Forests are complex ecological systems in which trees are the dominant life-form. Together, forests of all types cover nearly 30 percent of Earth’s land surface. Tree-dominated forests can occur wherever the temperatures rise above 10°C (50°F) in the warmest months and the annual precipitation is more than 200 mm (8 inches). They can develop under a variety of conditions within these climatic limits, and the kind of soil, plant, and animal life differs according to the extremes of environmental influences. In cool, high-latitude subpolar regions, forests are dominated by hardy conifers like pines, spruces, and larches. These taiga (boreal) forests have prolonged winters and between 250 and 500 mm (10 and 20 inches) of rainfall annually.

Reindeer (Rangifer tarandus), such as this grazer looking for grass near Torinen, Sweden, are well adapted for life in taiga forests in northern Europe and Asia. Olivier Morin/AFP/Getty Images
In more temperate high-latitude climates, mixed forests of both conifers and broad-leaved deciduous trees predominate. Broad-leaved deciduous forests develop in middle-latitude climates, where there is an average temperature above 10 °C (50 °F) for at least six months every year and annual precipitation is above 400 mm (16 inches). A growing period of 100 to 200 days allows deciduous forests to be dominated by oaks, elms, birches, maples, beeches, and aspens. In the humid climates of the equatorial belt, tropical rainforests develop. There heavy rainfall supports evergreens that have broad leaves instead of needle leaves, as in cooler forests. In the lower latitudes of the Southern Hemisphere, the temperate deciduous forest reappears.
Forest types are distinguished from each other according to species composition (which develops in part according to the age of the forest), the density of tree cover, types of soils found there, and the geologic history of the forest region.

Mixed evergreen and hardwood forest on the slopes of the Adirondack Mountains near Keene Valley, New York. Jerome Wyckoff
Soil conditions are distinguished according to depth, fertility, and the presence of perennial roots. Soil depth is important because it determines the extent to which roots can penetrate into the earth and, therefore, the amount of water and nutrients available to the trees. The soil of taiga forests is sandy and quickly drained. Deciduous forests have brown soil, richer than sand in nutrients, and less porous. Rainforests and savanna woodlands have a soil layer rich in iron or aluminum, which give the soils either a reddish or yellowish cast. The amount of water available to the soil, and therefore available for tree growth, depends on the amount of annual rainfall. Water may be lost by evaporation from the surface or by leaf transpiration. Evaporation and transpiration also control the temperature of the air in forests, which is always slightly warmer in cold months and cooler in warm months than the air in surrounding regions.
The density of tree cover influences the amount of both sunlight and rainfall reaching every forest layer. A full-canopied forest absorbs between 60 and 90 percent of available light, most of which is absorbed by the leaves for photosynthesis. The movement of rainfall into the forest is considerably influenced by leaf cover, which tends to slow the velocity of falling water, which penetrates down to the ground level by running down tree trunks or dripping from leaves. Water not absorbed by the tree roots for nutrition runs along root channels, so water erosion is therefore not a major factor in shaping forest topography.
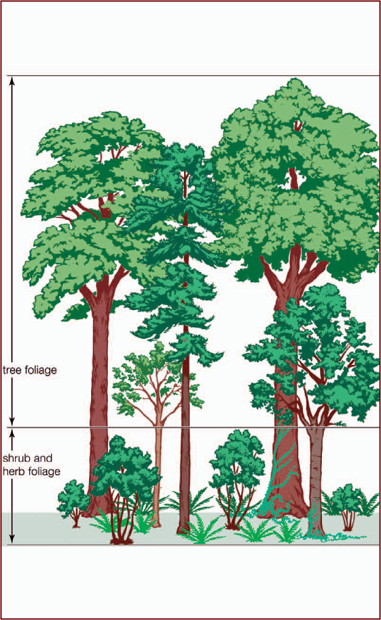
Forests are among the most complex ecosystems in the world, and they exhibit extensive vertical stratification. Conifer forests have the simplest structure: a tree layer rising to about 30 metres (98 feet), a shrub layer that is spotty or even absent, and a ground layer covered with lichens, mosses, and liverworts. Deciduous forests are more complex; the tree canopy is divided into an upper and lower story, while rainforest canopies are divided into at least three strata. The forest floor in both of these forests consists of a layer of organic matter overlying mineral soil. The humus layer of tropical soils is affected by the high levels of heat and humidity, which quickly decompose whatever organic matter exists. Fungi on the soil surface play an important role in the availability and distribution of nutrients, particularly in the northern coniferous forests. Some species of fungi live in partnership with the tree roots, while others are parasitically destructive.
Animals that live in forests have highly developed hearing, and many are adapted for vertical movement through the environment. Because food other than ground plants is scarce, many ground-dwelling animals use forests only for shelter. In temperate forests, birds distribute plant seeds and insects aid in pollination, along with the wind. In tropical forests, fruit bats and birds effect pollination. The forest is nature’s most efficient ecosystem, with a high rate of photosynthesis affecting both plant and animal systems in a series of complex organic relationships.
A rainforest is a luxuriant forest, generally composed of tall, broad-leaved trees and usually found in wet tropical uplands and lowlands around the Equator.
Rainforests usually occur in regions where there is a high annual rainfall of generally more than 1,800 mm (70 inches) and a hot and steamy climate. The trees found in these regions are evergreen. Rainforests may also be found in areas of the tropics in which a dry season occurs, such as the “dry rainforests” of northeastern Australia. In these regions annual rainfall is between 800 and 1,800 mm (31 and 70 inches) and as many as 75 percent of the trees are deciduous.
Tropical rainforests are found primarily in South and Central America, West and Central Africa, Indonesia, parts of Southeast Asia, and tropical Australia. The climate in these regions is one of relatively high humidity with no marked seasonal variation. Temperatures remain high, usually about 30 °C (86 °F) during the day and 20 °C (68 °F) at night. Where altitude increases along the borders of equatorial rainforests, the vegetation is replaced by montane forests, as in the highlands of New Guinea, the Gotel Mountains of Cameroon, and in the Ruwenzori mass of Central Africa. Tropical deciduous forests are located mainly in eastern Brazil, southeastern Africa, northern Australia, and parts of Southeast Asia.
Other kinds of rainforests include the monsoon forests, most like the popular image of jungles, with a marked dry season and a vegetation dominated by deciduous trees such as teak, thickets of bamboo, and a dense undergrowth. Mangrove forests occur along estuaries and deltas on tropical coasts. Temperate rainforests filled with evergreen and laurel trees are lower and less dense than other kinds of rainforests because the climate is more equable, with a moderate temperature range and well-distributed annual rainfall.
The topography of rainforests varies considerably, from flat lowland plains marked by small rock hills to highland valleys crisscrossed by streams. Volcanoes that produce rich soils are fairly common in the humid tropical forests.
Soil conditions vary with location and climate, although most rainforest soils tend to be permanently moist and soggy. The presence of iron gives the soils a reddish or yellowish colour and develops them into two types of soils: extremely porous tropical red loams, which can be easily tilled; and lateritic soils, which occur in well-marked layers that are rich in different minerals. Chemical weathering of rock and soil in the equatorial forests is intense, and in rainforests weathering produces soil mantles up to 100 metres (330 feet) deep. Although these soils are rich in aluminum, iron oxides, hydroxides, and kaolinite, other minerals are washed out of the soil by leaching and erosion. The soils are not very fertile, either, because the hot, humid weather causes organic matter to decompose rapidly and to be quickly absorbed by tree roots and fungi.
Rainforests exhibit a highly vertical stratification in plant and animal development. The highest plant layer, or tree canopy, extends to heights between 30 and 50 metres (98 to 164 feet). Most of the trees are dicotyledons, with thick leathery leaves and shallow root systems. The nutritive, food-gathering roots are usually no more than a few centimetres deep. Rain falling on the forests drips down from the leaves and trickles down tree trunks to the ground, although a great deal of water is lost to leaf transpiration.
Most of the herbaceous food for animals is found among the leaves and branches of the canopy, where a variety of animals have developed swinging, climbing, gliding, and leaping movements to seek food and escape predators. Monkeys, flying squirrels, and sharp-clawed woodpeckers are some of the animals that inhabit the treetops. They rarely need to come down to ground level.
The next lowest layer of the rainforest is filled with small trees, lianas, and epiphytes, such as orchids, bromeliads, and ferns. Some of these are parasitic, strangling their host’s trunks; others use the trees simply for support.
Above the ground surface the space is occupied by tree branches, twigs, and foliage. Many species of animals run, flutter, hop, and climb in the undergrowth. Most of these animals live on insects and fruit, although a few are carnivorous. They tend to communicate more by sound than by sight in this dense forest strata.
Contrary to popular belief, the rainforest floor is not impassable. The ground surface is bare, except for a thin layer of humus and fallen leaves. The animals inhabiting this strata, such as rhinoceroses, chimpanzees, gorillas, elephants, deer, leopards, and bears, are adapted to walking and climbing short distances. Below the soil surface, burrowing animals, such as armadillos and caecilians, are found, as are microorganisms that help decompose and free much of the organic litter accumulated by other plants and animals from all strata.
The climate of the ground layer is unusually stable. The upper stories of tree canopies and the lower branches filter sunlight and heat radiation, as well as reduce wind speeds, so that the temperatures remain fairly even throughout the day and night.
Virtually every group of animals except fishes is represented in the rainforest ecosystem. Many invertebrates are very large, such as giant snails and butterflies. The breeding seasons for most animals tend to be coordinated with the availability of food, which, although generally abundant, does vary seasonally from region to region. Climatic variations, however, are slight and thus affect animal behaviour very little. Those animals that do not have highly developed modes of quick locomotion are concealed from predators by camouflage or become nocturnal feeders.
Although similar to temperate rainforests, tropical rainforests are found in wet tropical uplands and lowlands around the Equator.
Rainforests are vegetation types dominated by broad-leaved trees that form a dense upper canopy (layer of foliage) and contain a diverse array of vegetation. Contrary to common thinking, not all rainforests occur in places with high, constant rainfall; for example, in the so-called “dry rainforests” of northeastern Australia the climate is punctuated by a dry season, which reduces the annual precipitation. Nor are all forests in areas that receive large amounts of rainfall true rainforests; the conifer-dominated forests in the extremely wet coastal areas of the American Pacific Northwest are temperate evergreen forest ecosystems. Therefore, to avoid conveying misleading climatic information, the term rainforest is now preferred over rain forest.
This section covers only the richest of rainforests—the tropical rainforests of the ever-wet tropics.
Tropical rainforests represent the oldest major vegetation type still present on the terrestrial Earth. Like all vegetation, however, that of the rainforest continues to evolve and change, so that modern tropical rainforests are not identical with rainforests of the geologic past.

Rainforest vegetation along the northern coast of Ecuador. © Victor Englebert
Tropical rainforests grow mainly in three regions: the Malesian botanical subkingdom, which extends from Myanmar (Burma) to Fiji and includes the whole of Thailand, Malaysia, Indonesia, the Philippines, Papua New Guinea, the Solomon Islands, and Vanuatu and parts of Indochina and tropical Australia; tropical South and Central America, especially the Amazon basin; and West and Central Africa. Smaller areas of tropical rainforest occur elsewhere in the tropics wherever climate is suitable. The principal areas of tropical deciduous forest are in India, the Myanmar–Vietnam–southern coastal China region, and eastern Brazil, with smaller areas in South and Central America north of the Equator, the West Indies, southeastern Africa, and northern Australia.
The flowering plants (angiosperms) first evolved and diversified during the Cretaceous Period about 100 million years ago, during which time global climatic conditions were warmer and wetter than those of the present. The vegetation types that evolved were the first tropical rainforests, which blanketed most of the Earth’s land surfaces at that time. Only later—during the middle of the Paleogene Period, about 40 million years ago—did cooler, drier climates develop, leading to the development across large areas of other vegetation types.
It is no surprise, therefore, to find the greatest diversity of flowering plants today in the tropical rainforests where they first evolved. Of particular interest is the fact that the majority of flowering plants displaying the most primitive characteristics are found in rainforests (especially tropical rainforests) in parts of the Southern Hemisphere, particularly South America, northern Australia and adjacent regions of Southeast Asia, and some larger South Pacific islands. Of the 13 angiosperm families generally recognized as the most primitive, all but two—Magnoliaceae and Winteraceae—are overwhelmingly tropical in their present distribution. Three families—Illiciaceae, Magnoliaceae, and Schisandraceae—are found predominantly in Northern Hemisphere rainforests. Five families—Amborellaceae, Austrobaileyaceae, Degeneriaceae, Eupomatiaceae, and Himantandraceae—are restricted to rainforests in the tropical Australasian region. Members of the Winteraceae are shared between this latter region and South America; those of the Lactoridaceae grow only on the southeast Pacific islands of Juan Fernández; members of the Canellaceae are shared between South America and Africa; and two families—Annonaceae and Myristicaceae—generally occur in tropical regions. This has led some authorities to suggest that the original cradle of angiosperm evolution might lie in Gondwanaland, a supercontinent of the Southern Hemisphere thought to have existed in the Mesozoic Era (251 to 65.5 million years ago) that consisted of Africa, South America, Australia, peninsular India, and Antarctica. An alternative explanation for this geographic pattern is that in the Southern Hemisphere, especially on islands, there are more refugia—that is, isolated areas whose climates remained unaltered while those of the surrounding areas changed, enabling archaic life-forms to persist.
The first angiosperms are thought to have been massive, woody plants appropriate for a rainforest habitat. Most of the smaller, more delicate plants that are so widespread in the world today evolved later, ultimately from tropical rainforest ancestors. While it is possible that even earlier forms existed that await discovery, the oldest angiosperm fossils—leaves, wood, fruits, and flowers derived from trees—support the view that the earliest angiosperms were rainforest trees. Further evidence comes from the growth forms of the most primitive surviving angiosperms: all 13 of the most primitive angiosperm families consist of woody plants, most of which are large trees.
As the world climate cooled in the middle of the Cenozoic, it also became drier. This is because cooler temperatures led to a reduction in the rate of evaporation of water from, in particular, the surface of the oceans, which led in turn to less cloud formation and less precipitation. The entire hydrologic cycle slowed, and tropical rainforests—which depend on both warmth and consistently high rainfall—became increasingly restricted to equatorial latitudes. Within those regions rainforests were limited further to coastal and hilly areas where abundant rain still fell at all seasons. In the middle latitudes of both hemispheres, belts of atmospheric high pressure developed. Within these belts, especially in continental interiors, deserts formed. In regions lying between the wet tropics and the deserts, climatic zones developed in which rainfall adequate for luxuriant plant growth was experienced for only a part of the year. In these areas new plant forms evolved from tropical rainforest ancestors to cope with seasonally dry weather, forming tropical deciduous forests. In the drier and more fire-prone places, savannas and tropical grasslands developed.
Retreat of the rainforests was particularly rapid during the period beginning 5 million years ago leading up to and including the Pleistocene Ice Ages, or glacial intervals, that occurred between 2.6 million and 11,700 years ago. Climates fluctuated throughout this time, forcing vegetation in all parts of the world to repeatedly migrate, by seed dispersal, to reach areas of suitable climate. Not all plants were able to do this equally well because some had less-effective means of seed dispersal than others. Many extinctions resulted. During the most extreme periods (the glacial maxima, when climates were at their coldest and, in most places, also driest), the range of tropical rainforests shrank to its smallest extent, becoming restricted to relatively small refugia. Alternating intervals of climatic amelioration led to repeated range expansion, most recently from the close of the last glacial period about 10,000 years ago. Today large areas of tropical rainforest, such as Amazonia, have developed as a result of this relatively recent expansion. Within them it is possible to recognize “hot spots” of plant and animal diversity that have been interpreted as glacial refugia.
Tropical rainforests today represent a treasure trove of biological heritage. They not only retain many primitive plant and animal species but also are communities that exhibit unparalleled biodiversity and a great variety of ecological interactions. The tropical rainforest of Africa was the habitat in which the ancestors of humans evolved, and it is where the nearest surviving human relatives—chimpanzees and gorillas—live still. Tropical rainforests supplied a rich variety of food and other resources to indigenous peoples, who, for the most part, exploited this bounty without degrading the vegetation or reducing its range to any significant degree. However, in some regions a long history of forest burning by the inhabitants is thought to have caused extensive replacement of tropical rainforest and tropical deciduous forest with savanna.
Not until the past century, however, has widespread destruction of tropical forests occurred. Regrettably, tropical rainforests and tropical deciduous forests are now being destroyed at a rapid rate in order to provide resources such as timber and to create land that can be used for other purposes, such as cattle grazing. Today tropical forests, more than any other ecosystem, are experiencing habitat alteration and species extinction on a greater scale and at a more rapid pace than at any time in their history—at least since the major extinction event at the end of the Cretaceous Period, some 65.5 million years ago.
The equatorial latitude of tropical rainforests and tropical deciduous forests keeps day length and mean temperature fairly constant throughout the year. The sun rises daily to a near-vertical position at noon, ensuring a high level of incoming radiant energy at all seasons. Although there is no cold season during which plants experience unfavourable temperatures that prohibit growth, there are many local variations in climate that result from topography, and these variations influence and restrict rainforest distribution within the tropics.
Tropical rainforests occur in regions of the tropics where temperatures are always high and where rainfall exceeds about 1,800 to 2,500 mm (about 70 to 100 inches) annually and occurs fairly evenly throughout the year. Similar hot climates in which annual rainfall lies between about 800 and 1,800 mm and in which a pronounced season of low rainfall occurs typically support tropical deciduous forests—that is, rainforests in which up to about three-quarters of the trees lose their leaves in the dry season. The principal determining climatic factor for the distribution of rainforests in lowland regions of the tropics, therefore, is rainfall, both the total amount and the seasonal variation. Soil, human disturbance, and other factors also can be important controlling influences.
The climate is always hot and wet in most parts of the equatorial belt, but in regions to its north and south seasonal rainfall is experienced. During the summer months of the Northern Hemisphere—June to August—weather systems shift northward, bringing rain to regions in the northern parts of the tropics, as do the monsoon rains of India and Myanmar. Conversely, during the Southern Hemisphere’s summer, weather systems move southward, bringing rain from December to February to places such as northern Australia. In these hot, seasonally wet areas grow tropical deciduous forests, such as the teak forests of Myanmar and Thailand. In other locations where conditions are similar but rainfall is not so reliable or burning has been a factor, savannas are found.
Topographic factors influence rainfall and consequently affect rainforest distribution within a region. For example, coastal regions where prevailing winds blow onshore are likely to have a wetter climate than coasts that experience primarily offshore winds. The west coasts of tropical Australia and South America south of the Equator experience offshore winds, and these dry regions can support rainforests only in very small areas. This contrasts with the more extensively rainforest-clad, east-facing coasts of these same continents at the same latitudes. The same phenomenon is apparent on a smaller scale where the orientation of coastlines is parallel to, rather than perpendicular to, wind direction. For example, in the Townsville area of northeastern Australia and in Benin in West Africa, gaps in otherwise fairly continuous tracts of tropical rainforest occur where the prevailing winds blow along the coast rather than across it.
Mean temperatures in tropical rainforest regions are between 20 and 29 °C (68 and 84 °F), and in no month is the mean temperature below 18 °C (64 °F). Temperatures become critical with increasing altitude; in the wet tropics temperatures fall by about 0.5 °C (0.9 °F) for every 100 metres (328 feet) climbed. Vegetation change across altitudinal gradients tends to be gradual and variable and is interpreted variously by different authorities. For example, in Uganda tropical rainforest grows to an altitude of 1,100 to 1,300 metres (3,600 to 4,300 feet) and has been described as giving way, via a transition forest zone, to montane rainforest above 1,650 to 1,750 metres (5,400 to 5,700 feet), which continues to 2,300 to 3,400 metres (7,500 to 11,200 feet). In New Guinea, lowland tropical rainforest reaches 1,000 to 1,200 metres (3,300 to 3,900 feet), above which montane rainforests extend, with altitudinal variation, to 3,900 metres (12,800 feet). In Peru, lowland rainforest extends upward to 1,200 to 1,500 metres (3,900 to 4,900 feet), with transitional forest giving way to montane rainforest above 1,800 to 2,000 metres (5,900 to 6,600 feet), which continues to 3,400 to 4,000 metres (11,200 to 13,100 feet). These limits are comparable and reflect the similarities of climate in all regions where tropical rainforests occur. Plant species, however, are often quite different among regions.
Although the climate supporting tropical rainforests is perpetually hot, temperatures never reach the high values regularly recorded in drier places to the north and south of the equatorial belt. This is partly due to high levels of cloud cover, which limit the mean number of sunshine hours per day to between four and six. In hilly areas where air masses rise and cool because of the topography, the hours of sunlight may be even fewer. Nevertheless, the heat may seem extreme owing to the high levels of atmospheric humidity, which usually exceed 50 percent by day and approach 100 percent at night. Exacerbating the discomfort is the fact that winds are usually light; mean wind speeds are generally less than 10 km (6.2 miles) per hour and less than 5 km (3.1 miles) per hour in many areas. Devastating hurricanes (cyclones and typhoons) occur periodically in some coastal regions toward the margins of the equatorial belt, such as in the West Indies and in parts of the western Pacific region. Although relatively infrequent, such storms have an important effect on forest structure and regeneration.
The climate within any vegetation (microclimate) is moderated by the presence of plant parts that reduce incoming solar radiation and circulation of air. This is particularly true in tropical rainforests, which are structurally more dense and complex than other vegetation. Within the forest, temperature range and wind speed are reduced and humidity is increased relative to the climate above the tree canopy or in nearby clearings. The amount of rain reaching the ground is also reduced—by as much as 90 percent in some cases—as rainwater is absorbed by epiphytes (plants that grow on the surface of other plants but that derive nutrients and water from the air) and by tree bark or is caught by foliage and evaporates directly back to the atmosphere.
Soils in tropical rainforests are typically deep but not very fertile, partly because large proportions of some mineral nutrients are bound up at any one time within the vegetation itself rather than free in the soil. The moist, hot climatic conditions lead to deep weathering of rock and the development of deep, typically reddish soil profiles rich in insoluble sesquioxides of iron and aluminum, commonly referred to as tropical red earths. Because precipitation in tropical rainforest regions exceeds evapotranspiration at almost all times, a nearly permanent surplus of water exists in the soil and moves downward through the soil into streams and rivers in valley floors. Through this process nutrients are leached out of the soil, leaving it relatively infertile. Most roots, including those of trees, are concentrated in the uppermost soil layers where nutrients become available from the decomposition of fallen dead leaves and other organic litter. Sandy soils, particularly, become thoroughly leached of nutrients and support stunted rainforests of peculiar composition. A high proportion of plants in this environment have small leaves that contain high levels of toxic or unpalatable substances. A variant of the tropical rainforest, the mangrove forest, is found along estuaries and on sheltered sea coasts in tidally inundated, muddy soils.

