7Reconstruction Principles: Facial Tissues
‘ Undoubtedly the eyes of any living creature indicate to a greater or lesser extent the inner feelings and emotions by which the bodily actions are controlled’.
Previous chapters have focused on attributes of tissue reconstruction which affect multiple body components. In this chapter, we will zero in on the anatomy of ancient animal faces (Fig. 7.1). These aspects should not be considered divorced from the points and observations made in preceding chapters (such as methods of muscle reconstruction, predicting integument types and so on) but reconstructing facial anatomy presents enough unique considerations that it warrants dedicated discussion.
Fig. 7.1 The face of the dome-headed dinosaur Pachycephalosaurus wyomingensis, restored based on epidermal correlates. Animals with armoured or otherwise reinforced facial tissues lend themselves well to such restorations because tough skin greatly influences the surface texture of animal skulls. (M. Witton)
Sclerotic rings and eyes
Eyes are located in sockets known as orbits and, though most eye tissues do not survive as fossils, we can make some useful observations on modern animals to guide our restorations. Firstly, eyeballs are rarely completely flush with the margins of the skull, instead bulging somewhat from their sockets and being held in place by soft tissue. This is particularly so with small animals: close inspection of living individuals will reveal eyes which protrude from the sides of the head, often further than the skin and integument. The position of the orbits has an important role in dictating which way eyes are directed. For many tetrapods, orbits and eyes face laterally but others – particularly predators and climbers – have forward-facing eyes that provide overlapping visual fields (sometimes called ‘binocular’ vision). Many semi-aquatic animals have orbits which face dorsally, allowing their eyes to project somewhat above their skulls. Similar orbital configurations are present in fossil semi-aquatic animals.
Pupil shape is varied among modern animals and probably differed in the past too. Living species have converged on similar pupil shapes multiple times and recent studies show strong correlation between pupil shape, ecological niche and animal size (Banks et al. 2015). Ergo, if we understand these aspects of fossil animals, we can make a reasonable inference of their pupil shape. Round pupils are good ‘generalist’ morphologies and might be considered a safe ‘default’ assumption for any fossil vertebrate. Vertical slit pupils are a common alternative, but the idea that these only occur in animals which are adapted for nocturnal activity is not entirely accurate. Slit pupils allow for great control of light entering the eye and shading of sensitive eyes from bright lights, but ecology and behaviour have an important, sometimes dominant influence on pupil shape too. Vertical slit pupils are most commonly correlated to predators adapted for ambushing prey, as they permit greater capacity to judge distances and gauge depth of field without cranial motion – an obvious boon to a predator hoping to remain undetected. Such pupils are especially common in animals with forward-facing eyes as well as smaller predators, because detecting depth of field via slit pupils is more effective at reduced angles to the ground (for instance, in shorter animals). Ergo, taller predators receive less benefit from vertical slit pupils, and tend to have rounder ones (Banks et al. 2015). We might assume that this phenomenon also explains why almost all birds and bats have round pupils – perhaps most extinct fliers had round pupils, too. Horizontally-expanded pupils are generally associated with laterally-facing eyes and permit panoramic vision while providing shade against sunlight. They are suited to grazing prey species that need to be aware of predators. Artists should note that horizontally elongated pupils can remain aligned to the horizon even as the head moves – they literally rotate in their sockets to stay level as the head moves up and down (Banks et al. 2015). This fact is worth considering when drawing extremely-posed animals with horizontal pupils.
Fig. 7.2 Restoring the eyes of reptiles. (A) Skull and mandible of Eurhinosaurus longirostris showing large orbit and sclerotic ring (after McGowan and Motani 2003). (B) Diagrammatic cross-section of a reptile eye: note that the sclerotic ring is buried under soft tissue, and is not visible on the living animal. (C) Restoration of E. longirostris with erroneous eye proportions, where the visible eye fills the entire orbit. (D) Restoration of E. longirostris with correctly restored eye proportions, dictated by the inner margin of the sclerotic ring.
Rings of bony plates known as sclerotic rings support the eyeballs of many animals (they are absent in living mammals and crocodylians), and their extent provides a minimal eyeball size. In some animals, like ichthyosaurs and raptorial birds, sclerotic rings can be huge and dominate the entire eye socket (Fig. 7.2), while in others they are small and dwarfed by the orbital housing. In these instances, and assuming the ring has not moved, the position of the sclerotic ring gives an indication of where the eye sat in the orbit: it is not always the centre of the eye socket. Sclerotic rings extend across the front of the eyeball and their size provides a minimum dimension for the eyeball tissues. This does not correspond to the maximum size of the visible eye, however. Rather, the bony part of the ring is covered by facial tissues, and the iris and pupils are only discernible through the opening at the centre. It is very common for artists to ignore sclerotic bones when restoring animals like ichthyosaurs, restoring the entire orbit as filled with a gigantic eye. Sclerotic ring proportions suggest that their visible eye tissues were much smaller than the orbit dimensions however: their giant eyeballs were only fractionally visible in life.
(C) Giraffatitan brancai. (D) Darwinopterus robustodens. (E) Stagonolepis robertsoni.
Nostrils, proboscides and trunks
The nasal openings of animal skulls – the nares – are morphologically variable. In their simplest form they are small, paired openings into the snout, and at their most complex they are intricately structured, cavernous openings into the skull. They can house a variety of nostril types and nasal tissues: simple, immobile openings, sealable apertures, large cartilaginous structures as well as dextrous proboscides and trunks. Accurately predicting the configuration of nasal tissues is thus an important part of restoring an animal’s face.
Fig. 7.3 Nostril position in fossil reptiles following Witmer (2001), which predicts that nostrils are typically placed at the front and base of the nasal opening. (A) Tyrannosaurus rex. (B) Diabloceratops eatoni.
Our first concern is the placement of nostrils (Fig. 7.3). Often, particularly in large nosed species like sauropods or pterosaurs, artists seemingly place nostrils arbitrarily, perhaps guided by what intuitively looks best or an assumption about functional performance. Research into living reptiles shows that the nostrils of most lizards, crocodylians and birds are placed in the anterior region of the nasal cavity, even when the nasal opening itself is large (Witmer 2001). There are exceptions, such as certain monitor lizards and marine birds, which have dorsally or posteriorly displaced nostrils to avoid influxes of water (in aquatic species) or debris (in terrestrially foraging species), but these are secondarily-developed specializations of the nasal tissues. It seems that nostrils are also placed at the tip of osseous nasal openings in mammals too, or at the tip of nasal soft tissues in species with augmented nose anatomy. Thus, unless we have good reason to do so, we should place the nostrils of fossil animals in the anteriormost region of the nasal tissues. This seemingly minor point can have a surprising impact on the appearance of some fossil species, moving the nostrils from one side of the skull to another, or implying the presence of additional soft tissues to house the nostril in its correct place (Witmer 2001).
