2
Darwin’s Toolkit

One of the most disturbing realizations of the 2016 American presidential election—for Democrats and Republicans alike—was the fragility of truth. Most of us are exhorted to tell the truth throughout our lives. We know that we sometimes fail, since lying can be personally advantageous in a thousand different ways, but we also know that if everyone lied then something precious that we all rely upon—trustworthy information—would be lost. That’s why most of us commit to telling the truth, expect to be punished when we lie, and punish other liars with zeal.
All that seemed to change with the 2016 American presidential election. Some people and organizations seemed to abandon the norm of truth-telling for themselves, wantonly generating “fake news” in order to achieve their own partisan ends. Worse, the societal response to lying seemed to break down. When fake news and other deceptions were brought to light—nobody was punished. In no time, fake news spread through the body politic like a cancer.
If you want a vision of hell on earth, you can’t do much better than a world where no facts can be trusted. Against this background, the norms of scholarship and science begin to look downright heavenly. Scholars such as Geoffrey Hodgson and Robert Richards, whom we met in the last chapter, and the many scientists we will meet in this book are not saints—but they have a strong personal commitment to finding the truth. Even more important, they are held accountable by truth-telling norms that are strongly enforced. To communicate as a scholar or a scientist, you must document everything that you say and your work must be reviewed by your peers before it can be published. Even minor transgressions, such as failing to cite previous relevant work, can damage your reputation. Flagrant lying, such as falsifying data, results in expulsion. In this sense, scholars and scientists regard Truth as their religion.1
That said, we don’t contemplate this often. We in academia and science are more like plumbers and carpenters on a day-to-day basis, showing up with a toolkit. We size up the job, reach for the appropriate tools, and get to work figuring out the facts of the matter. Some of the tools are physical objects, but there are also conceptual tools—certain ways of parsing information that are exceptionally insightful. In this chapter I will introduce you to the major conceptual tools in Darwin’s toolkit, which were developed mostly for the study of genetic evolution in nonhuman species but which can also be used to study human genetic and cultural evolution, including the formulation of public policy.
NIKO TINBERGEN AND HIS FOUR QUESTIONS
Niko Tinbergen (1907–1988) was a Dutch biologist who shared the Nobel Prize in medicine with Konrad Lorenz and Karl von Frisch in 1973 for pioneering the field of ethology, or the study of animal behavior.2 Back then, the idea that a behavioral trait such as aggression can evolve in the same way as an anatomical trait such as a deer’s antlers or a physiological trait such as glucose metabolism was not widely accepted. In his effort to establish ethology as a branch of biology, Tinbergen called attention to four questions that must be addressed to fully understand any product of evolution. These questions are the most important tools in an evolutionary scientist’s repertoire and are essential for understanding the arguments in this book.
First, what is the function of a given trait (if any)? Why does it exist compared to many other traits that could exist? Second, what is the history of the trait as it evolved over multiple generations? Third, what is its physical mechanism? All traits, even behavioral traits, have a physical basis that must be understood in addition to their functions. Fourth, how does the trait develop during the lifetime of the organism? Recognizing these as separate questions and studying them in conjunction with each other form the foundational concepts in Darwin’s toolkit.

Tinbergen pioneered the study of animal behavior.
For example, your hand is an adaptation that evolved by natural selection to grasp objects (its function). It evolved as part of the vertebrate lineage and is anatomically similar to the fins of fish (its history). Physically, it is composed of muscles, bones, tendons, and nerves put together in just the right way to grasp objects (its mechanism). It begins to appear as early as the fifth week of gestation (its development). A complete explanation of your hand requires answers to all four questions.
Fair enough, you might be thinking, but how can Tinbergen’s four questions be used as tools in a policymaker’s toolkit? Let’s take a look at each question in more detail.
WHAT IS THIS FOR?
