ACCORDING TO THE ENACTIVE APPROACH, the human mind emerges from self-organizing processes that tightly interconnect the brain, body, and environment at multiple levels. The key ideas on which this proposition is based are those of autonomous systems and emergence or emergent processes. In this chapter, I explain these ideas. In the next chapter, I explore some connections between these ideas and phenomenological ideas about form, in particular forms or structures of behavior. These two chapters will lay the groundwork for the enactive strategy of addressing the explanatory gap by going back to the roots of mind in life and then working forward to subjectivity and consciousness.
In the first section of this chapter, I review some basic ideas about dynamic systems that form a background for the enactive approach. In the second section, I explain the notion of an autonomous system. A distinctive feature of the enactive approach is the emphasis it gives to autonomy. In brief, an autonomous system is a self-determining system, as distinguished from a system determined from the outside, or a heteronomous system. On the one hand, a living cell, a multicellular animal, an ant colony, or a human being behaves as a coherent, self-determining unity in its interactions with its environment. An automatic bank machine, on the other hand, is determined and controlled from the outside, in the realm of human design. The paradigm for interaction with a heteronomous system is input/processing/output, in which deviations from desired outputs are seen as system errors. The paradigm for interaction with an autonomous system is a conversation, in which unsatisfactory outcomes are seen as breaches of understanding (Varela 1979, p. xii). According to the enactive approach, living beings and cognitive agents need to be understood as autonomous systems. I discuss the implications of this autonomy perspective for how we think about information in the third section.
In the fourth section, I turn to emergence, a now familiar notion that describes the arising of large-scale, collective patterns of behavior in complex systems as diverse as cells, brains, ecosystems, cities, and economies. Emergence is closely related to self-organization and circular causality, both of which involve the reciprocal influence of “bottom-up” and “top-down” processes. For example, a tornado emerges through the self-organization of circulating air and water particles; it reciprocally sucks those particles into a particular macroscopic configuration, with devastating effect for anything in its path. In this section, I sketch a way of thinking about emergence that I call dynamic co-emergence. Dynamic co-emergence means that a whole not only arises from its parts, but the parts also arise from the whole. Part and whole co-emerge and mutually specify each other. A whole cannot be reduced to its parts, for the parts cannot be characterized independently of the whole; conversely, the parts cannot be reduced to the whole, for the whole cannot be characterized independently of the parts. I discuss philosophical issues related to this conception of emergence in Appendix B.
In recent years growing interest in the dynamics of cognition and emotion has given rise to a distinct dynamical approach in mind science (Kelso 1995; Lewis and Granic 2000; Port and van Gelder 1995; Thelen and Smith 1994). The central idea of the dynamical approach is that natural cognition—cognition in evolved, living agents—is a dynamic phenomenon and accordingly needs to be understood from the perspective of the science of dynamic systems. This perspective includes dynamic-systems theory (a branch of pure mathematics), dynamic-systems modeling (mathematical modeling of empirical systems), and experimental investigations of biological and psychological phenomena informed by these tools.
The first important concept we need to introduce in this context is that of a dynamic system.1 In simple terms a dynamic system is one that changes over time. The term system, however, is ambiguous, in that it can refer either to an actual system in the world, such as the solar system, or to a mathematical model of an actual system. In the case of the actual world, the term system does not admit of precise definition. In general, a system is a collection of related entities or processes that stands out from a background as a single whole, as some observer sees and conceptualizes things. The classic example from the history of science is the solar system. Its components are the sun, moon, and planets, and its states are their possible configurations. What changes over time is the state of the system. A dynamic system in the sense of a model, however, is a mathematical construction that aims to describe and predict the way an actual system changes over time (the paths of the planets, and events such as eclipses, in the case of the solar system). To this end, some aspects of the actual system are singled out as being especially important and are mathematically represented by quantitative variables. Specifying the numerical values of all the variables at a given time indicates the state of the system at that time. A dynamic system includes a procedure for producing such a description of the state of the system and a rule for transforming the current state-description into another state-description for some future time. A dynamic system is thus a mathematical model for the way that a system changes or behaves as time passes.
