4
The Problem of Goodness

The idea that harmony and order exist at large scales, such as human societies, the biosphere, and the physical universe, is deeply entrenched in Western thought and many other cultural traditions. It suffuses the Christian worldview and shows up in places that are not typically associated with Christianity, such as economics and complex systems science. In Christian thought, the belief that the universe was created by a benign and all-powerful god leads to a conundrum: If true, how can we explain the existence of evil? This is called the problem of evil and much ink has been spilled by theologians trying to resolve it—including Thomas Malthus’s belief that famine and disease are divinely imposed to teach virtuous behavior.
The evolutionary worldview turns the problem of evil on its head. The behaviors that we associate with evil are easy to explain from an evolutionary perspective, because they typically benefit the evildoer at the expense of others. The problem is to explain how the behaviors that people associate with goodness, which typically benefit others and society as a whole, can evolve by a Darwinian process. Much ink also has been spilled by evolutionary thinkers to resolve the problem of goodness, but they have made more progress than theologians, for the simple reason that evolution provides a better theory of living processes than creationism. In other words, we are in a position to provide a scientific account of how the behaviors associated with goodness can triumph over the behaviors associated with evil—or vice versa—depending upon environmental conditions.
In this chapter I will tell three stories about the eternal contest between good and evil and how we can tilt the playing field in favor of goodness. The first two stories are purely biological, although they are highly relevant to human welfare. The third story gets to the heart of what human morality is and how it can be strengthened. Before we begin, I must say a little more about the progress that evolutionists have made resolving the problem of goodness.1
MULTILEVEL SELECTION
Take a moment to imagine a morally perfect individual. Don’t worry too much about the precise definition of morality. Just trust your own intuition. What words would you use to describe him or her? Now take a moment to imagine that person’s opposite. What words would you use to describe evil incarnate? In short, what is your version of Dr. Jekyll and Mr. Hyde?

Robert Louis Stevenson’s story explores the eternal conflict between good and evil.
I have played this game with audiences around the world and all levels of expertise, from elementary school students to theologians and philosophy professors. The answers are so consistent that I list them in a slide that is part of my presentation, knowing it is what I am going to hear. Words used to describe the morally perfect individual include loving, honest, brave, giving, unselfish, loyal, and so on. Words used to describe the opposite include greedy, murderous, selfish, deceptive, manipulative, uncaring, and so on. Was I able to read your mind as well?
Darwin encountered a problem when he tried to explain how the traits associated with goodness can evolve by natural selection. In every case, they seem vulnerable to exploitation by the traits associated with evil. If natural selection favors individuals who survive and reproduce better than others, and if goodness involves helping others to survive and reproduce even at one’s own expense, then how can goodness evolve?
While goodness posed a problem for Darwin, the solution was not far to seek. Social behaviors are almost invariably expressed in groups that are small compared to the total evolving population—a fish school, a bird flock, a pride of lions, or a human tribe. This means that an evolving population is not just a population of individuals but also a population of groups. If individuals vary in their propensity for good and evil, then this variation will exist at two levels: variation among individuals within groups, and variation among groups within the entire population. While goodness might be vulnerable to evil within any particular group, groups whose members are loving, honest, brave, etc., to each other will robustly outcompete groups whose members are greedy, murderous, selfish, etc., to each other. Here is one of the passages from Darwin’s Descent of Man where he describes natural selection as a two-level process and relates it to human morality:
It must not be forgotten that although a high standard of morality gives but a slight or no advantage to each individual man and his children over other men of the same tribe, yet that an increase in the number of well-endowed men and advancement in the standard of morality will certainly give an immense advantage to one tribe over another. There can be no doubt that a tribe including many members who, from possessing in a high degree the spirit of patriotism, fidelity, obedience, courage, and sympathy, were always ready to aid one another, and to sacrifice themselves for the common good, would be victorious over most other tribes; and this would be natural selection. At all times throughout the world tribes have supplanted other tribes; and as morality is one important element in their success, the standard of morality and the number of well-endowed men will thus everywhere tend to rise and increase.
