CHAPTER 3
Comparative Anatomy and Physiology
From their iridescent feathers to their tiny skeletons, the hummingbirds offer an amazing number of specializations that can be matched by few if any other bird groups. On the basis of their wings alone, the hummingbirds are preeminent among birds. The remarkable elongation of the hand bones and the associated length of the ten primary feathers are examples. The secondary feathers, however, are greatly reduced in both length and number. There typically are only six feathers, but sometimes a rudimentary seventh is present. The rectrices are also almost invariably ten, although one species—the marvelous spatuletail—has only four, two of which are highly specialized.
The skeletal characteristics (Figure 7) of hummingbirds exhibit many obvious specializations for their unique mode of flight. The sternum, which is greatly enlarged in comparison with more typical flying birds (Figure 8), is also deeply keeled, and the eight pairs of ribs (versus six in most land birds) protect it during the great stresses of hummingbird flight. Further, the unusually strong coracoid is attached to the sternum by a shallow ball-and-cup socket, in a manner that is unique to hummingbirds and swifts (Ridgway, 1890).
The tongue of a hummingbird is essentially as long as the species’ bill, yet, by virtue of the elongated hyoid bones, it can be greatly extended from the tip of the bill, thus increasing its effective length for foraging in deeply tubular flowers. For about the anterior half of its total length, the tongue is divided into two separate units, which are fringed along the membranous outer edges, probably in conjunction with the amount of insect foods usually consumed (Figure 7). The bill itself varies in length from less than a centimeter in Ramphomicron to sometimes more than 10 centimeters in Ensifera; it is often slightly decurved and is very rarely recurved. In a few genera, such as Heliothryx and Schistes, the bill is greatly compressed laterally, forming a fine tip when viewed from above. In nearly all hummingbirds the nostril is to some extent covered from above by a distinct shelf-like scale, or operculum. Whether this is related to protection of the nasal cavity from inhaling pollen and the like or has some other unrelated function is unknown.
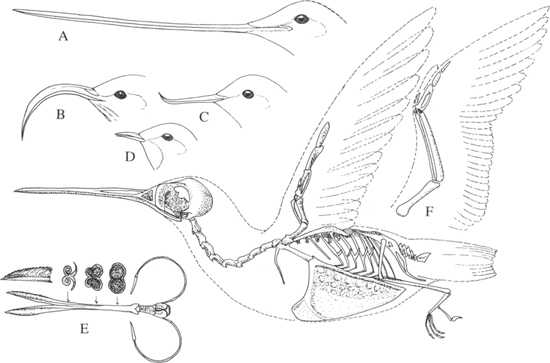
7. Skeleton, bill, and tongue characteristics of hummingbirds, including bills of (A) Andean swordbill, (B) white-tipped sicklebill, (C) avocet-bill, and (D) thornbill. The (E) tongue of the green-throated carib is also diagramed. The wing of a typical flying bird (duck)—drawn to the same scale—is included for comparison (F). (Partly after Ridgway, 1890)
In nearly all species, the primary feathers are progressively longer from the inside outwardly, and in all but a very few the outermost primary is the longest. The exceptions have an outermost primary that has obviously been modified in structure to generate sound, yet in the streamertail it is shorter than the second primary but not specialized in shape. In some genera, particularly Campylopterus and Aphantochroa, the shafts of one to three of the outermost primaries are greatly thickened in males, possibly related to a general strengthening of the outer primaries or perhaps for some unknown display purposes.
Although there is remarkable consistency in the number of tail feathers, the shape of the tail varies substantially among hummingbirds, with differential lengthening of the feathers resulting in forked, scissor-like, wedge-shaped, rounded, graduated or pointed tails, or other variations (Figure 9). In several species the outer tail feathers are appreciably narrower than the more centrally located ones, and these probably are set into vibration during aerial display; the Costa and Anna are examples among North American species. On the other hand, in the rufous male the pair of rectrices adjacent to the middle pair are curiously notched near the tip. They possibly also generate special sounds, although Baptista and Matsui (1979) suggested that, in the case of the Anna, the dive-noise is largely or entirely vocal in origin.