We can also consider that nostrils are not always simple oval-shaped openings. Their shape often depends on ecology: many ground-feeding birds have flaps of skin covering most of the nostril to keep out debris, and burrowing species may have small nostrils for the same reason. Diving or aquatic species have narrowed, sealable, or completely closed nostrils that prevent water entry. Thus, if something of the palaeoecology of a fossil reptile is known, we can restore it with appropriate nostril form. Most animals have a nasal septum, a piece of cartilage which divides the nasal chamber into left and right divisions and prevents light penetrating the rostrum through the nostrils. Nasal septa are delicate structures and rarely preserve in fossils, but evidence of their presence occurs in well-preserved skulls (Frey et al. 2003; Bourke et al. 2014): they were probably as common in extinct animals as they are today. An alternative to this morphology are the pervious nostrils of certain birds (Fig. 7.4), which lack a septum between the nostril openings. Unfortunately, there seems to be little correlation between lifestyle and pervious nostrils, so it is difficult to know when to apply this to fossil animals unless the septal anatomy can be determined through internal examination of the nasal cavity.
Many nares are small, paired openings at the end of the snout and present few opportunities for erroneous interpretation, but several lineages present much larger nasal openings that require more consideration. This especially applies to mammals where the nares are vast and filled with muscular and cartilaginous tissues. Extra complications arise when these tissues are linked to the lip and cheek anatomy, creating mobile or even prehensile nasal anatomy. Fleshy mammalian nasal tissues are predictable through the development of a large, conjoined and anteriorly-open nasal opening and a large infraorbital opening – this is a perforation seen below the orbit in mammal-line animals which passes blood vessels and nervous tissue to fleshy lips, noses and cheeks. This configuration occurs in the cynodonts, extinct close relatives of true mammals, but is not seen in more rootward members of the mammal line. Presumably, these non-cynodont species had more reptile-like nasal tissues.

Fig. 7.5 Reconstructing proboscides and trunks in fossil mammals: proboscis features are demonstrated for Paraceratherium transouralicum (top left) and Deinotherium giganteum (right); trunks are demonstrated on an African elephant (Loxodonta africana).
Mobile combinations of nasal, cheek and labial tissue – trunks and proboscides – are muscular, complex organs which have a suite of osteological correlates, allowing artists to predict their presence and some aspects of their basic shape (Fig. 7.5). We can infer proboscides through the combination of retracted nasal openings; the presence of large muscle scars, bony knobs and other muscle attachment markers around the nasal opening (particularly in the dorsal region); retraction of the nasal bone (the ‘roof’ of the nasal opening); deepening of the premaxillary bone; anterior migration of the orbit; a large infraorbital opening; and strengthening of the posterior skull regions related to supporting the weight of the head (Wall 1980). The criteria for elephant-like trunks are similar, but they require further modification to the skull. The front of the skull is shortened and sheer, with particularly complex knobs and sculpting above the large nasal opening for trunk muscle attachment, and the infraorbital opening is especially large (Markov et al. 2001). With these criteria, we can see that many fossil mammals likely bore elaborate nasal tissues that may not be evident in their modern relatives. For instance, the giant indricotheres – enormous relatives of rhinoceros – almost certainly sported a tapir-like proboscis in life (Fig. 7.6), and many other fossil rhino-line mammals may have been similarly equipped.
Fig. 7.6 Reconstruction of the giant indricotherine Paraceratherium transouralicum with a short proboscis, a feature strongly indicated by its skull anatomy (see Fig. 7.5) (M. Witton).
We also see that some fossil animals must have had proboscis morphologies unlike anything around today. Deinotherids – large relatives of elephants, famous for the downturned tusks on their lower jaws – have skull morphologies consistent with the presence of a proboscis (Markov et al. 2000), but the configuration of the skull implies a unique morphology. The nasal region, which anchors dorsal aspects of the proboscis (for example, levator nasalis and levitor labii maxillaris muscles – see Goldfinger 2004) is broad but relatively shallow, suggesting the dorsal muscles of the proboscis were wide but low. Conversely, the region under the nares is deep and complex, suggesting the lower proboscis component – derived from the cheek (buccinator) muscles – was much deeper and perhaps better developed. Put together, these anatomies suggest a proboscis with a flat top but wide, deep underside: not what we see in living elephants or any other living animal. Looking closely at fossil mammal skulls allows us to reconstruct nasal tissues appropriate to their skulls, and not just substitute nasal tissues from their modern relatives.
Fig. 7.7 The surprising extent of nasal tissues in reptiles. (A) Water monitor (Varanus salvator) skull with large, dorsally-facing ‘basins’ as narial openings. (B–C) Cross-sections of savannah monitor (Varanus exanthematicus) snout showing extent of soft tissue occupying nasal region (B). Compared to how we might reconstruct the same region in palaeoart (C). (D–F) Skulls and soft tissues of living species in lateral profile showing impact of nasal tissues on face shape (D) Savannah monitor. (E) Dromedary (Camelus dromedarius). (F) Giraffe (Giraffa camelopardalis). (G) How the cranial profile of Giraffatitan brancai is altered by varying extents of nasal tissue. (H–I) As for (G), with Stagonolepis robertsoni.