Darwin doesn’t comment on the glaring fact that in his scenario, moral behaviors are confined to members of one’s own tribe and are often directed against members of other tribes. Group selection doesn’t eliminate immoral behaviors so much as it elevates them to the level of between-group interactions. Altruism within groups can become a form of collective selfishness toward other groups. Nevertheless, we need to explain how within-group morality can evolve before we attempt to tackle more expansive forms of morality.
In a 2007 review article that I wrote with the famed evolutionary biologist Edward O. Wilson, we summarized Darwin’s theory of two-level selection this way:
Selfishness beats altruism within groups. Altruistic groups beat selfish groups. Everything else is commentary.2
While this solution to the problem of goodness is simple, the ramifications are far-reaching. In the first place, for within-group morality to evolve, between-group selection pressures must be stronger than within-group selection pressures. There is no guarantee that this will always be the case. When within-group selection is the stronger force, then evil triumphs over good. There is no warrant for a worldview that claims that nature writ large embodies goodness. Goodness only emerges when between-group selection pressures outweigh within-group selection pressures.
In the second place, nature is more complicated than two-level selection, or a hierarchy of only individuals within groups. Individuals are themselves groups of cells and genes. Single-species social groups such as fish schools, bird flocks, and lion prides exist in ecosystems composed of many species and a nested hierarchy of scales, ultimately making up the whole biosphere. In the human world we have genes, individuals, families, villages, cities, provinces, and nations, all nested within what Marshall McLuhan dubbed the Global Village. The tug-of-war between levels of selection that I described for individuals in groups exists for all levels. What’s good for me can be bad for my family. What’s good for my family can be bad for my clan—all the way up to what’s good for my nation can be bad for the Global Village.
In short, two-level selection needs to be expanded into a theory of multilevel selection (MLS), from genes to the planet. The problem of goodness can potentially be resolved at any level, but the conditions for higher-level selection to prevail over lower-level selection become more challenging as we ascend the rungs of the multitier hierarchy, as we shall see with my three stories of the eternal contest between good and evil.
THE EVIL OF CANCER
Each of us consists of trillions of cells that are differentiated into hundreds of cell types. Every cell is descended from a parent cell, all the way back to the original sperm and egg. Each cell division requires copying the DNA, which consists of thousands of genes and roughly four billion base pairs—the nucleotide “letters” that make up the genetic “alphabet.”
With a few exceptions, all of our cells have the same genes. The way that cells differentiate is by expressing some genes and silencing others. When differentiated cells divide, the patterns of gene expression must be copied in addition to copying the entire complement of genes. The inheritance of patterns of gene expression is called epigenetics.3
Some of our cells have long lifetimes. Many of the cells in your brain or in your ovaries (if you are a woman) were present when you were a child. Other cells have a lifetime of weeks or even days. If you are a man, for example, the sperm cells in your testes are no more than a few weeks old. Skin cells, liver cells, the cells that line our gut, and the cells that make up our immune system have especially high turnover rates. An estimated 500 billion cell divisions take place in our bodies every day!
Cells must divide and differentiate in just the right way to play their appointed role in a multicellular organism. They must express only some genes and not others. They must stop dividing when an organ is fully developed. Cells even routinely commit suicide (called “programmed cell death”) when their absence benefits the organism more than their presence. This symphony of cooperation is produced by natural selection. Very simply, organisms whose cells work together for the common good survive and reproduce and their properties are inherited by their offspring more often than organisms whose cells fail to do their part. The result is a living snowflake: a purely physical process (addressed by Tinbergen’s mechanism and development questions) that—unlike real snowflakes—results in a reliable working copy time after time.
The replication of genes and patterns of gene expression during every cell division are amazingly accurate, but not perfect. Imagine that every book had to be transcribed by hand from another book, which was true before the invention of the printing press. Even the most careful transcriber is likely to make a few errors, which will then be perpetuated in future copies. So it is with our cells. With billions of base pairs that need to be replicated, almost every new cell contains a few copying errors, or mutations.