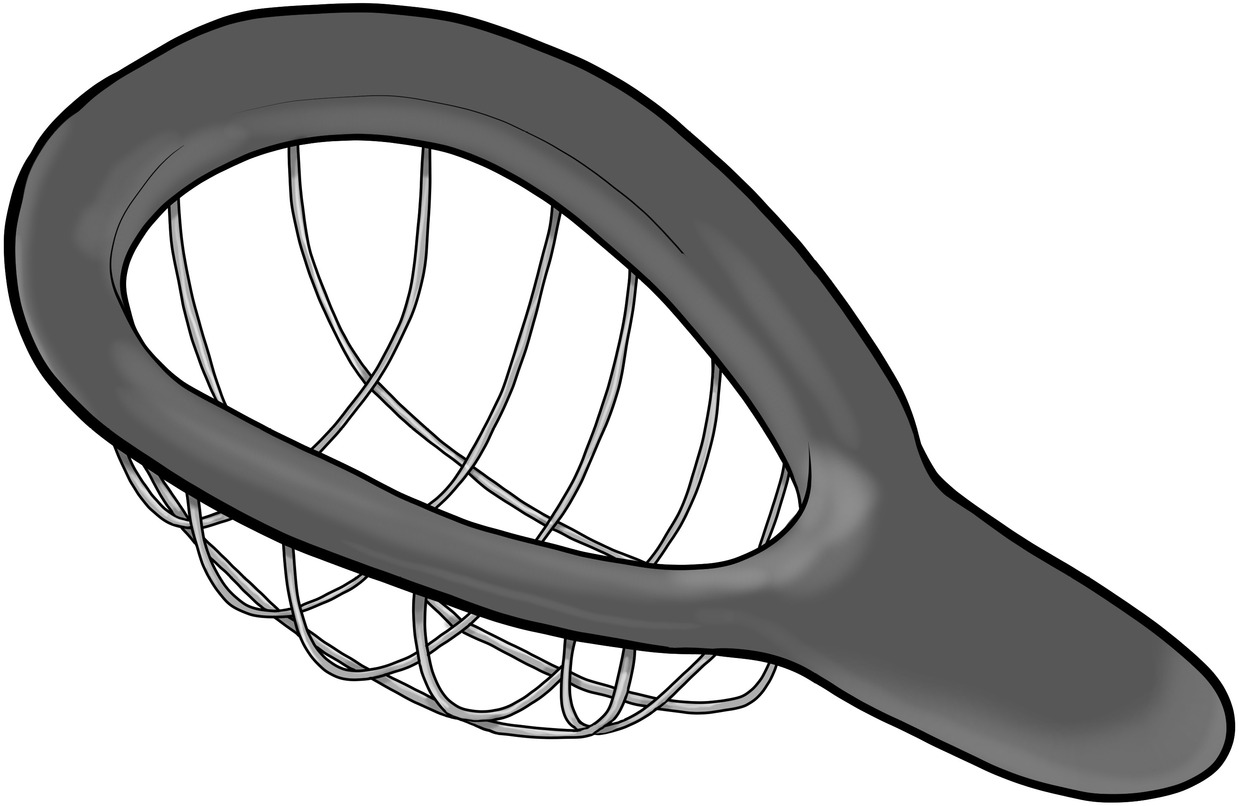
What is this for?
This gadget is clearly designed to do something, but what? Take a moment to think about it. Maybe it is a sieve, although most sieves don’t have such a large mesh size. Maybe it’s for sifting the poop from kitty litter, although I personally would like a longer handle for that job. Maybe it’s a well-ventilated cup for protecting the private parts of male athletes. Wrong, wrong, wrong. It’s an avocado cuber!
Whenever I play this game with an audience, a game show atmosphere develops. People who are normally shy about speaking in public eagerly raise their hands. There is a burst of laughter and satisfaction when they learn the right answer, which explains the details of the object so much better than the wrong answers. The size of the mesh and length of the handle are no longer a mystery. The overall shape (to conform to an avocado) and width of the rim (to push into the avocado) become important parts of the design, whereas before they might have appeared arbitrary.
Here’s another puzzle for you. What is a snowflake designed to do? The one shown here is much more intricate than an avocado cuber. Nevertheless, the correct answer to my question is that a snowflake isn’t designed to do anything. It wasn’t made by people and it isn’t a life form. Instead, it is the result of a physical process that takes place when water molecules crystallize around tiny particles in cold temperatures.
To say that a snowflake has no purpose doesn’t mean that it lacks value. Value is in the eye of the beholder, and I truly value the beauty of a snowflake, along with sunsets, cloudscapes, and the night sky. But none of these is designed for a purpose like a human implement or a life form that is a product of evolution. If you try to explain a snowflake or a physical process such as the weather as if they were designed for a purpose, confusion will result. If you try to explain a human-designed implement such as an avocado cuber or a biological adaptation such as a bird’s wing as if it is not designed for a purpose, confusion will also result.

A snowflake has beautiful structure but no function.