Epiphytic orchids (Dendrobium). E.R. Degginger
Even within the same area, however, there are likely to be significant variations in soil related to topographic position and to bedrock differences, and these variations are reflected in forest composition and structure. For example, as altitude increases—even within the same area and on the same bedrock—soil depth decreases markedly and its organic content increases in association with changes in forest composition and structure.
Only a minority of plant and animal species in tropical rainforests and tropical deciduous forests have been described formally and named. Therefore, only a rough estimate can be given of the total number of species contained in these ecosystems, as well as the number that are becoming extinct as a result of forest clearance. Nevertheless, it is quite clear that these vegetation types are the most diverse of all, containing more species than any other ecosystem. This is particularly so in regions in which tropical rainforests not only are widespread but also are separated into many small areas by geographic barriers, as in the island-studded Indonesian region. In this area different but related species often are found throughout various groups of islands, adding to the total regional diversity. Exceptionally large numbers of species also occur in areas of diverse habitat, such as in topographically or geologically complex regions and in places that are believed to have acted as refugia throughout the climatic fluctuations of the past few million years. According to some informed estimates, more than a hundred species of rainforest fauna and flora become extinct every week as a result of widespread clearing of forests by humans. Insects are believed to constitute the greatest percentage of disappearing species.
All major groups of terrestrial organisms are represented abundantly in tropical rainforests. Among the higher plants, angiosperms are particularly diverse and include many primitive forms and many families not found in the vegetation of other ecosystem types. Many flowering plants are large trees, of which there is an unparalleled diversity. For example, in one area of 23 hectares (57 acres) in Malaysia, 375 different tree species with trunk diameters greater than 91 cm (35.8 inches) have been recorded, and in a 50-hectare (124-acre) area in Panama, 7,614 trees belonging to 186 species had trunk diameters greater than 20 cm (7.8 inches). New species of plants—even those as conspicuously large as trees—are found every year. Relatively few gymnosperms (conifers and their relatives), however, are found in rainforests; instead, they occur more frequently at the drier and cooler extremes of the range of climates in which tropical rainforests grow. Some plant families, such as Arecaceae (palms), are typically abundant in all tropical rainforest regions, although different species occur from region to region. Other families are more restricted geographically. The family Dipterocarpaceae (dipterocarps) includes many massive trees that are among the most abundant and valuable species in the majority of tropical rainforests in western Malesia; the family, however, is uncommon in New Guinea and Africa and absent from South and Central America and Australia. The Bromeliaceae (bromeliads), a large family consisting mainly of rainforest epiphytes and to which the pineapple belongs, is entirely restricted to the New World.
Monsoon forests, which are also known as dry forests or tropical deciduous forests, are open woodland in tropical areas that have a long dry season followed by a season of heavy rainfall. The trees in a monsoon forest usually shed their leaves during the dry season and come into leaf at the start of the rainy season. Many lianas (woody vines) and herbaceous epiphytes (air plants, such as orchids) are present. Monsoon forests are especially well developed in Southeast Asia and are typified by tall teak trees and thickets of bamboo.

Monsoon forest in the Anaimalai Hills, Western Ghats, Tamil Nadu state, India. Tropical deciduous forests grow in seasonally dry areas of the tropics. These forests occur mainly in India, the Myanmar–Vietnam–southern coastal China region, eastern Brazil, smaller areas in South and Central America north of the Equator, the West Indies, southeastern Africa, and northern Australia. Gerald Cubitt
Tropical rainforests, which contain many different types of trees, seldom are dominated by a single species. A species can predominate, however, if particular soil conditions favour this occurrence or minimal disturbance occurs for several tree generations. Tropical deciduous forests are less diverse and often are dominated by only one or two tree species. The extensive deciduous forests of Myanmar, for example, cover wide areas and are dominated by only one or two tree species—teak (Tectona grandis) and the smaller leguminous tree Xylia xylocarpa. In Thailand and Indochina deciduous forests are dominated by members of the Dipterocarpaceae family, Dipterocarpus tuberculatus, Pentacme suavis, and Shorea obtusa.
Ferns, mosses, liverworts, lichens, and algae are also abundant and diverse, although not as well studied and cataloged as the higher plants. Many are epiphytic and are found attached to the stems and sometimes the leaves of larger plants, especially in the wettest and most humid places. Fungi and other saprophytic plants (vegetation growing on dead or decaying matter) are similarly diverse. Some perform a vital role in decomposing dead organic matter on the forest floor and thereby releasing mineral nutrients, which then become available to roots in the surface layers of the soil. Other fungi enter into symbiotic relationships with tree roots (mycorrhizae).
Interacting with and dependent upon this vast array of plants are similarly numerous animals. Like the plants, most animal species are limited to only one or a few types of tropical rainforest within an area, with the result that the overall number of species is substantially greater than it is in a single forest type. For example, a study of insects in the canopy of four different types of tropical rainforest in Brazil revealed 1,080 species of beetle, of which 83 percent were found in only one forest type, 14 percent in two, and only 3 percent in three or four types. While the larger, more conspicuous vertebrates (mammals, birds, and to a lesser degree amphibians and reptiles) are well known, only a small minority of the far more diverse invertebrates (particularly insects) have ever been collected, let alone described and named.
As with the plants, some animal groups occur in all tropical rainforest regions. A variety of fruit-eating parrots, pigeons, and seed-eating weevil beetles, for example, can be expected to occur in any tropical rainforest. Other groups are more restricted. Monkeys, while typical of tropical rainforests in both the New and the Old World, are entirely absent from New Guinea and areas to its east and south. Tree kangaroos inhabit tropical rainforest canopies only in Australia and New Guinea, and birds of paradise are restricted to the same areas.
To a large extent these geographic variations in tropical rainforest biota reflect the long-term geologic histories of these ancient ecosystems. This is most clearly demonstrated in the Malesian phytogeographic subkingdom, which has existed as a single entity only since continental movements brought Australia and New Guinea northward into juxtaposition with Southeast Asia about 15 million years ago. Before that time the two parts were separated by a wide expanse of ocean and experienced separate evolution of their biota. Only a relatively small sea gap lies between them today; Java, Bali, and Borneo are on one side, and Timor and New Guinea are on the other, with islands such as Celebes and the Moluccas forming an intermediate region between. The gap is marked by a change in flora and, especially, fauna and is known as Wallace’s Line. The contrast is particularly stark with respect to mammals. To the west the rainforests are populated—or were populated until recently—by monkeys, deer, pigs, cats, elephants, and rhinoceroses, while those to the east have marsupial mammals, including opossums, cuscuses, dasyurids, tree kangaroos, and bandicoots. Only a few groups such as bats and rodents have migrated across the line to become common in both areas. Similar contrasts, albeit less pronounced, can be seen in many other animal and plant groups across the same divide.
Tropical rainforests are distinguished not only by a remarkable richness of biota but also by the complexity of the interrelationships of all the plant and animal inhabitants that have been evolving together throughout many millions of years. As in all ecosystems, but particularly in the complex tropical rainforest community, the removal of one species threatens the survival of others with which it interacts. Some interactions are mentioned below, but many have yet to be revealed.
Plants with similar stature and life-form can be grouped into categories called synusiae, which make up distinct layers of vegetation. In tropical rainforests the synusiae are more numerous than in other ecosystem types. They include not only mechanically independent forms, whose stems are self-supporting, and saprophytic plants but also mechanically dependent synusiae such as climbers, stranglers, epiphytes, and parasitic plants. An unusual mix of trees of different sizes is found in the tropical rainforest, and those trees form several canopies below the uppermost layer, although they are not always recognizably separate layers. The upper canopy of the tropical rainforest is typically greater than 40 metres (131 feet) above ground.
The tropical rainforest is structurally very complex. Its varied vegetation illustrates the intense competition for light that goes on in this environment in which other climatic factors are not limiting at any time of year and the vegetation is thus allowed to achieve an unequaled luxuriance and biomass. The amount of sunlight filtering through the many layers of foliage in a tropical rainforest is small; only about 1 percent of the light received at the top of the canopy reaches the ground. Most plants depend on light for their energy requirements, converting it into chemical energy in the form of carbohydrates by the process of photosynthesis in their chlorophyll-containing green tissues. Few plants can persist in the gloomy environment at ground level, and the surface is marked by a layer of rapidly decomposing dead leaves rather than of small herbaceous plants. Mosses grow on tree butts, and there are a few forbs such as ferns and gingers, but generally the ground is bare of living plants, and even shrubs are rare. However, tree seedlings and saplings are abundant; their straight stems reach toward the light but receive too little energy to grow tall enough before food reserves from their seeds are exhausted. Their chance to grow into maturity comes only if overhanging vegetation is at least partially removed through tree death or damage by wind. Such an occurrence permits more solar radiation to reach their level and initiates rapid growth and competition between saplings as to which will become a part of the well-lit canopy.
As in most tropical forests, the trees of Panama exhibit a variety of different adaptations to aid dispersal of their seeds. These adaptations involve substantial investment of the trees’ material, but they are worthwhile because seed dispersal increases both the seeds’ and the species’ chances of survival. Seed destroyers such as herbivores, fungi, and bacteria often concentrate their activities in the vicinity of the parent tree. Therefore, seeds that can come to rest some distance away from the parent tree are more likely to germinate and grow.
Dispersal efforts that take advantage of air currents can be elaborate. Because the rainforest canopy effectively blocks wind from reaching the environment below, aerial seed dispersal is not as widely afforded as in other, more open ecosystems. Even so, many trees have managed to exploit this strategy. For example, the kapok tree, found in tropical forests throughout the world, is an emergent—a tree whose crown rises well above the canopy. The kapok’s towering height enables it to gain access to winds above the canopy. The tiny seeds of the kapok are attached to fine fibres that, when caught by the wind, enable distribution far from the parent tree. The balsa tree also uses fibrous seeds to distribute its progeny, but it is not an emergent. Instead, balsa grows quickly as a colonizer of gaps in the forest, giving its seeds access to wind while the gap in the trees is still open.

Woolly seeds produced by the seed pods of the kapok tree (Ceiba pentandra). Norman Myers—Bruce Coleman Inc.
Other trees grow aerodynamic structures to make use of the wind. The canopy trees Platypodium elegans and Tachigalia versicolor produce single-winged fruits similar to those of maple trees common in temperate zones. In the case of P. elegans, each fruit is attached to a twig by the tip of its wing and has a dry weight of about 2 grams (0.07 ounce)—only about 20 percent of which is the seed’s weight. They remain unripe for many months, but when Panama’s dry season arrives (January through March) the fruits dry out and are dispersed by strong seasonal winds. Seeds often are blown 50 metres (160 feet) or more. Shaded seedlings within about 30 metres (100 feet) of the parent tree tend to die from fungal attack, but fruits landing farther than 30 metres from the tree or in canopy gaps fare much better. The suicide tree encloses its seeds in elliptical wings that can measure nearly 15 cm (6 inches) long. The tree’s name comes from the fact that, after producing seeds, the tree dies.
Gaps in the canopy of a tropical rainforest provide temporarily well-illuminated places at ground level and are vital to the regeneration of most of the forest’s constituent plants. Few plants in the forest can successfully regenerate in the deep shade of an unbroken canopy; many tree species are represented there only as a population of slender, slow-growing seedlings or saplings that have no chance of growing to the well-lit canopy unless a gap forms. Other species are present, invisibly, as dormant seeds in the soil. When a gap is created, seedlings and saplings accelerate their growth in the increased light and are joined by new seedlings sprouting from seeds stored in the soil that have been stimulated to germinate by light or by temperature fluctuations resulting from the sun’s shining directly on the soil surface. Other seeds arrive by various seed-dispersal processes. A thicket of regrowth rapidly develops, with the fastest-growing shrubs and trees quickly shading out opportunistic, light-demanding, low-growing herbaceous plants and becoming festooned with lianas. Through it all slower-growing, more shade-tolerant but longer-lived trees eventually emerge and restore the full forest canopy. The trees that initially fill in the gap in the canopy live approximately one century, whereas the slower-growing trees that ultimately replace them may live for 200 to 500 years or, in extreme cases, even longer. Detailed mapping of the trees in a tropical rainforest can reveal the locations of previous gaps through identification of clumps of the quicker-growing, more light-demanding species, which have yet to be replaced by trees in the final stage of successional recovery. Local, natural disturbances of this sort are vital to the maintenance of the full biotic diversity of the tropical rainforest.
Just as tropical rainforest plants compete intensely for light above ground, below ground they vie for mineral nutrients. The process of decomposition of dead materials is of crucial importance to the continued health of the forest because plants depend on rapid recycling of mineral nutrients. Bacteria and fungi are primarily responsible for this process. Some saprophytic flowering plants that occur in tropical rainforests rely on decomposing material for their energy requirements and in the process use and later release minerals. Some animals are important in the decomposition process; for example, in Malaysia termites have been shown to be responsible for the decomposition of as much as 16 percent of all litter, particularly wood. Most trees in the tropical rainforest form symbiotic mycorrhizal associations with fungi that grow in intimate contact with their roots; the fungi obtain energy from the tree and in turn provide the tree with phosphorus and other nutrients, which they absorb from the soil very efficiently. A mat of plant roots explores the humus beneath the rapidly decomposing surface layer of dead leaves and twigs, and even rotting logs are invaded by roots from below. Because nutrients are typically scarce at depth but, along with moisture, are readily available in surface layers, few roots penetrate very deeply into the soil. This shallow rooting pattern increases the likelihood of tree falls during storms, despite the support that many trees receive from flangelike plank buttresses growing radially outward from their trunk bases. When large trees fall, they may take with them other trees against which they collapse or to which they are tied by a web of lianas and thereby create gaps in the canopy.

An earthstar (Geastrum) puffball, growing on moist soil among mosses. Larry West—The National Audubon Society/Photo Researchers
Tree growth requires substantial energy investment in trunk development, which some plants avoid by depending on the stems of other plants for support. Perhaps the most obvious adaptation of this sort is seen in plants that climb from the ground to the uppermost canopy along other plants by using devices that resemble grapnel-like hooks. Lianas are climbers that are abundant and diverse in tropical rainforests; they are massive woody plants whose mature stems often loop through hundreds of metres of forest, sending shoots into new tree crowns as successive supporting trees die and decay. Climbing palms or rattans (Calamus) are prominent lianas in Asian rainforests, where the stems, which are used to make cane furniture, provide a valuable economic resource.
Lianas in a tropical rainforest. The vascular tissues of lianas are modified primarily for water conduction, which leaves these tall plants dependent on other plants for support. © Gary Braasch
Epiphytes are particularly diverse and include large plants such as orchids, aroids, bromeliads, and ferns in addition to smaller plants such as algae, mosses, and lichens. In tropical rainforests epiphytes are often so abundant that their weight fells trees. Epiphytes that grow near the upper canopy of the forest have access to bright sunlight but must survive without root contact with the soil. They depend on rain washing over them to provide water and mineral nutrients. During periods of drought, epiphytes undergo stress as water stored within their tissues becomes depleted. The diversity of epiphytes in tropical deciduous forests is much less than that of tropical rainforests because of the annual dry season.
Parasitic flowering plants also occur. Hemiparasitic mistletoes attached to tree branches extract water and minerals from their hosts but carry out their own photosynthesis. Plants that are completely parasitic also are found in tropical rainforests. Rafflesia, in Southeast Asia, parasitizes the roots of certain lianas and produces no aboveground parts until it flowers; its large orange and yellow blooms, nearly 1 metre (3.28 feet) in diameter, are the largest flowers of any plant.
Stranglers make up a type of synusia virtually restricted to tropical rainforests. In this group are strangler figs (Ficus), which begin life as epiphytes, growing from seeds left on high tree branches by birds or fruit bats. As they grow, they develop long roots that descend along the trunk of the host tree, eventually reaching the ground and entering the soil. Several roots usually do this, and they become grafted together as they crisscross each other to form a lattice, ultimately creating a nearly complete sheath around the trunk. The host tree’s canopy becomes shaded by the thick fig foliage, its trunk constricted by the surrounding root sheath and its own root system forced to compete with that of the strangling fig. The host tree is also much older than the strangler and eventually dies and rots away, leaving a giant fig “tree” whose apparent “trunk” is actually a cylinder of roots, full of large hollows that provide shelter and breeding sites for bats, birds, and other animals. Stranglers may also develop roots from their branches, which, when they touch the ground, grow into the soil, thicken, and become additional “trunks.” In this way stranglers grow outward to become large patches of fig forest that consist of a single plant with many interconnected trunks.
Some of the tallest trees and lianas, and the epiphytes they support, bear flowers and fruits at the top of the rainforest canopy, where the air moves unfettered by vegetation. They are able to depend on the wind for dispersal of pollen from flower to flower, as well as for the spreading of fruits and seeds away from the immediate environment of the parent plant. Ferns, mosses, and other lower plants also exploit the wind to carry their minute spores. However, a great many flowering plants, including many that grow in the nearly windless environment of the understory, depend on animals to perform these functions. They are as dependent on animals for reproductive success as the animals are on them for food—one example of the mutual dependence between plants and animals.
Many rainforest trees have sizable seeds from which large seedlings emerge and thrust their way through the thick mat of dead leaves on the dark forest floor. They develop tall stems, using food reserves in the seed without having to rely on sunlight, which is usually too dim, to meet their energy requirements. Because large seeds cannot be dispersed by the wind, these plants depend on a variety of animals to perform this function and have evolved many adaptations to encourage them to do so. Fruit bats are attracted by fragrant, sweet fruits typically borne conspicuously and conveniently on the outer parts of the tree canopy; the mango (Mangifera indica), native to the rainforests of India, provides a good example. The bats not only feed on fruits as they hang from the trees but also may carry a fruit away to another perch, where they eat the flesh and drop the seed. Smaller fruits may be swallowed whole, the seeds passing through the gut intact and being voided at a distance. The ground beneath trees used by fruit bats as a roost is commonly thick with seedlings of fleshy, fruit-bearing trees.
A variety of birds eat fleshy fruits also, voiding or regurgitating the unharmed seeds. Birds of different sizes are typically attracted to similarly scaled fruits, which are carried on stems of appropriate thickness and strength. For example, large pigeons in New Guinea feed preferentially on larger fruits borne on thicker stems that can bear not only the weight of the fruit but also the weight of the large bird; smaller pigeons tend to feed on smaller fruits borne on thinner twigs. In such a manner, the diverse plant community is matched by a similarly diverse animal community in interdependence.
Terrestrial mammals also help to disperse seeds. In many cases this has favoured the positioning of flowers and fruits beneath the canopy on the trunks of trees accessible to animals unable to climb or fly, an adaptation called cauliflory. In some cases fruits are grown in the canopy but drop as they ripen, opening only after they fall to attract ground-dwelling animals that will carry them away from the parent tree. The durian fruit (Durio zibethinus) of Southeast Asian rainforests is an example; its fruits are eaten and its seeds dispersed by a range of mammals, including pigs, elephants, and even tigers.