Fig. 7.8 Aetosaur Stagonolepis robertsoni restored with generous quantities of nasal cartilage. (M. Witton)
Somewhat surprisingly, it is not only mammals that have enormous and complex nasal skeletons. Many fossil reptiles – examples include pterodactyloid pterosaurs, sauropod, ceratopsid and hadrosaurid dinosaurs, aetosaurs, and several others – also have huge nares, often with complex underlying anatomy (Fig. 7.3). Exactly what these enormous narial openings signify is not clear. There are strong indications that some were lined with complex cartilaginous passages and chambers involved in lightening their skulls, olfaction and heat exchange, but we have not gleaned enough details of their structure to infer their actual life appearance. Traditionally, palaeoartists have simply made reptile nostril tissues confluent with the surrounding skull structure, an approach validated by living birds with cavernous nasal skull openings. But another reptile group, the varanid (or monitor) lizards, show a rather different anatomy (Fig. 7.7). These lizards have proportionally large nares which occupy the entire length of their snouts, and these act as shallow trays for large cartilaginous chambers – ‘nasal capsules’ – that visibly enhance the height and width beyond the margins of the underlying skull. Indeed, it’s these structures which produce the characteristic bugles and ‘boxy’ appearances of monitor lizard snouts, not their bony anatomy. In this respect varanids are not dissimilar to mammals, where – albeit with differently shaped noses – we see a similar correlation between large nasal openings and a reliance on cartilage for nasal form and shape. These modern species suggest our traditional approach of simply ‘smoothing over’ extensive nares may not always be appropriate and we might consider giving some species with cavernous nasal openings fuller, more prominent and maybe even bulbous nasal tissues (Fig. 7.8).
‘Extra-oral tissues’ and tooth exposure
It is something of a trope that prehistoric animals must bare their teeth in palaeoart, even when their mouths are closed. This is a relatively recent invention, however. Historically, the majority of palaeoartists covered the teeth of their subjects with extra-oral tissues (lips, cheeks and so on – Fig. 7.9), and only select species – sabre-toothed carnivorans or mammoths – were depicted with exposed tusks or sabre-canines. Nowadays, whether animal teeth should be exposed or covered in palaeoart is the subject of some debate, and one made all the more frustrating by complications of correlating skeletal anatomy to – the facial tissues which cover the mouth.
Several approaches have been used to investigate tooth coverage in fossil animals, but all have issues which preclude their use unreservedly. One method involves measuring how far teeth overlap jaw skeletons when the mouth is tightly shut (Ford 1997). These efforts have not accounted for post-mortem tooth slippage however (Chapter 4), not calibrated the impact that jaw soft tissues have on mouth closure, or noted that x-rays and CT scans of living animals show that jaws often are only loosely closed even when mouths look, externally, tightly shut. Without full knowledge of oral tissues, we may not be able to tell exactly when fossil animals considered their mouths shut, and thus how much their teeth protruded over the jaw skeleton. These models need a stronger experimental basis to contribute meaningful data to this discussion.
Reisz et al. (2016) have argued that teeth can only be exposed in semi-aquatic species, as aquatic settings prohibit the enamel tooth component from desiccating and becoming damaged. This model assumes that exposed teeth among terrestrial species lack enamel covers, which would give artists a simple way to predict tooth coverage: enamel teeth are covered, enamel-less teeth (for instance, those with exposed dentine) may not be. This idea is complicated by two factors, however. The first is that the idea of only enamel-less teeth being exposed in living animals is untrue (Hilson 2005). While some exposed teeth lack significant amounts of enamel (such as the tusks of elephants), there are many fossil and living species that do have substantial enamel components on their exposed teeth. For example, the upper tusks of gomphotheres (fossil elephant relatives) have broad bands of enamel along their lateral surfaces (Padro and Alberdi 2008), and the canines of male muntjac deer are largely covered with enamel, save for the medial surface (Hilson 2005). The tusks of male wild boars and warthogs only bear dentine on the posterior surface and wear facet, with enamel covering the other surfaces. To confuse things further, walruses have mostly dentine tusks, despite their aquatic habits negating the purported desiccation risk, while the spiralling tusks of another marine mammal, the narwhal, are entirely covered in enamel (Hilson 2005). If there is a relationship between enamel, desiccation and tooth exposure, it is not immediately obvious.
A further reason to be sceptical of the enamel desiccation hypothesis is that many crocodylians – some of the only living animals with numerous exposed teeth – aestivate for months at a time in dry underground burrows without access to water (Grigg and Kirshner 2015). This either implies that the idea of dental desiccation is incorrect, or that the requirements for dental hydration are so relaxed – literally months can pass without getting the teeth wet – that it has little influence on facial anatomy. Work in this area is still ongoing as enamel microstructure is varied across vertebrates and its properties are poorly understood. However, an inescapable fact seems to be that enamelled teeth can be exposed, and that the proposed relationship between enamelled teeth and extra-oral tissues is complex, if it even exists.
Fig. 7.10 The relationship between jaw bone foramina and extra-oral tissues in living animals. A face value reading of this intriguing data suggests correlation between foramina counts and the likelihood of pliable jaw tissues, though further research is needed to test this link (from Morhardt 2009, reproduced with permission).
A somewhat more promising line of research involves labial foramina, small openings on jaw bones adjacent to the teeth that allow passage of nerves and blood vessels between the skull and jaw soft tissues. Some (Knoll et al. 2008) have found little correlation between labial foramina and soft tissue morphology, as their number and position can vary among species with similar jaw tissues, between individuals of the same species, and even between the left and right jaws of an individual. Morhardt (2009), however, found that there may be a coarse link between foramina count and tooth coverage, where the likelihood of tooth exposure increases the number of foramina (Fig. 7.10). Whereas animals with very few jaw foramina have lips or cheeks covering their teeth, animals with high foramina counts – 100 or more in each jaw bone (premaxilla, maxilla and dentary – see Fig. 4.5) – generally have exposed teeth. Morhardt’s proposal is based on studies of a wide variety of extant animals and remains one of the more promising investigative pathways on this topic, but is not without caveats. For instance, some of the foramina counted in this study may have been related to specialized facial sensory tissues, not extra-oral tissues, as is certainly the case for crocodylians and sea lions (Morhardt 2009). This data has also yet to be factored against differences in nervous and vascular pathways in animal skulls or be subjected to phylogenetic correction (Morhardt, personal communication 2017). For the time being, this study remains an intriguing insight into animal life appearance that artists should consider for their work, but it also remains a work in progress.
While we wait for a more definitive mechanism to detect extra-oral tissues, we can consider some basic observations of living animal faces that have bearing on the likelihood of tooth exposure in fossil species. Firstly, a simple tally of species with completely covered teeth against those with exposed teeth shows that our convention of maximizing tooth exposure in palaeoart is unusual. The overwhelming majority of living animal species have covered teeth, including all amphibians, most mammals and most reptiles. Crocodylians and some species of freshwater dolphin are the only living tetrapods with mostly or fully exposed dentition, while lips – either mere margins of skin, or skin overlying muscle – are so universal among tetrapods that we should assume they were ancestral to Tetrapoda itself, and that exposed teeth are a derived condition limited to select species. With living animals being our best insight into their fossil relatives, this observation suggests covered teeth should be our ‘default’ assumption for extinct animal appearance and that, instead of asking ‘does this animal have extra-oral tissues?’, we should ask ‘why doesn’t this animal have extra-oral tissues?’.
