10 Semiotic freedom: an emerging force
The term “information” has become nearly omnipresent in modern biology (and medicine). One would probably not exaggerate if
the famous saying of evolutionary biologist Theodosius Dobhzhansky, that “nothing in biology makes sense except in the light
of evolution” should nowadays be reframed as “nothing in biology makes sense except in the light of information.” But are
those two concepts, evolution and information, somehow internally related? And if so, how?
10.1 Information in evolution
In textbooks, newspapers, and even scientific papers, the meanings of the terms “evolution” and “information” are generally
supposed to be well known, and they are rarely explained. And yet, there is no general consensus in science, or even in biology,
about what they really mean. For instance, the preferred exemplar (in the Kuhnian sense) of evolution by natural selection
is that of industrial melanism. In woodlands, where industrial pollution has killed the lichens and exposed the dark brown
tree trunks, dark forms of the peppered moth – melanics – are supposedly better camouflaged against predation from birds than
are the light gray forms that predominated before the Industrial Revolution. The observation by 1950 that darker forms had
largely displaced lighter forms was thus taken as evidence for natural selection in action. This exemplar does indeed illustrate
the effect of natural selection, but whether it shows evolution depends on your idea of evolution. Thus, if by the term “evolution”

Fig. 10.1 According to the “central dogma,” the information flow in a cell is one-way. Information is passed from DNA to RNA and further
to protein, but never in the reverse direction.
is meant something like “the origin of species,” then it is remarkable that no speciation has actually taken place here, and
it is not obvious that any speciation would indeed occur in a case like this. So, populations do indeed change as a result
of natural selection, but is this mechanism also behind speciation and evolution at large? The majority of biologists certainly
feel assured that macroevolution – that is, evolution above the level of species – is in fact the tardy result of an infinitely
ongoing microevolution (adaptation in populations), but there remains serious disagreement on this (Depew and Weber, 1995; Gould, 2002).
Likewise, the meaning of the term “information” is loaded with ambiguities. Whereas theorists may eradicate such ambiguities
by rigorous definition, it is often less than clear how well-defined concepts of information relate to the actual use of information
as an explanatory tool in biology. When talking about information, most biologists probably have in mind the kind of information
expressed in the so-called “central dogma,” as stated by Francis Crick in the following terms: “Once information has passed
into protein it cannot get out again”; that is, the flow of information in the cell is unidirectional; it originates in the
gene and ends up as protein (see Figure 10.1).
Writers of textbooks are anxious to point out that the kind of information we are talking about here is “sequential information,”
which is “replicated” when the cell divides, is “transcribed” into mRNA, and is “expressed” by genes. Following transcription
to mRNA, the information is said to be “processed” or “edited,” whereupon it “migrates” from the nucleus to the cytoplasm,
where it is finally “read” by a ribosome that “translates” it into protein. However, as was shown by Sahorta Sarkar, nothing really goes on here that could not be expressed by decent biochemical processes
exhibiting traditional efficient causality (Sarkar, 1996). So, why do molecular biologists prefer to talk about “information”?
There can, certainly, be no doubt about the heuristic value of the information metaphor; something is added to our understanding
when we talk about information rather than just about chemistry. In fact, without the information metaphor, it would be hard
to understand modern biochemistry at all.1 But whatever this added understanding consists of, it is not part of the cellular reality according to molecular biologists.
At the cellular level chemistry exhausts what goes on. The reason for this insistence on the reducibility of information to
chemistry, I shall suggest, is that the heuristic value of the information concept is connected to the role that history (evolution)
plays in the life of cells and organisms. What happens is that “history talks”, but history is not considered part of biochemistry
or molecular biology. As shown by Terrence Deacon, in Chapter 8 of this volume, the contextual (historical) aspect of information is due ultimately to the fact that informative signals
are necessarily caused by externally derived perturbations of some medium away from its expected state. The receipt of an
informative signal ipso facto provides evidence of the material influence of something other than the signal itself, linking
its resultant form to this “absent cause”: the immediate or mediating object of reference. Although history is, of course,
not “doing” anything, and most certainly does not “talk”, the present form of the informative substrate – for example, the
actual sequence of nucleotides in a section of the DNA string – does nevertheless refer to absent causes: that is, causes
that are connected to the evolutionary past of the species. This referential aspect of information furthermore provides for the inherent intentionality
of biological information to the extent that it reflects the workings of natural selection. As Deacon would put it, genetic
and phenotypic representations with the least correlation with environmental regularities will not be transmitted (reproduced).