If the passage of time is considered to be continuous (like the sweeping second hand of an analogue clock), then the dynamic system is a differentiable one: the variables change in a smooth and continuous way, and the rules or “evolution equations” that govern the changing state of the system take the form of differential equations. If time is considered to pass in evenly spaced, discrete jumps (like a digital clock), then the system is described by a difference equation or a mapping (a function repeatedly applied or iterated in discrete time steps). Some differential equations have an analytical solution, which means they can be exactly solved by mathematical formulas. Given the starting values of the variables (the initial conditions), then all future states of the system can be known without recalculating the state of the system for each time increment. Most differential equations, however, cannot be solved in this way. When the equations contain nonlinear terms—functions in which the value of the output is not directly proportional to the sum of the inputs—then such a solution is impossible. Therefore a different mathematical approach has to be taken from that of finding a formula that makes possible the prediction of a future state from a present one.
This other approach, introduced by Henri Poincaré in the nineteenth century, is known as the qualitative study of differential equations (or of nonlinear differentiable dynamic systems). One thinks of the space of all possible states of the system as a geometric space, known as state space or phase space, and the way that the system changes or behaves over time as curves or trajectories in this space. Instead of seeking a formula for each solution as a function of time, one studies the collection of all solutions (corresponding to trajectories in phase space) for all times and initial conditions at once (Norton 1995, p. 46). This approach is said to be qualitative because it uses topological and geometrical techniques to study the general or global character of the system’s long-term behavior (its behavior in phase space), instead of seeking to predict the system’s exact future state (the specific values of its variables at a future time). It is precisely this qualitative approach to dynamics that goes by the name of dynamic systems theory.
We need to introduce one more related notion—that of complexity. The term complexity describes behavior that is neither random nor ordered and predictable; rather, it is in between, exhibiting changing and unstable patterns. Of particular importance in the context of recent nonlinear dynamic-systems approaches to the brain and behavior is the notion of complexity as dynamic instability or metastability—“the successive expression of different transient dynamics with stereotyped temporal patterns being continuously created and destroyed and reemerging again” (Friston 2000b, p. 238). Recent science indicates that complexity of this sort can be found at numerous scales and levels, from the molecular and organismic to the ecological and evolutionary, as well as the neural and behavioral.2 In every case the message seems to be that complexity, instability, or metastability is necessary for self-organization and adaptive behavior.
We can now return to the dynamical approach in mind science. The fundamental dynamical hypothesis of this approach is that natural cognitive agents (people and other animals) are dynamic systems (or, more precisely, that the cognitive systems agents instantiate are dynamic systems), and that accordingly action, perception, and cognition should be explained in dynamic terms (van Gelder 1998). Proponents of the dynamical hypothesis contrast it with the cognitivist hypothesis, which states that cognitive agents (or the cognitive systems they instantiate), whether natural or artificial, are digital computers or physical symbol systems and that accordingly cognition should be explained in symbol-processing terms.
To illustrate these ideas, we can turn to research on neural and behavioral coordination dynamics by Haken, Kelso, and colleagues (Bressler and Kelso 2001; Kelso 1995). One case they have studied is rhythmic finger movement (Haken, Kelso, and Bunz 1985). The experimental task was to move the two index fingers at the same frequency from side to side. At low speeds, there are two comfortable coordination patterns (the system is bistable): either the fingers move in-phase (equivalent muscle groups in each hand contract simultaneously) or anti-phase (equivalent muscle groups alternate in their contraction and expansion). As the speed gradually increases, the in-phase pattern becomes unstable, and eventually at a certain critical frequency the fingers spontaneously switch to an anti-phase pattern (the system undergoes a bifurcation). As the speed decreases, the in-phase pattern becomes stable again, but it does so below the original switching point (this delayed return to a previous state is known as hysteresis).
Haken, Kelso, and colleagues devised a dynamic-systems model to describe and predict these properties of motor behavior. The model describes how the relative phase relation between the two fingers evolves over time. Relative phase is an example of a “collective variable”—one whose value is set by the relation between the values of other variables, in this case those describing the individual finger movements. A collective variable describes a high-level or global characteristic of a system that emerges as a coherent and ordered pattern from the interactions of the system’s components. This macrolevel pattern is also known as an order parameter because it reduces the degrees of freedom of the system’s components by organizing them into a coherent and ordered pattern. When the fingers move in-phase, the collective variable or order parameter of relative phase is zero; once the critical transition or bifurcation to anti-phase happens, the relative phase becomes nonzero up to some maximum value. Because the phase transition occurs at a certain critical frequency of finger oscillation, the frequency acts as a “control parameter” for the system. The control parameter does not dictate or prescribe the collective variable or order parameter (the emergent pattern of relative phase). Rather, its changing values lead the system through a variety of possible patterns or states (Kelso 1995, p. 7). Thus the model mathematically describes how the control parameter of finger-movement frequency leads the system through different patterns of finger coordination.