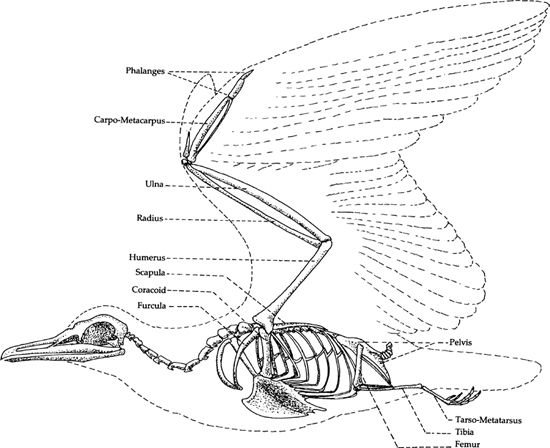
8. Skeletal characteristics of a typical flying bird (gull), for comparison with Figure 7.
Generally, neither the remiges nor rectrices of hummingbirds are extensively iridescent, perhaps because the structural specializations in the barbules responsible for this coloration seem to inhibit the ability of the separate feather barbs to “knit” together and maintain an unbroken airfoil when subjected to stresses. However, with the exception of these feathers, nearly all of the other feather regions of hummingbirds are relatively iridescent. Their body feathers are also extremely small and closely packed. Aldrich (1956) reported that a male Allen hummingbird had 1459 feathers and a female had 1659. By comparison, a brown thrasher (Toxostoma rufum) had 1920, but this species has a skin surface area about ten times larger than a ruby-throat; thus, the density of feathers in the hummingbird is approximately five times greater (Greenewalt, 1960a).
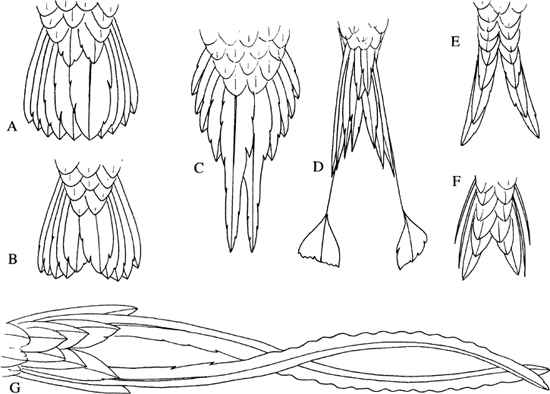
9. Variations in tail shapes of hummingbirds, including (A) rounded, (B) double-rounded, (C) pointed (long-tailed hermit), (D) racket-like (racket-tailed coquette), (E) scissor-like (amethyst woodstar), (F) forked (white-bellied woodstar), and (G) streamer-like (streamertail). (After Ridgway, 1890, 1911)
The highly iridescent feathers of the hummingbird gorgets are among the most specialized of all bird feathers. But even in the male’s gorget of a species such as the Allen hummingbird, only about the distal third of each feather is modified for iridescence; the close overlapping of adjacent feathers thus generates the unbroken color effect. The iridescence is produced by the proximal parts of the barbules, which are smooth, flattened, and lack hook-like barbicels or hamuli. Beyond the color-producing portion, the barbule is strongly narrowed and curved toward the distal tip of the feather (Figure 10). The barbicels in this area help to hold together the barbules on one side of the barb, but do not unite the barbules of adjacent barbs (Aldrich, 1956).