Some mutations don’t make any detectable difference in the performance of the cell or organism. Others impair the performance of the cell without harming the organism. In fact, one reason that programmed cell death evolved is to rid the body of mutated cells. Still other mutations cause the cell to grow inappropriately at the expense of neighboring cells and to evade programmed cell death. The tissues that form from these cells are called neoplasms.
Neoplasm cells are not good for the organism but they are perversely good for themselves. After all, natural selection favors any entity that survives and reproduces better than neighboring entities. Neoplasm cells fit that description. It doesn’t matter that they might turn into malignant tumors that result in the death of the organism, and therefore their own death. Natural selection has no foresight. It is simply a physical process of replacement that takes place on the basis of differential survival and reproduction in the here and now, regardless of the long-term consequences. The only way that long-term consequences become salient at the cell level is through a process of natural selection operating at the level of multicellular organisms, which can be thought of as group selection among tribes of cells.
Neoplasms are much more common than malignant cancers. In fact, we are only in the process of discovering how common. Look at this image of an eyelid. It is not a new eye shadow. It is a pictorial of neoplasms on the eyelid of a person between fifty-five and seventy-three years of age. Each circle represents a mutated skin cell that grew at the expense of the normal skin cells to create a tiny patch of its own kind. This person did not have skin cancer, but rather a condition called drooping eyelid that is corrected by surgically removing some of the skin of the eyelid, affording an opportunity for a team of scientists led by Peter Campbell at the University of Cambridge and the Wellcome Trust Sanger Institute in Britain to conduct a genetic analysis of skin samples like this at a fine spatial scale.4 They estimated that each skin cell had an average of about four mutations and over 20 percent of the cells had mutations associated with skin cancer. These are the cells that tended to grow at the expense of neighboring cells to form tiny neoplasms.
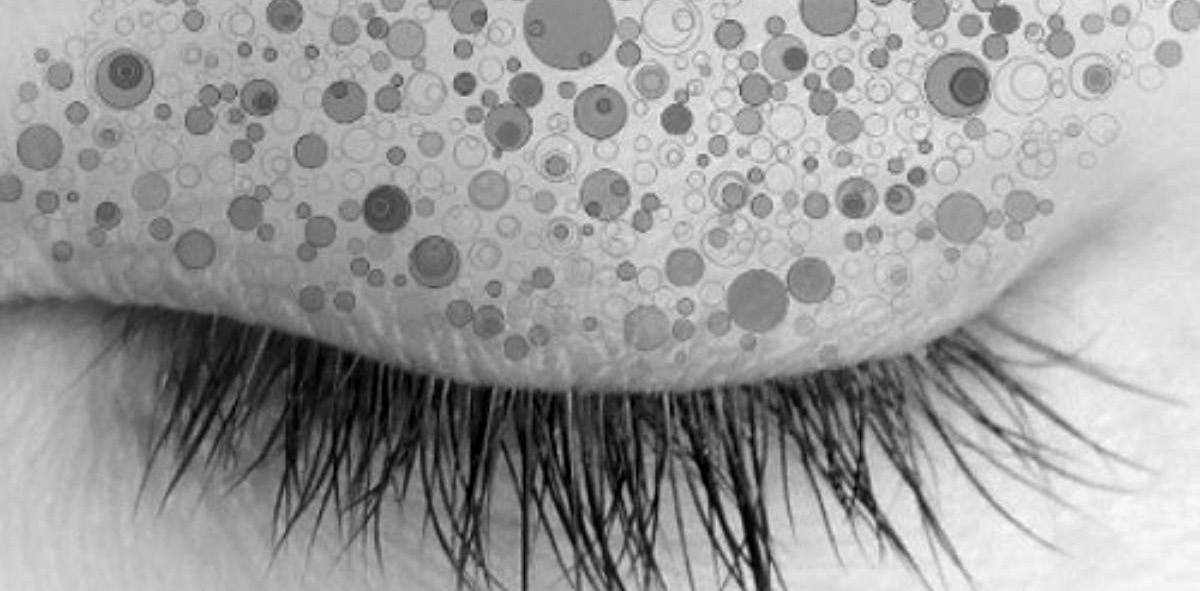
Each dot on this person’s eyelid represents a group of mutant cells that has proliferated at the expense of the normal cells.