In the language of dynamic-systems theory, this kind of description gives the state space of the system—the abstract and multidimensional space that represents all possible states of the system by specifying all possible values of the system’s variables. The temporal evolution of the system corresponds to its trajectory through this space. The model predicts the observed switching from one phase to another without positing any internal motor program that directs the switches by issuing symbolic instructions. Instead, the phase transitions occur spontaneously as emergent properties of the system’s self-organizing dynamics. Kelso and colleagues have extended and developed this type of phase-transition model to apply to a wide variety of cognitive domains, such as motor skill learning, speech perception, visual perception, and the dynamic coordination of activity among cortical areas of the brain (Bressler and Kelso 2001; Kelso 1995).
One of the key points relating to the dynamical approach is its emphasis on time. Traditional computational models are static in that they specify only a sequence of discrete states through which the system must pass. In contrast, dynamic-systems models specify how processes unfold in real time. As Tim van Gelder states, “Although all cognitive scientists understand cognition as something that happens over time, dynamicists see cognition as being in time, that is, as an essentially temporal phenomenon (van Gelder 1999a, p. 244). Van Gelder (1998) describes this contrast as one between change versus state; geometry versus structure; structure in time versus static structure; time versus order; parallel versus serial; and ongoing versus input/output.
Whereas computationalists focus primarily on discrete states and treat change as what happens when a system shifts from one discrete state to another, dynamicists focus on how a system changes state continuously in time. Dynamicists conceive of state changes geometrically, in terms of their position and trajectory in phase space, whereas computationalists focus on the internal formal or syntactic structure of combinatorial entities. Computationalists think of these structures as laid out statically (like snapshots), as either present all at once or not, and hence of cognition as the rule-governed transformation of one such static structure into another. For dynamicists, cognitive structures are laid out as temporally extended patterns of activity, and cognition is seen as the flow of complex temporal structures mutually and simultaneously influencing each other. Dynamicists are therefore interested in the timing (rates, periods, durations, synchronies) of processes, whereas computationalists have traditionally been interested only in the order of cognitive states. Moreover, computationalists tend to think of this order as being the serial or sequential progression of sense → perceive → think → act, whereas for dynamicists cognition unfolds as the continuous coevolution of acting, perceiving, imagining, feeling, and thinking. Finally, whereas computationalists think of cognitive processes as having an input-output structure—the system receives an input, proceeds through a sequence of internal operations, produces an output, and then halts—dynamicists think of processes as always ongoing, with no clear starting or end points. The goal is not to map an input at one time onto an output at a later time, but always to maintain appropriate change (van Gelder 1998).
The dynamicist idea that cognitive processes are always ongoing with no clear starting or end points can be deepened by introducing the distinction between autonomous and heteronomous systems. Autonomy and heteronomy literally mean, respectively, self-governed and other-governed. A heteronomous system is one whose organization is defined by input-output information flow and external mechanisms of control. Traditional computational systems, cognitivist or connectionist, are heteronomous. For instance, a typical connectionist network has an input layer and an output layer; the inputs are initially assigned by the observer outside the system; and output performance is evaluated in relation to an externally imposed task. An autonomous system, however, is defined by its endogenous, self-organizing and self-controlling dynamics, does not have inputs and outputs in the usual sense, and determines the cognitive domain in which it operates (Varela 1979; Varela and Bourgine 1991).
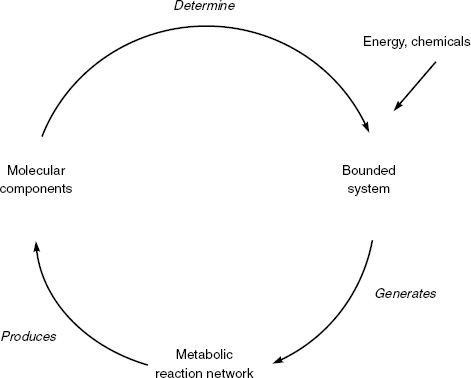
In general, to specify any system one needs to describe its organization—the set of relations that defines it as the system it is. In complex systems theory, the term autonomous refers to a generic type of organization. The relations that define the autonomous organization hold between processes (such as metabolic reactions in a cell or neuronal firings in a cell assembly) rather than static entities. In an autonomous system, the constituent processes (i) recursively depend on each other for their generation and their realization as a network, (ii) constitute the system as a unity in whatever domain they exist, and (iii) determine a domain of possible interactions with the environment (Varela 1979, p. 55). The paradigm is a living cell. The constituent processes in this case are chemical; their recursive interdependence takes the form of a self-producing, metabolic network that also produces its own membrane; and this network constitutes the system as a unity in the biochemical domain and determines a domain of possible interactions with the environment. This kind of autonomy in the biochemical domain is known as autopoiesis (Maturana and Varela 1980). Figure 3.1 illustrates the basic organization required for autopoietic autonomy.