We can further observe that, as a general rule, teeth employed in processing food are covered. Mammal tusks and the exposed canines of certain deer are not directly involved in food processing, although they are not non-functional overall (for instance, elephants use their tusks to break branches, dig, topple trees; deer fight with their large canines). It seems that only teeth of extreme size relative to the rest of the dentition are likely to escape oral margins, though they can remain covered even when their tips extend to the dorsal or ventral limits of the jaw skeleton, so long as they are aligned more or less vertically within the jaw. Chances of tooth exposure are boosted when teeth grow obliquely or directly away from the jawline.
Fig. 7.11 The relationship between extra-oral coverings and tooth size in living and fossil animals. Blue lines shows extent of lip tissues. (A-C) Extinct species typically restored with exposed dentition. (A) Gorgonopsian Inostrancevia alexandri. (B) Tyrannosaurus rex. (C) Smilodon fatalis. (D-K) Large-toothed living animals and the shapes of their lips. (D) Lion (Panthera leo). (E) Mandrill (Mandrillus sphinx). (F) Hippopotamus (Hippopotamus amphibius). (G) Crocodile monitor (Varanus salvadorii). (H) Tuatara (Sphenodon punctatus). (I) Asian elephant (Elephas maximus); (J) muntjac deer (Muntiacus reevesi). (K) Common warthog (Phacochoerus africanus). Only I–K have exposed teeth.
Fig. 7.12 Black water monitors (Varanus salvator komaini) are equipped with substantial dentition (see Fig. 7.7A) but this is not obvious in living individuals, even those with open mouths. The same might have been true for ancient animals.
But although large tooth size seems to correlate with tooth exposure, the threshold is much higher than we intuitively assume (Fig. 7.11). Animal jaw tissues can cover surprisingly large teeth, sheathing large fangs and tusks in species that look – from the outside – to have unremarkable dentition. Rhinoceroses, sloths, tapirs, numerous primates (especially the papionines – geladas, baboons and so on) camels, tuataras, snakes, peccaries, bullfrogs, hippopotamuses, monitor lizards, leopards, numerous rodents and many other living species have surprisingly ferocious teeth which are hidden without even a bulge in facial tissues or change in lip direction. Indeed, in some cases the large teeth remain hard to spot even in open mouths thanks to lips, cheeks, gums and tongues overwhelming them in soft tissue (Fig. 7.12). Even a simple lip can hide large teeth through the aid of sheaths and pockets, or simply by being large. Amphibians, lizards and many mammals have upper and lower lips of similar size which meet over the teeth so that sheaths can form on either jaw, but some mammals – including most carnivorous forms – have very large, fleshy upper lips over thinner, tightly-bound soft tissues of the lower jaw. In these species, the canine teeth overbite the lower lip but the tissues of the upper jaw are large enough to obscure them (Fig. 7.9).
Fig. 7.13 Gorgonopsian Inostrancevia alexandri is typically restored with exposed canine teeth, but its dentition is within the size range of covered teeth among living species. (M. Witton)
Assumptions that large or conspicuous teeth in fossil animals should be exposed are thus directly challenged by modern animals. We should not underestimate the capacity for facial tissues to obscure even very large, sharp or ferocious-looking teeth, and should note that the dentitions of fossil animals frequently shown with exposed teeth – particularly theropod dinosaurs, gorgonopsians and other carnivorous stem-mammals – are relatively no larger than those enclosed inside the oral tissues of living animals (especially once the problem of tooth slippage is corrected). Frankly, for most species, it is hard to justify why their teeth should not be covered (Fig. 7.13). It’s only under specific circumstances that we should seriously consider rendering exposed teeth.
The first is that especially long teeth, which project a considerable distance from the margins of the skull and lower jaw, are good candidates for permanent exposure. Examples might include the canines of sabre-toothed mammalian carnivores (such as Smilodon and other machairodont felids), the tusks of fossil elephants and their relatives, and the larger tusks of dicynodonts. We should also note those fossil reptiles – such as certain Crocodyliformes, pterosaurs and marine reptiles – where entire toothrows are composed of dentition so long that their tips extend well beyond the jaw skeleton. Such extensive dental apparatus would seem to preclude the development of any sheathing soft tissues (or, at least, any sheaths akin to those exhibited by modern animals).
A second justification for tooth exposure might be the nature of the facial tissues. For instance, if there is evidence that the subject species has taut facial skin even at the jawline (such as in modern crocodilians – see Chapter 8), there is an increased chance that its teeth were exposed. We might predict tight skin coverings using a combination of high foramina counts in the tooth-bearing jaw bones (Morhardt 2009), rugose and sculpted bone textures (particularly those associated with epidermal scales – see Chapter 6), and the presence of robust jaw bones with small or absent openings (Chapter 8). Such characteristics are seen on the skulls of many fossil Crocodyliformes and ‘amphibians’, and they might imply that, like their living descendants, they likely had exposed teeth.
Phylogenetic bracketing gives a third consideration for rendering exposed teeth. For example, the ‘over-lip’ configurations of living mammalian carnivores (Fig. 7.9) are common enough in the group to assume this was their ancestral condition, and this must be our default assumption for the lip anatomy in all their fossil relatives. This can feed into specific predications of tooth exposure in carnivorans with particularly impressive teeth: reconstructions of machairodont cats like Smilodon with perpetually bared fangs are reasonable because we know carnivorans generally lack extensive lower lip tissues and, unless Smilodon and kin had lip anatomy completely unlike their modern relatives, their canine teeth must have been exposed.
Fig. 7.14 Tooth exposure in the Tasmanian devil (Sarcophilus harrisii). Despite having every correct condition to predict fully sheathed dentition, devil canines are exposed perpetually (A) and much of the dentition is visible when the mouth is opened (B). Examples such as these undermine any hard rules concerning extra-oral tissue reconstruction.