The (subconscious?) effect of the apparently unavoidable “information talk” in molecular biology may thus be to smuggle in
the intentional (semiotic) aspect of information by the back door, so to say. At the surface, genetic information is treated
as if it was just a simple causal factor, but its deep appeal to our understanding derives from its hidden connotation of
an otherwise tabooed intentionality. The exclusion of history may be relatively innocent at the level of molecular biology
itself, but when molecular biology is understood as the basis for genetics and evolution, this ahistorical understanding serves
to reify processes that are in fact embedded in contextual constraints that should be accounted for in our theories. Genes
are supposed to specify particular traits or characters such as missing eyes in a fruit fly or Huntington’s chorea in human
beings. So the question is, what does it mean that genes are carrying information about such properties?
The identification of genetic information with “sequential information,” however, reinforces and is reinforced by the belief
in genocentric versions of Darwinism. If information is nothing but a molecular property of DNA, it may be replicated and
transported down through generations in well-defined and unambiguous units open to modification by the combined processes
of mutation and selection. Natural selection will then gradually optimize the set of instructional units carried in the gene
pool of a population. What is pushed aside in this simplified view of gene function once again is the burden of context. The
concrete functioning of genes is dependent not only on the genetic background on which they happen to be placed, but also
on a host of circumstances normally described collectively as the milieu (Griffiths and Gray, 1994; Gray, Griffiths, and Oyama, 2001). The conception of genes as unambiguous or autonomous functional units does not even come true in those monogenetic diseases
that originally served as models for our ideas of gene function. Thus, the monogenetic disease PKU has now been shown to exhibit
quite unexpected phenotypic variation (Scriver and Waters, 1999). Not all untreated carriers of the “disease gene” exhibit disturbed cognitive development, probably because the build-up
of toxic concentration of the amino acid phenylalanine in the brain is influenced by unknown factors. Genetic information
does not simply “cause” things to happen.
Needless to say, proponents of the “intelligent design” movement use this kind of criticism to attack the legitimacy of evolutionary
theory. And Darwinists, on their side, respond with deep suspicion towards any criticism of the general scheme of evolutionary
theory. A third possibility exists though, and this is the possibility I am going to present and discuss in the remaining
part of this chapter: namely, the approach called biosemiotics.
10.2 Semiosis and life
Biosemiotics suggests that living systems should be studied as semiotic2 systems in their own right. This idea is based on the belief that the poverty of the information discourse in biological
sciences results from the reductive neglect of the interpretative aspect of biological information. By introducing the concept
of the sign, as developed by US chemist and philosopher Charles Sanders Peirce (1839–1914) as a substitute for information,
it will be assured that the interpretative side of information is not neglected. In everyday parlance, a sign is simply “something
that refers to something else”, like smoke refers to a fire. This reference, however, cannot be brought about without a process
of interpretation. Thus, a baby will not know that smoke signifies fire. A deer, on the other hand, will perhaps not know
that smoke refers to fire, but it definitely takes smoke as a sign of danger.
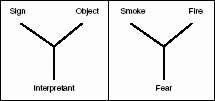
Fig. 10.2 Sign action. Left: a graphical illustration of Charles Sanders Peirce’s conception of a sign as a triadic relation connecting
the primary sign to its object through the production of an interpretant. Right: Smoke provokes the formation of an interpretant
in the brain of an animal, causing it to flee. The animal is seized by alarm, and this being “seized by alarm” is the interpretant.
In this figure, the three instances of the sign relation are connected by a tripod rather than by a triangle in order to emphasize
the internal logic of the sign relation, which should never be confused with a mere summation of three relations between corners
in a triangle.
And adult people, of course, usually turn their head in order to see where the fire might be.3 An “interpretant” is constructed by brain processes that mediate the connection between the sense impression of smoke and
the presumed existence of something burning. Thus, we get the Peircean triadic sign concept, as given in Figure 10.2.