Mango (Mangifera indica). Robert C. Hermes from the National Audubon Society Collection/Photo Researchers—EB Inc.
Numerous plants depend on animal dispersers to transport seeds either internally or externally. Birds generally disperse seeds internally by eating the fruits, which are often small and red and the numerous seeds of which easily pass through the birds’ digestive systems. Some seeds actually have higher rates of germination after passing through animal gut; others benefit from being deposited in nutrient-rich dung. Fruit bats such as the Jamaican, or common, fruit bat (Artibeus jamaicensis) are important seed dispersers in Panama, feeding on many fruits, including those of figs (genus Ficus) and cecropias (genus Cecropia), and distributing some seeds internally and others externally. The bat homes in on the smell of ripe fruit and transports it to a feeding roost away from the source tree. Small seeds are eaten and later excreted in flight, whereas larger seeds are discarded at the feeding site.

Barbados cherry (Malpighia glabra). Douglas David Dawn
Other examples of external seed transport by animals are also common. Some trees provide rich fruit that is attractive to foraging animals. As a consequence, organisms ranging from ants to bats to rodents such as the agouti unwittingly disperse the trees’ seeds. For example, the wild cashew (Anacardium excelsum) bears nuts on a sweet, green stem enlargement (hypocarp) that is a favourite food of many bats, which disperse the nuts while feeding.
The seed dispersal process can be complex, involving the activity of more than one animal, or it may depend on specific animal behaviours. The bright orange fruits of the black palm (Astrocaryum standleyanum), for example, comprise a seed covered by a tough woody layer forming a nut, or stone, which is in turn covered by a layer of pulp. When the fruit ripens and drops to the forest floor, many animals come to eat the sweet pulp, sometimes moving the seeds about in the process. Since weevils lay eggs on nearly all black palm fruits, unless agoutis peel the orange flesh from the palm nuts and bury them, the newly hatched weevil larvae destroy the seeds. Therefore, despite the fact that they eat large numbers of the seeds themselves, agoutis provide a net benefit to the palm. In the absence of agoutis it is likely that a tract of forest with Astrocaryum would offer few prospects for new trees.
Agoutis are also important to the almendro tree (Dipteryx panamensis), which attracts many dispersers because it fruits at the end of Panama’s dry season, when fruit is in short supply. A single seed is encased in a thick, hard wooden pod covered with a thin layer of green pulp. When a fruit crop ripens, numerous arboreal animals flock to it, including kinkajous, bats, monkeys, coatis, and squirrels. In addition, ground dwellers such as agoutis, peccaries, pacas, spiny rats, and tapirs seek out fruits that fall to the forest floor. Most of these animals simply eat the sweet pulp covering the fruit, but for the almendro seed to germinate it must first be carried far from its parent tree and buried. In the case of the almendro, the process is initiated by 70-gram (2.5-ounce) fruit bats (Artibeus lituratus), which first disperse a large number of fruits by carrying them off to feeding roosts away from the parent tree, where they chew off the pulp and drop the seeds. Then agoutis, which are less likely to bury almendro seeds found near parent trees, carry off seeds that the bats have dropped and bury some of them. Normally, agoutis consume most of these seeds or eat the seedlings when they germinate, but in a year of abundant fruit buried seeds will often germinate and grow. Thus, the almendro may need two animals, the fruit bat and the agouti, to give its seeds the opportunity to become new trees. Such findings strongly suggest that, in order to conserve many of the tree species in a tropical forest, it is also important to protect animal populations.

Cashew apples (hypocarp) and nuts of the domesticated cashew tree (Anacardium occidentale). W.H. Hodge
Many other animals, from ants to apes, are involved in seed dispersal. In the Amazon basin of Brazil, where large areas of tropical rainforest are seasonally flooded, many trees produce fruit attractive to fish, which swallow them whole and void the seeds. Squirrels are also important seed dispersers in parts of South America. In the tropical rainforests of northeastern Australia, cassowaries are responsible for generating mixed clumps of tree seedlings of several species that grow from their dung sites.
It is important for seeds to be spread away from parent plants, both to allow seedlings to escape competition with the parent and to expand the range of the species. Another capacity important to seed survival, particularly in the diverse tropical rainforest community, involves the evasion of seed predators. Many different beetles and other insects are specialized to feed on particular types of seed. Seeds concentrated beneath a parent plant are easy for seed predators to locate. Seeds that are carried away to areas occupied by different plant species—and different seed predators—are more likely to survive.
In addition to dispersing seeds, animals are vital to tropical rainforest reproduction through flower pollination. Many insects such as bees, moths, flies, and beetles as well as birds and bats carry out this activity. Birds such as the hummingbirds of South and Central America and the flower-peckers of Asia have adaptations that allow them to sip nectar from flowers. In the process they inadvertently become dusted with pollen, which they subsequently transport to other flowers, pollinating them. The plants involved also show special adaptations in flower structure and colour. Most flowers pollinated by birds are red, a colour highly visible to these animals, whereas flowers pollinated by night-flying moths are white or pink, and those pollinated by insects that fly during the day are often yellow or orange. Bats are important pollinators of certain pale, fragrant flowers that open in the evening in Asian rainforests.
When two or more species in an ecosystem interact to each other’s benefit, the relationship is said to be mutualistic. The production of Brazil nuts and the regeneration of the trees that produce them provide an example of mutualism, and in this case the interaction also illustrates the importance of plant and animal ecology in maintaining a rainforest ecosystem.
Euglossine bees (most often the females) are the only creatures regularly able to gain entrance to the Brazil nut tree’s flowers, which have lids on them. The bees enter to feed on nectar, and in the process they pollinate the flower. Pollination is necessary to initiate the production of nuts by the tree. Thus, the Brazil nut tree depends on female euglossine bees for pollination.
Male euglossines have a different role in this ecological process. To reproduce, the males must first prove themselves to the females. The males accomplish this by visiting orchids for the single purpose of gathering fragrant chemicals from the flowers. These fragrances are a necessary precondition of euglossine mating. Without the orchids of the surrounding rainforest, the euglossine population cannot sustain itself, and the Brazil nut trees do not get pollinated. For this reason, Brazil nuts used for human consumption must be collected from the rainforest; they cannot be produced on plantations.
Once the Brazil nut pods are formed, the tree then depends on the agouti, a rodent, to distribute and actually plant the seeds. The agouti is one of the few animals capable of chewing through the very hard pod to reach the nuts inside. Agoutis scatter and bury the nuts for future consumption, but some nuts manage to sprout and grow into mature trees.
Of all vegetation types, tropical rainforests grow in climatic conditions that are least limiting to plant growth. It is to be expected that the growth and productivity (total amount of organic matter produced per unit area per unit time) of tropical rainforests would be higher than that of other vegetation, provided that other factors such as soil fertility or consumption by herbivorous animals are not extremely low or high.
Various methods are employed to assess productivity. Gross primary productivity is the amount of carbon fixed during photosynthesis by all producers in the ecosystem. However, a large part of the harnessed energy is used up by the metabolic processes of the producers (respiration). The amount of fixed carbon not used by plants is called net primary productivity, and it is this remainder that is available to various consumers in the ecosystem—such as the herbivores, decomposers, and carnivores. Of course, in any stable ecosystem there is neither an accumulation nor a diminution in the total amount of organic matter present, so that overall there is a balance between the gross primary productivity and the total consumption. The amount of organic matter in the system at any point in time, the total mass of all the organisms present, is called the biomass.
The biomass of tropical rainforests is larger than that of other vegetation. It is not an easy quantity to measure, involving the destructive sampling of all the plants in an area (including their underground parts), with estimates made of the mass of other organisms belonging to the ecosystem such as animals. Measurements show that tropical rainforests typically have biomass values on the order of 400 to 700 metric tons per hectare, greater than most temperate forests and substantially more than other vegetation with fewer or no trees. A measurement of biomass in a tropical deciduous forest in Thailand yielded a value of about 340 metric tons per hectare.
Increase in biomass over the period of a year at one rainforest site in Malaysia was estimated at 7 metric tons per hectare, while total litter fall was 14 metric tons, estimated mass of sloughed roots was 4 metric tons, and total live plant matter eaten by herbivorous animals (both invertebrate and vertebrate) was about 5 metric tons per hectare per year. These values add up to a total net production of 30 metric tons per hectare per year. Respiration by the vegetation itself was estimated at 50 metric tons, so that gross primary productivity was about 80 metric tons per hectare per year. Compared with temperate forests, these values are approximately 2.5 times higher for net productivity and 4 times higher for gross productivity, the difference being that the respiration rate at the tropical site was 5 times that of temperate forests.
Despite the overall high rates of productivity and biomass in tropical rainforests, the growth rates of their timber trees are not unusually fast; in fact, some temperate trees and many smaller herbaceous plants grow more rapidly. The high productivity of tropical rainforests instead results in their high biomass and year-round growth. They also have particularly high levels of consumption by herbivores, litter production, and especially plant respiration.
As recently as the 19th century tropical forests covered approximately 20 percent of the dry land area on Earth. By the end of the 20th century this figure had dropped to less than 7 percent. The factors contributing to deforestation are numerous, complex, and often international in scope. Mechanization in the form of chain saws, bulldozers, transportation, and wood processing has enabled far larger areas to be deforested than was previously possible. Burning is also a significant and dramatic method of deforestation. At the same time, more damage is being done to the land that is the foundation of tropical forest ecosystems: heavy equipment compacts the soil, making regrowth difficult; dams flood untouched tracts of wilderness to produce power; and mills use wood pulp and chips of many tree species, rather than a select few, to produce paper and other wood products consumed primarily by the world’s industrialized nations. Although political, scientific, and management efforts are under way to determine means of slowing the destruction of tropical forests, the world’s remaining acreage continues to shrink rapidly as demand for wood and land continues to rise.
The implications of forest loss extend far beyond the borders of the states in which the forests grow. The role that rainforests play at the global level in weather, climatic change, oxygen production, and carbon cycling, while significant, is only just beginning to be appreciated. For instance, tropical rainforests play an important role in the exchange of gases between the biosphere and atmosphere. Significant amounts of nitrous oxide, carbon monoxide, and methane are released into the atmosphere from these forests. This metabolism is being changed by human activity. More than half the carbon monoxide derived from tropical forests comes from their clearing and burning, which are reducing the size of such forests around the world.
Another consequence of deforestation must be examined. In the upper Amazon River basin of South America, the rainforest recycles rains brought primarily by easterly trade winds. Indeed, surface transpiration and evaporation supply about half the rainfall for the entire region, and in basins of dense forest far from the ocean such local processes can account for most of the local rainfall. Should the Amazon Rainforest, which accounts for 30 percent of the land area in the equatorial belt, disappear, drought would likely follow, and the global energy balance might well be affected.
The primary forces causing tropical deforestation and forest degradation can be tied to economic growth and globalization and to population growth. Population growth drives deforestation in several ways, but subsistence agriculture is the most direct in that the people clearing the land are the same people who make use of it. Rural populations must produce what food they can from the land around them, and in the rainforest this is most often accomplished via slash-and-burn agriculture. Forest is cleared, the cuttings are burned, and crops are planted for local consumption. However, the infertile tropical soils are productive for only a few years, and so it is soon necessary to repeat the process elsewhere. This form of shifting agriculture has been practiced sustainably among aboriginal cultures worldwide for centuries. Small patches of forest are cleared and abandoned when they become unproductive. The community then settles another isolated part of the forest, thus allowing previously settled land to regenerate.
However, in areas throughout the tropics larger populations than before now live at the forest margins. As subsistence agriculture progresses onto adjacent land, there is no opportunity for regeneration, especially if the shifting population is increasing. In some regions lowland forests have already been exhausted, and upland forests have been cleared. Land located on the slopes of hills and mountains is particularly susceptible to erosion and, therefore, to loss of the topsoil needed to sustain vegetation—arboreal or agricultural. Lowland tropical forests are not immune to erosion, however, as the heavy rainfall washes away unprotected soils.
Another subsistence-related factor in deforestation is demand for fuelwood, which is the main source of energy for 40 percent of the world’s population. As population increases, this demand exerts significant and growing pressure on tropical forests, particularly in Africa.
Urban population growth has led to the establishment of resettlement programs in several countries. Governments have made land available to poor families in overcrowded cities, who then have attempted to begin new lives from cleared forest. In Brazil the Transamazonian highway system was begun in the 1960s to enable development and settlement of the Amazon Rainforest. Part of the Transamazonian highway, called BR 364, penetrates the remote state of Rondônia in west-central Brazil. Since the highway’s construction, this region has undergone significant deforestation. Main roads are cut into the forest, and parallel sets of access roads allow access to individual plots of land that are settled by farmers. This method of settlement results in a characteristic “fishbone” pattern when the land is viewed from above.
Small farms line the slopes in the highlands of Burundi, one of the most densely populated regions in Central Africa. Dr. Nigel Smith/The Hutchison Library
Brazil’s resettlement program, while extensive, is by no means the largest. Population resettlement to provide agricultural employment and access to land is also important in some Southeast Asian countries, notably Indonesia, Malaysia, and Vietnam. By far the largest program has been conducted in Indonesia, where more than four million people have been voluntarily resettled from Java and Bali to the less-populated islands, especially to the province of Irian Jaya on the island of New Guinea. Despite considerable success, the program has been plagued by such problems as improper site selection, environmental deterioration, migrant adjustment, land conflicts, and inadequate financing. A program in Malaysia has been quite successful, in part because it set much smaller settlement targets and was better funded. Vietnamese development policy also utilized the resettlement of people in an effort to revitalize areas outside the major population centres.
While resettlement in Malaysia or Indonesia entails sea travel to isolated islands, roads connect South American population centres to the Amazon, where frontier cities draw both unsuccessful farmers from rural areas and migrants from established cities. The Amazon basin has long been relatively uninhabited, but improved diets and sanitation and the greater ease of transportation are making it more attractive for human settlement. From the mid-1940s onward, a number of “penetration roads” have been built from the populous highlands of Colombia, Ecuador, Peru, and Bolivia into Amazonia, often in conjunction with Brazil’s Transamazonian highway. These roads have funneled untold numbers of landless peasants into the lowlands. Its vast area notwithstanding, the Amazon basin by the late 20th century had a predominantly urban population. Almost one-third of the estimated nine million Brazilians living in the 4.9 million-square-km (1.9 million-square-mile) area officially designated as Legal Amazonia were concentrated in Belém and Manaus, each with more than one million inhabitants, and in Santarém. These cities, which are logistic bases of operations for cattle ranching, mining, timber, and agroforestry projects, are still growing rapidly, with modern residential towers and shantytowns standing side by side. Even frontier trading centres in the interior, such as Marabá, Pôrto Velho, and Rio Branco, have 100,000 or more inhabitants. In the upper reaches of the drainage area, places such as Florencia in Colombia, Iquitos and Pucallpa in Peru, and Santa Cruz in Bolivia have become significant urban centres.
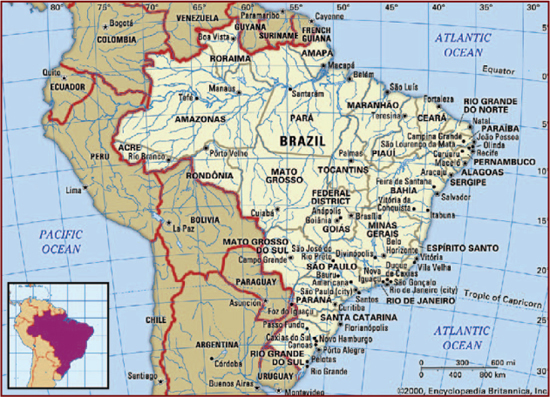
This map shows Brazil and its extensive river systems. Encyclopædia Britannica, Inc.
Most of those who come to the Amazon in resettlement programs are ill-prepared to become frontier farmers in an environment so naturally unsuitable to field agriculture, and the plots are soon abandoned. But the forest does not often reclaim the land; it is usually taken over by cattle ranchers first. In the Amazon and Central America the single largest use of cleared land is beef production—most of it for export. Cattle ranching thus illustrates how economic growth and globalization drive deforestation; other examples include logging and mining.
Tropical forests throughout the world often grow atop rich mineral deposits that are most easily mined by first clearing away the forest. The minerals are then extracted and sold in the global marketplace by the governmental or corporate enterprises involved. Even small tropical islands such as Fiji and New Caledonia have not been immune to deforestation by mining. In addition to clearing forests to gain access to deposits, mining also adds to deforestation by taking wood from the surrounding forest for ore processing. Such is the case in the Carajás region of Brazil, where tropical forest trees fuel iron smelters.
Gold deposits have been found in Indonesia and Papua New Guinea, as well as in the tropical forests north and south of the Amazon River. The resulting Amazon “gold rush” has brought as many as a half million transient miners (garimpeireos) equipped with picks, shovels, and sluice boxes to search for the mineral in alluvial deposits. Brazil’s annual production peaked in 1987 at nearly 90 tons, declining thereafter. Meanwhile, the mercury used in extracting the gold polluted waterways, causing the fish that are so important in the local diet to become inedible. On the Madeira River teams operating from rafts pump auriferous sediments from the riverbed; the sediments are subjected to a similar treatment.
Ostensibly, countries possessing tropical forests seek sources of trade, such as mining and logging, and income to raise their populations’ standard of living. It is often argued, however, that the underlying cause of economic dilemmas facing these governments is that control of resources is too concentrated among a wealthy few. Furthermore, these decision makers are not always from the developing countries, as multinational corporations can wield substantial influence on developing or unstable economies.
A common denominator in the destruction of tropical forests worldwide has been the pursuit of short-term gains at the expense of long-term prospects, both economic and environmental. By the end of the 20th century the importance of tropical forests had been realized, and conservation had become a subject of international politics. The institutional arrangements controlling tropical forests began to change significantly as the roles of environmental and other nongovernmental organizations (NGOs) at local, national, and international levels expanded. Recent changes have resulted in some measure of progress: development projects have been halted; sustainable management programs have become a focus of research; developing countries have established governmental departments to oversee the use of natural resources; and a broader range of interest groups, such as indigenous tribal peoples, are being considered. Protected areas are being set aside throughout the world as cooperation between institutions at the international level is realized. In 1997, for example, Brazil established 57,000 square km (22,000 square miles) of land as protected rainforest in the state of Amazonas, creating the world’s largest rainforest reserve.