Greenewalt (1960a) reviewed at length the aspects of optical theory required for an adequate understanding of iridescence in hummingbird feathers; the following is a brief overview: First, the colors do not directly depend on selective pigment absorption and reflection, as do browns and blacks produced by the melanin pigments of non-iridescent feathers. Rather, they depend on “interference coloration,” such as that resulting from the colors seen in an oil film or a soap bubble. Basically, the colors depend on light being passed through a substance with a different refractive index than that of air (1.0), and being partially reflected back again at a second interface. The percentage of light that is reflected back increases with the difference in the refractive indices of the two media; in addition, the thickness of the film through which the light is passing strongly influences the wavelengths of light that are reflected back. Put simply, red wavelengths are longer than those at the violet end of the spectrum and generally require films that are thicker or have higher refractive indices than those able to refract bluish or violet light. Thus, the optimum refractive index for red feathers is about 1.85; for blue feathers it is about 1.5.
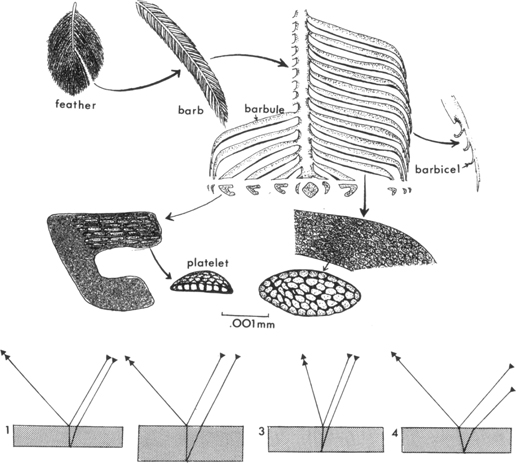
10. Iridescent feather structure of a hummingbird feather, based on photographs in Greenewalt (1960a). Below, diagrams of light pathways of optical films having a higher refractive index than that of air, showing effects of variations in film thickness (1 and 2) and in angle of viewing (3 and 4) on reinforcement of particular wavelengths. (In part after Greenewalt, 1960a)
Hummingbird feathers may attain any refractive index within this range because the iridescent portions of the barbules are densely packed with tiny, tightly packed layers of platelets. These platelets are only about 2.5 microns in length and average about 0.18 microns in thickness, but they vary in thickness and are differentially filled with air bubbles. The platelet’s matrix, probably of melanin, evidently has a refractive index of about 2.2, whereas the air bubbles inside have a refractive index of 1.0. Varying the amount of air in the platelets provides a composite refractive index that ranges from the red end of the spectrum (1.85) to the blue (1.5). An analysis by Greenewalt (1960a) indicated that the theoretical structure of the plates should have an “optical thickness” equal to one-half of the particular color’s peak wavelength, or an actual thickness equal to the optical thickness divided by the average refractive index. If, then, half the wavelength of red light (0.6 to 0.7 micron) should be divided by the average refractive index, “red” plates should be about 0.18 micron in average thickness, or the same as the actual thickness indicated by electron microscope examination.
Thus, the actual thickness of the platelets not only significantly determines the quality of the perceived light, but it also affects the amount of air held within the pigment granules and the consequent variations in interference effects. Further, a single pigment granule can produce different color effects according to the angle at which it is viewed. When an optical film is viewed from above, it reflects longer wavelengths than when viewed from angles progressively farther away from the perpendicular (Figure 10). Thus, a gorget may appear ruby red when seen with a beam of light coming from directly behind the eye, but as the angle is changed the gorget color will shift from red to blue and finally to black, as the angle of incidence increases (Greenewalt, 1960a).
In hummingbirds, the color-producing pigment platelets are closely packed into a mosaic surface, and 8 to 10 such layers are then tightly stacked on top of one another in typical iridescent feathers. Far from confusing the visual effects, such stacking actually tends to intensify and purify the resulting spectral color, which is probably why hummingbirds have possibly the most intensively iridescent feathers known in birds (Greenewalt, 1960a).
Iridescence is only one of the many respects in which hummingbirds outdo other birds. When flying, all birds expend a high amount of energy. In most strongly flying birds, the two pairs of muscles originating on the sternum’s keel occupy about 15 to 25 percent of total body weight, but in hummingbirds these muscles comprise 25 to 30 percent. Further, the muscle that elevates each wing is approximately half the weight of that responsible for the downstroke; it is usually only 5 to 10 percent in more typical birds (Greenewalt, 1960a, 1962).