Signs, however, are not causes in the traditional sense of (Aristotelian) efficient causality, for the effect of a sign is
not compulsory but depends upon a process of interpretation, and the interpretation may well be – and probably most often
is – “mistaken.” An example of this is when a predator is lured away from a bird’s nest because it misinterprets the clumsy
behavior of the bird as a sign for an easy catch. The bird, however, just pretended to have a broken wing and flew away the
minute the predator had missed the nest. In this semiotically interesting case, the bird profits from its “knowledge” of the
predator’s predictable habit of going for the easy catch. Whether the bird’s “knowledge” in this case is phylogenetically
(instinctually) or ontogenetically (learned) rooted does not change the general logic of the situation – what differs is only
a question of the implied mechanism and the time span of the interpretative act. On the one hand, if the bird’s behavior is
instinctually based, it is the result of an evolutionary interpretation, and we are then talking of time spans of perhaps
millions of years during which this particular habit developed in the bird as an interpretative response to the predictable
behavior of predators. If, on the other hand, the behavior is based on learning, the interpretative act is a product of brain
processes not far from the brain processes involved in our own human interpretative activity.
I said above that the heuristic value of the information concept derives from the historical nature of living systems. Although
nobody in science denies that living creatures are the result of an evolutionary process, this process is normally not conceived
as historical in the usual sense of this term, because it is assumed to have obeyed the deterministic rule of natural laws.
The metaphysics of determinism, however, seem less and less supported by the findings of modern science. It cannot, of course,
be excluded right away, but there no longer seems to be a strong reason to adopt it. In Stuart Kauffman’s book Investigations (2000), an important part of the analysis turns on the question of the non-ergodicity of the universe, meaning that the universe
never had the time it would have needed should its present state of affairs in any way be representative of its in-built possibilities.
The persistent movement of the universe into the “adjacent possible” precludes it from ever reaching a state that depends
on statistical likelihood. Instead, the universe is historical, for “History enters when the space of the possible that might
have been explored is larger, or vastly larger, than what has actually occurred” (Kauffman, 2000, p. 152).
The historical nature of the world has profound consequences for the study of life, because it confronts us with the problem
of organization in a new way. If the complex forms of organization exhibited by living systems – from the cell to the ecosystem
– are not the inescapable result of predictable lawfulness, they must instead have emerged through processes that are still in need
of discovery. The principle of natural selection, of course, greatly helps us in explaining the widespread adaptedness of
biosystems, but we need an additional principle that would solve the fundamental question of the “aboutness” of life, the
never-ending chain of attempts by living systems to come to terms with their conditions of life. As Stuart Kauffman and Philip
Clayton have formulated this: “it is a stunning fact that the universe has given rise to entities that do, daily, modify the
universe to their own ends. We shall call this capacity agency” (Clayton and Kauffman, 2006, p. 504). This “agency” necessarily involves a kind of measuring process, whereby the agent is enabled to modify its environment
in a selective way, furthering its own ends, and this measuring process is the core of the striking “aboutness” that characterizes
living systems. “Aboutness,” thus, is not derivable from the principle of natural selection for the simple reason that it
is required for natural selection to operate in the first place.4 If organisms did not exhibit aboutness, if they did not “take an interest” in the world around them (if they did not “strive”
– to use Darwin’s own term), there would be no “competition for survival” but only disorganized activity leading nowhere.
However, if natural selection is not responsible for this aboutness, what then is the basis of it? Kauffman and Clayton propose
a tentative five-part definition:
a minimal molecular autonomous agent: such a system should be able to reproduce with heritable variation, should perform at least one work cycle, should have boundaries such that it can be individuated naturally, should engage in self-propagating work and constraint construction, and should be able to choose between at least two alternatives(Clayton and Kauffman, 2006)
Terence Deacon has suggested an even more simple system as a candidate for the prebiotic emergence of life and agency, a system
that he calls the “autocell” (Deacon and Sherman, 2008). Autocells are self-assembling molecular structures that derive their individuality from a synergistic relationship between
two kinds of self-organizing processes that reciprocally depend upon one another’s persistence. Such autocells could have
been an important stepping stone in the process leading from the non-living to the living. However, as Deacon and Sherman
themselves point out, autocells are not yet full-blown living systems. They lack several features that are generally considered
criteria for being alive, such as the possession of the replicative molecules of RNA or DNA, and differential survival through
replications. Furthermore, autocells will not meet the set of criteria put forward by Kauffman and Clayton. The autocell model,
however, does demonstrate a possible unbroken continuity from thermodynamics to evolvability.