Each neoplasm is like one of the populations in Richard Lenski’s E. coli experiment. Instead of being selected to digest glucose, they are selected to grow at the expense of neighboring cells. Additional mutations are required to turn a neoplasm into life-threatening cancer. The right combination of mutations to develop cancer only occurs in a small fraction of neoplasms. The rest are benign and do not threaten our health.
One key adaptation of a malignant cancer is to increase the rate of mutations. A fast-growing tumor isn’t just one type of cancer cell that is rapidly proliferating. It is a boiling cauldron of hyper-mutating cell lines that compete against each other. Another adaptive strategy for a cancer cell is to disperse so that it can compete against normal cells rather than against other cancer cells—metastasis.
Cancer provides an example of multilevel selection and the eternal contest between the behaviors associated with good and evil. With cancer, the group is the multicellular organism and the individuals are the cells. Just as the traits associated with good are vulnerable to the traits associated with evil for organisms living in social groups, normal cells are vulnerable to cancer cells within multicellular organisms. In the same way groups of morally virtuous individuals outcompete groups crippled by selfishness, multicellular organisms free of cancer outcompete multicellular organisms riddled by cancer. The eternal struggle between good and evil takes place within our own bodies and has since the origin of multicellular organisms roughly a billion years ago.
Not only does cancer provide an excellent example of multilevel selection, but it also illustrates how the theory decides what we can observe. Cancer research is enormously sophisticated, but almost all of it is concentrated on Tinbergen’s mechanism and development questions. The very concept of cancer as natural selection taking place within multicellular organisms wasn’t proposed until the 1970s and still accounts for a tiny fraction of cancer research.5 A fully rounded four-question approach, one that includes the function and history questions, challenges the wisdom of some current practices and suggests new possibilities for cancer treatments that were previously invisible.
Take the current practice of aggressive chemotherapy, which has the goal of entirely eliminating a tumor. This might make sense if all of the tumor cells are alike, but if they are a rapidly mutating and evolving population of cells, then aggressive chemotherapy becomes an extremely strong selection pressure for tumor cells that are resistant to chemotherapy. We already know that trying to blast bacterial diseases with antibiotics merely results in the selection of resistant strains that become ecologically dominant because other species in the ecosystem, such as predators and competitors, are completely wiped out. A more promising approach, informed by evolution and ecology, is to keep the ecosystem intact and enlist the aid of other species to help control the pest or disease species. A similar approach for cancer, led by the few researchers who are employing an evolutionary and ecological perspective, is called adaptive therapy.
Since the risk of cancer increases with every cell division, we might expect large and long-lived species such as elephants to be more prone to cancer than small and short-lived species such as mice. But this isn’t the case—mice and elephants have roughly the same cancer rates. The reason appears to be that cancer is a greater selection pressure for elephants, so they have evolved more effective defenses against cancer than mice have. We should be studying large and long-lived species to learn how they suppress cancer so well and whether we can base therapies for humans on the same mechanisms. Cross-species comparisons, which rely heavily on Tinbergen’s history question, provide a rich vein of information on cancer from an evolutionary perspective, but almost all cancer research is conducted on only two species, humans and laboratory mice—more blindness from the failure to employ the right theory.
Cancer from an evolutionary perspective is a fascinating and important topic in its own right and provides a great example of the contest between good and evil in unexpected places. Now let’s see what chickens can teach us about the problem of goodness.