Here is another way to emphasize the crucial difference between an explanation that assumes functional design and one that doesn’t. Suppose that you’re walking up a mountain and you hear a crashing noise ahead of you. It’s a falling boulder that will crush you if you don’t move! You quickly calculate its trajectory and take note of objects that might change its path before you dart out of its way. That was a close call! Now suppose that the crashing noise is a charging grizzly bear rather than a boulder. If you thought about the bear the same way as you did a boulder, you would be dead. Avoiding the bear requires a completely different line of thought than avoiding the boulder.
Let’s call the way we think about avocado cubers, bird wings, and charging bears “functional reasoning mode” and the way we think about snowflakes, the weather, and falling boulders “physical reasoning mode.” All of us have a lifetime of experience reasoning in both modes, but we are also prone to misclassifications. We mistake a purely physical object such as a snowflake for a functionally designed object such as a bird’s wing, or vice versa. We correctly classify an object as functionally designed but attribute the wrong function to it, like mistaking an avocado cuber for an athletic cup. When these mistakes happen, we become as blind to reality as Darwin and Sedgwick were bumbling around that Welsh valley looking for fossils.
THE PRE-DARWINIAN WORLDVIEW
Mistakes took place on a grand scale prior to Darwin’s theory of evolution. According to the Christian worldview, the entire universe was created by a benign and all-powerful god. Everything from the tiniest insect to the stars in heaven plays a role in his grand plan. Even human suffering and the appearance of evil have a purpose that we must attempt to decipher. That’s why Malthus interpreted famine and disease as divinely imposed to teach virtuous behavior. Based on what we know today, most of these errors were the result of confusing purely physical processes for designed processes or attributing the wrong functions to designed objects.
The emergence of science during the Enlightenment started to correct some of these errors but left others alone. Isaac Newton stunned the world with his ability to predict the motions of the planets with mathematical equations, but he also saw purpose and design in the heavens. For him and most other Enlightenment thinkers, scientific inquiry provided a new way to understand God by studying his creation. Scientific knowledge often disagreed with biblical accounts, which led to a conflict between Enlightenment thinkers and the established church. Even the scientists, however, assumed that their inquiry would reveal an orderly and harmonious universe, a giant clock where every part contributes to the working of the whole.
The concept of laissez-faire in economic theory and practice is a direct descendant of Christian and Enlightenment thinking prior to Darwin. Laissez-faire refers to a policy or attitude of letting things take their own course without interfering. One use of the phrase in the early eighteenth century was “on laissez faire la nature,” or “let nature run its course,”3 a prescription that only makes sense if nature writ large is harmonious and self-regulating. Ecologists have largely abandoned the idea that nature left undisturbed achieves a harmonious balance.4 Instead, ecological systems are frequently out of equilibrium and can settle into many “basins of attraction” (locally stable configurations) that vary in their desirability from a human perspective. Wise ecological policy requires active management with complex systems in mind. The same is true for wise economic policy, but the pre-Darwinian concept of laissez-faire gets in the way of seeing this clearly.5 I will have much more to say about economic policy as we proceed. For now, the important point to make is that we still make massive errors in functional reasoning.
Darwin’s theory of evolution was a breakthrough in how to think properly about the presence and absence of functional design in the natural world. Virtually every trait that appears well designed in nature—the wing of the bird, the fang of the tiger, the speed of the gazelle—exists due to a historical process whereby individuals possessing those traits survived and reproduced better than individuals possessing different traits (Tinbergen’s history question). In evolutionary parlance, they were more fit. Enhanced fitness compared to alternative traits is the only source of design in nature.
This simple statement leads to one of the most powerful tools in Darwin’s toolkit, which is often called “adaptationist thinking” or “natural selection thinking.” If you want to understand the features of an organism in the same way as the features of an avocado cuber, ask how the organism would need to be structured to survive and reproduce in its environment (Tinbergen’s function question). That’s how I reasoned about the feeding behavior of copepods as a graduate student in the 1970s—my first introduction to natural selection thinking.