The difficult problem to solve in any theory of the origin of agency and life is how to unify two normally quite separate
kinds of dynamics: a dynamics of chemical interaction patterns and a dynamics of signification or semiosis. This immediately
places this question in the contextual situation of the environment. The Kauffman–Clayton criteria clearly do so, but they
also presuppose a far more complex beginning than Deacon’s autocell model. It remains to be seen if, or how, these two approaches
might be reconciled.
Biosemiotics, of course, immediately reminds us that “to ask for the origin of life is to ask for the origin of the environment”
(Hoffmeyer, 1998). Living organisms are inscribed in their environments much like patterns woven into a carpet – the two cannot get apart.
From a semiotic point of view, the decisive step in the process that led to the origin of life was the appearance in the world
of a new kind of asymmetry, an asymmetry between inside and outside. The formation of a closed membrane around an autocatalytically closed system of components (Kauffman, 1993) might have been an initial step. Such a membrane would have created what is probably the most essential and unique characteristic of life: the never-ending interest of the insides into their outsides or, in other words, cellular aboutness. I have suggested that this “interest” should be understood as a property that ultimately was derived from the primordial
membrane itself. A closed membrane sheet necessarily has two distinct kinds of exterior: the inside exterior and the outside
exterior. For agency to appear, such membranes must have managed not only to canalize a selective flow of chemicals across
themselves, but also to subsume their interior system of components to help them resist the flow of perturbations from the
outside exterior. Or, in other words, for the membranes to persist, they would have to function as interfaces connecting their
inside worlds to the outside world. At some point in prebiotic development, a self-referential digital description (in RNA
or DNA) of constitutive “cell” components was established. For a prebiotic system to become a true living system, however,
this self-referential description of the system had to be integrated (for example through signal transduction) into the other-referential
system of receptors at the surface of the cell. Such a stable integration of a self-referential digitally coded system into an other-referential analogically coded system
may perhaps be seen as a definition of life (Hoffmeyer, 1998).
10.3 Semiotic freedom
When a bacterial cell finds itself in a gradient of nutrients and swims right instead of left, the cell is making a choice.5 The choice is of course based on a complicated chemotactical machinery (comprising some scores of different protein species), but a biochemical analysis of the chemotactic system does not exhaust our need for understanding.
We also need to know why this apparatus was developed in the first place. Biochemistry essentially helps us to construct an
image of cellular life as chains or webs of chemical reactions taking place inside a cell or an organism. This of course is
extremely useful in many contexts, but in itself, it does not contain information about the structural logic that has ruled
the organization of this apparent mess of millions of chemical reactions taking place in a cell. As Nobel laureate and biochemist
Alfred Gilman once told Scientific American:
I could draw you a map of all the tens of thousands of components in a single-celled organism and put all the proper arrows connecting them (and even then) I or anybody else would look at that map and have absolutely no ability whatsoever to predict anything.(Gibbs, 2001, p. 53)
The key to cellular or organismic organization must be searched for in the historical (evolutionary) constraints on the interaction
of simple biosystems with each other and with their environments. In the semiotic understanding outlined above, the chemotactic
machinery serves to integrate the sensing of the outer world to the reality of the inner world as this reality is described
in the self-referential, or genetic, system. Natural selection, of course, has modulated this system all the way down from
the first cells on Earth; and for all we know, the system is very well safeguarded. The possibility remains, however, that
mutations will spoil or change it and, more importantly perhaps, that external factors may fool it. Researchers may, for instance,
easily fool the bacterial chemotactic system by adding nutrient analogs (such as artificial sweeteners instead of glucose)
to the medium; and most likely nature itself will from time to time “invent” comparable kinds of dupe.
In such cases, it seems appropriate to say that the cell misinterprets the chemical signs of its environment. Such misinterpretations are dangerous, and natural selection will favor any solution that helps the organism to better interpret the situations it
meets. Indeed, selection would be expected to favor the evolution of more sophisticated forms of “semiotic freedom” in the
sense of an increased capacity for responding to a variety of signs through the formation of (locally) “meaningful” interpretants.