Autopoiesis is the paradigm case of biological autonomy for two reasons. It is empirically the best understood case, and it provides the core “biologic” of all life on Earth. To qualify as autonomous, however, a system does not have to be autopoietic in the strict sense (a self-producing bounded molecular system). An autopoietic system dynamically produces its own material boundary or membrane, but a system can be autonomous without having this sort of material boundary. The members of an insect colony, for example, form an autonomous social network, but the boundary is social and territorial, not material.
In exploring the notion of autonomy, we can take two complementary approaches—a top-down approach and a bottom-up one (Ruiz-Mirazo and Moreno 2004). Both approaches see autonomy as a relational, system-level property, but there is a critical difference between the two. Whereas the top-down approach focuses on the relational organization proper to autonomy, the bottom-up approach emphasizes the energetic and thermodynamic requirements for autonomy.
Varela takes the top-down approach in his 1979 book, Principles of Biological Autonomy.3 In this work he defines an autonomous system as a system that has organizational closure (later called operational closure) (Varela 1979, pp. 55–60). Here closure does not mean that the system is materially and energetically closed to the outside world (which of course is impossible). On the contrary, autonomous systems must be thermodynamically far-from-equilibrium systems, which incessantly exchange matter and energy with their surroundings. Organizational closure refers to the self-referential (circular and recursive) network of relations that defines the system as a unity, and operational closure to the reentrant and recurrent dynamics of such a system.4 An autonomous system is always structurally coupled to its environment. Two or more systems are coupled when the conduct of each is a function of the conduct of the other. (In dynamic-systems language, the state variables of one system are parameters of the other system, and vice versa.) “Structural coupling” refers to the history of recurrent interactions between two or more systems that leads to a structural congruence between them (Maturana 1975; Maturana and Varela 1987, p. 75). Thus the state changes of an autonomous system result from its operational closure and structural coupling. The result of any state change is always further self-organized activity within the system, unless its closure is disrupted and it is no longer able to carry on its coupling, in which case it disintegrates. Systems described as autonomous in this sense abound throughout the living world—single cells, microbial communities, nervous systems, immune systems, multicellular organisms, ecosystems, and so on. Such systems need to be seen as sources of their own activity, specifying their own domains of interaction, not as transducers or functions for converting input instructions into output products. In other words, the autonomous character of these systems needs to be recognized.
The second, bottom-up approach to autonomy builds on these notions of organizational and operational closure, but tries to work out the energetic and thermodynamic requirements for the instantiation of “basic autonomy” in the physical world. From this perspective, basic autonomy is “the capacity of a system to manage the flow of matter and energy through it so that it can, at the same time, regulate, modify, and control: (i) internal self-constructive processes and (ii) processes of exchange with the environment” (Ruiz-Mirazo and Moreno 2004, p. 240). This capacity brings with it specific and demanding physical-implementation requirements: the system must have certain types of components, specifically a semipermeable active boundary (a membrane), an energy transduction/conversion apparatus (an energy currency such as adenosine triphosphate (ATP) in living cells, which transfers energy from chemical bonds to energy-absorbing reactions within the cell), and at least one type of component that controls and facilitates the self-construction processes (catalysts) (Ruiz-Mirazo and Moreno 2004, p. 252).
Figure 3.1 depicts the basic autopoietic organization for a living cell. A cell stands out of a molecular soup by creating the boundaries that set it apart from what it is not and that actively regulate its interactions with the environment. Metabolic processes within the cell construct these boundaries, but the metabolic processes themselves are made possible by those very boundaries. In this way, the cell emerges as a figure out of a chemical background. Should this process of self-production be interrupted, the cellular components no longer form a unity, gradually diffusing back into a molecular soup.