Large iron mine in the Serra dos Carajás, Pará state, Brazil. © Tony Morrison/South American Pictures
The recent emergence of the ecotourism industry is a phenomenon that relies on the cooperation of various groups with interests in tropical forests. Ecotourism is recreational travel for the purposes of observing and experiencing natural environments. Rainforests are popular destinations, and these sites are often jointly operated by a combination of governmental, private, environmental, and indigenous groups. Ecotourism facilities also serve as biological research stations, and vice versa. In this way ecotourism can be seen as contributing to conservation efforts.
Such changes, while encouraging, are only beginning to work against the continuing decrease in acreage. International agreements among governments and businesses are highly dependent on the cooperation and commitment of the parties involved. Enforcement of policies at all levels of government, both within and between countries, is problematic. The record extent of fires in Amazonia and Indonesia in 1997–98 underscored profound problems in spite of recent progress. The relationships between oftentimes competing groups—local, national, and international; economic and environmental; governmental and nongovernmental—are what will determine the future of the planet’s tropical forests.
A temperate forest is a vegetation type with a more or less continuous canopy of broad-leaved trees. Such forests occur between approximately 25° and 50° latitude in both hemispheres. Toward the polar regions they grade into boreal forests, which are dominated by evergreen conifers, so that mixed forests containing both deciduous and coniferous trees occupy intermediate areas. Temperate forests usually are classified into two main groups: deciduous and evergreen.
Deciduous forests are found in regions of the Northern Hemisphere that have moist, warm summers and frosty winters—primarily eastern North America, eastern Asia, and western Europe. In contrast, evergreen forests—excepting boreal forests, which are covered in boreal forest—typically grow in areas with mild, nearly frost-free winters. They fall into two subcategories: broad-leaved forests and sclerophyllous forests. (Sclerophyllous vegetation has small, hard, thick leaves.) The former grow in regions that have reliably high, year-round rainfall; the latter occur in areas with lower, more erratic rainfall. Broad-leaved forests dominate the natural vegetation of New Zealand; they are significantly represented in South America, eastern Australia, southern China, Korea, and Japan; and they occur in less well-developed form in small areas of southeastern North America and southern Africa. Sclerophyllous forests occur particularly in Australia and in the Mediterranean region.
Temperate forests originated during the period of cooling of world climate that began at the start of the Cenozoic Era (65.5 million years ago). As global climates cooled, climatic gradients steepened with increasing latitude, and areas with a hot, wet climate became restricted to equatorial regions. At temperate latitudes, climates became progressively cooler, drier, and more seasonal. Many plant lineages that were unable to adapt to new conditions became extinct, but others evolved in response to the climatic changes, eventually dominating the new temperate forests. In areas that differed least from the previously tropical environments—where temperate evergreen forests now grow—the greatest numbers of plant and animal species survived in forms most similar to those of their tropical ancestors. Where conditions remained relatively moist but temperatures dropped in winter, deciduous trees evolved from evergreen rainforest ancestors. In areas that became much more dry—though not to the extent that tree development was inhibited and only scrubland or desert environments were favoured—sclerophyllous trees evolved.
During the rapid climatic fluctuations of the past two million years in which conditions alternated between dry, cold glacial states—the ice ages of some northern temperate regions—and warmer, moister interglacial intervals, tree species of temperate forests had to migrate repeatedly to remain within climates suitable for their survival. Such migration was carried out by seed dispersal, and trees that were able to disperse their seeds the farthest had an advantage. In the North American and European regions where ice-sheet development during glacial intervals was most extensive, the distances that had to be traversed were greatest, and many species simply died out. Extinctions occurred not only where migration distances were great but also where mountains or seas provided barriers to dispersal, as in southern Europe. Thus, many trees that were formerly part of the European temperate forests have become extinct in the floristically impoverished forest regions of western Europe and are restricted to small refuge areas such as the Balkans and the Caucasus. For example, buckeye (Aesculus) and sweet gum (Liquidambar) are two trees that no longer occur naturally in most parts of Europe, having disappeared during the climatic turmoil of the past two million years.
Human activities have had pronounced effects on the nature and extent of modern temperate forests. As long ago as 8,000 years, most sclerophyllous forests of the Mediterranean region had been cut over for timber or cleared to make space for agricultural pursuits. By 4,000 years ago in China the same process led to the removal of most broad-leaved and deciduous forests. In Europe of 500 years ago the original deciduous forests had disappeared, although they are remembered in nursery tales and other folklore as the deep, wild woods in which children and princesses became lost and in which dwarfs and wild animals lived.
The deciduous forests of North America had been cleared almost completely by the end of the 19th century. Australia and New Zealand experienced similar deforestation about the same time, although the earlier activities of pre-European peoples had substantial impacts. The character of the Australian sclerophyllous forests changed in response to more than 38,000 years of burning by the Aboriginal people, and the range of these forests was expanded at the expense of broad-leaved forests. In New Zealand about half the forested area, which previously had covered almost the entire country, was destroyed by fire brought to the island by the Polynesian inhabitants who arrived 1,000 years before the Europeans.
Winter in the temperate latitudes can present extremely stressful conditions that greatly affect the vegetation. The days are shorter and temperatures are low, so much so that in many places leaves are unable to function for long periods and are susceptible to damage from freezing. These conditions reduce the photosynthetic activity of the trees. In regions where winter temperatures regularly fall well below the freezing point and where soil moisture and nutrients are not in short supply, many trees have evolved a type of leaf that is relatively delicate and thin with a life span of a single growing season. Because such deciduous leaves do not require a large input of chemical energy, it is not too wasteful for the plant to shed them after a single growing season.
The “throwaway” leaves of temperate deciduous trees are shed as the days shorten in autumn and are replaced by new leaves in the spring. The forests dominated by these trees, therefore, have extremely pronounced seasonal changes in appearance, function, and climate. Most trees in temperate deciduous forests follow this habit, although some evergreen species usually are scattered among them. In particular, several broad-leaved evergreen shrubs are found in the understory of temperate deciduous forests that have less delicate, longer-lasting leaves than their deciduous neighbours; these leaves have adaptations that allow them to survive the freezing winter temperatures, and they can carry out photosynthesis for more than a single summer.
In areas where milder conditions prevail, however, photosynthesis may be possible at any season without need for protective mechanisms against frost damage. In these relatively unstressful circumstances most trees may benefit from retaining their leaves throughout the year, and heavy use of resources through frequent leaf replacement is thereby avoided. In environments such as these that also have a sufficient supply of moisture, temperate broad-leaved forests are found.
In mild but drier temperate environments, moisture shortage necessitates that trees develop thickened leaves. These leaves often have a reduced surface area or they dangle pendulously from limbs, two strategies employed to slow the loss of water (transpiration). High levels of energy and nutrients are needed to produce these thick leaves, which, therefore, cannot be replaced easily at annual intervals, an added reason for sclerophyllous trees to retain their leaves throughout the year. With foliage perennially present, these trees can carry out photosynthesis whenever moisture becomes available, provided temperatures are warm enough; this characteristic is advantageous where rainfall is infrequent and unpredictable.
Soils in temperate sclerophyllous forests are frequently poor in mineral nutrients. This poverty of nutrients accounts in part for the nature of the vegetation, because the annual production of leaves or the development of a dense broad-leaved canopy requires a significant input of nutrients. In contrast, soils in regions of deciduous and broad-leaved evergreen forest are generally fertile. (The forests that occupied the best soils in most regions, however, have been cleared almost completely to make way for agriculture.) Typical temperate deciduous forest soils are mull soils, which have a high level of organic matter especially close to the surface that is well mixed with mineral matter. Variations in soil materials and fertility have a strong influence on the types of trees that will dominate the forest. For example, in northwestern Europe, the European beech (Fagus sylvatica) dominates deciduous forests on shallow soils that overlie chalk, while oak (Quercus) is dominant on deeper, clay soils. The richness of the ground flora beneath the trees generally increases with soil fertility.
Intimate associations, or mycorrhizae, between tree roots and fungi are important and occur in most tree species. Although important in all forest types, these interactions have been studied more thoroughly in temperate deciduous forests. The fungal component of this symbiotic partnership grows on or in the fine roots of trees and benefits by obtaining nutrition in the form of carbohydrates from the tree root; the tree, in turn, is better nourished because a mycorrhizal root is more efficient at absorbing dissolved mineral nutrients from the soil than is an uninfected root.
Waterlogging of soils in temperate deciduous woodlands commonly occurs in regions with higher rainfall and humidity in late winter and spring, such as the British Isles. This occurs not only because in winter rainfall is higher and evaporation lower but also because the trees, barren of foliage, transpire a minimal amount of moisture. Areas subject to waterlogging include clay-rich soils, places that have low slope angles, depressions, and spots along watercourses. These places tend to have a richer ground flora but a less luxuriant tree canopy, which consists of only a few species that are tolerant of wet soils.
The depth of tree roots in temperate deciduous forests varies, but in many instances roots are shallow, with few reaching 1 metre (3.28 feet) below the surface. In the European beech, for example, shallow lateral growth of roots predominates over the development of a deep taproot, leading to growth of a “root plate” just beneath the soil surface. This enables the tree to exploit nutrients released at the surface by litter decomposition efficiently, while avoiding deeper layers that may become waterlogged. However, trees with root plates are more prone to being blown over in gales, especially after heavy rain has made the soil more plastic.