Semiotic freedom (or interpretance) allows a system to “read” many sorts of “cue” in the surroundings, and this would normally
have beneficial effects on fitness. Thus, from the modest beginnings we saw in chemotactic bacteria the semiotic freedom of
organic systems would have tended to increase, and although it has not been easy to prove that any systematic increase in
complexity, as this concept has traditionally been defined, has in fact accompanied the evolutionary process, it is quite
obvious that semiotic complexity or freedom has indeed attained higher levels in later stages, advanced species of birds and
mammals in general being semiotically much more sophisticated than less advanced species (Hoffmeyer, 1996).
Allowing for semiotic freedom in the organic world significantly changes the task of explaining emergent evolution, because
semiotic freedom has a self-amplifying dynamic. Communicative patterns in assemblies of cells or individuals may often have
first appeared as a simple result of the trial-and-error process of normal interaction, and may then endure for considerable
periods of time. If such patterns are advantageous to the populations (cells or organisms), they may eventually become scaffolded
by later mutational events. Through this “semi-Baldwinian” mechanism, the evolutionary process will enter a formerly forbidden
area of goal-directedness (Hoffmeyer and Kull, 2003).
Biosemiotics presents a strong argument for an emergentist view of life. By semiotic emergence, I mean the establishment of
macro-entities or higher-level patterns through a situated exchange of signs between sub-components. The important point here
is that whereas the emergence of higher-level patterns may seem to be slightly mysterious (often raising suspicion of vitalism),
as long as only physical interactions between entities are considered, the same outcome becomes quite understandable when based on semiotic interactions among entities at the lower level. Most importantly, semiotic emergence in this sense may stand as a possible alternative candidate to natural selection as a mechanism for explaining the evolution
of purposive behavior.
The biosemiotic understanding also implies that semiosis cannot be used to mark off the human species from the rest of the
world’s creatures. Our species’ linguistic skills make us very different from other species on Earth, indeed, but as shown
in the work of Terrence Deacon, the capacity for semiotic reference is not in itself the distinctive mark between humans and
animals. What is distinctive is our unmatched talent for that particular kind of semiotic activity that is symbolic reference
(Deacon, 1997), and thus languaging. We share the semiotic capacity as such (that is, the iconic and indexical referencing) with all life
forms, and biosemiotics thus puts us back into nature in the same time as it reconstructs nature as a place for humans to
belong. Nature is much more like us than science – in its obedience to the anthropocentric taboo – has allowed. Instead of
the Cartesian either-or thinking, biosemiotics institutes a more-or-less thinking.
John Deely has called the human being “the semiotic animal” (Deely, 2007). Semiosis – sign action – takes place all over the life sphere, but only humans know the difference between signs and things; only humans are semiotic animals.
10.4 Semiotic emergence and downward causation
A key question in discussions of emergence concerns the ontological reality of causative influences upon lower-level entities
exerted by the macrostate of the system. The expression “downward causation” is often used in this type of causal relationship
in which a macrostate acts upon the very microstates of which it consists (Campbell, 1974). The term may be seen as an attempt to express parts of what used to be called “final causation” without linking ourselves
to the cultural inheritance of Aristotelianism and all the baggage this implies. As we shall see, semiotic emergence and downward causation
are two sides – or rather two aspects – of the same coin, which I would prefer to call “semiotic causation,” bringing about
effects through a process of interpretation.
Deborah Gordon’s laborious and highly rewarding work with ants of the species Pogonomyrmex barbatus (who live in a harsh zone bordering the deserts between Arizona and New Mexico) is illustrative here. Gordon’s work revealed
that the survival of colonies of this species is deeply dependent on the regulation of a sophisticated pattern of semiotic
interactions between individual ants, which then raises the question of whether the ant colony deserves to be seen as a “superorganism.”