The unusually large size of the muscles that elevate the wings is related to the fact that hummingbirds generate power during the upstroke as well as the downstroke, with the wings operating like a variable-pitch rotor. The pitch during the upstroke can modify the thrust generated on the downstroke, so that hummingbirds are readily capable of forward, hovering, and backward flight. These remarkable abilities are achieved by a combination of rotary movements of the outer portion of the wing as well as changes in the plane of the wing movement. Thus, during forward flight the tips of the wings describe a vertical oval in the air, not very different from that of other flying birds. However, during hovering the wings are moved in a manner approaching a horizontal figure eight, with the plane of the movement essentially parallel to the horizon (Figure 11). By a slight backward tilting of the plane the bird can easily move upward and backward, and likewise by tilting the plane downward the bird can begin to proceed slowly forward, much in the manner of a helicopter (Greenewalt, 1960a).
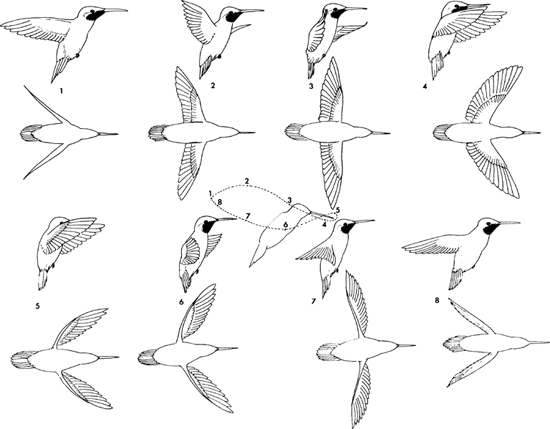
11. Stages in wing action during hovering flight in a hummingbird, in side and dorsal views. Numbered points on the central diagram indicate location of wingtip at successive stages. (Modified from Greenewalt, 1960a)
The wings of hummingbirds therefore act like aerial “oars,” lacking the flexing at the wrist and forearm joints typical of other birds and operating like mechanical oscillators that have constant-speed motors. Thus, they have a close relationship between wing length and the rate of wing beats, as do other flying birds as well as insects. In fact, hummingbirds appear between insects and other groups of birds when their wing length is plotted against their rate of wing beats, or when their wing length is plotted against average body weight (Greenewalt, 1960a). Their wing length is relatively long relative to body size, and on the average body weight is proportional to the 1.5 power of the wing length (Greenewalt, 1962). Average adult hummingbirds weigh from about 2.1 grams in the vervain species (Lack, 1976) to a maximum of about 22 grams in the giant hummingbird (Lasiewski et al., 1967). The wingbeat rate of the former (as well that of the slightly smaller bee hummingbird) is still undetermined, but in the very slightly larger amethyst woodstar it averages 80 per second (Greenewalt, 1960a). However, that of the giant hummingbird ranges from about 10 to 15 per second. In spite of this species’ remarkably large size, it is perfectly capable of controlled hovering and backward flight (Lasiewski et al., 1967).