GOOD AND EVIL CHICKENS
In chapter 1 we met Francis Galton, Darwin’s half cousin, who thought that people should be bred for their abilities in the same way that domesticated plants and animals are. Even Darwin thought that eugenics might work for humans, objecting only because he thought it would violate our instincts for sympathy and compassion, which he regarded as important human adaptations.
With this in mind, consider an experiment that was performed on chickens by William Muir and his colleagues at Purdue University’s Department of Animal Sciences in the 1990s.6 Their goal was to increase the egg-laying productivity of hens. Chickens evolved to live in flocks, but in the modern poultry industry they are often housed in cages with five to nine hens per cage, so the study was focused on maximizing production in this environment. The design was simple: they monitored how many eggs each hen laid, the most productive hens from each cage were used to breed the next generation, and so on for a number of generations. If the trait of egg productivity is heritable, then this method should substantially increase egg productivity over a number of generations, much in the same way that later generations of E. coli in Richard Lenski’s experiment evolved to digest glucose more efficiently.
But that’s not what happened. Instead, the subsequent generations laid fewer eggs and became more aggressive toward each other. The image opposite shows one of the cages after the experiment had been in progress for five generations. The cage originally housed nine hens, but six were murdered and the survivors had plucked each other’s feathers. No wonder they weren’t laying many eggs!
What happened to produce this ghoulish result? The most productive hens in each cage achieved their productivity by bullying the other hens. Bullying behavior is heritable in chickens, so selecting the biggest bullies led to a strain of hyper-aggressive hens within five generations. The energy expended and stress induced by their constant attacks on each other caused all of them to lay fewer eggs, despite the fact that the most productive had been selected to breed, generation after generation.
In an experiment that was performed in parallel with the first, egg productivity was monitored at the level of whole cages. Instead of breeding the most productive hens within each cage, all of the hens from the most productive cages were selected to breed the next generation. The image on page 86 shows a cage from this experiment after five generations. All nine chickens are alive and fully feathered, and their egg productivity increased 160% during the course of the experiment.

This is what happened when the “best” egg layer within each group was selected to breed the next generation.
These two experiments provide beautiful examples of within-group and between-group selection as envisioned by Darwin. The first one highlights the advantage that selfish traits have over cooperative traits within single groups. The chickens in the first image, tormenting and killing each other for their own gain, exhibit traits we would certainly call evil. The second experiment highlights the need for selection at the level of groups to evolve the traits that enable everyone within the group to thrive. The chickens in the second image, living amicably with each other, exhibit behaviors we would certainly call good.
Francis Galton assumed a simple relationship between individual abilities and societal warfare. Able societies are built by able individuals. Ability is an individual trait that is inherited by offspring from their parents. Selecting the most able individuals therefore must result in the most able society.

This is what happened when all hens from the best groups were selected to breed the next generation.
But the chicken experiments suggest that this logic is flawed—even for farm animals where eugenics is a common practice. It seems Francis Galton was deeply mistaken about the relationship between individual abilities and societal welfare. The number of eggs laid by an individual hen is not an individual trait so much as it is a social trait, because it depends upon how members of the group behave toward each other. If the individuals who profit most from a social group do not contribute to the group’s welfare, and if their traits are heritable, then selecting for them results in the collapse of the society. The relationships between single genes, observable traits measured in individuals, and the performance of whole groups are sufficiently complex—even for caged chickens—that selecting at lower levels often fails to produce the expected group-level outcomes. Selecting whole groups on the basis of their success proves to be more effective because the next generation will inherit some combination of all the individual traits that, in previous generations, resulted in fruitful interactions and contributed to the success of the group.