(Gordon, 1995, 1999). Seen from a semiotic point of view, a superorganism might be understood as an assembly of organisms that collectively interacts
with its environments in a way that depends on a finely elaborated internal semiotic activity among the individual organisms
– a “proto-endosemiotics” (Hoffmeyer, 2008). Gordon found that a particularly important element in the colony’s behavior and growth process is what she calls “job allocation”
– and she shows that although this task does indeed rely on a quite schematic interaction pattern between different groups
of ants, an element of unpredictability persists:
An ant does not respond the same way every time to the same stimulus; nor do colonies. Some events influence the probabilities that certain ants will perform certain tasks, and this regularity leads to predictable tendencies rather than perfectly deterministic outcomes.(Gordon, 1999, p. 139)
Gordon’s experiments in this area may be seen as a response to experiments performed by the founder of sociobiology, Edward
O. Wilson, that were claimed to show a full-blown determinism in the response pattern of ants to chemical signals (for example
oleic acid (Wilson, 1975)). Gordon’s experiments, on the contrary, showed that “Just as the same word can have different meanings in different situations … so the same chemical cue can elicit different responses in different social situations” (Gordon, 1999, p. 97). Physiological, social, and ecological processes are simultaneously at work, says Gordon, and none of them is more
basic than the others: “Living levels of organization is central to any study of social behavior. For humans and other social
animals, an individual’s behavior is always embedded in a social world” (ibid., p. 96).
The semiotic competence of subunits, then – whether these subunits are human individuals in a society, plants in an ecosystem,
cells in a multicellular organism, or ants in an ant colony – is the medium through which the behavior and integrity of the
higher-level entity is produced and maintained. To the extent that such a system’s endosemiotic relations perceive and utilize
cues and signs that indicate (are indices for) the state of the holistic unit and its “needs,” it seems justified to talk
about these processes as genuinely endosemiotic – and consequently, the holistic system itself deserves to be ascribed a status as an autonomous unit: a superorganism.
The evolutionary formation of this kind of autonomous macro-entity is the quintessence of what is called “downward causation”
as defined above, and it is suggested that the connection between semiotic emergence and downward causation is taken as constitutive
for both phenomena, as shown in Figure 10.3.
The semiotic relations between subunits that collectively account for the stability of the large-scale or holistic system
(for instance the ant hill, the multicellular organism, or perhaps the symbiotic system of bobtail squids and light-emitting
Vibrio bacteria: see below) must necessarily be geared to respond to changes in the environment in ways that do not threaten the
integrity of the large-scale system. Sub-units, for their part, must receive messages telling them how to uphold the macro-system,
and the easiest way to do this is probably to distribute the needs of the macro-system via “indexical” signs.
An interesting example is the so-called “quorum sensing.” Quorum sensing has become the designation for a kind of
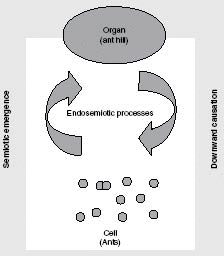
Fig. 10.3 The connection between semiotic emergence and “downward causation.” Downward causation operates through indexical sign relations;
that is, the values of system parameters are interpreted by lower-level agents as indexical signs. But this state of affairs
in itself presupposes the formation in the first place of a large-scale pattern with a behavior that stabilizes the semiotic
interaction between parts (from Hoffmeyer, 2008).
communicative activity in bacteria in which the density of bacteria present is a causal factor. In short, quorum sensing is
the result of a process in which each single bacterium excretes a certain chemical compound such that the concentration of
this compound in the medium will reflect the number of bacteria per unit volume. Quorum sensing occurs if the compound, after
having reached a threshold concentration, binds to a regulatory protein in the cell and thereby initiates the transcription
of specific genes. An illustrative case of quorum sensing occurs in a species of squid, Euprymna scolopes, which hunts small fish by night on the coral reefs off the coast of Hawaii.6 Moonlight causes the squid to cast a shadow that makes them an easy catch for predators. As a defense strategy, E. scolopes has evolved a sophisticated way of emitting light that effectively “hides” its own shadow. “Counter-illumination” is the
name given to this kind of camouflage, and it is only made possible by the squid’s symbiotic relationship with luminous bacteria
called Vibrio fischeri that live in the mantel cavity of the squid. Living on food provided by the digestive system of the squid, the bacteria emit
light of the exact same intensity and color as the light reaching the squid from the Moon, and this prevents predators from
seeing the squid from below (McFall-Ngai and Ruby, 1998).