It must be stressed that these observations are guidelines, not hard rules, and several modern species defy these attempts to characterize extra-oral tissues and dental sheathing. For example, restoring sabre-toothed cats with exposed teeth is complicated by one living cat species – the clouded leopard – which sheaths a long set of canines in its lower jaw. We would not predict this based on other cat species and, if known only from fossils, clouded leopards would probably be restored with exposed canine teeth. Likewise, Tasmanian devils have exposed upper canines despite their relatively modest length and, if only known from fossils, there would be no reason to consider them unsheathed (Fig. 7.14). These examples remind us that palaeoart is ultimately a game of prediction and probability, attempting to restore what is likely and defensible rather than what looks best, matches our gut feelings, or is our favourite interpretation.
Types of extra-oral tissues: lips, cheeks and rictal tissues
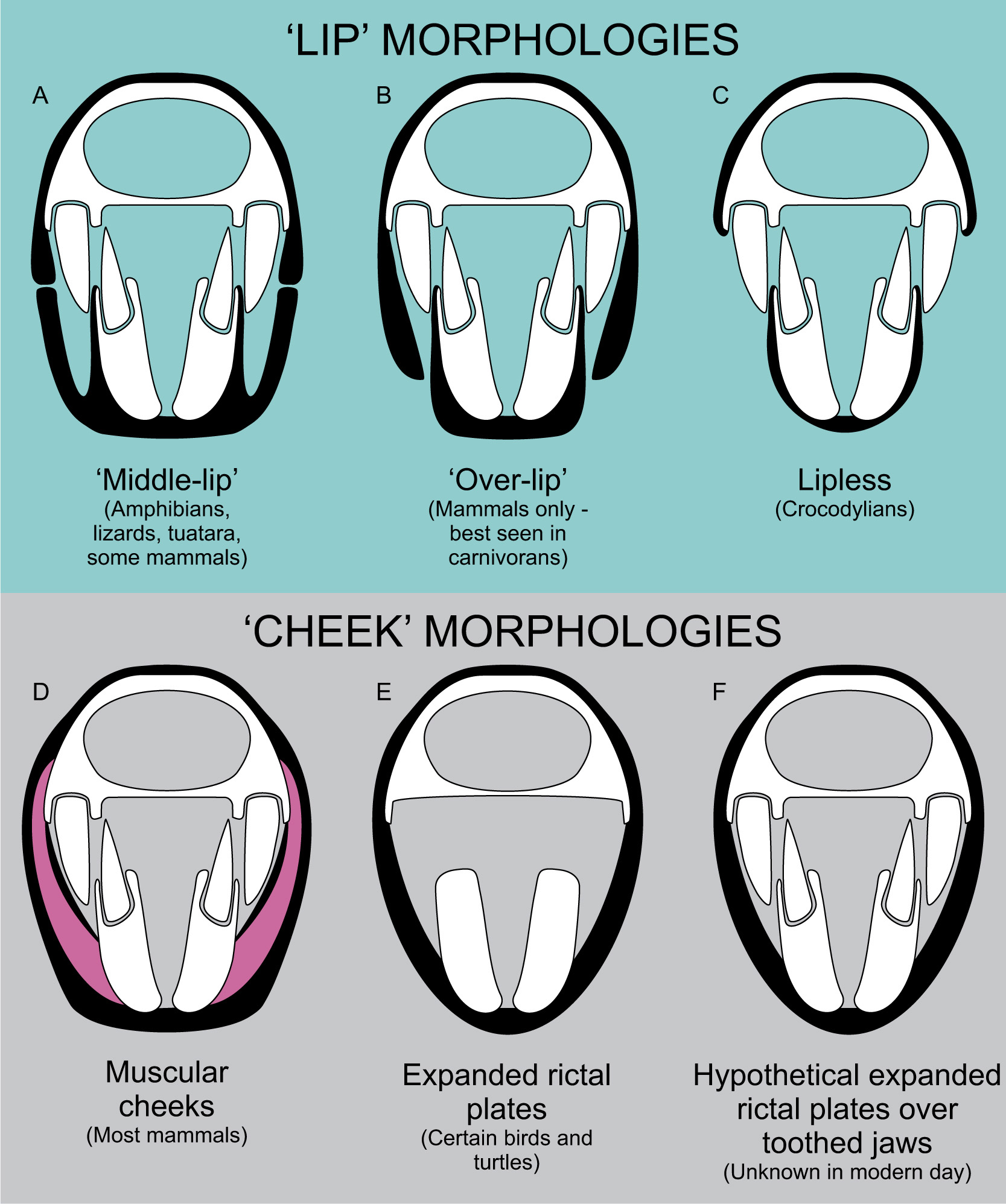
Broadly speaking, extra-oral tissues are divided into two groups: lips – sets of tissues which extend along the jaws to seal the mouth when closed, and cheeks – tissues attached to both jaws which stretch when the mouth is opened, covering the jaws regardless of mouth posture. We must be careful when discussing these tissues as lips and ‘cheeks’ in reptiles and amphibians are structurally different to those of mammals. This has implications for life appearance, so the right extra-oral tissues must be applied to our subjects to ensure credible restoration.
The most basic form of lip, common in reptiles and amphibians, is a ridge of tissue that grows around the jaw margins. A similar condition, though independently developed, occurs in whales and dolphins. Labial foramina mark the base of the lip, allowing us to predict deep or large-lipped animals versus relatively shallow-lipped species. Generally speaking, we do not see osteological correlates for epidermal tissues between the labial foramina and toothrow as the skin is separated from the bone in these regions. These tissues are not muscled so they cannot be pulled into expressive poses: lizards and amphibians cannot snarl like an angry dog. It can be surprising how much of the dentition can be obscured by these lips: the large fangs of butterfly lizards, for instance, are only slightly visible below their lip margins, even in individuals with wide-open mouths. This form of lip should be considered the starting point for all tetrapod reconstructions, and only not applied when we have good indications of liplessness, or reasons to assume more derived lip anatomy. Most fossil reptiles and many non-mammalian synapsids probably had lips of this kind.
Mammalian lips have a muscular component that makes them malleable. The principle muscle of the lip is the orbicularis labia, a complex of muscles that form a ring around the entire mouth. Mammalian lips are further distinguished from those of reptiles in being intrinsically involved with the muscles of the cheek, creating a very dynamic set of oral tissues. The configuration of mammal lips varies. Some species have fairly symmetrical upper and lower lips, while others have large, fleshy top lips which overhang reduced lower lips. Many living mammals have large cheek components that displace the lips to the front of the mouth, while others have reduced cheeks and long lips which expose a lot of the oral structure when opened (Fig. 7.14). The size and muscular nature of lips can be hard to predict from fossils, but is aided by phylogenetic bracketing among crown-group mammals. In stem-mammals, the development of the infraorbital opening versus a reduction in labial foramina is probably a good correlate for the development of muscular lip anatomy.