Bracket fungi, which grow on tree trunks, are among some of the largest fungi. Some species may reach 40 cm (16 inches) in diameter. H.S. Knighton
The principal regions of deciduous forest all occur in the Northern Hemisphere and have historical connections between them. Thus, many similarities exist among their biota. The same plant and animal genera tend to occur in all regions, although different species are found in each region. However, the European deciduous forest flora is poorer than that of eastern North America and East Asia. Many plants are common and widespread in the forests of North America and Asia, but in Europe they are present only as restricted relict populations or fossils. Examples include hickory (Carya), Magnolia, sour gum (Nyssa), and sweet gum.
Deciduous forests are composed primarily of broad-leaved trees that shed all their leaves during one season. Deciduous forest is found in three middle-latitude regions with a temperate climate characterized by a winter season and year-round precipitation: eastern North America, western Eurasia, and northeastern Asia. Deciduous forest also extends into more arid regions along stream banks and around bodies of water.
Oaks, beeches, birches, chestnuts, aspens, elms, maples, and basswoods (or lindens) are the dominant trees in mid-latitude deciduous forests. They vary in shape and height and form dense growths that admit relatively little light through the leafy canopy. Shrubs are found primarily near clearings and forest edges, where more light is available, and herbaceous flowering plants are abundant within the forest in the spring, before the trees come into full leaf.
The soils upon which deciduous forests thrive are gray-brown and brown podzols. They are slightly acidic and have a granular humus layer known as mull, which is a porous mixture of organic material and mineral soil. Mull humus harbours many bacteria and invertebrate animals such as earthworms.
Snails, slugs, insects, and spiders are common inhabitants of the deciduous forest, and many cold-blooded vertebrates, such as snakes, frogs, salamanders, and turtles, are also present. Birds are represented by warblers, flycatchers, vireos, thrushes, woodpeckers, hawks, and owls. Prominent mammals include mice, moles, chipmunks, rabbits, weasels, foxes, bears, and deer.
Differences in temperature, moisture, and elevation may cause the formation of distinct plant associations within the deciduous-forest pattern. The dominance of beeches and maples in the northern part of the eastern North American deciduous forest and that of oaks and hickories along the southern extension of this vegetation are typical examples.
Most of the areas of North American deciduous forest are dominated by oaks (several species of Quercus) and/or beech (Fagus grandifolia), with maples (Acer) and species of hickory and linden or basswood (Tilia) also widespread. Beech and basswood are rare in other North American vegetation types, but oaks, hickories, and maples are more widespread.
In addition to these widespread species, many other trees are important components of the North American deciduous forests on a local scale. The rich, mixed mesophytic (adapted to environments that are neither too dry nor too wet) forest type found north of the Appalachians includes buckeye and tulip tree (Liriodendron), while the southern floodplain forest of the Mississippi valley is made up of oaks mixed with sour gum and the evergreen conifer swamp cypress (Taxodium). Southward and eastward of the Appalachians, oaks and hickories mingle with another conifer, pine (Pinus), while east of the Great Lakes, beech mix with maples, birch (Betula), and hemlock (Tsuga). Elsewhere, ash (Fraxinus), hop hornbeam (Ostrya), poplar (Populus), elm (Ulmus), and, until its decimation by fungal infection, chestnut (Castanea), are also important. A wide range of understory shrubs and small trees includes dogwood (Cornus), holly (Ilex), Magnolia species, and serviceberry (Amelanchier).
Fragments of the North American deciduous forest also occur on mountains in Mexico and Guatemala, where many of the same trees—such as Fagus, Fraxinus, Juglans (walnut), Liquidambar, Quercus—occur as identical or closely related species. Commonly these trees are accompanied by an understory of evergreen shrubs of tropical affinity.
In Japan, Korea, and China, north of the evergreen broad-leaved forests there is a gradual transition to deciduous forests. In Japan deciduous forests are dominated by beech (Fagus crenata and F. japonica), oak (Quercus crispula), and maple (Acer carpinifolium and other species); other trees mingle with these, including cherries (several species of Prunus), ash, Magnolia, and, in the east, the evergreen conifer fir (Abies). The leaves of many deciduous trees in Japan, like those in North America but unlike most in Europe, turn to bright shades of red and yellow before they are shed in autumn, the maples being particularly spectacular. Below the trees a dense layer of dwarf bamboo (Sasa) commonly grows; it may be so thick that it prevents the canopy trees from regenerating from seedlings. Thus, rapid, dense regrowth by dwarf bamboo may seriously interfere with reforestation after logging. Many small flowering herbs such as Aconitum, Shortia, Mitchella, and Viola grow at ground level. Much of the area of Chinese deciduous forests is dominated by various oaks, frequently mixed with other trees including maples, alder (Alnus), ash, walnut, poplar, and many others. A varied understory includes many small trees or shrubs such as hornbeam (Carpinus), dogwood, service-tree (Sorbus), and the shrubs Acanthopanax and Aralia, which are relatives of ivy. Deciduous forests of birch fringe the oak forests at their northern and montane margins.
The dominant trees of the European deciduous forests are generally closely related to their equivalents in North America and Asia, consisting of different species of common genera. In well-drained areas, such as those on sloping ground or permeable soils, most deciduous forests are dominated by European beech or by one of a few species of oak. Beech is overwhelmingly dominant across large areas that have an oceanic climate; these regions are damper and milder than most other regions in which deciduous forests grow because of the influence of prevailing winds from the Atlantic Ocean. Eastward from Britain and western France through central Europe into Russia there is a progressive decrease in this oceanic influence. Along this gradient the forest flora changes. For example, beech, durmast oak (Quercus petraea), and European hornbeam (Carpinus betula) all reach their eastward limits in the area north of the Carpathian Mountains, while linden (Tilia cordata) is typical of a small number of deciduous forest trees that extend east beyond Moscow.
Places subject to seasonal waterlogging in European deciduous forests—and which are also nutrient rich—often are dominated by alder (Alnus glutinosa), growing above a rich ground flora including ferns (Athyrium and Dryopteris), sedges (Carex), and forbs (Caltha and Filipendula).
The only significant temperate deciduous forests in the Southern Hemisphere occur in a small area of Chile around Valdivia, between about 36° and 41° S. Forests here are dominated by a deciduous species of beech, Nothofagus obliqua, which usually grows amid evergreen trees more typical of the broad-leaved forests bordering this area to the south.
The milder environments that support temperate evergreen forests generally lie closer to the Equator than do areas with temperate deciduous forest. They have richer biotas than the sclerophyllous or deciduous forests that grow in more stressful environments at similar latitudes, although they are less rich than the tropical rainforests where environmental stress is at a minimum throughout the year.
In the southernmost regions of Japan and Korea, which enjoy a warm, wet climate, the natural vegetation is evergreen broad-leaved forest dominated by oaks and their near relative Castanopsiscuspidata and by the laurel Machilus thunbergii. Camphor laurel (Cinnamomum camphora), figs (Ficus retusa), pandans (Pandanus boninensis), palms (Livistona subglobosa), and other plants that require year-round warmth also occur in the warmest places, whose vegetation is described as subtropical by some authorities and as warm temperate forest by others. Shrubs in these forests include species of Aucuba, Camellia, and Eurya, and orchids and ferns are commonly found at ground level.
Similar forests in southern China, now almost completely replaced by farmland or tree plantations, had an even richer flora, with oaks, members of the laurel family, and the tea relative Schima being prominent examples of a large diversity of trees. The remnants of these Chinese evergreen broad-leaved forests extend west to the foothills of the Himalayas, where similar forests at lower altitudes include many trees such as Alcimandra, Castanopsis, Machilus, Magnolia, and Mallotus. At higher altitudes there are more sclerophyllous forests that contain fewer species and are dominated by oaks.
In Australia a variety of temperate broad-leaved forests occur, usually as small patches in moist, sheltered, and fire-protected areas in mountainous and coastal terrain along the east coast. They are described as temperate rainforests owing to similarities with the flora, structure, and ecology of tropical rainforests that are found in similar environments to the immediate north. The warm temperate rainforests of milder, more northern areas display a high diversity of trees, including coachwood (Ceratopetalum apetalum), crab apple (Schizomeria ovata), and yellow carabeen (Sloanea woollsii). Palms are often present, as are various climbing plants and epiphytes (plants that grow on other plants but that derive moisture and nutrients from rain), although not to the extent that they occur in tropical rainforests. Ferns are typically abundant, and many large, graceful tree ferns grow there.
A similar warm temperate rainforest grows in northern parts of New Zealand; it contains a mixed broad-leaved canopy of trees such as Elaeocarpus, Metrosideros, and Weinmannia, which is frequently penetrated and overtopped by tall conifers, including the massive kauri (Agathis australis). Palms (Rhopalostylis sapida) and various lianas are often present.
Cool temperate evergreen broad-leaved forests in the southernmost areas of eastern Australia, particularly Tasmania, and in New Zealand and the southern portion of South America are usually dominated by evergreen species of beech (Nothofagus), with different species occurring in each region. Few other trees typically coexist with Nothofagus in these cool forests, which also lack climbers and vascular epiphytes, although they may have a great abundance of mosses on tree trunks, branches, and sometimes leaves. In Australia other trees that may be present include sassafras (Atherosperma moschatum), Elaeocarpus holopetalus, and leatherwood (Eucryphia lucida), while in New Zealand conifers in the plum pine family (Podocarpaceae) commonly emerge above the broad-leaved canopy, especially on sites with a history of natural disturbance such as landslides. In Chile other trees growing with Nothofagus, or in some cases forming temperate evergreen forests without it, include Eucryphia, Chilean laurel (Laurelia), and Persea, with bamboos (Chusquea) becoming abundant at some sites after forest disturbance.
South of the European deciduous forests lie areas that were occupied by temperate sclerophyllous forests before the effects of human manipulation of the environment were felt. These areas extend as a narrow ring around the coastline of the Mediterranean Sea. Typical evergreen trees are oaks (several species including the cork oak, Quercus suber) and the pistachio relative Pistacia lentiscus, commonly mixed with various deciduous species near their northern limits and with pines elsewhere. Forests similar to the Mediterranean sclerophyllous forests extend east, discontinuously, as far as the western slopes of the Himalayas, although throughout their range they have been extensively altered by human activities and nowhere are considered to be in their natural state.
Temperate sclerophyllous forests are most widespread in Australia. It is remarkable that across the major portion of a very large area, the most common trees are various species of one genus, Eucalyptus. About 500 species are known of this archetypal Australian tree, which occurs naturally in few other regions. However, gum trees, as they are often called, are well known in many regions to which they have been introduced. Two other diverse and widespread trees in these forests are Acacia and Casuarina. Many other trees, shrubs, and grasses occupy subordinate positions. A sharp boundary separates the temperate sclerophyllous forests from the tropical and temperate rainforests of moister sites in the east. This abrupt transition is maintained by fire, which prevents or reverses invasion of the sclerophyllous forest by rainforest plants. However, where fires are infrequent, especially in southern regions, rainforest vegetation can form an understory beneath the Eucalyptus canopy. To the north, temperate sclerophyllous forests grade into similar tropical savanna and sclerophyllous forests; inland they merge into shrublands and deserts.
The fauna of temperate forests resembles the regional fauna. However, the structure of the vegetation provides similar ecological niches in all regions of the same vegetation type, so that, although different species inhabit different forests, they are of a similar type. Tree holes provide homes and nest sites for arboreal mammals and birds in most regions of temperate forest but with pronounced variations. For example, apart from bats no native mammals are found in the New Zealand forests. In Australia the arboreal mammals are all marsupials or bats, including gliders such as the greater glider (Petaurus volans) and opossums such as the common ringtail (Pseudocheirus peregrinus), which nests in holes, and the well-known koala (Phascolarctos cinerea), which is free-living and feeds mainly or entirely on young tree foliage.
In temperate forests of the Northern Hemisphere, squirrels are widespread. Local additional arboreal forms in Asian forests include monkeys, most of which are predominantly seedeaters. This feeding niche is particularly appropriate in Northern Hemisphere forests, which include more trees with large seeds, such as the acorn-producing oaks, than do their Southern Hemisphere equivalents.
Birds are less regionally distinct, with families such as those of the owl and pigeon being well represented in almost all temperate forest regions. Nevertheless, there are still some pronounced regional variations. The tits (Paridae) dominate the foliage-gleaning insectivore guild in Europe, where warblers (Sylviidae) are less varied; this situation is reversed in North America. More fundamental contrasts are apparent in Australia, where honeyeaters, which feed on nectar, and parrots, which feed on small, hard seeds, are diverse and common in the sclerophyllous forests. In the Northern Hemisphere few plants provide nectar for birds, and tree seeds are usually eaten by squirrels and pigeons.
At mid-latitudes the sun never rises to the near-vertical position in the sky as it does in the tropics. In winter, when the sun appears particularly close to the horizon throughout the short period of daylight, its direct radiant energy impinges most on slopes facing the Equator—southward in the Northern Hemisphere, northward in the Southern Hemisphere. In deciduous forest regions, primarily in the Northern Hemisphere where the trees are without leaves in winter, direct sunlight bathes the forest floor. The ground surface and the vegetation on southern slopes are quickly warmed, paradoxically creating a more stressful environment for plants than exists on the shadier, cooler northern slopes. The stress is due to more extreme temperature fluctuations than the southern slope sustains, which affects the speed at which the plant tissues, frozen during the cold night, are thawed. On the southern slope the thawing is much more rapid, which is more damaging than the gradual thaw that the vegetation on the northern slopes experiences. As a result, herbaceous plants of the forest floor that retain green leaves throughout winter, such as Hepatica, Hydrophyllum, and Tiarella in eastern North America, are more common on northern slopes, while sunnier slopes become bare and brown during the winter months.
As days lengthen and temperatures warm in spring, new, green photosynthetic shoots develop rapidly from buds that formed and food reserves that were laid down in storage tissues during the previous growing season. Annuals growing from seed are rare. The most rapid development of all typically occurs in the herbaceous plants of the forest floor. These plants must take immediate advantage of the spring warmth and sunlight before the new tree foliage casts its heavy shade over the ground, drastically reducing available light energy and slowing the rate of photosynthesis. Many plants of the forest floor have underground energy-storing organs such as bulbs, corms, or fleshy rhizomes that allow them to grow rapidly and strongly in spring to gain maximum advantage from the short, warm, shade-free period. They frequently also produce strong, pointed shoots that are able to emerge above the thick layer of dead leaves that dropped from the trees of the canopy the previous autumn. Thus, the ground of temperate deciduous forests in spring typically is covered by a green carpet of foliage that often includes dramatic displays of colourful flowers before the buds on the trees above have opened. Later, after the leaves of the tree canopy have regrown, the ground cover declines. Typical low-growing plants in North American deciduous forests include species of Cypripedium, Erythronium, Hydrophyllum, Trillium, and Viola. In Europe bluebells (Hyacinthoides non-scripta), daffodils (Narcissus pseudonarcissus), and wood anemones (Anemone nemorosa), among many other herbaceous species, provide similarly spectacular spring flower displays.
In autumn the delicate leaves of the deciduous trees senesce and start to die. As this happens, they lose their green colour and turn various shades of brown, yellow, and red. Dramatic displays are created, becoming significant tourist attractions in the areas in which the colours are brightest—that is, eastern North America and western Asia. A weak layer of tissue called the abscission layer develops at the base of each leaf stalk, and at this point the stalk breaks and the leaf is shed. The massive leaf drop that ensues during autumn has earned the season its alternate designation, fall.
As in other forests, the composition of the temperate deciduous forest is commonly determined by the influence of disturbances—natural as well as human—on tree regeneration. For instance, in eastern North America, the tulip tree (Liriodendron tulipifera) produces seeds that can remain dormant in the soil for up to seven years. When they germinate, subsequent establishment of the quick-growing saplings is most successful on bare mineral soil in full light. Stands of this species therefore tend to be of the same age and act as markers of the time at which a catastrophe destroyed the former tree cover, baring the ground. Beech that occur in the same region are shade tolerant and occupy places not subject to catastrophe, regenerating beneath undamaged tree canopy.
There are still a few temperate forests that have been disturbed little by human activities, and the interrelationships between the vegetation and the large animals are of interest. A rare, relatively intact area of deciduous forest that contains some evergreen conifers is found in Poland. The most common trees include linden (species of Tilia), oak (Quercus robur), hornbeam (Carpinus betulus), maple (Acer platanoides), and spruce (Picea abies). Until 1923 large areas were dominated by linden, but most tree regeneration at that time was by spruce, hornbeam, and maple. By 1973 these latter species had become common as canopy trees, and lindens began regenerating freely, as they had not at the earlier time. Such changes result in part from the varying regeneration requirements of the trees. Most successful regeneration takes place where gaps of light reach the forest floor. Competition between saplings of different species is intense and has different outcomes depending in part on soil and light intensities and to a large degree on the effects of mammals. Rooting by wild pigs, although it destroys many small plants, creates suitable conditions for seedling establishment. Saplings are subject to browsing by deer, which also feed on herbaceous plants that compete with tree seedlings. European bison similarly eat foliage and, in addition, can debark trees with their horns. The combined variable influences of these and other animals interact to produce a mosaic forest of different tree ages and composition, with longer-term changes resulting from population fluctuations due, in part, to human impact.
Most temperate forests, where they still survive at all, have been so exploited and disturbed by human influences that their natural condition is difficult to discern. However, some areas outside the old agricultural regions of Asia and Europe have sustained much less human disturbance, making it possible to define natural disturbances responsible for some variations. For example, temperate broad-leaved forests in Chile have been shown to vary in structure and composition according to their history of natural catastrophe in the form of earthquake-induced landslides; similar relationships between ecosystem alteration and natural disturbances have been demonstrated in New Zealand with volcanic eruption and in Tasmania with wildfire. In all these regions, sites undisturbed for many centuries have forests dominated by shady, highly competitive species of Nothofagus, often with few seedlings of any kind beneath the large, old trees. However, in the wake of natural catastrophe, other trees can invade the sites, and only gradually does Nothofagus reestablish itself and slowly resume dominance during subsequent tree generations. Therefore, in areas that have suffered many instances of disturbance, there exists today a variety of forest types. Comparable variations resulting from storm damage and wildfire occurrence have been recognized in North American deciduous forests.
The total aboveground biomass (dry weight of organic matter in an area) for temperate deciduous forests is typically 150 to 300 metric tons per hectare; values for temperate broad-leaved forests are generally higher, and those for sclerophyllous forests are lower. The subterranean component is more difficult to measure, but it appears to approximate a value of about 25 percent of the aboveground component in deciduous forests and rather more in temperate evergreen forests. Total biomass in temperate deciduous forests is, therefore, about 190 to 380 metric tons per hectare. These values refer to mature, undamaged forests; when disturbances also are considered, the range would include lower values.
For temperate forests gross primary productivity (the total biomass fixed by the vegetation in a unit area within a unit time) has been estimated at 16 to 50 metric tons per hectare per year. Net primary productivity, gross primary productivity less that used by plants in respiration, is approximately 10 metric tons per hectare per year; it is greatest in young forests where the trees are rapidly growing toward full size, and it declines in forests of old trees. While the gross primary productivity of temperate forests is considerably lower than that of tropical rainforests, the net primary productivity is not so different, reflecting the lower diversity and complexity of the consumer component of the temperate forest ecosystem.
Temperate forests have been useful to human populations in many diverse ways. Although in most places they have been replaced by simpler agricultural systems, large areas still remain, especially on poorer soils, and are important sources of timber. The total yield as well as the quality of timber is maximized by keeping forests in a condition of greatest net primary productivity—that is, by harvesting trees before they reach their age of declining growth. Old growth forests have a high biomass and considerable conservational significance but are not efficient in terms of total sustained timber yield.
Boreal forests, or taiga, are a vegetation type composed primarily of cone-bearing, needle-leaved, or scale-leaved evergreen trees, found in regions that have long winters and moderate to high annual precipitation.
The boreal (meaning northern) forest region occupies about 17 percent of the Earth’s land surface area in a circumpolar belt of the far Northern Hemisphere. Northward beyond this limit, the boreal forest merges into the circumpolar tundra. The boreal forest is characterized predominantly by a limited number of conifer species—that is, pine (Pinus), spruce (Picea), larch (Larix), fir (Abies)—and to a lesser degree by some deciduous genera such as birch (Betula) and poplar (Populus). These trees reach the highest latitudes of any trees on the Earth. Boreal plants and animals are adapted to short growing seasons of long days that vary from cool to warm. Winters are long and very cold, the days are short, and a persistent snowpack is the norm. The boreal forests of North America and Eurasia display a number of similarities, even sharing some plant and animal species. The northern forests of Russia, especially Siberia, are referred to as taiga, meaning “little sticks,” a term now widely accepted as an alternative to boreal forest.
During the final period of maximum cold temperatures (23,000 to 16,500 years ago), in the latter part of the Pleistocene Ice Age (which ended 11,700 years ago), species that now constitute the boreal forest were displaced as far south as 30° N latitude by the continental glaciers of Europe, Asia, and North America and by the hyperarid and extremely cold environments of unglaciated Asia and North America. As the glaciers began to retreat gradually about 18,000 years ago, species of the boreal forest began to move northward in Europe and North America. In eastern and central North America the northward movement of the forest was relatively steady and gradual. An exception to this progression occurred about 9,000 years ago in western Canada when white spruce spread rapidly northward across 2,000 km (1,240 miles) of newly deglaciated land in only 1,000 years. This rapid migration resulted from seed dispersal facilitated by strong northward winds caused by clockwise atmospheric circulation around the remnant ice cap of northern Quebec and the western part of Hudson Bay.
Because so much of the Earth’s water was bound up in ice at this time, sea levels were lower than they are today, and this allowed migrations of various terrestrial species to occur. Many areas that are now islands were then connected to the nearby mainland—for example, the British Isles were linked to Europe. As the climate warmed during the last stages of the glacial period, but before the sea level rose to its current position, some plants and animals of the mainland European boreal forest migrated to Britain. This biota exists today as part of the boreal forest in the highlands of Scotland. The areas of lowland central Alaska, the central Yukon territory, and far eastern Russia, which had climates too arid to permit the formation of ice sheets, were connected by the Bering Land Bridge, across which many species migrated. As a result, today across Alaska a gradient in plant characteristics can be observed, ranging from typical North American forms in the east to those with Eurasian characteristics in the west.
The boreal forests of North America and Eurasia are broad belts of vegetation that span their respective continents from Atlantic to Pacific coasts. In North America the boreal forest occupies much of Canada and Alaska. Although related transition forest types are present in the northern tier of the lower 48 United States, true boreal forest stops just north of the southern Canadian border. The vast taiga of Asia extends across Russia and southward into northeastern China and Mongolia. In Europe most of Finland, Sweden, and Norway are covered with boreal forest. A small, isolated area of boreal forest in the Scottish Highlands lacks some continental species but does contain the most widespread conifer of the Eurasian boreal forest, Scotch pine (Pinus sylvestris).
The position of the boreal forest zone generally is controlled by the degree of warmth experienced during the growing season, the temperature of the soil, and the extreme minimum winter temperature. The boreal forest belt consists of three roughly parallel zones: closed canopy forest, lichen woodland or sparse taiga, and forest-tundra. The closed canopy forest is the southernmost portion of the taiga. It contains the greatest richness of species, the warmest soils, the highest productivity, and the longest growing season within the boreal zone. North of the closed canopy forest is the lichen woodland—a smaller parallel zone of sparse forest or woodland in which tree crowns do not form a closed canopy. Lichen mats and tundralike vegetation make up a significant portion of the ground cover. To the north of the lichen woodland lies forest-tundra, which occurs along the northern edge of tree growth (tree line). Patches of trees consisting of only a few species dot restricted portions of the landscape, forming a complex mosaic with tundra. Many trees in the forest-tundra zone have never been known to produce viable seeds or have done so only sporadically. These trees were established during warmer climatic episodes from a few hundred to a few thousand years ago and have persisted since, usually by vegetative (asexual) reproduction. Forest fires in this zone remove trees, and because of the lack of reproduction, only unburned patches of trees remain.
The closed forest, or southern taiga zone, on both continents is not distributed along a strictly east-west axis. At the western margin of Europe the warming influence of the Gulf Stream allows the closed canopy forest to grow at its northernmost location, generally between about 60° and 70° N. In western North America the Kuroshio and North Pacific currents likewise warm the climate and cause the northward deflection of the forest into Alaska and Yukon in Canada. On the eastern margin of the continents the boreal forest is deflected southward to between about 50° and 60° N by the cold polar air masses that flow south along these coasts. This is the southernmost limit of the boreal forest, to the south of which, in humid eastern North America and Europe, lies a northern deciduous broad-leaved transition forest. In this forest small stands of boreal conifers are distributed on cooler or less productive sites such as peaty wetlands. In the arid centre of both continents the closed canopy boreal forest is bordered to the south by a forest parkland of trees and grassland.
The central portions of Eurasia and North America are regions of flat or gently rolling topography. There, the northern and southern boundaries of the boreal forest are broad and gradual; they have fluctuated by as much as 200 km (125 miles) during the past few thousand years. A well-defined but complex boundary is formed between taiga and alpine tundra on the mountains of the Pacific edge in western North America and the far eastern region of Russia. Generally the boreal forest does not come into contact with the humid temperate or subpolar rainforest of coastal Alaska and British Columbia because of high mountain barriers, but some low-elevation regions have a transition zone often characterized by trees that are a hybrid of Sitka spruce (Picea sitchensis) and white spruce (P. glauca). In Norway and Scotland a variant form of the boreal forest occupies extremely humid environments.
Practically all the large river systems of the taiga of Siberia, including the Ob, Yenisey, and Lena rivers, are northward flowing. The Ob in western Siberia forms a great lowland basin with a considerable percentage of the land surface covered with poorly drained peaty wetlands. In such situations within the boreal zone a closed canopy forest is generally absent.
Coldness is the dominant climatic factor in boreal forest regions, although a surprising diversity of climates exists. Several factors—namely, the solar elevation angle, day length, and snow cover—conspire to produce this cold climate. In the boreal region the Sun is never directly overhead (90°) as it can be in the tropics. The maximum solar angle decreases with increasing latitude. At latitude 50° N in the southern boreal region the maximum solar angle is 63.5° and at the Arctic Circle it is only 47°. As a result solar energy is less intense in the boreal regions because it is spread out over a greater area of the Earth’s surface than it is in equatorial regions. Day length also affects temperature. Long winter nights at high latitudes allow radiation emitted by the surface of the Earth to escape into the atmosphere, especially in continental interiors where cloud cover is less abundant than it is near the coast. Snow cover, too, affects the climate, because it reflects incoming solar radiation and amplifies cooling. During winter, a snowpack persists for at least five months in the southern portion of boreal forest regions and for seven or eight months in the northern reaches. The boreal forest actually mitigates this cooling because it roughens and darkens what would otherwise be a smooth, snow-covered, energy-reflecting surface for much of the year. It has been estimated that the Earth would be significantly colder without the boreal forest.
The northern limit of the North American boreal forest coincides with the mean position of the Arctic front—the boundary between Arctic and mid-continental air masses—in the summer; its southern limit coincides with the mean frontal position in the winter. Mean annual temperatures in the boreal forest range from a few degrees Celsius above freezing to −10 °C (14 °F) or more. Areas with a mean annual temperature below freezing are susceptible to the formation of permafrost soils (frozen ground).
The mean temperature of January, the coldest month, is generally less than −10 °C (14 °F) across the boreal region. The boreal forest includes areas that experience some of the lowest temperatures on the Earth, excluding Antarctica. At the height of winter an intensely cold pocket of air develops over inland areas of far eastern Siberia; mean temperatures of −50 °C (−58 °F) have been recorded in this region. As this Siberian cold air flows over the unfrozen northern Pacific Ocean a great temperature contrast develops that results in strong, westward-moving storm systems. The movement, position, and strength of these storms control much of the weather in the Northern Hemisphere.
Boreal forests do not grow on areas surrounding the Bering Strait. A rigorous cold climate with a very short snow-free season precludes the growth of trees on the Russian side of the Bering Strait in the Chukotka region of the Russian Far East. On the North American side in western Alaska summers are too cool for trees to grow because of cold air masses moving off the Bering Sea.
The growing season in the boreal forest is generally cool; the mean temperature of the warmest month, July, is generally between 15 and 20 °C (59 and 68 °F). Summer daytime high temperatures are typically cool to warm—20 to 25 °C (68 to 77 °F)—for much of the growing season in the boreal forest. Cool summer temperatures can actually produce higher photosynthetic efficiency in plants than can warmer conditions. Species adapted to cold respire less (use up less of their food stores) when photosynthesizing at cool temperatures in intense summer light than they do at higher temperatures, allowing a greater net gain in biomass (dry mass of organic matter).
Areas of the boreal forest located in the centre of continents generally receive 30 to 50 cm (12 to 20 inches) of annual precipitation. Precipitation totals are relatively modest in these locations because they are a significant distance from unfrozen oceans that supply moisture. Some boreal forests are semiarid and may even include grasslands interspersed with the forest. These forests are found in regions of Yukon and Alaska that occur on the leeward side of mountains, which are sheltered from moisture-bearing winds, as well as in some portions of the interior of far eastern Russia. Annual precipitation in low elevations of these regions is 30 cm (12 inches) or less. The highest annual precipitation total in the boreal forest, which can exceed 100 cm (39 inches), is in eastern North America and northern Europe. During ancient eras of colder climate these regions also received relatively abundant precipitation, which resulted in the buildup of glacial ice sheets. Today these once heavily glaciated regions support extensive lakes, streams, and wetlands.
Coniferous forests constitute a vegetation type composed primarily of cone-bearing, needle-leaved, or scale-leaved evergreen trees, found in regions of the world that have long winters and moderate to high annual precipitation. The northern Eurasian coniferous forest is called the taiga, or the boreal forest. Both terms are used to describe the entire circumpolar coniferous forest with its many lakes, bogs, and rivers. Coniferous forests also cover mountains in many parts of the world. Pines, spruces, firs, and larches are the dominant trees in coniferous forests. They are similar in shape and height and often form a nearly uniform stand with a layer of low shrubs or herbs beneath. Mosses, liverworts, and lichens cover the forest floor.
The light-coloured, usually acidic soils of coniferous forests are called podzols (podsols) and have a compacted humus layer, known as mor, which contains many fungi. These soils are low in mineral content, organic material, and number of invertebrate animals such as earthworms.
Mosquitoes, flies, and other insects are common inhabitants of the coniferous forest, but few cold-blooded vertebrates, such as snakes and frogs, are present because of the low temperatures. Birds include woodpeckers, crossbills, warblers, kinglets, nuthatches, waxwings, grouse, hawks, and owls. Prominent mammals include shrews, voles, squirrels, martens, moose, reindeer, and wolves.
Eurasian coniferous forest is dominated in the east by Siberian pine, Siberian fir, and Siberian and Dahurian larches. Scots pine and Norway spruce are the important species in western Europe.
North American coniferous forest is dominated throughout by white spruce, black spruce, and balsam fir, although lodgepole pine and alpine fir are important species in the western section.
A distinct subtype of the North American coniferous forest is the moist temperate coniferous forest, or coast forest, which is found along the west coast of North America eastward to the Rocky Mountains. This subtype is sometimes called temperate rain forest, although this term is properly applied only to broad-leaved evergreen forests of the Southern Hemisphere. Warm temperatures, high humidity, and often misty conditions encourage the development of a mossy, moisture-loving plant layer under the giant trees of Sitka spruce, western red cedar, western hemlock, Douglas fir, and coast redwood.
Other subtypes of coniferous forest occur at various elevations in the Rocky Mountains of North America, in Central America, and in eastern Asia. They are known as subalpine and montane forests and are dominated by combinations of pine, spruce, and fir species.
Extended periods of clear, dry weather in the boreal region are caused by persistent strong polar high pressure systems. If strong high pressure persists during the long days near the summer solstice, temperatures can warm to 30 °C (86 °F) or higher. Intense heating at the ground surface often produces convective storms with lightning but little rain, causing forest fires.
Boreal forest conifer litter is highly acidic. Soils of the more humid and southern boreal forest are highly leached spodosols, which are characterized by the leaching of iron, aluminum, and organic matter from the chemically and biologically distinct surface layer—horizon A—to the next layer—horizon B. Much of the soil of central and eastern Canada—granitic Canadian Shield—has been repeatedly scraped clean by glacial advances. Thus, productive forests often are restricted to portions of the landscape where soil material has been deposited by glaciers. Peaty wetlands occur where surface drainage is impeded by permafrost, youthful glacial topography, or aggraded rivers; their soils are characteristically organic soils, or histosols. Soils in much of boreal western North America and Asia are inceptisols, which have little horizon development. Very thin surface salt deposits are found in the most arid portions of the boreal forest.
Cold soils are characteristic of the boreal forest region, which overlaps the zone of permafrost. Permafrost is soil or earth material that remains below 0 °C (32 °F) for at least two years. The surface, or active, layer of permafrost thaws in the warm season and freezes in the winter, but the soil below the active layer remains continuously frozen. Because the plant rooting zone is restricted to the active layer, nutrient supply is limited and secure anchoring for roots is lacking. Some trees and other plants of the taiga (especially black spruce [Picea mariana] and tamarack [Larix laricina] in North America and larches in Siberia) can grow on permafrost if the active layer is sufficiently deep, but several species are eliminated from permafrost.
The boreal forest itself is an important contributing factor to the development of permafrost. The latter stages of forest growth—characterized by development of an intact forest canopy, growth of an insulating moss cover in summer, and accumulation of forest litter—may cool the soil to such an extent that permafrost develops. Warming of the soil is promoted by forest fires, which remove the canopy, moss, and forest litter layers. In the absence of an intact canopy, a deeper and more effective insulating layer of snow accumulates in the winter. The presence of dark ash following a fire increases solar energy absorption on the site for several years.
The boreal forest of Europe generally lacks permafrost, but east of the Ural Mountains and from central Canada northward permafrost is common. In southern and central boreal forests, permafrost occurs sporadically and occupies only a small percentage of the landscape that experiences the coldest temperatures. The northern portion of closed canopy forest and the lichen woodland zone are in a region of discontinuous permafrost, where permafrost is found on north-facing slopes and in cold air drainage basins but is absent from south-facing slopes and newly deposited alluvial sites. Most of the forest-tundra is within the continuous permafrost zone.
Forest productivity in the middle and northern taiga zones is directly related to soil temperature. Warmer soils decompose organic matter more quickly, releasing nutrients for new plant growth and creating a more productive site. Productive forest types occupy warmer, south-facing slopes and river terraces, and less productive dwarf or sparse forest occupies the north-facing and basin permafrost sites.
Floodplains throughout the boreal forest regions are free of permafrost, high in soil fertility, and repeatedly disturbed in ways that renew the early, rapid growth stages of forest succession. Floodplains are a mosaic of productive shrubland and forest that serve as a major habitat for moose (Alces alces), which influence ecosystem structure and function.
South-central Alaska and adjacent Yukon and British Columbia support the most extensive ice sheets and glaciers in the world outside the polar desert regions of Antarctica and Greenland. Glacial meltwater is a large part of the flow of larger rivers such as the Yukon and Tanana in Alaska and the Yukon territory. Glacial meltwater carries a heavy load of suspended sediment that deposits in riverbeds and causes frequent channel shifts. Glacial river floodplains are extensive, very dynamic, and constantly renewed with fertile soil material. In the ancient past exposed deposits of glacial silt were picked up by strong winds and deposited on surrounding hillsides. Fertile soils, known as loess, resulted, on which highly productive upland forests are found today. Because the beds of glacially fed rivers are rising, the landscape through which they flow is partially drowned from the impeded drainage, often preventing forest growth and favouring the development of marshes and mires.
Nearly all major taxonomic groups have fewer species of animals and plants in the boreal forest than they have in other terrestrial ecosystems at lower latitudes. This accords with the species diversity gradient that is observed from lower to higher latitudes, with numbers of species decreasing in a poleward direction.
Scotch pine is the most widely distributed pine species in the world, growing from northern Scotland to the Russian Pacific shore. The relatively humid and productive taiga of northern Europe and south-central Siberia is dominated by this species. Forest management has greatly favoured this species in Scandinavia and Finland. It is a thick-barked species and easily survives light ground fires, often reaching ages of 350 to 400 years, with some individuals being older than 700 years. European aspen and Siberian spruce are essentially transcontinental in distribution as well.
The species composition of Eurasian taiga is different east of central Siberia from that which prevails westward into Europe. Distinctive European species include Norway spruce (Picea abies), a large dominant species of the productive humid boreal forest, and Sukaczev larch (Larix sukaczewii), an early successional species (one of the first species to colonize an area after a disturbance) of European Russia. Gray (Betula populifolia) and white birch (B. pendula) occur across northern Europe and well into central Siberia. The birches often form dense stands of light- or white-barked trees that are considered a characteristic feature of the boreal forest. Siberian larch (Larix sibirica) and Siberian fir (Abies sibirica) are restricted to north-central Asia. Species restricted to northeastern Asia include chosenia (Chosenia arbutifolia), an early successional broad-leaved tree of floodplains; Siberian stone pine (Pinus sibirica), a short shrub or tree; and Asian spruce (Picea obovata).
Vegetation profile of a boreal forest. The tree layer consists mainly of conifers, and mosses are the predominant ground cover. Encyclopædia Britannica, Inc.
All North American tree species are distributed across the continent except jack pine (Pinus banksiana), lodgepole pine (Pinus contorta), and balsam fir (Abies balsamea). Jack pine is a relatively small, short-lived, early successional tree occurring in the eastern and central parts of boreal forests east of the Rocky Mountains. Lodgepole pine is a longer-lived, early successional species growing in western Canada and along the Rocky Mountain axis from central Yukon southward to well south of the boreal forest limit. Balsam fir is a shade-tolerant, late successional, but relatively short-lived tree that occurs only in the eastern and central boreal forest.
Major taiga tree species are well adapted to extreme winter cold. The northernmost trees in North America are white spruce that grow along the Mackenzie River delta in Canada, near the shore of the Arctic Ocean. The northernmost trees in the world are Gmelin larch (Larix gmelinii) found at latitude 72°40′N on the Taymyr Peninsula in the central Arctic region of Russia.
A distinctive feature of the flora of boreal forests is the abundance and diversity of mosses; about one-third of the ground cover under boreal forest is dominated by moss. Much of the ground cover in older conifer stands is moss, which grows on rocks, on tree trunks, and in the pits formed by upturned trees. Extensive peaty wetlands in the boreal region are often thick accumulations of dead sphagnum and other mosses, sedges, and other plants; a living moss layer continually grows at the surface.
Lichens (a symbiotic association of a fungus and algae) constitute a significant part of the ground cover in the lichen woodland or sparse taiga. Lichens are also generally well distributed on tree trunks and especially in the canopy of older conifers throughout the boreal forest. Because lichens and mosses are dispersed by airborne spores that can travel long distances, many species of both groups are found across the entire circumpolar boreal forest.
Many vascular plants are also widespread across the circumpolar north. Some forest understory species dominate their habitats; they include twinflower (Linnaea borealis), lingonberry (Vaccinium vitis-idaea), baneberry (Actaea rubra), and Swedish and Canadian dwarf cornel (Cornus suecica and C. canadensis). Several boreal forest plants are adapted to rapid colonization and growth in recently burned areas, such as fireweed (Epilobium angustifolium). The extensive peatlands of the boreal north support a typical flora that usually includes species such as Labrador tea (Ledum palustre), cloudberry (Rubus chamaemorus), cotton grass (Eriophorum species), and crowberry (Empetrum nigrum or E. hermaphroditum). In northern Europe crowberry also grows as shrub mats under Scotch pine forests or woodlands. Crowberry has been shown to produce secondary chemical compounds that inhibit or kill Scotch pine seedlings. Periodic light ground fires reduce the abundance and vigour of crowberry and allow tree regeneration.
Specialized orchids in the forest understory include calypso (Calypso bulbosa), coral root (Corallorrhiza trifida), and lady’s slipper (Cypripedum species). The roots of these plants form particular associations with fungi (mycorrhizae). Willow shrubs (Salix species) are one of the first plants to emerge following disturbances on floodplains and occasionally on uplands as well. Important grasses across the boreal region include species of bromegrass (Bromus species), bluegrass (Poa species), reed bent grass (Calamagrostis species), and vanilla grass (Hierochloe odorata). Many freshwater aquatic plants such as sedges (Carex species) and pondweeds (Potamogeton species) are distributed widely across the boreal zone of both continents because migratory waterfowl and shorebirds are effective in dispersing their seeds. Several species of ferns are common to the boreal forests of the two continents, especially in regions of higher precipitation.
Because a winter snowpack is a dependable feature of the taiga, several mammals display obvious adaptations to it. The snowshoe, or varying, hare (Lepus americanus), for example, undergoes an annual change in colour of its pelage, or fur, from brownish or grayish in the summer to pure white in the winter, providing effective camouflage. Its feet are large in proportion to its body size, a snowshoelike adaptation for weight distribution that allows the hare to travel over the surface of snow rather than sink down into it. The lynx (Lynx canadensis) is the principal predator of the snowshoe hare. It, too, has large feet, with fur between the toes, enabling the lynx to remain on the snow’s surface. Most animals of the boreal forest are well adapted to the cold and survive it easily if they have enough food to maintain an energy balance through the winter.
Moose are the largest browsing animals in the boreal forest. In the summer they eat willow and broad-leaved trees and also wade in lakes and ponds to consume aquatic plants. Throughout the winter moose eat large quantities of woody twigs and buds. Moose depend on high-quality feeding areas in the shrub zone along river floodplains and on the early successional growth of woody plants in burned or cutover forest. Intensive browsing by moose can alter the composition of the forest in its early stages of growth, often increasing the dominance of conifers, which they do not consume in as great amounts as they do broad-leaved trees. Harvesting a moose for winter food is an important and even critical element of survival for humans living in isolated rural areas of the taiga.
Moose populations are controlled by various means. Wolves (Canis lupus) prey on moose across most of the boreal forest, and some scientists and game managers believe that once moose numbers are depressed wolf predation can keep moose populations low. As a result, wolf trapping or shooting programs are carried out as a game management measure to increase prey numbers. The natural regulation of moose populations by wolf predation and the presence of wolves themselves is valued as well. As a result programs to control wolf populations are often the subject of intense debate. Other factors control moose numbers, such as the restriction of access to plants during years of deep snow and lack of early successional woody plant growth caused by forest maturation. Where the boreal forest is extensively cut for forest products, moose numbers have increased greatly, often to levels that are considered undesirable for forest regeneration. Subsistence and sport hunting of moose are widely used tools of moose population management.
Cyclical fluctuations in the population density of the snowshoe hare and its effect on the population of its predator, the lynx. The graph is based on data derived from the records of the Hudson’s Bay Company. Encyclopædia Britannica, Inc.
Another large-hoofed browsing mammal that is present seasonally in the boreal forest is the reindeer (Rangifer tarandus) in Eurasia and the closely related caribou in North America. A large portion of the reindeer population is semidomesticated and herded by nomadic peoples such as the Sami of Scandinavia and several native peoples in northern Russia. Caribou migrate the greatest distances of any large land mammal in North America. They often move in vast herds of 500,000 animals or more, seldom stopping or pausing because they must constantly forage in these environments of generally low productivity. During the early winter, reindeer and caribou migrate south from their summer ranges in the tundra to the forest-tundra or lichen woodland, where they graze primarily on lichens. Later in winter caribou typically move to open forests and sedge-rich lake margins or to snow-free wind-swept mountains. In April and May, caribou form long columns and migrate back north to the tundra.