In the morning, the squid bury themselves in the sand and excrete 90–95% of the bacteria, which brings the density of the
remaining bacteria well below the threshold level. The bacteria continue producing the particular substance, N-acyl homoserine lactone, which triggers the induction of light emission, but because the density of bacteria is low the concentration
of this substance remains too low to elicit any induction of the involved lux genes. Apparently, the squid is in full control
of the bacterial growth rate by adjusting the supply of oxygen, and at sunset the population of bacteria reaches the threshold
level once again. When night approaches, the squid makes sure that the bacterial density in its mantel cavity (and thus the
N-acyl homoserine lactone concentration) is high enough for the bacteria to respond by allowing for the transcription of lux
operon genes – and thus for light emission – to start. The point I want to make here is that the semiotic emergence whereby
this system was gradually established in the course of evolution necessarily also demanded the invention of semiotic means
for assuring the stability of the system, and these “semiotic means” are precisely what we understand by “downward causation.”
And here I have only touched upon the surface of the complexities of the communicative network operative in bringing this symbiotic interaction
between the squid and the bacterium to work.
Downward causation and semiotic emergence are thus two interwoven, but not identical, aspects of the same process. That this
mechanism, based on indexical semiosis, is indeed coupled to the circadian rhythms of the squid was confirmed by the finding
that the squid has means at its disposal to fine-tune light emission. It may, for instance, change the wavelength of the emitted
light by help of a “yellow filter,” and it may weaken the intensity of light by opening a bag of “ink” (Bruni, 2007). Presumably, the indexical sign process itself is too slow in its effects to compensate for momentary variations in background
light (as occurs under cloudy conditions), and evolution therefore had to provide the macro-system (the squid) with a number
of “additional screws” for fine-tuning.
10.5 Biosemiotics and God
Contrary to the genocentric neo-Darwinian orthodoxy, the view of animate nature as profuse with signs does not contradict
the fact that our Earth has supported the evolution of conscious human beings with moral feelings. Biosemiotics may potentially
account for the appearance of such creatures through immanent natural processes. The biosemiotic approach thereby overturns
the need for, or legitimacy of, the argument for intelligent design. Biosemiotics does not logically entail any stance on
the presence or absence of a transcendental creator, but it may be seen as rescuing the scientific world view from the need
for admitting such a transcendental power, a need that seems rather inescapable if one adheres to a traditional scientific
perspective. As Deacon and Sherman have put it:
In the debate between science and fundamentalist religion that the world’s citizens watch so attentively, the burden is on science to demonstrate how purposive processes can emerge in the absence of antecedent intelligence, carefully selected prior conditions, or intrinsically teleological components.(Deacon and Sherman, 2008, pp. 59–76)
Deacon and Sherman have made an impressive attempt at “tracking the emergence of the fledgling precursor to these intentional
relations,” but although their approach is perfectly naturalistic, its emphasis on theories of complexity and semiotics unfortunately
brings it well outside the narrow space of accepted causative agencies in mainstream scientific world view.
The present author remains agnostic towards the questions of a transcendent or immanent deity, but notes that such agnosticism
would have been hard or impossible to maintain, had it not been for the biosemiotic solution to the hard problems of natural
intentionality. For in the classical scientific image of the world as ruled by unbreakable natural laws, only a deity could
possibly have created such “unnatural” beings as you and me.
References
Bruni, L. E. (2007). Cellular semiotics and signal transduction. In Cellular Semiotics and Signal Transduction, ed. Barbieri M.. Dordrecht: Springer, 365–407.
Campbell, D. T. (1974). “Downward causation” in hierarchically organised biological systems. In Downward Causation, eds F. I. Ayala and T. Dobzhansky. Berkeley: University of California Press, 179–186.
Clayton, P., and Kauffman, S. (2006). On emergence, agency, and organization. Biology and Philosophy, 21: 501–521.
Deacon, T. (1997). The Symbolic Species. New York: Norton.
Deacon, T., and Sherman, J. (2008). The pattern which connects pleroma to creatura: The autocell bridge from physics to life. In A Legacy for Living Systems: Gregory Bateson as Precursor to Biosemiotics, ed. J. Hoffmeyer. Dordrecht: Springer, 59–76.