I will use the chicken experiments as a parable for human social interactions throughout the rest of this book, but for now let’s examine the implications for agriculture and animal welfare, which are important policy areas in their own right. The most common commercial method of rearing hens is to cram them into standing-room-only cages, where they are unable to escape each other or perform their normal behaviors. Their beaks must be “trimmed” to prevent them from injuring each other, and their bones break from lack of use. If that’s not injurious to animal welfare, what would be? Many people (including myself) are willing to pay more for eggs from hens that are allowed to range more freely, but free-range social environments have their own problems. Fighting still takes place and dominant birds prevent subordinate birds from accessing food, water, and nesting sites, resulting in low productivity for the group as a whole. Providing more space does not solve the problem of evil triumphing over good within groups, for chickens any more than ourselves!
A final lesson is that genetic evolution will take place in domesticated animals and plants, whether we want it to or not. It is sobering to contemplate the malleability of life. It only takes five generations to turn a population of mild-mannered chickens into a population of psychopaths. If we don’t manage evolutionary processes, they will very likely take us where we don’t want to go.
Thus ends my second story of the eternal contest between good and evil in unexpected places. Now it is time to bring the concepts to bear upon our own species.
WHAT IS MORALITY, ANYWAY?
At the beginning of this chapter, I wrote that there is no warrant for the deeply entrenched view that harmony and order exist at all scales of nature. We can expect harmony and order only when higher-level selection manages to prevail against lower-level selection. In animal societies, there are many cases of natural selection going the way of the first chicken experiment. The traits that we associate with evil triumph over the traits that we associate with good within groups, and the counterforce provided by between-group selection is not strong enough to save the day. These are “life’s a bitch and then you die” societies. We would not want to live in them.
There are also cases where natural selection went the way of the second chicken experiment. Between-group selection is strong enough to prevail against within-group selection, favoring the traits that we associate with goodness. Many social species are mosaics of both kinds of traits, some maintained in the population by within-group selection, others by between-group selection. However, the balance between levels of selection is not static but can itself evolve. In rare cases, mechanisms evolve that largely suppress the potential for disruptive forms of selection within groups, making between-group selection the primary evolutionary force for most traits of the species. Then something magical happens: the group evolves to be so cooperative that it is transformed into a higher-level organism in its own right.
This transformation is called a major evolutionary transition, and it was first proposed in the 1970s by the cell biologist Lynn Margulis to explain how nucleated cells evolved from bacterial cells.7 The former are much more complex, with internal structures such as mitochondria, chloroplasts, and ribosomes that are called “organelles” because of their specialized roles in maintaining the cell, similar to the organs of our body. Despite the difference in their complexity, it was assumed as a matter of course that nucleated cells must have evolved by small mutational steps from bacterial cells. Margulis’s radical proposal was that nucleated cells originated as cooperative communities of bacterial cells. Decades were required for her symbiotic cell theory to be accepted, but now it is widely regarded as fact.
The idea that individuals can evolve from groups rather than from other individuals was then generalized in the 1990s by two theoretical biologists, John Maynard Smith and Eörs Szathmáry, to include the evolution of the first bacterial cells, the evolution of multicellular organisms, and the evolution of insect colonies.8 In all cases, the higher-level entity evolves the properties of an organism by suppressing the potential for disruptive selection from within. Even the origin of life itself might be explained in this way as groups of cooperating molecular interactions.9
Multicellular organisms and their ability to suppress cancer provide some of the best examples of major evolutionary transitions, as we saw with my first story. Most genes that evolve in multicellular organisms do so by benefitting the whole organism relative to other organisms (or groups of organisms relative to other groups). It is comparatively rare for genes to evolve at the expense of other genes within the same organism, but only thanks to elaborative mechanisms that evolved to ensure that this does not happen. Higher-level selection is so much stronger than lower-level selection that we use a different word to describe the higher-level entity. Instead of calling it a society of cells, we call it an organism. This name change should not obscure the fact that a multicellular organism is nothing more than a highly regulated society of cells that evolved thanks to a very strong imbalance between levels of selection.