American mink (Mustela vison). Karl H. Maslowski
Several mammals of the boreal region are valued for their furs, and trapping and trade in furs has been an important part of the culture, economy, and history of the region as long as humans have lived there. Important furbearing species include lynx and marten (Martes americana) and in wetland habitats beaver (Castor canadensis), mink (Mustela vison), and muskrat (Ondatra zibethica).
In the North American boreal forest the northern flying squirrel (Glaucomys sabrinus) is adapted to consume fungi, especially underground fruiting bodies (sporocarps) of fungi that form mutually beneficial relationships (mutualism) with trees by colonizing their roots. The flying squirrel’s consumption and dispersal of these underground fungi provide a significant benefit to the forest as a whole.
The boreal forest is the migratory destination of large numbers of birds for the summer breeding season. These include several passerine songbirds typical of shrub and forest habitats, such as thrushes, flycatchers, and warblers. Many of these species consume insects in the canopy of the boreal forest and other habitats. Predators of these birds occur in the forest as well, such as the sharp-shinned hawk (Accipiter striatus) and the northern goshawk (A. gentilis). Populations of several boreal forest-breeding migratory thrushes, flycatchers, and warblers may be declining because of the loss of their wintering habitats in the tropical forests of the world and the changes to or loss of forest habitats in the temperate zones along their migratory routes.
Birds of the boreal forest fill a variety of niches. Some are seed consumers or dispersers, others are insect consumers. They carry out other specialized roles as well. For example, the yellow-bellied sapsucker (Sphyrapicus varius) drills evenly spaced rows of small holes in the bark of trees and then visits these “wells” to obtain sap and the insects it attracts. Various other birds, mammals, and insects benefit from the sap wells, too.
Woodpeckers excavate tree cavities, which subsequently are used by many species of birds and mammals. Woodpeckers are specialized predators of wood- and bark-inhabiting insects; they are thought to be important in the control of the spruce beetle (Dendroctonus rufipennis) population. In searching for insects, woodpeckers chisel or strip the bark off dead or dying trees, promoting more rapid decay and the release of nutrients from dead trees. As large old trees have become rarer through forest cutting, some year-round resident woodpeckers such as the northern three-toed woodpecker (Picoides tridactylus) and the great spotted woodpecker (Dendrocopos major) have lost their habitats and declined in numbers.
Few bird species remain in boreal forests through the long cold winters because of limited opportunities for food, although some undertake only a short migration south. Resident bird species include the common raven (Corvus corax) and the boreal and black-capped chickadees of North America and the Siberian tit (Parus species).
The extensive areas of lakes, ponds, and wetlands—especially in the glaciated part of the boreal forest—provide a large habitat for waterfowl and shorebirds, although the birds primarily occur in low densities across the landscape. North American shorebirds that breed in forested peatlands include common snipe (Gallinago gallinago) and yellowlegs (Tringa species). Commonly encountered waterfowl are northern pintail (Anas acuta), scaup (Aythya species), and scoters (Melanitta species).
The boreal forest is the home of relatively few species of insects, but extensive and usually uniform areas of habitat periodically support high populations of species that do live there. The boreal forest lacks the elaborate complexes of invertebrate predators and parasites that serve as stabilizers of the insect populations in warmer regions. As a result, boreal insect populations occasionally increase rapidly and cause outbreaks. Some outbreaks can injure or kill trees across widespread areas of the boreal forest. Once an outbreak reaches a certain size it can become self-sustaining, much like a forest fire; the effects of the spruce budworm and spruce beetle in North America are well-documented examples. Outbreaks can be triggered by unusual weather or physical injuries that stress trees and make them vulnerable to the insects; they can end for a variety of reasons, including production of defensive chemicals by the host plants or depletion of susceptible host plants.
Perhaps the insects most noticeable to humans in the boreal forest are mosquitoes, which belong to several species. Mosquitoes feed on and are fed upon by many of the birds of the boreal forest. Wetland areas of the boreal region, such as sites having poor drainage because of permafrost, provide extensive mosquito breeding sites. Where well-oxygenated, flowing water is found, biting flies are abundant. Almost all food webs that support fish in boreal forest streams are dependent on insects.
Conifers serve as hosts for a variety of wood-boring beetles, spruce beetles, bark beetles, and ips beetles (Ips species). These insects aid in wood decomposition and nutrient release. Some beetles have outer shells with specialized indentations specifically matched to the shape and size of the spores of wood-decomposing fungi. Fungal spores become securely lodged in these cuplike structures. As the beetles burrow into wood they inoculate it with fungi.
A variety of lepidopterans (moths and butterflies) are adapted to feeding on the leaves of boreal trees. These include defoliators and leaf rollers.
The species richness and total biomass of soil organisms are significantly lower in the boreal forest than they are at lower latitudes. Dominant soil organisms are protozoans, nematodes, rotifers, and tardigrades. These organisms live primarily in soil water film and soil pore water. The soil fauna of the boreal forest region is distinctive because it generally lacks large invertebrates such as millipedes, isopods (springtails), and earthworms, especially in the middle and northern taiga. Larger soil invertebrate animals perform the function of biting off (shredding) pieces of leaf litter in forest soils and passing them through their guts. As a result of this activity thick layer of several years’ accumulation of only partially decomposed plant material is characteristic of boreal forest soils.
Fungi are the dominant organisms in the task of decomposition of boreal forest litter, but flushes of bacterial growth occur in response to triggering factors. The soil animals generally do not attack the forest litter directly but instead exert their influence by grazing on the fungi and bacteria. The rate of decomposition in boreal forest soils does not keep pace with the rate of production, causing the progressive accumulation of organic matter. At middle depths of the forest floor small invertebrates, especially dipteran larvae, partially consume or skeletonize leaf litter before emerging as adults.
The structure of the biological community in boreal forests is determined by the severity and frequency of natural and human-caused disturbances. Forest fires and logging are the largest sources of ecological disturbance in this biome.
The boreal forest is well adapted to development following natural disturbances, which include fire, floods, snow breakage, and insect outbreaks. Characteristic of the boreal forest is the general lack of late successional species that develop under an intact forest canopy.
Fire is the primary agent responsible for natural disturbances in the boreal forest. It can result from natural causes, such as lightning, or it can be set by humans. Large-scale insect outbreaks can weaken or kill trees over vast areas, thus creating an environment less resistant to fire. In the period between 1981 and 1989 an estimated 3 million hectares (7.4 million acres) burned annually in the former Soviet Union, almost all of which occurred within the taiga region of Russia. The so-called Black Dragon Fire of 1987 in China and Russia may have been the largest single fire in the world in the past several hundred years. During the 20th century, about 1 million hectares (2.5 million acres) of taiga in Canada have burned annually; a great majority of the burning occurred in the less accessible boreal forest of the northern and western parts of the country. In Alaska in years that have prolonged hot and dry periods of summer weather, millions of hectares burn, primarily in a few very large fires. Intervals of about 200 years occur between fires in the uplands of northwestern Canada and in the interior of Alaska. In much of the central and western boreal forest of North America replacement of vegetation on upland sites, presumably by fire, appears to be necessary for forest regeneration. Floodplain islands usually do not burn and contain white spruce trees as old as 400 years. In the northern boreal forest of Europe a pattern of periodic light ground fires in Scotch pine forests was typical before the era of fire control. The thick bark of these mature trees allowed them to survive these fires. In much of the boreal forest only wildland fires that threaten high-value resources are actively suppressed. Complete fire suppression would cause soil temperature to decline gradually, promoting permafrost development that would cause a significant decrease in site productivity.
Jack pine and lodgepole pine have cones that remain closed on the tree (serotinous), and black spruce has semiserotinous cones; these cones do not open to release their seeds until a wax layer is melted by the heat of fire. White spruce seedlings require the bare mineral soil produced by burning of thick organic layers of the forest floor for proper establishment; they may time their periodic production of seed to dry periods when fire is more likely.
Different degrees of forest development have had various effects on biodiversity around the circumpolar boreal forest zone.
A highly developed forest industry based on intensive forest utilization is maintained in boreal Scandinavian countries and Finland. About 95 percent of the productive forest types of Finland and the Scandinavian countries have been harvested at least once. Finland is located almost entirely within the boreal region and is one of the most forested nations in the world. About 6.5 percent of Finnish land, which includes large areas of marginal forest, woodland, and tundra, is protected from human modification. Only about 3 percent of Swedish forests are protected, most of which is concentrated in marginal forests of mountainous regions. Between 10 and 15 percent of species in Swedish forests are threatened.
The Canadian boreal forest represents nearly 7.5 percent of the Earth’s forested area. Much of the harvesting of Canadian forest has been carried out in primary (previously unlogged) forest. Nearly all the mature first-growth timber, especially of southern, central, and eastern boreal Canada, is anticipated to be removed by the late 1990s. Considerable effort has been devoted to forest regeneration and tending of new stands, although a certain amount of land does not meet reforestation goals.
In Alaska the amount of land with at least 10 percent forest cover in the boreal region is estimated at about 46 million hectares (114 million acres), or 12 percent of the state, only 5.5 million hectares (13.6 million acres) of which is considered productive timberland. Of all areas in the world, Alaska probably has the largest percentage of its surface area, about 40 percent, devoted to strict protection of natural habitats and species. Local-scale logging traditionally has been carried out for much of the 20th century. Plans to accelerate logging are being considered.
The taiga of Siberia covers 680 million hectares (1.7 billion acres) and represents nearly 19 percent of the world’s forested area and possibly 25 percent of the world’s forest volume. About 400,000 hectares (990,000 acres) of the Russian taiga are logged annually, and nearly an equal area is burned, with perhaps half of the burned area resulting from destructive fires of human origin. Social and economic problems in the early postcommunist era have slowed the amount of logging by one-third to one-half; however, several large-scale joint ventures between Russian organizations and foreign partners to harvest the forests are under way. The fate of the Siberian boreal forest has become a matter of international concern.
Large areas, perhaps exceeding 2 million hectares (4.9 million acres), of the Russian taiga near Norilsk and the Kola Peninsula have been destroyed by air pollution. Many oil pipelines are leaking in Siberia, and repairs and maintenance are minimal. In July through September 1994 more than 150,000 metric tons of crude oil were spilled in the Kolva, Usa, and Pechora river basins of the republic of Komi in Russia. Spring meltwater could carry spilled oil in this region into the northward-flowing rivers that empty into the Arctic Ocean.
Primary productivity (the rate at which photosynthesis occurs) of boreal forest ecosystems often is limited by cold soil temperatures. Net annual primary production (the total amount of productivity less that used by photosynthetic organisms in cellular respiration) in boreal forest types varies greatly, from slightly more than 2 metric tons per hectare near the polar tree limit to about 10 metric tons per hectare along its southern margin. Boreal forests are estimated to contain about 18 percent of the Earth’s total biomass (the dry weight of organic matter). The boreal forest or taiga of Siberia alone represents 57 percent of the Earth’s coniferous wood volume. Ecosystems and soils of the boreal region store a significant amount of the Earth’s carbon in the form of dead but undecomposed or partially decomposed organic matter. Global warming or land use changes could enhance decomposition, leading to the release of increased amounts of stored carbon into the atmosphere in the form of the greenhouse gas carbon dioxide.
Although most of the areas cleared for crops and grazing represent permanent and continuing deforestation, deforestation can be transient. About half of eastern North America lay deforested in the 1870s, almost all of it having been deforested at least once since European colonization in the early 1600s. Since the 1870s the region’s forest cover has increased, though most of the trees are relatively young. Few places exist in eastern North America that retain stands of uncut old-growth forests. In addition, while some forests are being cleared, some are being planted. The United Nations Food and Agriculture Organization (FAO) estimates that there are approximately 1.3 million square km (500,000 square miles) of such plantations on Earth. These are often of eucalyptus or fast-growing pines—and almost always of species that are not native to the places where they are planted.
Elsewhere, forests are shrinking. The FAO estimates that the annual rate of deforestation is about 1.3 million square km (500,000 square miles) per decade. About half of that is primary forest—forest that has not been cut previously (or at least recently). The greatest deforestation is occurring in the tropics, where a wide variety of forests exists. They range from rainforests that are hot and wet year-round to forests that are merely humid and moist, to those in which trees in varying proportions lose their leaves in the dry season, and to dry open woodlands. Because boundaries between these categories are inevitably arbitrary, estimates differ in how much deforestation has occurred in the tropics.
Dry forests in general are easier to deforest and occupy than moist forests and so are particularly targeted by human actions. Worldwide, humid forests once covered an area of about 18 million square km (7 million square miles). Of this, about 10 million square km (3.9 million square miles) remained in the early 21st century. Given the current annual rates of deforestation, most of these forests will be cleared within the century. Indeed, in some places, such as West Africa and the coastal humid forests of Brazil, very little forest remains today.
The human activities that contribute to tropical deforestation include commercial logging and land clearing for cattle ranches and plantations of rubber trees, oil palms, and other economically valuable trees. Another major contributor is the practice of slash-and-burn agriculture, or swidden agriculture. Small-scale farmers clear forests by burning them and then grow their crops in the soils fertilized by the ashes. Typically, the land produces for only a few years and then must be abandoned and new patches of forest burned.
The Amazon Rainforest is the largest remaining block of humid tropical forest, and about two-thirds of it is in Brazil. (The rest lies along that country’s borders to the west and to the north.) Detailed studies of Amazon deforestation from 1988 to 2005 show that the rate of forest clearing has varied from a low of about 11,000 square km (4,200 square miles) per year in 1991 to a high of about 30,000 square km (11,600 square miles) per year in 1995. The high figure immediately followed an El Niño, a repeatedly occurring global weather anomaly that causes the Amazon basin to receive relatively little rain and so makes its forests unusually susceptible to fires. Studies in the Amazon also reveal that 10,000 to 15,000 square km (3,900 to 5,800 square miles) are partially logged each year, a rate roughly equal to the low end of the forest clearing estimates cited above. In addition, each year fires burn an area about half as large as the areas that are cleared. Even when the forest is not entirely cleared, what remains is often a patchwork of forests and fields or, in the event of more intensive deforestation, “islands” of forest surrounded by a “sea” of deforested areas.
The effects of forest clearing, selective logging, and fires interact. Selective logging increases the flammability of the forest because it converts a closed, wetter forest into a more open, drier one. This leaves the forest vulnerable to the accidental movement of fires from cleared adjacent agricultural lands and to the killing effects of natural droughts. As fires, logging, and droughts continue, the forest can become progressively more open until all the trees are lost.
Although forests may regrow after being cleared and then abandoned, this is not always the case. About 400,000 square km (154,000 square miles) of tropical deforested land exists in the form of steep mountain hillsides. The combination of steep slopes, high rainfall, and the lack of tree roots to bind the soil can lead to disastrous landslides that destroy fields, homes, and human lives. Steep slopes aside, only about one-fourth of the humid forests that have been cleared are exploited as croplands. The rest are abandoned or used for grazing land that often can support only low densities of animals, because the soils underlying much of this land are extremely poor in nutrients. (To clear forests, the vegetation that contains most of the nutrients is often burned, and the nutrients literally “go up in smoke” or are washed away in the next rain.)
Deforestation has important global consequences. Forests sequester carbon in the form of wood and other biomass as the trees grow, taking up carbon dioxide from the atmosphere. When forests are burned, their carbon is returned to the atmosphere as carbon dioxide, a greenhouse gas that has the potential to alter global climate, and the trees are no longer present to sequester more carbon. In addition, most of the planet’s valuable biodiversity is within forests, particularly tropical ones. Moist tropical forests such as the Amazon have the greatest concentrations of animal and plant species of any terrestrial ecosystem. Perhaps two-thirds of Earth’s species live only in these forests. As deforestation proceeds, it has the potential to cause the extinction of increasing numbers of these species.