To say that adaptationist thinking is a fruitful line of inquiry does not mean that it is simple or leads directly to the correct interpretation of functional design. It also doesn’t mean that every detail of an organism is adaptive.6 Consider the red blood cells that are coursing through your veins. Their primary function is to carry oxygen to your other cells. They also happen to be colored red. Is their color part of their functional design? The question is not totally farfetched. Perhaps the color of blood has signal value in a social species such as our own. When we bleed, it is highly visible so that others can come to our aid. But if that were the case, then the color of blood would probably be different in species where individuals provide aid to each other, compared to less cooperative species. Since blood is red in both cooperative and non-cooperative vertebrate species, the social signaling hypothesis for its color is unlikely to be correct. The most judicious conclusion is that the color of blood has no function. It is just a by-product of the hemoglobin molecule’s main job of carrying oxygen. To advance further down this line of inquiry, it would be helpful to know more about hemoglobin as a physical molecule, which requires pivoting to Tinbergen’s mechanism question. Evolutionists form, test, and accept or reject hypotheses like this all the time as part of our job of arriving at the facts. It’s part of our conceptual toolkit.
As another example, consider the behavior of birds on remote oceanic islands. When Darwin visited the Galápagos Islands, he was surprised when birds alighted on his shoulders and arms as if he were a tree. They were well adapted to island life in the absence of people, but poorly adapted to the presence of people and other mammals such as mice, rats, goats, pigs, cats, and dogs. The moment that people and their mammalian companions set foot on remote oceanic islands, the environment changed radically for all of the native inhabitants and what was adaptive in their old environment became tragically maladaptive in their new one. Unless the native fauna and flora are protected by people or adapt to their new environment, they are likely to go extinct. Adaptations must be understood in terms of survival and reproduction in the historical environments that gave rise to the adaptations (Tinbergen’s history question). When the environment changes, there is no reason to expect organisms to be well adapted to their new circumstances, and generations are required for new adaptations to evolve.7
Evolutionary mismatches don’t just happen to other species. In many respects, we are like those bird species on oceanic islands, adapted to environments that have ceased to exist and sadly maladapted to our current environments. Three detailed examples will be provided in the next chapter. For now, I hope you are beginning to see how Tinbergen’s function and history questions—two important tools in the evolutionist’s toolkit—might also come in handy as tools in a policymaker’s toolkit.
LIVING SNOWFLAKES
If we want to understand the nature of a snowflake, there is only a physical process to consider. A snowflake is born when water molecules crystallize around a tiny airborne particle such as dust or pollen in freezing temperatures. Water molecules are shaped in a way that creates a hexagon when they join together to form a crystal. As the crystal grows, new molecules are added unevenly to the outer edge based on irregularities, creating arms that themselves provide edges for continuing growth. Since a typical snowflake includes an estimated 1019 water molecules (a mind-boggling number), every snowflake is indeed unique, as many of us are told as children. The fact that crystalline growth depends upon temperature and other weather conditions, with each snowflake experiencing different conditions as it falls toward earth, contributes to its uniqueness.
Understanding the formation and growth of a snowflake is an example of thinking in physical reasoning mode, based entirely on the interaction between the chemical properties of water and its physical environment. Functional reasoning is not needed and indeed would be misleading, since a snowflake is neither a human artifact nor a product of evolution.
For life forms and human artifacts, it is necessary to think in both physical and functional reasoning modes, which is why Tinbergen’s mechanism and development questions are needed to complement his function and history questions. Like a snowflake, this beautiful image of an anglefish skeleton is the product of a physical process that grows and develops over a period of time. Unlike a snowflake, living physical processes have been shaped by natural selection over the course of billions of years to replicate with astonishingly high fidelity—and the more one learns about the details, the more wondrous they become. The mechanistic branches of the biological sciences, such as biochemistry, molecular biology, genetics, and neurobiology, are devoted to revealing the physical basis of life.
It is remarkable how the function of an object and its physical basis can be studied independently of each other. Suppose I challenged you to make an avocado cuber out of physical materials that are completely different from the one shown in the illustration. You could probably meet my challenge without much difficulty. Likewise, consider the mundane fact that many desert-living species are sandy colored to conceal themselves from their predators and prey. This observation holds for desert-living snails, insects, amphibians, reptiles, birds, and mammals. The reason in each case is clear enough—individuals varied in their coloration and those that matched their background the best survived and reproduced the most. But wait! The exteriors of these species are made of completely different physical materials—calcium carbonate for snails, chitin for insects, keratin for reptiles, and so on. We didn’t need to know anything about the physical makeup of each species to predict that they should be sandy colored. As long as the physical makeup of each species results in heritable variation, then it becomes a kind of malleable clay to be shaped by selection.8

Unlike a snowflake, this angelfish skeleton has both beautiful structure and function.