Beyond lips, we have ‘cheeks’. We have to be careful in our use of this term as all vertebrates have structures stretching between their jaws that cover at least some of the oral cavity, but not all animals have ‘cheeks’. True ‘cheeks’ are best considered a mammalian feature: a complex of fleshy, muscular and elastic tissue between the posterior jaw and lips, formed primarily by the buccinator muscle, and overlain by fatty tissues and other muscles. The infraorbital foramen also supplies nutrients and sensitivity to cheeks, so this structure is probably a good correlate for fleshy cheeks as well as muscular lips. So far as we can tell, this complicated set of tissues has never developed outside of mammals and our close ancestors.
Fig. 7.15 Rictal plates, or ‘pseudocheeks’. (A) Alligator snapping turtle (Macrochelys temminckii) demonstrating a large rictal plate. (B) Placodont Henodus chelyops restored with a rictal plate, a soft tissue which would be functionally advantageous to this filter-feeding reptile.
However, this is not to say that cheek-like structures are unique to mammals. The jaws of vertebrates are filled at least partly by their own jaw muscles and, when these are large (as they are in crocodylians, for instance), they can add an expanse of stretchy tissue of cheek-like appearance to the jaw. All animals also have a rictus (sometimes termed ‘rictal plate’), a piece of highly vascularized and sensitive skin at the junction between upper and lower jaw tissues. In most species this is only a tiny web of flesh, but the rictus is large and sheet-like in some birds and turtles (good examples include condors, flamingos and alligator snapping turtles), extending some way along the jaw to form a cheek-like structure which terminates at the base of their beak tissues (Fig. 7.15). It is possible that many fossil animals had these expanded rictal plates or ‘pseudocheeks’, but there is no known correlate for such structures. They do not seem to have any relationship to labial foramina (Knoll et al. 2008; Morhardt 2009) or obvious attachment sites. It has been assumed that medially-offset toothrows are a correlate for a cheek-like structure in dinosaurs (Galton 1973), it being reasoned that the teeth have been moved inwards to allow room for soft tissues at the side of the face. This is demonstrably false however, as many lizards have medially-displaced toothrows while mammals show no medial dental displacement. Perhaps the only possible indicator of expanded rictal plates are osteoderms, which can be found overlying the jaws of heavily armoured animals, such as ankylosaurid dinosaurs (for example, Panoplosaurus). The assumption that these structures sat in a pseudocheek is not unreasonable, but remember that osteoderms underlie scales, and scales cover the lips of virtually all living reptiles; outlandish as it might seem, these osteoderms may simply represent enormous lip scales. And if armoured lips seem ridiculous, consider that some ankylosaurs possessed bony, armoured eyelids – if any clade were to develop bony lips, it would be these dinosaurs.
The addition of pseudocheeks to fossil animals is thus currently a game of speculation and educated inference, although it is not a foolish one: expanded rictal plates are common enough in modern reptiles and birds to assume they may have cropped up multiple times in evolutionary history. We do not have any living species where large rictal plates cover toothed jaws, but there is no reason to think these anatomies are mutually exclusive. It is not unreasonable to use functional interpretations to aid inference of pseudocheeks. Species which chewed food repetitively and continuously – such as hadrosaurid and ceratopsid dinosaurs – may have found that pseudocheeks contained food in their mouths. Aquatic animals that employed suction feeding – which might include some extinct marine reptiles and mammals – may have benefited from large rictal plates, as they enhance sucking pressure gradients between the mouth and surrounding water (Lemell et al. 2010). Animals that did not use these strategies may have benefited little from pseudocheeks, and might have enjoyed greater utility from the increased gape and food gathering potential of a cheekless mouth.
Toothless jaws and beaks
Beaks are composed of hard-wearing cornified sheaths that grow around animal jaws, often in replacement of teeth. They are not common to all toothless species (some turtles, for instance, lack them) but have appeared numerous times in animal evolution. As noted in the previous chapter, cornified sheaths have a strong relationship with closely underlying bone and this enables palaeoartists to predict their distribution without the help of soft tissue preservation (Fig. 7.16). This includes the development of compound beak tissues – where beaks are composed of multiple sheets – versus entirely continuous beak structures (Chapter 6). Among tetrapods, beaks seem to be the sole remit of reptile-line animals, the ‘beaks’ of mammals like platypuses being soft, spongy tissue which only superficially resembles the true beaks of birds, turtles and their many extinct relatives.
Fig. 7.16 Restoring beaks. (A) Skull of a fulmar (Fulmarus glacialis) with compound beak tissues labelled. (B) Interpretation of the beak tissues in the Cretaceous bird Hesperornis regalis, in which the toothed regions lack beak tissues (based on Hieronymus and Witmer 2010). (C) Osteological correlates for cornified sheaths on the lower jaw of a pterosaur, Serradraco sagittirostris, suggesting sheath tissue was present proximal to the dentition.
Beak tissues – which are sometimes called rhamphotheca – are not homogenous. Rather, they have different properties depending on their position and function. Beak tips (including the ‘nails’ of compound bird beaks) tend to be the hardest, most heavily keratinized tissue (Hieronymus and Witmer 2010) and are likely to be the most conspicuous, thickest part of the beak structure. Softer, rubbery rhamphothecal tissue occurs in regions which are prone to experiencing movement, such as the corners of the mouth, or in beaks overlying flexible jaw bones (such as the lower jaws of pelicans – see Field et al. 2011). Firmer, tougher parts of beaks are often gnarlier and/or more lustrous than those of more leathery and flexible regions.