The coastal forest of Rio de Janeiro state, Braz., badly fragmented as portions were cleared for cattle grazing. Courtesy, Stuart L. Pimm
The world’s forests range from the Equator to the high latitudes. Several individual forests are notable for the role they play in maintaining the global ecological balance, the economic and recreational resources they provide, or their value as settings of significant events in human history.
The Amazon and Ituri forests are textbook examples of tropical rainforests. The Amazon Rainforest is arguably the largest rainforest in the world, covering a vast portion of the northern part of South America. The Ituri, on the other hand, encompasses only a small part of Africa’s vast Congo River basin. Despite their differences in size, both regions possess tremendous biological diversity.
The Amazon Rainforest is a large, tropical rainforest occupying the drainage basin of the Amazon River and its tributaries in northern South America, and covering an area of 6 million square km (2.3 million square miles). Comprising about 40 percent of Brazil’s total area, it is bounded by the Guiana Highlands to the north, the Andes Mountain Ranges to the west, the Brazilian central plateau to the south, and the Atlantic Ocean to the east.
Amazonia is the largest river basin in the world, and its forest stretches from the Atlantic Ocean in the east to the tree line of the Andes in the west. The forest widens from a 320-km (200-mile) front along the Atlantic to a belt 1,900 km (1,200 miles) wide where the lowlands meet the Andean foothills. The immense extent and great continuity of this rainforest is a reflection of the high rainfall, high humidity, and monotonously high temperatures that prevail in the region.
Canoe on the Negro River in the Amazon Rainforest, Amazonas state, northern Brazil. Union Press/Bruce Coleman, Inc., New York
The Amazon Rainforest is the world’s richest and most varied biological reservoir, containing several million species of insects, plants, birds, and other forms of life, many still unrecorded by science. The luxuriant vegetation encompasses a wide variety of trees, including many species of myrtle, laurel, palm, and acacia, as well as rosewood, Brazil nut, and rubber tree. Excellent timber is furnished by the mahogany and the Amazonian cedar. Major wildlife includes jaguar, manatee, tapir, red deer, capybara and many other types of rodents, and several types of monkeys.
In the 20th century, Brazil’s rapidly growing population settled major areas of the Amazon Rainforest. The Amazon forest shrank dramatically as a result of settlers’ clearance of the land to obtain lumber and to create grazing pastures and farmland. In the 1990s the Brazilian government and various international bodies began efforts to protect parts of the forest from human encroachment, exploitation, and destruction.
The overwhelmingly dominant feature of the Amazon basin is the tropical rainforest, or selva, which has a bewildering complexity and prodigious variety of trees. Indeed, as many as 100 arboreal species have been counted on a single acre of forest, with few of them occurring more than once. The Amazon forest has a strikingly layered structure. The sun-loving giants of the uppermost reaches, the canopy, soar as high as 40 metres (120 feet) above the ground; occasional individual trees, known as emergents, rise beyond the canopy, frequently attaining heights of 60 metres (200 feet). Their straight, whitish trunks are splotched with lichens and fungi. A characteristic of these giant trees is their buttresses, the basal enlargements of their trunks, which help stabilize the top-heavy trees during infrequent heavy winds. Further characteristics of the canopy trees are their narrow, downward-pointing “drip-tip” leaves, which easily shed water, and their cauliflory (the production of flowers directly from the trunks rather than from the branches). Flowers are inconspicuous. Among the prominent members of the canopy species, which capture most of the sunlight and conduct most of the photosynthesis, are rubber trees (Hevea brasiliensis), silk-cotton trees (Ceiba pentandra), Brazil nut trees (Bertholletia excelsa), sapucaia trees (Lecythis), and sucupira trees (Bowdichia). Below the canopy are two or three levels of shade-tolerant trees, including certain species of palms—of the genera Mauritia, Orbignya, and Euterpe. Myrtles, laurels, bignonias, figs, Spanish cedars, mahogany, and rosewoods are also common. They support a myriad of epiphytes (plants living on other plants)—such as orchids, bromeliads, and cacti—as well as ferns and mosses. The entire system is laced together by a bewildering network of woody ropelike vines known as lianas.
In addition to the rainforests of the terra firme, there are two types of inundated rainforests, várzea and igapó, which constitute about 3 percent of the total Amazon Rainforest. Várzea forests can be found in the silt- and nutrient-rich floodplains of whitewater rivers such as the Madeira and the Amazon, with their ever-changing mosaic of lakes, marshes, sandbars, abandoned channels, and natural levees. They are generally not as high, diverse, or old as those of the terra firme, and they are subject to periodic destruction by floods and human manipulation. (The várzea and its flood-free margins attract the most human settlement.) Wild cane (Gynerium) and aquatic herbs and grasses, as well as fast-growing pioneer tree species of the genera Cecropia, Ficus, and Erythrina, are conspicuous.
Igapó forests grow along the sandy floodplains of blackwater and clearwater rivers such as the Negro, the Tapajós, and the Trombetas. These forests may reach a maximum flood level of 12 metres (40 feet) for up to half the year, but they can be accessed by canoe.
The lowland rainforest on the Andean fringe grades into a discontinuous, tangled montane or cloud forest of misshapen trees cloaked with mosses, lichens, and bromeliads. There the cinchona, or fever-bark tree, once exploited for its antimalarial agent quinine, can be found. At still higher elevations is the grass and shrub growth of the páramo zones and cold Antiplano region.
Along the drier, southern margin of the Amazon basin, high forest gives way to the immense cerrado (scrub savanna), campo (grassy savanna), and caatinga (heath forest). The latter is characteristic of parts of the Mato Grosso Plateau, where taller forest is restricted to the stream courses and swales (marshy depressions) that dissect the upland surface. On the sandy soils of the lower Negro and the Branco drainage areas, and locally in Amapá, grassy savannas dotted with stunted trees replace the high forest. Large areas of grassy savannas can be found on the Mato Grosso Plateau, Marajó Island, the Llanos, and elsewhere.
To give a succinct overview of the complete fauna of Amazonia is as impossible as it is to adequately describe the great diversity of its flora; in part this is because many of the region’s species have yet to be identified. The rivers and streams of the basin teem with life, and the forest canopy resonates with the cries of birds and monkeys and the whine of insects. There is a notable paucity of large terrestrial mammal species; indeed, many of the mammals are arboreal.
More than 8,000 species of insects alone have been collected and classified. Myriads of mosquitoes may transmit diseases including malaria and yellow fever. Leaf-cutter ants (of the genera Atta and Acromyrmex) are prevalent, as are the ubiquitous small black flies known as piums in Brazil. Fireflies, stinging bees, hornets, wasps, beetles, cockroaches, cicadas, centipedes, scorpions, ticks, red bugs, and giant spiders are abundant. Most spectacular, however, are the hundreds of species of brilliantly coloured butterflies; sometimes thousands of butterflies gather in the afternoon on riverside sandbanks.
About 1,500 fish species have been found within the Amazon system, but many more remain unidentified. Most fish are migratory, moving in great schools at spawning time. Fish represent a critical source of protein in the often meat-poor diet of the caboclo population. (The term caboclo refers to people of mixed European and Indian ancestry in Brazil who live off the rivers and forests.) Among the more important commercial species are the pirarucu (Arapaima gigas), one of the world’s largest freshwater fish, and various giant catfish. The small, flesh-eating piranha generally feeds on other fish but may attack any animal or human that enters the water; its razor-sharp teeth cut out chunks of flesh, stripping a carcass of its meat in a few minutes. Some fish species have become locally threatened as the worldwide demand for frozen and dried fish has increased. Tropical fish of the Amazon are also some of the most desired for food and breeding and for use as aquarium specimens. Iquitos, Manaus, and the Colombian port of Leticia are centres of these trades.
Alligators are hunted for their skins; river turtles and their eggs are considered a delicacy; and the giant sea cow, or manatee, is sought for its flesh and for oil. All are threatened by overhunting, and the manatee has been listed as an endangered species. Aquatic animals also include river dolphins (Inia geoffrensis); the semiaquatic capybara, the largest rodent in the world (weighing up to 80 kg [170 pounds]); and the nutria, or coypu, valued especially for its pelt. Other common rodents are the paca, agouti, porcupine, and local species of squirrels, rats, and mice.
The tapir, the white-lipped peccary, and several species of deer are native to the Amazon basin and much sought for their meat. Water buffalo, introduced from Southeast Asia as work and dairy cattle, predominate in the remote, swampy parts of Marajó Island.
Especially characteristic of the Amazon forest are several species of monkeys. Of note are the howler monkeys, which make the selva resound with their morning and evening choruses. The small, agile squirrel monkey, the most ubiquitous of Amazonia’s monkeys, is used in laboratories, as is the larger spider monkey. Among a host of other primate species are woolly monkeys, capuchin monkeys, titis, sakis, and marmosets. All species are used for food and frequently are seen for sale in local markets. As the human population increases and the shotgun replaces the blowgun, hunting of the wild fauna has been mounting.
Large cats, such as the jaguar and ocelot, have become rare, though pumas may be found in larger numbers in the Andean fringe of the basin. Smaller carnivores include coati, grisons, and weasels. Countless bats inhabit the Amazonian night, including the blood-eating vampire bat.
Other animals of the forest include two varieties of arboreal sloths, three types of anteaters, armadillos, and iguanas, the last especially prized for their flesh. Among snakes the nonpoisonous boa constrictor and anaconda are notable for their size, the latter reaching lengths up to 9 metres (30 feet).
The Amazon basin is exceedingly rich in birdlife. Morning and evening, the parrots and macaws fly to and from their feeding grounds, their brilliant plumage flashing in the sunlight and their raucous voices calling out their presence. Throughout the day the caciques quarrel in trees where their hanging nests swing by the dozens. Hoatzins screech in noisy flocks from streamside brush, while solitary hawks and eagles scream from tree stumps. Everywhere is heard the twittering of small birds, the sound of woodpeckers, and the guttural noises of such waterbirds as herons, cormorants, roseate spoonbills, and scarlet ibises. Parakeets, which are more common in the Amazon than sparrows are in the United States, fly around in great flocks. At dusk, toucans cry a discordant plaint from the treetops and are joined by ground-dwelling tinamous and quail. The night air is filled with the cries of various species of nightjars.
The Ituri Forest is a dense tropical rainforest lying on the northeastern lip of the Congo River basin in the Central African nation of the Democratic Republic of the Congo. Situated between 0° and 3° N latitude and 27° and 30° E longitude, the precise geographic limits of the Ituri are poorly defined, especially along its southern and western extensions. The Ituri is bounded to the north and northeast by savanna and in the east by the fertile highlands of the Western Rift Valley, while to the south and west it is contiguous with the lowland rainforest, where its rivers drain into the Congo River. The total area of the Ituri Forest is approximately 62,900 square km (24,300 square miles). The forest, which is inhabited by both Bantu-speaking and Pygmy peoples, owes its name to the Ituri River, which flows east-west across the forest into the Aruwimi River and thence to the Congo.
The magnificence of the tropical rainforest of the Ituri cannot help but inspire the modern-day observer with the same poetic enthusiasm displayed by the famous Welsh explorer Henry Morton Stanley when he described his crossing of the area in 1887–88. The trees of the forest range in size from small saplings just inches in diameter to gigantic hardwoods reaching to heights of 52 metres (170 feet). Like the pillars of a Gothic cathedral, these giant trees are buttressed; roots run down their sides and extend great distances across the forest floor, making the ground a labyrinth of roots that anchor the trees and grab scarce nutrients from the shallow forest soil. In places where the high canopy is nearly continuous, only small, elusive patches of sunlight reach the forest floor. The lack of light at lower levels is accentuated by the darkness of the foliage of the few shrubs and small trees that can grow under such shaded conditions. Where gaps occur in the upper canopy, herbaceous plants with long leaves resembling those of the banana plant take advantage of the available light and grow in dense stands. In many places the forest has been disturbed, either by human activity or by natural treefalls that cut large swathes through the canopy and open up the forest to the strong equatorial sun. There, the vegetation near the ground is a dense tangle of nettles, creepers, and competing species of fast-growing, short-lived trees, which make walking difficult if not impossible. Everywhere on the ground there is a profusion of fallen nuts and fruits, some as large as basketballs and many partially eaten by monkeys, antelope (duikers), and rodents. During some seasons the air is filled with the nectar of numerous species of flower, including many epiphytes, which cling to the surface of other plants and draw their sustenance from the air. Always there is the sound of myriad insects. Cicadas perch on tree trunks and emit an irritating buzz that seems designed to drive any intruder to madness. Army ants advance in columns, audibly cracking the bodies of their insect prey. Seemingly endless lines of migrating butterflies flutter through the understory and sometimes congregate in colourful displays along streambeds. The buzzing of bees, busily plying the treetops in search of sweet nourishing nectar, is ever present. While magnificent, the forest with its constant high humidity and dark interior may seem oppressive to some. Certainly Joseph Conrad thought so when he referred to the forest as the “heart of darkness.” But the overwhelming impression for even the most squeamish visitor is not of darkness, not of oppressive gloom, but of life in its most vibrant and exciting form.

