustacea, of the acoustic organs and the antennæ, and seeing the very backward position (figs. 2 and 4) of the one great pair of antennæ, I have always imagined that these orifices probably marked the normal position of the anterior pair of antennæ, which, since the earlier larval stages, have disappeared. And I now find that Schödler affirms, that in most, if not in all Daphnidæ, there is a black spot in front of the eye, which is connected with an opening in the basal portion of the anterior antennæ, and he concludes that it is an organ of hearing.
Quoted by Dana, ‘Crustacea of United States Exploring Expedition,’ .
Antennæ. — These, from their present position, and from standing, in their earlier stages whilst within their envelopes or horns, exteriorly to the small medial pair (since aborted), I believe to be the second pair; and this is Mr. Dana’s opinion. In my former description of these very singular and important organs (Pl. 30, figs. 4 and 8), I have fallen into some considerable mistakes: the two plates or segments (fig. 4, N), of which the posterior margins are inflected as apodemes (n), carrying the eyes, are certainly, as may be clearly seen in the pupa of Alcippe, Pl. 23, fig. 16, and as affirmed by Burmeister, the basal segments of the antennæ. The second or main segment (formerly called by me the basal segment) has in some species an upper portion of the membrane of which it is composed, next to the body, excessively thin, and separated from the rest of the membrane composing the segment, by an oblique line (fig. 8, o), which I mistook for its articulation with the body. We then come to the disc or third segment; and lastly to the fourth, or ultimate segment. This ultimate segment, generally, has its external corner projecting up, as a step; and this sometimes, as in Dichelaspis Warwickii, gives the appearance of its consisting of two segments; but a careful examination of this part in Ibla, in which the step-like structure is carried to an extreme, makes me believe that there is only one segment. The prehensile antennæ, therefore, like the natatory legs, are formed of four consecutive segments, of which the basal segments give rise to the singular apodemes, presently to be noticed (fig. 7), that carry the great compound eyes. This basal segment, in all Cirripedes, is moulted with the eyes, the three other segments invariably remaining cemented to the surface of attachment.
‘Beiträge zur Naturgeschichte der Rankenfüsser,’ . In tab. 1 of this work there are good drawings of the general structure of the pupa of a species of Lepas, probably L. australis. I believe this author was the first who made out the structure of the abdomen of the pupa.
In the table of measurements of the antennæ of the several genera and species of the Lepadidæ () of my former volume, the articulation, called by me basal, I now know to be really the articulation between the basal and second segment. In the fourth column, headed “Length from end of the disc to the inner margin of the basal articulation,” the term inner margin really applies to the oblique curved line separating the thin and scarcely visible membrane from the thicker membrane of that segment. These corrections do not in the least affect the object for which the table was given.
In a sketch, sent me by Mr. Dana, of this organ in the pupa of a Lepas from the Antarctic Ocean, I observe that he divides my ultimate segment into two segments.
In the Southern Atlantic I took some specimens of the pupa of Lepas australis, not yet attached, and therefore with the muscles of the antennæ, not having suffered any of that absorption, which they undergo, as soon as the pupa is permanently cemented to some floating object. In my former volume I noticed a pair of strong muscles, attached to the tips of the middle forks (Pl. 30, fig. 7) of the apodemes, and I now find two pairs attached to the bases of the two outer forks, and directed dorso-anteriorly; and two other pairs, also attached to their bases, but directed dorso-posteriorly, so that altogether there are five pairs of muscles attached to the apodemes; their chief function, I should think, was to draw the antennæ posteriorly and upwards within the carapace; but as the apodemes cannot be moved without the great compound eyes being likewise moved, the muscles probably serve a double purpose. When the pupæ were alive, I noticed that their eyes were constantly kept in a state of vibratory movement. Flexor and extensor muscles are attached at one end to the posterior margin of the basal segment, and at the other end to the second or main segment; other powerful muscles attached to this latter segment, are prolonged by ligaments into the disc. In Cryptophialus I observed that the disc-segment had a movement almost like that of the wrist. Whether any muscles enter the small terminal segment, I know not.
The drawing in Pl. 30, fig. 8, of part of the second segment, of the third or disc segment, and of the fourth or ultimate segment, in Lepas australis, is, I think, very accurate. The second segment articulates on the upper or dorsal surface of the disc, and has the articulation on one side constricted and formed of thin and flexible membrane; the little terminal segment, which is turned outwards at right angles, also, articulates on the disc. That the disc forms a true segment is shown clearly in Cryptophialus (Pl. 24, fig. 18), where the articulation with the second segment is not in such close contact. The disc is either circular, as in Lepas, or hoof-shaped, as in Ibla: in B. balanoides the disc is rather hollowed out on the inner side. It has the power of adhering even to so smooth a substance as glass, placed vertically. It is surrounded by a rim of transparent membrane. On the hinder margin some spines arise from the central and more opaque part: in Lepas australis, there are no less than seven of these spines (fig. 8): in Conchoderma virgata there are only four, in Scalpellum and Ibla only one. When the disc is placed on the surface of attachment, these spines lie parallel to it. The middle part of the disc is, almost always, nearly opaque; and in tracing the cement-ducts from within the body of the pupa, or of the young Cirripede, I in many cases traced them as far as this point, but here lost them. From this same obscure central part of the disc, in most, if not in all species of the Lepadidæ, spokes radiate, which sometimes are branched, and are not regular, not always even resembling each other on the opposite sides of the same individual. Round the proper membranous border of the disc, a second one may be observed (fig. 8, p), which differs in shape and extent in different specimens: under favorable circumstances, and very high powers, it may be seen to have a reticulated structure, and to be of a very pale brown colour; towards the exterior margin, the reticulations become finer, and are blended together and lost; on the inner margin, the substance forming this membrane may be seen to come out of the spokes. This substance is the cement, which has the power of adhering to whatever substance it grows against; and thus the disc of each antenna becomes cemented down, and soon both the antennæ are surrounded by a common border of cement, which gradually increases, after the metamorphosis, in extent. Occasionally the cement forms little projections, like short spines, on the edges opposite to the orifices of the spokes.