Modern beaked reptiles are entirely toothless, but many fossil species – aetosaurs, many dinosaurs, pterosaurs and others – are only partly edentulous. Should we reconstruct their teeth emerging through beak tissues? Hieronymus and Witmer (2010) suggest that teeth and beaks do not mix, at least in some toothed birds. Correlates for compound beak tissues are found at the jaw tips of toothed birds such as Hesperornis but are not found adjacent to the toothrows, suggesting true rhamphothecal tissues were limited to edentulous parts of the mouth (Hieronymus and Witmer 2010). This relationship might not be universal, however. Some toothed pterosaurs, such as the Cretaceous pterosaur Serradraco, have correlates of cornified sheaths directly adjacent to their teeth, suggesting that their teeth did grow through a beak-like tissue (Fig. 7.16). Whether this was the case for all pterosaurs is not clear, however. Correlates for beak tissues are not as common in pterosaurs as current palaeoart suggests, being mostly restricted to certain toothless species, and there may be more variation in their jaw appearance than we currently realize: lips, partial beak covers or simply tough, leathery skin may have lined some pterosaur jaws instead of bird-like beaks. Perhaps this explains why beak tissues are relatively rare in specimens of pterosaurs which otherwise preserve abundant soft tissues, including delicate structures like fibres and wing membranes, and why our preserved beak tissues tend to be restricted to jaw tips (see Witton 2013 for a review). There is clearly scope for looking closely at extinct animal jaws to make our most educated inference about their covering: the presence or absence of teeth or an inherent ‘birdiness’ of a fossil animal does not necessarily equate to a bird-like beak.
Ears
Artists of ancient animals often restore ears incorrectly. The most common mistake is to place ear tissues within the temporal skull openings, presumably because of assumptions that these openings are ‘ear holes’. Ears are very anatomically variable because they evolved independently in each major tetrapod line (Christensen-Dalsgaard and Carr 2008), and many aspects of their evolution remain the subject of ongoing study. We know enough to say that development of ear tissues is far from simple in any group, and that some structures historically associated with ear anatomy are being reinterpreted as being related to jaw mechanics or nerve anatomy. Our situation is somewhat aided by the fact that artists only really need to concern themselves with tympanic ears, structures which are either openings on the side of animal heads covered with the tympanic membrane (the ear drum) or, when situated internally, are linked to external openings via tubes that allow sound to reach the ear. Several landmarks and features are relevant to credibly reconstructing external ear anatomy, but visible ear tissues are not present in all tetrapods, in some cases because they were never developed (in most early tetrapods), and in others because they were lost for adaptive purposes (particularly among aquatic or digging species).
Fig. 7.17 Ear position in early tetrapods and reptiles. Featured skulls (from top): Seymouria baylorensis; common tegu (Tupinambis teguixin); Caviramus schesaplanensis; Tyrannosaurus rex; Giraffatitan brancai.
The nature of the ear of the first tetrapods and their immediate relatives is somewhat mysterious, but it seems likely that they lacked visible ears. The temporal notch (historically known as the ‘otic notch’), a narrow invasion of the posterior skull margin (Fig. 7.17), has typically been taken as a sure-fire indicator of a tympanic membrane, but recent work suggests the first tetrapods used this structure for respiration (Clack 2002). Visible ears are likely in some temnospondyls and fossil lissamphibians however, their temporal notches being associated with a stapes (a bone involved in the detection of sound) configured to detect vibrations on a membrane (Clack 2002). This variation in non-amniote ears is reflected in living lissamphibians. Salamanders have relatively rudimentary ear structures with no external openings or markers, while many frogs have round, visible tympanic membranes in the temporal notch. When fossil frogs evolved this feature is unknown, but it seems likely that it was present in at least some Mesozoic frogs (Christensen-Dalsgaard and Carr 2008). The fact that modern frogs are quick to lose their external ear apparatus undermines confidence that it would be present in all fossil species, however.
The ears of reptile-line animals are, for artists, relatively simple to consider, being situated behind the skull in a notch formed by the quadrate bone (Fig. 7.17). The external ear is an opening into the lateral tissues of the head/neck junction, the size of which is directly related to the size and shape of the posterior quadrate margin. The ears of reptile-line animals are variable in appearance. They may have deep external ear tissues resembling large holes; a tympanic membrane almost confluent with the surrounding skin; a partial or full scale cover (seen in crocodylians and turtles), and others bury them under feathers (most birds). The covering of ear tissues seems to correlate at least in some cases with lifestyles that could damage delicate ear tissues, such as swimming or burrowing, and sustained evolution along these pathways can lead to a complete loss of external ear tissues (snakes, probably mosasaurs – Evans 2016). We can assume that the same morphological variation and selection pressures applied to fossil reptile ear morphology.
Fig. 7.18 Ear position, soft tissues and adaptive considerations in synapsids. Featured skulls and reconstructions are Thrinaxodon liorhinus and a domestic horse (Equus ferus).
Mammal-line animals are perhaps the champions of ear complexity and variation (Fig. 7.18). Modern mammals have a large external ear apparatus – the pinna – which is a conspicuous and functionally significant element of their cranial anatomy, but the size and shape of pinnae are impossible to predict without soft tissue preservation. The formation of the mammalian ear structure from bones ancestrally part of the tetrapod jaw – a classic evolutionary story recounted in countless textbooks – represents another issue: what did the ears of stem-mammals look like?
The first mammal-line species lack space at the back of the skull for placement of visible ear tissues, and it’s likely that they lacked them altogether. But later species show development of a notch at the back of the lower jaw which probably housed a tympanic membrane (Kermack and Kermack 2013). This notch is composed of bones that, over time, would divorce from the rest of the jaw to create bones of the true mammalian ear. It is small in animals like Dimetrodon, but becomes increasingly large in therapsid- and cynodont-grade animals, and is retained even in some close relatives of true mammals, such as the famous mammaliaform Morganucodon (Kermack and Kermack 2013). The location of the tympanic apparatus seems relatively shallow in these animals, so we might assume that their ear apparatus was quite conspicuous (Fig. 7.18).
Ear skeletal anatomy is largely separated from the lower jaw – attaining a condition broadly akin to that of living mammals – in some especially mammal-like Mammaliaformes (Luo et al. 2001), but it is not clear if this osteological development was paralleled by the development of ear pinnae. The cartilages and muscles that comprise pinna in living mammals almost entirely anchor to other soft tissues, leaving little scarring on skull bones from which we might deduce their presence. We are thus largely left to phylogenetic bracketing and a handful of well-preserved mammal fossil species to suggest when pinnae developed. It is almost certain that the common ancestor of marsupials and placental mammals (the ‘therian’ mammal group) had ear pinnae because they are ubiquitous in living members of this group. Preserved external ear tissues in the Cretaceous eutriconodont mammal Spinolestes, a mammal more closely related to therians than the egg-laying monotremes, confirm this interpretation (Martin et al. 2015). But were pinnae present in animals off the therian line? Living monotremes – the platypus and echidna – lack or have only partly developed pinnae tissues, and the well-known semi-aquatic docodont Castorocauda – representing a pre-monotreme grade of mammal evolution – seems to lack ear pinnae altogether (Luo et al. 2016). There are two ways to interpret this limited data. The first is to assume that the semi-aquatic habits of platypuses and Castorocauda, and the defensive adaptations of echidna, promoted loss of visible ear anatomy from an ancestor that had typically mammalian ear tissues. This could be viewed as special pleading however; perhaps a more defensible second interpretation is that visible pinnae were not ancestral to all mammals, but only developed on the therian branch of mammal evolution. Ultimately, the lack of data on pinnae evolution precludes any robust conclusion on their development at present. But wherever they appeared in mammal evolution, we might assume that their earliest guises were not fully-formed pinnae like we see in living therians. Presumably, ‘protopinna’ were raised tissues or lobes around the tympanic membrane, akin to an intermediate condition between the near-invisible pinnae of echidnas and more developed ears of therians.