It’s an exaggeration to say that species can evolve in any direction. There are constraints imposed by their genes and developmental pathways. Hence, some features of organisms can only be understood in terms of their physical makeup and not their functional design. Also, full understanding of any particular adaptation requires mechanistic understanding. We might not need to know that the shells of desert snails are made from calcium carbonate to predict that they should be sandy colored, but we will need to know that fact to understand how it happens, how it sometimes goes wrong, and so on.
A MASTER EVOLUTIONIST AT WORK
The best way to learn a craft is by watching a master craftsperson at work. I will therefore close my introduction to Darwin’s toolkit by showing how one master evolutionist combines Tinbergen’s four questions to study evolution in action.9 Richard Lenski, the son of the sociologist Gerhard Lenski and the poet Jean Lenski, was born in 1956. His interest in ecology and evolution caused him to specialize in the study of insects in their natural environments. Shortly after he received his PhD from the University of North Carolina in 1982, however, Lenski had a change of heart. If he really wanted to study fundamental issues in ecology and evolution, he should be studying a microbe named Escherichia coli.
Why? Primarily because E. coli, a species of bacteria that inhabits our guts, reproduces every 20 minutes under optimum growth conditions. That works out to 72 generations every day and 504 generations every week. If we pick a rough figure of 20 years for a human generation, then a week’s worth of evolution for E. coli (at its maximum growth rate) is the equivalent of over 10,000 years of evolution for us, which would take us back to the dawn of human civilization.
Richard Lenski: A master craftsman at work
Also, E. coli is one of the best-studied organisms in the world. Biologists have wisely selected a few species that they call model organisms, based on the strategy that it is better to know a whole lot about a few organisms than a little about a lot of organisms. The A-list of model organisms includes the bacteria E. coli, the nematode worm Caenorhabditis elegans, the fruit fly Drosophila melanogaster, the mouse Mus musculus, and the rat Rattus norvegicus. It is difficult to fathom the degree to which these model organisms have been examined from stem to stern by biologists, including a complete inventory of their genes and extensive knowledge of how the genes interact to assemble the whole organism (Tinbergen’s mechanism and development questions).
A microbe such as E. coli has another advantage that is truly extraordinary for evolutionary research. The cells can be frozen for extended periods and brought back to life by thawing them. This meant that Lenski could place E. coli in a novel laboratory environment, periodically freeze samples, and bring the ancestors back to life to compare with their own descendants. A living fossil record!
The experiment that Lenski started in 1988 was simplicity itself. A single genetically identical clone of E. coli was used to create 12 populations inhabiting conical glass beakers called Erlenmeyer flasks in the laboratory. Each flask contained 10 ml of liquid growth medium with glucose as the main energy source. The flasks were placed on a shaking incubator that kept the E. coli well mixed under constant temperature and light conditions. Every day without fail, 0.1 ml of medium from each flask was transferred to a fresh flask with 9.9 ml of fresh sterilized medium. The E. coli from the 12 populations were never mixed. Even though the flasks were arranged in neat rows and columns on the shaking incubator, they were as genetically isolated from each other as if they were oceanic islands thousands of miles apart.
The E. coli grew well in their new laboratory environment, firing off about seven generations every day before depleting their resources. Every 500 generations, a sample from the flasks was frozen to become part of the living fossil record. Although the procedure sounds simple, it required monklike devotion on the part of Lenski and his associates. The transfers had to take place every day without fail. Someone always had to be on duty. Backup flasks and frozen samples had to be maintained. Periodic checks for contamination were required. Provisions for electrical outages and equipment failures had to be made. Day after day after day, right up to the present (nearly 70,000 generations as of this writing). In human terms, that is the equivalent of our history as a species!
In each of the 12 flasks, genetic mutations arose that caused some E. coli to grow faster than others. These strains increased in frequency until they replaced the slower-growing strains. Lenski knows this with certainty because he can compare the growth rates of the descendants with their own ancestors by unfreezing the earlier samples. When they are grown side by side in a horse race, the descendants grow 60 percent faster than their ancestors (Tinbergen’s function and history questions).
Using the techniques of molecular biology, Lenski can also pinpoint the genetic changes that took place and how the new genes altered the anatomy and physiology of E. coli to adapt it to its laboratory environment (Tinbergen’s mechanism and development questions). Do you think that the genetic changes were the same or different in the 12 populations? The answer to this question is not obvious. On one hand, each population inhabited exactly the same chemical and physical environment, so whatever anatomical and physiological changes work for one population should work for the others. On the other hand, genetic mutations are chance events and it is unlikely that the exact same mutation would occur in the different populations (Tinbergen’s history question).