The Congo River basin and its drainage network. Encyclopædia Britannica, Inc.
The Ituri Forest varies in altitude between 700 metres (2,300 feet) in its southern portions to 1,000 metres (3,300 feet) in the north. The topography is gently undulating in the south, but in the north there are frequent outcroppings of smooth granite that rise several hundred feet above the forest.
Steeped in the tannin-rich leaves covering the forest topsoil, the water flowing in the numerous streams that drain the Ituri is the colour of strong tea. Besides the Ituri River itself, there are many broad streams that flow generally from east to west. The most notable are the Nepoko in the north, the Epulu and Nduye in the centre, and the Ibina in the south. None of these rivers is navigable, even by pirogue, for more than a few miles. The streams are fed by rains that are highly variable from month to month and from year to year. Average annual rainfall is 1,900 mm (75 inches), and there are approximately 2,000 hours of sunshine per year. Average temperature at lower elevations is 31 °C (88 °F). There is a dry season that lasts roughly from December through February, when less than 7 inches of rain normally falls. By the end of the dry season humidity within the forest is reduced, and the smaller forest streams dry up. The heaviest rains fall in October and early November; rivers overflow their banks, and large areas of the forest become flooded, making walking through the forest or driving on the few available roads extremely difficult.
The soils of the Ituri Forest developed from granites, gneisses, and metamorphosed rock of Precambrian age. In most places the soil is sandy clay or sandy clay loam, ranging in colour from reddish brown through ochre to yellowish brown and even white. The soils are acidic, and the layer of humus is thin. If exposed to the strong equatorial sun and high rainfall, as when the forest vegetation is cleared by Bantu farmers, the soil deteriorates rapidly, recovering only if it is again taken over by secondary forest. Traditionally, farmers have practiced shifting cultivation to allow the fragile soils to regenerate.
The climax-forest vegetation left undisturbed by human occupation is characterized by three dominant species of tall, hardwood legumes in the subfamily Caesalpinioideae. In the south and west Gilbertiodendron deweverei dominates and can constitute 90 percent of the standing vegetation. The regions of the forest dominated by only a few plant species have less abundant and diverse animal life than the other, more botanically mixed areas, such as in the north and east. There, Cynometra alexandrii and Brachystegia laurentii, which together comprise less than 40 percent of the canopy, are interspersed with numerous other tall species (such as Albizia, Celtis, and Ficus).
For many generations, people residing in the Ituri have been practicing a form of agriculture that entails clearing and burning the forest, growing their crops, and then moving after several years to allow the forest to regenerate during a long fallow period. This method of shifting slash-and-burn cultivation has created a patchwork of climax vegetation interspersed with various successional stages of secondary forest on the sites of old gardens and abandoned villages. Some areas are a tangle of lianas and shrubs beneath emerging hardwood trees, while others are in less-advanced stages of succession, with large stands of umbrella trees (Musanga cecropoides). These various seral patches—combined with river valleys, swampy waters, rock outcroppings, and the most recent village and garden clearings near the roads—produce a mosaic of diverse habitats that provide cover and food for the greatest abundance of mammals in forested Africa.
Situated near the forest-savanna edge, Ituri fauna include not only species typical of the African equatorial forest but also forms, such as the hyena, that are usually found on the open savanna. The most notable species is the forest giraffe, called okapi, which is endemic to the Ituri. Numerous forest antelopes include five species of duiker, the water chevrotain, and the pygmy antelope. Leopards, genets, and mongooses are the main carnivores. The elephant, buffalo, and bongo (a kind of antelope) are present in forms slightly smaller than their savanna relatives. The Ituri also supports the greatest diversity of primates of any comparable area in the world. The many monkeys include the terrestrial anubis baboon, as well as the leaf-eating imperial black and white colobus and the owl-faced monkey. The only ape is the chimpanzee. Hundreds of species of birds have been recorded; among them, the shy Congo peacock, discovered in 1936, is perhaps the most famous.
Efforts to preserve the fauna and flora are largely confined to the Maiko National Park on the southern edge of the Ituri and the Okapi Wildlife Reserve (designated a UNESCO World Heritage site in 1996) to the northeast. Both offer some protection for such animals as the forest elephant, the okapi, the Congo peacock, the aardvark, and the chimpanzee, but poaching activities and destruction of forest habitat seriously threaten these and other species both outside and inside the park. Conservation efforts have also been disrupted by civil strife that began in the late 1990s and continued into the 21st century.
Beyond the Amazon and Ituri forests in the tropics, a number of other forests, notable for their historical, economical, or recreational significance, occur in temperate and high-latitude regions. The forested regions listed below include the Bavarian, Black, and Belovezhskaya forests of central and eastern Europe and the Tongass and Sierra National Forests of North America.
The Bavarian Forest, which is known in German-speaking regions as Bayerischer Wald, is a mountain region in east-central Bavaria Land (state) in southeastern Germany. The Bavarian Forest occupies the highlands between the Danube River valley and the Bohemian Forest along Bavaria’s eastern frontier with the Czech Republic. Located largely in the Regierungsbezirk (administrative district) of Niederbayern (Lower Bavaria), the highlands parallel the southeasterly flowing Danube for about 145 km (90 miles) from the Cham and lower Regen rivers to the Austrian border east of Passau.
The Bavarian Forest, occupying mainly granite and gneiss hills, is divided into two sections by a sharp quartz ridge known as the Pfahl. The ridge runs roughly along the Regen valley and ranges from 20 to 30 metres (65 to 100 feet) in height. The Vorderer Forest, or Danube Hills, a rolling plateau situated to the southwest between the Danube and the Pfahl, seldom rises more than 1,000 metres (3,300 feet) above sea level. Meadow, isolated farmsteads, and small hamlets dominate the landscape; only the higher and steeper slopes are still wooded. Northeast of the Pfahl is the Hinterer Forest, a higher and almost continuously forested mountain region where human settlement is confined to a few longitudinal valleys. Its highest peaks include the Grosser Arber, with an elevation of 1,456 metres (4,777 feet), and the Rachel, Lusen, Dreisesselberg, and Grosser Falkenstein.
The climate of the highlands is severe and wet, supporting only modest yields of rye, oats, and potatoes produced on small valley farms. Coniferous forest predominates, with spruce the main species at higher altitudes and a mixed woodland of spruce, silver fir, and beech found at lower levels. Lumbering, woodworking, and glass grinding are the principal industries. The tourist trade is expanding as the reputation of the Bavarian Forest as a beautiful and uncrowded holiday resort area spreads. Each year many visitors explore the Bavarian Forest National Park, where more than 98 percent of the park’s 130.8-square-km (50.5-square-mile) area is tree-covered and many species of plants, birds, and small animals thrive. Principal towns of the mountain region are Regen, Zwiesel, Waldkirchen, and Grafenau.
The Belovezhskaya Forest, which is also called the Belovezh Forest and Białowieża Forest, is a forested area in western Belarus and eastern Poland. It is one of the largest surviving areas of primeval mixed forest (pine, beech, oak, alder, and spruce) in Europe and occupies more than 1,200 square km (460 square miles). The Belovezhskaya Forest is located in Brest and Hrodna (Grodno) oblasti (provinces) of Belarus and in Podlaskie województwo (province) of eastern Poland near the headwaters of the Narev (Polish: Narew) and Lesnaya (Leśna) rivers, tributaries of the Bug. The forest has a wide range of flora (some conifers and hardwoods have attained ages of 350 to 600 or more years, heights in excess of 45 metres [150 feet], and diameters greater than 2 metres [6 feet]) and fauna (including elk, deer, lynx, and wild boar) from both western and eastern Europe. Hunted into extinction in the wild after World War I, the European bison, or wisent, was reintroduced to the Belovezhskaya with zoo-bred animals. The forest remains the European bison’s most notable home, though the animals are now also found again in other parts of Europe, including Lithuania, Russia, and Ukraine. Once the hunting grounds of kings and tsars, the Belovezhskaya is the oldest nature preserve in Europe. Both the Polish and Belarusian portions of the forest have become national parks, and both areas were designated as World Heritage sites (the Polish portion in 1979 and the Belarusian portion in 1992).
The Black Forest, which is called Schwarzwald in German, is a mountain region within Baden-Württemberg Land (state) in southwestern Germany. It is the source of the Danube and Neckar rivers. The Black Forest occupies an area of 6,009 square km (2,320 square miles) and extends toward the northeast for about 160 km (100 miles) from Säckingen on the Upper Rhine River (at the Swiss border) to Durlach (east of Karlsruhe). Its width varies from 16 to 40 km (10 to 25 miles). Structurally and topographically, it forms the counterpart of the Vosges, which lies west of the Rhine Valley. The Black Forest drops abruptly to the Rhine plain but slopes more gently toward the Neckar and Nagold valleys to the east.
It is mainly a granite highland with rounded summits, although its northern part comprises forested sandstone; and it is bordered to the south by a narrow band of lower and more fertile limestone. Divided into two parts by the deep Kinzig Valley, its highest summits—Feldberg (1,493 metres [4,897 feet]), Herzogenhorn, and Blössling—are to the south. Its northern half has an average height of 600 metres (about 2,000 feet).
The raw climate of the higher districts supports only hardy grains, but the valleys are mild with good pastureland. Oak and beech woods clothe the lower slopes, while the extensive fir forests, which gave the range its name, climb to 1,220 metres (4,000 feet). Traditional economic activities—such as lumbering, woodworking, and the manufacture of watches, clocks, and musical instruments—continue. Newer manufactures include electronic equipment and precision machinery. Tourism and winter sports are also prominent, and there are many mineral springs and spas, such as Baden-Baden and Wildbad. Principal cities are Freiburg im Breisgau, Offenburg, Rastatt, and Lahr.
This large area of evergreen coniferous forest in southwestern Idaho, U.S., is located north and east of the city of Boise. Established in 1908, it has an area of about 10,570 square km (4,080 square miles). Portions of both Frank Church–River of No Return Wilderness and Sawtooth Wilderness Area are located in the forest. Payette National Forest borders it on the north, and Challis and Sawtooth national forests adjoin it to the east.
The national forest is divided into two segments. The principal part lies east of the North Fork Payette River and is roughly teardrop-shaped, with maximum dimensions of some 175 km (110 miles) north-south and 90 km (55 miles) east-west. To the west and separated from it by the valley of the North Fork Payette River is a narrow outlying section of the forest, about 60 km (37 miles) north-south and 19 km (12 miles) east-west at its widest point; most of it, however, is much narrower. The main portion of the forest is generally steep and mountainous, the Sawtooth Range constituting much of the area; elevation decreases somewhat toward the Boise River valley in the southwest. The highest point within the forest is Big Baldy, 2,963 metres (9,722 feet) above sea level. The Middle Fork Salmon and South Fork Salmon rivers, the Middle Fork Payette River, and all three forks of the Boise River have their sources in the forest. Tree species include ponderosa and lodgepole pine, Douglas and grand fir, and Engelmann spruce. Lakes and rivers have populations of trout and salmon. Deer and elk, bobcats and lynx, wolves, bears, and otters are among the forest’s mammal species.
Following the discovery of gold north of Boise in 1862, numerous mining communities sprang up in the locality. As the deposits were depleted, many of the mining sites became lumbering or livestock-grazing outposts; ponderosa pine logging and sheep ranching were important to the local economy. Some of the mining towns, however, died out and became ghost towns of interest to tourists. Logging remains an important use of the forest.
The national forest is a popular recreation area, noted for its fishing and hunting, hiking, and rafting on the Middle Fork Salmon River. The two scenic wilderness areas on the eastern side of the forest, where commercial exploitation is forbidden, are kept virtually roadless, although the Frank Church wilderness has several airstrips. Bogus Basin Ski Resort is a winter-sports centre, 26 km (16 miles) north of Boise, which is the forest headquarters.
The Franconian Forest, which is called Frankenwald in German-speaking regions, is a forested highland region in extreme northeastern Bavaria Land (state) in east-central Germany. It forms a physical and geological link between the highlands of the Fichtel Mountains and the Thuringian Forest. About 50 km (30 miles) long, the forest descends gently north and east toward the Saale River but more precipitously west to the Bavarian Plain. Its highest point is Mount Döbra 795 metres [2,608 feet] high). Along the centre lies the watershed between the Main and the Saale basins and between the Rhine and the Elbe systems. The principal tributaries of the Main rising in the forest are the Rodach and the Hasslach; of the Saale, the Selbitz. Small hamlets lie in clearings in the heath, bog, and woods of the Franconian Forest. The chief city is Hof, to the east. Kulmbach, Kronach, and Bayreuth lie to the west.
The Mount Hood National Forest is a mountainous, heavily forested region in northwestern Oregon, U.S. The forest starts about 32 km (20 miles) east of Portland and extends southward along the Cascade Range from the Columbia River for more than 100 km (60 miles). Established in 1908 as the Oregon National Forest, it covers some 4,850 square km (1,875 square miles) of scenic mountains and streams. The forest provides timber, water, forage, wildlife habitats, and recreation; it is drained by the Columbia, Sandy, Clackamas, Hood, and White rivers and their tributaries. Douglas fir is the dominant tree species. Mount Hood (3,424 metres [11,235 feet]), near the centre of the forest, is Oregon’s highest point.

Punch Bowl Falls, Mount Hood National Forest, Oregon. B. Nelson—Shostal
Features of the national forest include Mount Hood Wilderness Area and four smaller wilderness areas, Timberline Lodge (built in 1937 on Mount Hood), Multnomah Falls (190 metres [620 feet]), Austin and Bagby hot springs, Timothy Lake, portions of the Oregon Trail, and Eagle Creek Trail, leading through a region of waterfalls. Hiking and skiing are two of the many activities available to tourists. Pacific Crest National Scenic Trail traverses the forest from north to south. The Warm Springs reservation of the Paiute, Wasco, and Warm Springs Indians adjoins the forest on the southeast, and the Columbia River Gorge National Scenic Area runs along its northern border. Headquarters are at Sandy.
The Ozark–Saint Francis National Forest is made up of forested areas mainly located in central and northwestern Arkansas, U.S., but it also includes a segment along the Mississippi River in the eastern part of the state. The forest consists of several separate units embracing parts of the Ouachita and Boston mountains and the southern end of the Springfield Plateau. The westernmost part of Ozark National Forest reaches the Oklahoma border, while St. Francis National Forest constitutes the eastern segment.
Established in 1908, Ozark National Forest covers approximately 4,048 square km (1,563 square miles). It is drained by tributaries of the Arkansas River. Hardwood trees, mainly oak and hickory, form the primary vegetation, and the undergrowth includes dogwood, maple, and redbud. Animal life is plentiful and includes white-tailed deer, black bear, rabbits, bobwhite quail, and wild turkey. The main unit constitutes a roughly rectangular area extending west-east through the southern Boston Mountains and surrounded by smaller units; the park Buffalo National River lies north of its eastern half. There are five designated wilderness areas, four in the main unit (Upper Buffalo, Hurricane Creek, Richland Creek, and East Fork) and one (Leatherwood) in the smaller unit northeast of the main unit. In the northernmost section of the forest, 23 km (14 miles) northwest of the city of Mountain View, is Blanchard Springs Caverns, a three-level cave system with two levels accessible to visitors. Magazine Mountain, at an elevation of 839 metres (2,753 feet), the highest point in Arkansas, is located in the smaller unit south of the main part.
Saint Francis National Forest, established in 1960 and administered jointly with Ozark National Forest, consists of 85 square km (33 square miles) of bottomland hardwood trees. It is named for the St. Francis River, which, along with the Mississippi River, forms the forest’s eastern boundary. The northwestern portion of the forest is located on hilly Crowley’s Ridge. Popular fishing areas and hiking trails are found in and around Storm Creek Lake and Bear Creek Lake.
Six national scenic byways cross the forests, including Arkansas Highway 7, often considered the most beautiful drive in the state. The 370 km (230 miles) of hiking trails include the 266-km (165-mile) Ozark Highlands National Recreational Trail, which begins in the southwest corner of the main unit at Lake Fort Smith, winds eastward, and terminates in the northeastern corner at Buffalo River Trail in Buffalo National River. Six waterways within the forests are designated as national wild and scenic rivers.
The Sequoia National Forest and Giant Sequoia National Monument is a large natural region of mountains and forestland in east-central California, U.S. The area is noted for its more than three dozen groves of big trees, or giant sequoias (Sequoiadendron giganteum), for which the national forest and the national monument are named.
The region lies at the southern end of the Sierra Nevada and is divided into two main sections of unequal size. The smaller, northern portion is separated from Sierra National Forest (north) by the Kings River and adjoins Kings Canyon (east) and Sequoia (south) national parks. The larger, southern section borders Sequoia National Park (north) and Inyo National Forest (northeast and east) and stretches south nearly to Bakersfield and the Mojave Desert (south). Two small, separate units of Sequoia National Forest lie immediately south and southeast of this section, and the Tule Indian Reservation adjoins its western border. The national forest, which was established in 1908, has a total area of 4,610 square km (1,780 square miles), with elevations ranging from 300 to above 3,700 metres (1,000 to above 12,000 feet). Giant Sequoia National Monument was established within Sequoia National Forest in 2000 to protect the groves of giant sequoias dotted throughout both sections of the forest. The national monument occupies a total of some 1,329 square km (513 square miles) of land and includes campgrounds and an interpretive trail.
The forest, which includes conifers, hardwoods, and chaparral as well as big trees, provides timber, water, forage, wildlife habitat, and recreation. In addition to the groves of giant sequoias, notable features include Kern River and Kings River canyons, Boyden Cavern, Balch Park (which has a notable grove of redwoods), many mountain lakes and well-stocked trout streams, and the Boole Tree, with a height of 82 metres (269 feet) and a circumference of 11 metres (35 feet), the largest known tree in any U.S. national forest. Dome Land Wilderness, one of five wilderness areas within the national forest, is a lofty region northeast of Bakersfield containing numerous rock outcroppings.
Sequoia National Forest is a popular hunting area, particularly for such large game as bears and mule deer. Trout fishing, camping, skiing, snowmobiling, and kayaking are also widely enjoyed, and the Kings and North Fork Kern rivers are major destinations for white-water rafting trips. The 1,450 km (900 miles) of trails are used by hikers, horseback riders, and off-road vehicle operators. The Pacific Crest National Scenic Trail runs through parts of the forest. Headquarters are at Porterville.
Sherwood Forest is a woodland and former royal hunting ground, county of Nottinghamshire, England, that is well known for its association with Robin Hood, the outlaw hero of medieval legend. Sherwood Forest formerly occupied almost all of western Nottinghamshire and extended into Derbyshire. Today a reduced area of woodland, mostly pine plantations, remains between Nottingham and Worksop. In the north the great ducal estates, or “dukeries,” of Welbeck, Clumber, and Thoresby have preserved parts of the forest. Many veteran oaks remain, and there is much heath. Agricultural encroachment has been limited by the poor, sandy soil. An underlying coalfield has been extensively developed since the mid-19th century.
The Sierra National Forest encompasses a region of forests and streams in central California, U.S., extending along the Sierra Nevada between Yosemite and Kings Canyon national parks (north and southeast, respectively) and bordered by Inyo (northeast), Sequoia (south), and Stanislaus (northwest) national forests. It was established in 1905 from an earlier (1893) forest reserve. It has an area of about 5,260 square km (2,030 square miles) and elevations ranging from 275 to 4,000 metres (900 to 13,000 feet). Notable features include the big trees (giant sequoias) of the Nelder and McKinley groves and the Kaiser, Dinkey Lakes, Monarch, Ansel Adams, and John Muir wilderness areas (the last two shared with Inyo National Forest). Headquarters are at Clovis.
Vegetation ranges from alpine meadows to stands of lodgepole pines, red and white firs, cedars, mountain hemlocks, and aspen. The forest and its streams provide timber, grazing land, water, and hydroelectric power, and there is some gold mining. Wildlife includes mule deer, black bears, coyotes, bobcats, foxes, marmots, porcupines, and quail. Trout, bass, and bluegills are favourite sportfishing species. The Mount Dana–Minarets Escarpment is one of the forest’s scenic highlights, many of its peaks exceeding 3,700 metres (12,000 feet). Hundreds of miles of trails are maintained in the Mammoth–High Sierra area. The highest peak, Mount Humphreys (4,263 metres [13,986 feet]), is on the Sierra Nevada crest. The Pacific Crest National Scenic Trail crosses the forest, and Devils Postpile National Monument is near its eastern edge. The Sierra Vista Scenic Byway runs for 160 km (100 miles) past some of the area’s natural highlights. The region offers fishing in numerous cold, clear streams and lakes, as well as bear, deer, and quail hunting. Winter sports such as snowmobiling and skiing are popular, as are aquatic sports such as kayaking and white-water rafting in warm weather.
The Teutoburg Forest, or Teutoburger Wald in German, is the westernmost escarpment of the Weser Hills (Weserbergland) in northeastern North Rhine-Westphalia Land (state) in northern Germany. Its wooded limestone and sandstone ridges curve from the Ems River valley southeastward in an arc approximately 100 km (60 miles) long and 6.5 to 9.5 km (4 to 6 miles) wide around the north and northeast sides of the Münsterland basin. The highest point in the Teutoburg Forest, the Velmerstot, rises to an elevation of 468 metres (1,535 feet) at the southeastern end where the range meets the Egge Mountains. The city of Bielefeld, a diversified industrial centre most famous for its linen textiles, is situated at an important pass through the hills. The Hermannsdenkmal, a colossal metal statue built in the 19th century to commemorate the Battle of the Teutoburg Forest (fought 9 CE), in which Germanic tribes led by Arminius (German: Hermann) annihilated three Roman legions, stands outside Detmold on the northeastern slope. Numerous health and holiday resorts are established in the small hill towns situated among beech and spruce forests.
The Thuringian Forest, known as Thüringerwald in German-speaking regions, is a range of forested hills and mountains in Germany, extending in an irregular line from the neighbourhood of Eisenach in west-central Thuringia southeastward to the Bavarian frontier, where it merges with the Franconian Forest. Its breadth varies from 10 to 35 km (6 to 22 miles). It nowhere rises into peaks, and, of its rounded summits, the highest, Beerberg, rises only 982 metres (3,222 feet). This range encloses many charming valleys and glens; the most prominent feature of its scenery is formed by the forests, chiefly of pines and firs. The northwest part of the system is the loftier and the more densely wooded as well as the more beautiful. The southeast part is the more populous and industrial.
The crest of the Thuringian Forest, from the Werra to the Saale rivers, is traversed by the Rennsteig, a broad path of unknown antiquity. The name probably means “frontier path,” and the path marked in fact the historical boundary between Thuringia and Franconia. It may be also regarded as part of the boundary line between north and south Germany, for dialect, customs, local names, and native costume have traditionally been different on the two sides. The area was once an iron-mining centre (until the 16th century) but is now largely given over to small industries (toy making, wood carving, and glass and china manufacturing) and to tourism in such resorts as Eisenach, Friedrichroda, Giessübel, and Oberhof.
The Tongass National Forest is a forest region and wilderness area in southeastern Alaska, U.S. It was established in 1907 by an executive order issued by President Theodore Roosevelt (formal legislation declaring it a national forest was signed into law in 1909). Tongass National Forest covers most of the Alaska panhandle and is the largest publicly owned forest in the United States. The forest was named for a Tlingit Indian group. Its approximately 68,790 square km (26,560 square miles), composed of (volcanic) mountainous offshore archipelagos and rugged fjord-indented coastline, include some of the most extensive intact remnants of virgin temperate rainforest in North America. Misty Fjords (or Fiords) and Admiralty Island national monuments are located within the forest.
Found in association with barren alpine tundra (above the tree line), lowland muskeg, and scores of tidewater glaciers that descend to the coastline, the forest is dominated by towering species of western hemlock and Sitka spruce. Its rich understory includes blueberries, skunk cabbages, and a profusion of ferns and mosses. Brown and black bears, Sitka black-tailed deer, wolves, mountain goats, river otters, mink, northern flying squirrels, seals, and numerous species of birds—including bald eagles, northern goshawks, and the elusive marbled murrelet—are part of the unusual variety of wildlife. Approximately one-third of Tongass has been protected as a national wilderness area, and about one-fifth has been designated for commercial development. The forest has long been the centre of intense conflict between conservationists and logging companies. Fishing and tourism are the forest’s most significant economic activities.