The small terminal segment usually bears on its truncated extremity six spines, some of which are occasionally hooked; in Scalpellum, two spines, rather longer than the others, are borne on a step some way down on the inner side of this segment; but in Lepas, two spines (fig. 8), very much longer than the others, arise from the outer corner of the extremity. These two are very different from the other four borne by this segment, or indeed any other spines on the body; for they are quite flexible, and are furnished with a double row of very long, straight, excessively fine hairs, which seem to be articulated on them — the whole presenting a very beautiful appearance. These spines are of considerable length, and in Conchoderma virgata they even equal in length the whole antenna. I can hardly doubt that these beautiful, plumose, flexible spines, into the thick bases of which the coloured corium could sometimes be seen to enter, serve as feelers. Owing to the facts immediately to be mentioned, I erroneously stated, in my former volume, that there were three long spines.
In three species of Lepas, in Dichelaspis Warwickii, and in Scalpellum Peronii, after having torn the lately-cemented antennæ from the surface of attachment, I observed proceeding from the end of the terminal segment, from between the above two groups of spines, what appeared to be a long narrow ribbon with its end torn off; and which, in the case of Lepas australis, I fancied was one of the plumose, long, flexible spines ripped open. But now that I have examined some of the pupæ of this species before their attachment, I find (as represented in Pl. 30, fig. 8, v′) a flattened tube, ending in a blunt point, and having a peculiar ringed structure. I have noticed similar appendages to the antennæ of specimens just attached of Lepas anatifera. In the Dichelaspis and the Scalpellum, the tube was very long, and seemed, from its torn appearance, to have been firmly attached to the supporting surface. In both these cases, the tube came out from within another slightly larger tube, which had been broken off close to the extremity of the terminal segment of the antenna. In the case of the Lepas anatifera, the tube expanded a little after leaving the antenna. In the Dichelaspis, it had exactly the same diameter as the cement-duct, which could be clearly distinguished within the two lower segments. From these several facts, and from the peculiar appearance of the tube itself, I believe it to be a tube of cement-tissue which thus, sometimes even before the pupa is attached, independently grows outwards. That the cement-tissue can grow outwards and assume definite forms, we know from the singular case of Lepas fascicularis, in which the cement proceeding from several apertures, forms a vesicular float round the peduncle of not only a single individual, but often of a group of specimens: we shall presently find a somewhat analogous fact in the case of Coronula. It is possible that this tube, proceeding from the extremity of the antenna, may be the channel through which cement continues to be poured forth during the continued growth of the above Cirripedes; and the manner in which this is effected, considering how firmly the end of the peduncle is cemented down, has always appeared to me a difficulty. In those pupæ of Lepas australis, which I caught swimming about unattached, it is surprising that the disc should have been edged with cement, and that a tube, similarly formed, should have grown out of the ultimate segment: it shows, I presume, that the cement-tissue will grow out, whether or no the pupa has succeeded in finding a proper object for attachment. Lastly, I have felt some surprise, in two or three instances in observing some dark purple pigment-cells, like those in the corium, within the terminal tube of cement; and likewise within the spokes of cement in the disc: this is the only fact which causes me the least doubt, whether I have rightly determined the nature of the terminal tube, as wholly formed of cement tissue; or whether it may not be covered by an outer integument, itself lined by true corium, coloured purple.
Finally, I may add, that, excepting in small details, the prehensile antennæ present no difference throughout the Order: I have minutely examined them in several genera of the Lepadidæ; and in the Balanidæ, I have seen them in Coronula, and in several species of Balanus. In B. balanoides I have examined them carefully; they are smaller and thicker than in Lepas, with the second or main segment bowed outwards, carrying its usual single spine; with the disc excised on its inner margin and apparently without the spoke-like vessels for the cement; and with the ultimate segment proportionably longer, and carrying, I believe, six spines, of which two appeared to be longer and more flexible than the other four shorter and somewhat hooked spines. In Coronula balænaris, also, the terminal segment is, proportionably to the others, of large size. Not only throughout the order, but throughout the whole Class, the antennæ are singularly uniform in structure, as will be seen, when the last two orders are described.