Predicting when pinnae evolved is one problem: predicting their shape is another. Modern mammals give us plenty of ideas about the scope and range of ear shapes, and it’s perhaps apt to take artistic guidance from modern analogues and close relatives where appropriate. But for more exotic or distantly related mammals, considering ear function might be our best way of inferring pinnae shape. Cupped ears are ideal structures for detecting the direction of sound, and are common among living mammals because of their utility in pinpointing prey or locating predators. Larger and convoluted cupped ears offer greater hearing sensitivity, and are suited to animals which may have relied on sound to explore their surroundings, detect predators or spent time in dark conditions.
As prominent structures with a large surface area to volume ratios, mammal ears are also capable of losing heat quickly. Thus, big ears might disadvantage cold-climate species, while large ears can cool warm-climate species – as is most famously demonstrated by elephants. Some authors (Prothero 2013) have assumed that elephant-like ears might be a requirement for other gigantic mammals because of the difficulties that all large animals have with radiating body heat. We must be cautious about this however, as elephants are surprisingly terrible at keeping cool, and are not great models for thermoregulation in other large mammals. They are stocky, compact animals with low surface area to volume ratios; they lack sweat glands (Wright and Luck 1984); do not pant; and they live in climates which are so warm that for much of the day they cannot shed heat through simple convection, big ears or not (Weissenböck et al. 2012). Elephants have to regulate their temperature through moistening their skin with mud bathing and trunk spraying (Wright and Luck 1984), maintaining a sparse set of body hair to aid thermal escape (Myhrvold et al. 2012), having a high tolerance for body temperature change (Weissenböck et al. 2012), using ‘thermal windows’ in their skin to shed heat (Weissenböck et al. 2010), exploiting highly wrinkled skin to boost surface area and – of course – fanning their blood-vessel rich ears to help lose heat, when ambient temperatures are low enough for this to make a difference. Other large mammals, such as hippos, giraffes and rhinos, use more conventional means to keep cool and lack proportionally large ears; perhaps, unless we have reason to assume compromised thermoregulatory capabilities, we should not assume oversize ears in large fossil mammals.
Tongues, throats and other intraoral tissues
Where the internal tissues of animal mouths are visible, they need as much consideration as the external parts of the body. The oral cavities of living animals are visually complex, reflecting the influence of jaw skeletal architecture, development of the tongue and gums, the expanse of the throat, and – most unexpectedly – the presence of soft tissue ridges and spines that aid food manipulation (Fig. 7.19). Insights into many of these elements are afforded by fossils.
Fig. 7.19 The slightly terrifying interior mouth tissues of a Humboldt penguin (Spheniscus humboldti). Awesome intraoral tissues of this nature are common in animal species, especially in toothless taxa.
The basic architecture of the mouth interior is determined by the shape of the jaw bones. The shape of the palate (the roof of the mouth) and the length of the mandibular symphysis (the fusion between the right and left lower jaw bones at the jaw tip) are critical here. Palatal surfaces are not always flat and featureless: they are often concave or convex, have ridges or bosses, and many fossil species have additional teeth on the roof of their mouths. Similarly, not all animals have human-like jaws where only the tips of the lower jaw touch to fuse the end of the jaw. Many tetrapods have long mandibular symphyses where left and right jaws are fused for considerable portions of the jaw length. The biting surface of these symphyses can vary in much the same way as the bones of the upper jaw can, with varying concavities, convexities and protruding structures. These are important details to note because details of the lower jaw are often exposed more than the upper, depression of the mandible being the primary means of opening the mouth (as opposed to elevation of the skull). However long the symphysis is, its posterior margin marks the maximum forward extent of the throat tissues.
Tongues and throats are supported by bone and cartilage structures known as the hyoid apparatus. These elements are often not preserved but can, when present, inform interpretations of tongue length and robustness, as well as the nature of the throat. Very long posterior projections of the hyoid, which might even wrap around the back of the skull, reflect longer tongues, and robust hyoid bones can typify species with large, muscular throat tissues which can expand capaciously when feeding. Tongues themselves are entirely soft tissue and almost unknowable without the most exceptional soft tissue preservation. Forked tongues, however, do have an osteological correlate. The forked tongue tips of lizards and snakes insert into a pair of pits on the palatal surface which lead to the vomeronasal organ (also known as the Jacobson’s Organ) and, if we see these in fossil reptiles, they likely had a forked tongue. Unfortunately, vomeronasal openings are not the easiest structures to identify even for experts (Schulp et al. 2005), so careful research is encouraged to make reasoned interpretation of the (sometimes numerous) paired openings in fossil reptile mouths.

Fig. 7.20 Restoration of the toothless pterosaur Thalassodromeus sethi with barbed intraoral tissues and showing the complex skeletal structure of the jaw biting surfaces. (M. Witton)
The internal surfaces of animal mouths can be augmented with soft tissue structures that help food manipulation and, though their presence in fossil animals can only be speculated about, they are common enough to count as reasonable additions to many extinct species (Fig. 7.20). Such structures seem especially common in toothless animals and carnivores, and this may have been true of extinct forms too. Among the commonest structures of this kind are recurved spines lining the upper surface of the tongue and palate. These can be incredibly numerous, sometimes even spreading to other internal oral tissues, and are conspicuous when well-developed, such as in penguins and turtles. Some mammalian carnivores have roughened or barbed upper tongue surfaces to help hook and strip flesh. These structures are less visible at a distance, but any close views of such tongues bring them into focus. The tongues of browsing herbivores are often long and dextrous, helping to bring vegetation into the mouth when foraging.