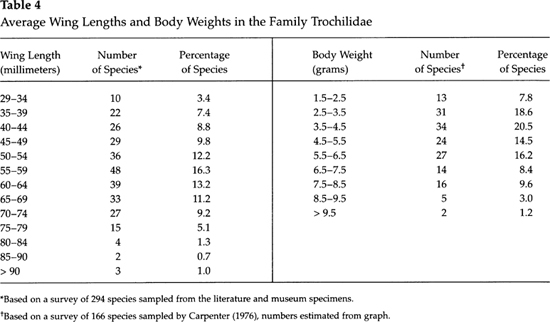
Plots of frequency distributions of average body weights and average adult wing lengths of hummingbirds reveal some interesting relationships. Table 4 presents the range of adult weights of 166 species as determined by Carpenter (1976), as well as the average adult wing lengths of nearly 300 hummingbirds, as obtained from museum specimens and extracted from literature. The latter total represents about 85 percent of the known species of hummingbirds (data on the remaining species were not readily available to me). According to the table, hummingbirds exhibit an apparent “adaptive aerodynamic peak” in weight at about 4 grams. Likewise there is a distinct peak for wing length between 55 and 60 millimeters, although ecologic factors may dictate other size constraints. Near the lower end of the scale, the average weights of female hummingbirds are somewhat greater than those of males, but among the larger species of hummingbirds the reverse is true. This may be because the effects of sexual selection in the larger species favor male dominance, whereas in the relatively small species the energy drain of egg-laying has fostered selection favoring larger female weights as compared with males. Thus, the female vervain hummingbird lays a clutch of two eggs of about 0.37 grams each, and their combined weight equals 34 percent of the average adult weight (Lack, 1976). In the larger species the relative egg weight is considerably less, and the females spend less energy in caring for the relatively smaller young (Brown et al., 1978).
Thus, the hummingbirds seem to have evolved along a very narrow evolutionary corridor, which has kept their body weights low enough to meet the energy requirements of their unique flight and associated hovering abilities and has prevented them from becoming larger than their limited food supplies would support. They have also been limited by constraints at the lower end of their potential body size by problems associated with regulating body heat; limits on miniaturization of body parts such as structural strength of bones and minimum brain size; and increasing vulnerability to all sorts of predators, including large insects.
Hummingbirds consume large amounts of oxygen, especially during flight, which often places their circulatory systems under special strains. They have the largest known relative heart size of all birds—up to 2.4 percent of their body weight (in the rufous-tailed)—and likewise the most rapid heartbeat in birds—1260 beats per minute (in the blue-throated hummingbird). Similarly, the density of their erythrocytes is the highest known among birds (6.59 million per cubic milliliter), perhaps as a result of the cells’ unusually small size, which is associated with efficient gas-transport capabilities. Body temperatures of active hummingbirds are generally close to 40° C, occasionally reaching as high as 43° C when struggling but restrained (Lasiewski, 1964; Morrison, 1962). The breathing rate in hummingbirds is also very high: approximately 250 per minute for a 3-gram hummingbird at rest.
The rate of metabolism of even a resting hummingbird, as measured by its oxygen consumption, is about 12 times greater than that of a pigeon (Columba livia) and 25 times greater than that of domestic fowl (Welty, 1975). A human, metabolizing energy at the rate of a hummingbird, would have to consume roughly double his weight in food such as meat every 24 hours, or about 45 kilograms of pure glucose, and his body temperature would rise to more than 400° C (Scheithauer, 1967). Hummingbirds and the comparably sized shrews among mammals have evidently reached the smallest sizes that are physiologically possible for warm-blooded animals; smaller animals simply could not eat enough food to avoid starvation.
Small birds endure a greater rate of heat loss from the body than do large birds; likewise, small birds are relatively more subject to overheating when placed in an environment that is warmer than their body temperature. Even temperate-zone hummingbirds lack down feathers; therefore, they are relatively unable to increase their insulation effectiveness by feather fluffing during exposure to cold. Thus, they can maintain body temperature only by increasing their heat production. Working on marginal energy balances, hummingbirds cannot accomplish such maintenance over a prolonged period; so, rather than make the attempt, many enter a state of torpor. The metabolic rate may then drop to about one-fiftieth of the basal rate at normal body temperature, and the rate of water loss by evaporation decreases by one-third to one-tenth as compared with that at normal body temperature; thus torpidity may be important as a water-conservation device. The rates of entry into and emergence from torpor are inversely related to the bird’s body size: The smaller the hummingbird, the more rapidly it may enter or emerge from a torpid state.