This combination of functional similarities (all strains good at digesting glucose) and genetic differences among the 12 populations is exactly what Lenski observed. The body size of E. coli increased in all 12 populations, for example, but the particular mutations that caused body size to increase in a mechanistic sense differed. There are many ways to skin a cat, and there are many genetic changes in E. coli that are functionally equivalent as far as increasing body size and other adaptations are concerned.
After the 12 populations had been isolated for 2,000 generations, Lenski started a new experiment while keeping the old experiment going. He switched the energy source for the 12 populations from glucose to another sugar called maltose. What do you think happened? All 12 populations were derived from a single clone and had adapted to the exact same laboratory environment with glucose as the energy source. Nevertheless, the populations varied greatly in how well they could grow on maltose (Tinbergen’s function question). They had genetically adapted to glucose as an energy source in different ways and these differences had consequences for how well they could grow on maltose. All of the populations were able to improve their growth rates on a maltose diet over the course of 1,000 generations, but their starting points were very different, unlike the start of the original experiment. Each population had become a distinctive genetic entity (Tinbergen’s history question).
Returning to the original experiment, you might think that the 12 populations would run out of ways of adapting to their unchanging laboratory environment. It’s true that they adapted more quickly at the beginning of the experiment, but they never reached a plateau. Even after many thousands of generations, new beneficial mutations arose to improve growth rate still further. These mutations might well have occurred during previous generations but failed to spread for lack of other genetic changes. In other words, as the 12 populations diverged, they provided different genetic backgrounds for subsequent mutations. The same mutation could be beneficial in one population and neutral or deleterious in other populations (Tinbergen’s mechanism and development questions).
After Lenski’s experiment had been in progress for fifteen years, something amazing happened. One population evolved the ability to digest a compound called citrate that had been present all along as part of the recipe of the growth medium. Citrate serves as an energy source for some bacteria but not for E. coli. For E. coli to start digesting citrate would be like for us to start digesting hay. Nevertheless, that is what one population started to do. Was this due to a once-in-a-trillion mutation that could have occurred in any of the 12 populations? Or had the genetic changes in one population during previous generations uniquely prepared it to adapt to an entirely new resource? Lenski and his associates were able to answer this question by reaching back into the frozen fossil record to a generation that preceded the evolution of the citrate-digesting mutation and restarting the experiment from that point. Sure enough, the same population that evolved the ability to digest citrate the first time did so again. Its unique genetic odyssey enabled it to do something that no population of E. coli had ever done before (Tinbergen’s history question).
Science often requires the use of physical tools. For a microbiologist, these include Erlenmeyer flasks, petri dishes, autoclaves, and the like, which came into use over a century ago. To these we can add the vastly more sophisticated tools of molecular biology that make it possible to read the genetic code letter by letter, surgically snip out genes and insert them into other genomes, and perform other wizardry. Training is required to master these tools, but it is a remarkable fact that Lenski didn’t start learning them until after he received his PhD. Retooling to become a microbiologist was little different from a carpenter retooling to become a plumber. Work is required but any motivated person can do it.
Far more important than the physical tools are the conceptual tools that Lenski used to design his experiments and interpret his results. First and foremost, he used adaptationist thinking to predict that E. coli would increase its growth rate in the laboratory environment that he created for them (Tinbergen’s function question). He could make this prediction without knowing anything about the physical makeup of E. coli because he could confidently assume heritable variation in growth rate. Since evolution is a historical process that relies upon chance mutations, he predicted that it would not take exactly the same course in each population and that the populations would increasingly diverge over time (Tinbergen’s history question). Studying the genetic changes that took place in each population required switching from functional reasoning mode to physical reasoning mode. Genes are physical entities and the symphony of genetic interactions that result in the development of an E. coli is a purely physical process, like the growth of a snowflake (Tinbergen’s mechanism and development questions).
Function. History. Mechanism. Development. These are the conceptual tools that Lenski learned as part of his PhD training in evolutionary biology, which he used to inform his research on insects before switching to E. coli. They are the conceptual tools that biologists use to study all aspects of all species, as Niko Tinbergen wisely noted. Once mastered they become second nature, like knowing how to ride a bicycle. They might seem so simple that they couldn’t be new—but only for those who have adopted evolution as their worldview. Now let’s see how the evolutionary toolkit can become part of the policymaker’s toolkit to address the problems of modern existence that everyone earnestly wants to solve.