Social insect colonies such as ants, bees, wasps, and termites provide another outstanding example of major evolutionary transitions.10 Unlike a multicellular organism, which has a clear physical boundary, the members of a social insect colony are physically separate from each other. On any given day, the honeybees from a single beehive can be dispersed over an area of several square kilometers. Nevertheless, their activities are so well coordinated that they invite comparison to a single multicellular organism. When searching for a new nest site, for example, a swarm of honeybees is every bit as discerning as a human house hunter, searching out and evaluating the alternatives on the basis of multiple criteria such as size, height, and exposure to the sun.11 Words such as “eusocial,” “ultrasocial,” and “superorganism” are used to designate their adaptedness at the level of the colony, based on the fact that between-colony selection is the dominant evolutionary force.
People have been fascinated by the social insects since antiquity. Even though we are unlike them in so many ways, we feel an affinity for them and even treat their industry on behalf of their groups as an ideal for us to emulate. We also sometimes express an affinity and yearning for our societies to be like a multicellular organism, as in this passage from a seventeenth-century religious tract:
True love means growth for the whole organism, whose members are interdependent and serve each other. That is the outward form of the inner working of the Spirit, the organism of the Body governed by Christ. We see the same thing among the bees, who all work with equal zeal gathering honey.12
These comparisons are metaphorical, but now they can be placed on a firm scientific foundation. We are evolution’s most recent major transition. Almost everything that sets us apart from other primate species can be explained as forms of cooperation that evolved by between-group selection, thanks largely to our ability to suppress disruptive within-group selection. In most primate societies, group members are cooperative to a degree but are also riven by within-group conflict. Even the cooperation that exists often takes the form of coalitions warring with other coalitions within the same groups. To the best of our current knowledge, our distant ancestors evolved the ability to suppress bullying and other disruptive self-serving behaviors within groups, like multicellular organisms evolved ways to suppress cancer cells, so that the primary way to survive and reproduce was through teamwork.13

Beehives have long been used as a symbol of cooperation and industry.
This brings us back to the topic of morality. At the beginning of this chapter, when I asked you to imagine the traits associated with good and evil, I encouraged you to trust your intuition and not to worry about formal definitions. Now we can do better. Few people are more authoritative on the subject than Simon Blackburn, who holds the Bertrand Russell Chair of Philosophy at Cambridge University in the UK. In an interview with Blackburn, I asked him to define morality as he would to students in a Philosophy 101 class, without reference to evolution. Here is what he said.
At its simplest, morality is a system whereby we put pressure on ourselves and others to conform to certain kinds of behavior. That’s the side of morality which is perhaps most obvious. It’s associated with rules, with boundaries to conduct, and limits to criminal behavior when those rules are transgressed. On top of that, there is an element of morality that concerns our sentiments and emotions; for example our capacity to feel sympathy for each other’s distress and a corresponding motivation to do something about it. So there are two sides to morality; one of them more gentle and humane and the other more coercive and to do with rules and social institutions designed to enforce those rules. For analytical purposes it is useful to separate them but in many contexts they merge into one another. For example, our sympathy for the distress of somebody being bullied might translate into a belief that the bully has transgressed social norms and the desire to punish them or to alleviate the distress in some way.14
This definition of morality, stated without reference to evolution, is exactly the system that we would expect to result from a major evolutionary transition. Our moral psychology is the societal equivalent of cancer-suppressing mechanisms in multicellular organisms. The coercive side of morality is required to suppress the potential for disruptive self-seeking behaviors within groups. Once the coercive side is established, then it becomes safe for group members to freely help each other without fear of exploitation.
This correspondence, between morality as already understood and morality as expected from an evolutionary perspective, was not lost on Blackburn. The rest of our interview explored the insights that can be gained from a more explicit study of human morality from an evolutionary perspective. No other theory comes close to explaining our odd mix of moral strengths and weaknesses: our intuitive sense of right and wrong; our virtuous behaviors and temptations to cheat; our zeal for monitoring and punishing the transgressions of others; and the ease with which we confine our virtuous behaviors to “us” and exclude “them.” The more we see the problem of goodness through the lens of the right theory, the more we will be able to construct moral communities that are adapted to our modern age.