During torpidity, the heartbeat rate varies with body temperature, from 50 to 180 per minute, and breathing becomes irregular, with long periods of nonbreathing at lower temperatures (Lasiewski, 1964). In many torpid birds the body temperature tends to approximate that of the environment, and in species such as the poorwill (Phalaenoptilus nuttallii) lowering the external temperature does not stimulate regulatory processes that prevent the body temperature from either falling further or from arousing. However, a few hummingbird species maintain a minimum body temperature of 18 to 20° C below that of the normal resting temperature in spite of even lower air temperatures. Apparently the regulated level of body temperature during torpidity is related to the minimum environmental conditions that are encountered under normal conditions in the wild (Wolf and Hainsworth, 1971). A study of several species of South American hummingbirds found that average body temperatures during daytime activities were about 39° C, with an increase of 2.2° C during maximum activity and a decrease of only 4° to 5° C during deep sleep or torpor conditions. Tropical species of hummingbirds thus seem more sensitive to a lowering of body temperature than are more temperate-adapted species (Morrison, 1962).
We have long wondered how species such as the ruby-throated hummingbird store enough energy to allow them to migrate across the Gulf of Mexico, a minimum overseas distance of about 800 kilometers, given their limited capacities for fat storage and the high rate of energy utilization during flight. Although earlier calculations suggested that the maximum flight range of this species might be about 616 kilometers, or far below the necessary minimum to make the flight, more recent estimates by Lasiewski (1962) have prompted new conclusions. He estimated that the average adult ruby-throat can store and use 2 grams of fat, which would be enough for about 26 hours flight in males and 24.3 hours in the larger females. Given an average air speed of 40 kilometers per hour, the maximum flight range of a male would thus be 1040 kilometers, and that of a female 975 kilometers—more than enough to make a nonstop flight across the gulf. This was based on the estimate that the birds would burn their energy reserves at the rate of 0.69 and 0.74 Calories per hour for the male and female sexes, respectively, and the fact that a gram of fat has an energy content of 9.0 Calories.
A hummingbird must spend a good proportion of its waking hours gathering food simply to stay alive. One wild male Anna hummingbird requires a minimum of 7.55 Calories (assuming torpidity at night) to 10.32 Calories (assuming sleep at night) during a 24-hour period. An average daily period of 12 hours and 52 minutes of activity would require an energy expenditure of 3.81 Calories in perching, 2.46 Calories in nectar flights, 0.09 Calories in insect-catching, and 0.30 Calories in defense of territory. The nectar production of about 1022 fuschia flowers could supply this daily need (Pearson, 1954). For the hummingbird to spend the night without becoming torpid, it must consume a substantial surplus of energy during the hours of activity; one study on the rufous-tailed hummingbird estimated that an excess daily intake of 4.07 Calories would enable the bird to survive the night without going into torpor (Schuchmann et al., 1979). Studies on a few species of hummingbirds indicated daily energy budgets of 7.78 to 12.4 Calories per 24 hours. These energy budgets are 3.1 to 3.6 times the standard (“basal”) metabolic rates of the individual species, or considerably greater than figures obtained for comparable nectar-feeders among passerine birds (MacMillen and Carpenter, 1977). For example, Powers and Nagy (1988) estimated the daily field metabolic rate of an Anna’s hummingbird weighing 4.48 grams to be about 32 kilojoules (or 7.65 Calories), which amounts to 5.2 times the estimated basal metabolic rate. During daytime hours the rate averaged 6.8 times the basal metabolic rate, but this dropped to 2.1 times greater during nighttime hours. The latter rate is similar to what might be expected in a resting bird experiencing normal body temperatures, but might higher than would be expected if the bird were in a state of torpor. Estimated daily intake of water was 164 percent of body mass (not eight times greater, as has been suggested in early literature). It is likely that enough sucrose is stored in the crop at night to provide a supplemental “energy storage depot” for the birds to draw upon for their nocturnal energy requirements (Powers, 1991).