Deely, J. (2007). Intentionality and Semiotics: A Story of Mutual Fecundation. Scranton, PA: University of Scranton Press.
Depew, D. J., and Weber, B. H. (1995). Darwinism Evolving: Systems Dynamics and the Genealogy of Natural Selection. Cambridge, MA: Bradford/The MIT Press.
Gibbs, W. (2001). Cybernetic cells. Scientific American, August, 285: 52–57.
Gordon, D. M. (1995). The development of organization in an ant colony. American Scientist, 83: 50–57.
Gordon, D. (1999). Ants at Work. How an Insect Society is Organized. New York: The Free Press.
Gould, S. J. (2002). The Structure of Evolutionary Theory. Cambridge, MA: The Belknap Press of Harvard University Press.
Gray, R., Griffiths, P., and Oyama, S. (2001). Cycles of Contingency. Developmental Systems and Evolution. Cambridge, MA: MIT Press.
Griffiths, P. E., and Gray, R. D. (1994). Developmental systems and evolutionary explanation. Journal of Philosophy, XCI(6): 277–304.
Hoffmeyer, J. (1996). Signs of Meaning in the Universe. Bloomington, IN: Indiana University Press.
Hoffmeyer, J. (1998). Semiosis and biohistory: A reply. Semiotica, 120 (3/4): 455–482.
Hoffmeyer, J., and Kull, H. (2003). Baldwin and biosemiotics: What intelligence is for. In Evolution and Learning, eds B. Weber and D. J. Depew. Cambridge, MA: MIT Press, 253–272.
Hoffmeyer, J. (2008). Biosemiotics. An Examination into the Signs of Life and the Life of Signs. Scranton, PA: University of Scranton Press.
Kauffman, S. (1993). Origins of Order: Self-Organization and Selection in Evolution. New York: Oxford University Press.
Kauffman, S. A. (2000). Investigations. New York: Oxford University Press.
McFall-Ngai, M. J., and Ruby, E. G. (1998). Sepiolids and vibrios: When first they meet. BioScience, 48: 257–65.
Sarkar, S. (1996). Biological information: A skeptical look at some central dogmas of molecular biology. In The Philosophy and History of Molecular Biology: New Perspective, ed. S. Sarkar. Dordrecht: Kluwer, 187–231.
Scriver, C. R., and Waters, P. J. (1999). Monogenic traits are not simple. Trends in Genetics, 15(7): 267–272.
Ulanowicz, R. E. (2009). A Third Window: Natural Foundations for Life. Philadelphia: Templeton Foundation Press.
Wilson, E. O. (1975). Sociobiology. The New Synthesis. London: Belknap Press.
Information and the Nature of Reality: From Physics to Metaphysics, eds. Paul Davies and Niels Henrik Gregersen. Published by Cambridge University Press © P. Davies and N. Gregersen 2010.
1 One promising candidate for an information concept that is both rigorous and biologically useful has been developed in ecology
as the so-called “average mutual information,” a measure for how well organized or determinate a configuration of ecological
relationships appears. Theoretical ecologist Robert Ulanowicz has developed this concept further to a concept of ascendency that represents: “the coherent power a system could bring to bear in ordering itself and the world around it” (Ulanowicz,
2009).
2 Semiosis = sign activity. Semiotics = the sciences studying sign activity.
3 But indeed, inside a theater the smoke might just be faked. Semiotics, as Umberto Eco once said, is the science of lying.
4 In the simplest case, the intentional dynamics of aboutness shows itself as fertility.
5 Again I am here using teleological language. The idea is not, of course, that the bacterium makes a conscious choice, but
only that it systematically favors a distinct response out of several equally possible behaviors. If it is objected that this
“preference” is not a real “preference” or “choice” because the bacterium could not possibly have failed to respond the way
it did, I will contend that: (1) considered as an evolving species the bacterium does in fact have a choice, in the sense
that it might have evolved differently; and (2) even single bacterial cells are complex systems that exhibit truly unpredictable
(chaotic) behavior and, in fact, mutant cells might behave differently in the same situation. It should be noticed here that
the idea of an original “wild-type” bacterium has now been given up: all bacterial cells are thus “mutants” or “normal,” depending
on how the context is defined.