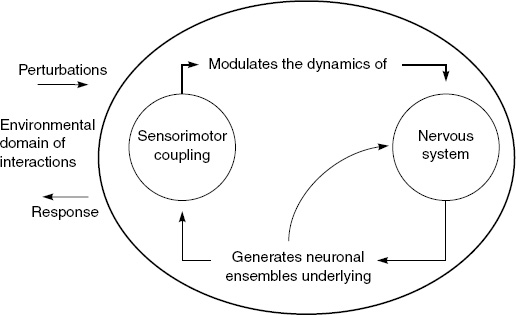
Figure 3.1 can be compared with Figure 3.2, which depicts the minimal form of organizational closure for a nervous system. Any nervous system operates according to a basic “neurologic,” a pattern that continues and elaborates the biologic of autopoiesis. The fundamental logic of the nervous system is to couple movement and a stream of sensory activity in a continuous circular fashion (Maturana and Varela 1987, pp. 142–176). Wherever movement is essential to a multicellular organism’s mode of life, there is the corresponding development of a nervous system. A nervous system links sensory surfaces (sense organs and nerve endings) and effectors (muscles, glands) within the body. In this way it integrates the organism, holding it together as a mobile unity, as an autonomous sensorimotor agent.
This neurologic underlies all the variations on sensorimotor coordination found in the animal kingdom. In all animals, neuronal networks establish and maintain a sensorimotor cycle through which what the animal senses depends directly on how it moves, and how it moves depends directly on what it senses. No animal is a mere passive respondent; every animal meets the environment on its own sensorimotor terms. Merleau-Ponty recognized this crucial point in his first work, The Structure of Behavior:
The organism cannot properly be compared to a keyboard on which the external stimuli would play and in which their proper form would be delineated for the simple reason that the organism contributes to the constitution of that form. When my hand follows each effort of a struggling animal while holding an instrument for capturing it, it is clear that each of my movements responds to an external stimulation; but it is also clear that these stimulations could not be received without the movements by which I expose my receptors to their influence . . . When the eye and the ear follow an animal in flight, it is impossible to say “which started first” in the exchange of stimuli and responses. Since all movements of the organism are always conditioned by external influences, one can, if one wishes, readily treat behavior as an effect of the milieu. But in the same way, since all the stimulations which the organism receives have in turn been possible only by its preceding movements which have culminated in exposing the receptor organ to the external influences, one could also say that the behavior is the first cause of the stimulations.
Thus the form of the excitant is created by the organism itself, by its proper manner of offering itself to actions from outside. Doubtless, in order to be able to subsist, it must encounter a certain number of physical and chemical agents in its surroundings. But it is the organism itself—according to the proper nature of its receptors, the thresholds of its nerve centers and the movements of the organs—which chooses the stimuli in the physical world to which it will be sensitive. “The environment (Umwelt) emerges from the world through the actualization or the being of the organism—[granted that] an organism can exist only if it succeeds in finding in the world an adequate environment.” This would be a key board which moves itself in such a way as to offer—and according to variable rhythms—such or such of its keys to the in itself monotonous action of an external hammer. (1963, p. 13)5
This passage clearly expresses an autonomy perspective. Organisms display patterns of behavior that require us to see them as autonomous. Varela tries to characterize this autonomy at an abstract level in terms of a generic dynamic pattern or form, namely, organizational and operational closure. Hence Varela gives us his “Closure Thesis,” which states, “Every autonomous system is organizationally closed” (Varela 1979, p. 58).6
Figure 3.1 illustrates the minimal form this closure takes for life at the single-cell level, and Figure 3.2 illustrates the minimal form it takes for the nervous system. Whereas autopoietic closure brings forth a minimal “bodily self” at the level of cellular metabolism, sensorimotor closure produces a “sensorimotor self” at the level of perception and action. In the one case the passage from network closure to selfhood (and correlative otherness) happens at the level of an active semipermeable boundary or membrane, which regulates interaction with the outside environment. In the other case it happens at the level of behavior and intentional action. In both cases we see the co-emergence of inside and outside, of selfhood and a correlative world or environment of otherness, through the generic mechanism of network closure (autonomy) and its physical embodiment (Varela 1997a; see also Moreno and Barandiaran 2004).7
In addition to these cellular and sensorimotor forms of selfhood, other forms of selfhood arise from other organizationally and operationally closed systems. The immune system, for instance—understood as an autonomous immune network that establishes a coherent somatic identity for the organism, rather than as a mere mechanism of defense—brings forth a dynamic, somatic identity at a distributed cellular and molecular level (Coutinho 2003; Varela and Coutinho 1991). The animate form of our living body is thus the place of intersection for numerous emergent patterns of selfhood and coupling. Whether cellular, somatic, sensorimotor, or neurocognitive, these patterns derive not from any homuncular self or agent inside the system organizing it or directing it, but from distributed networks with operational closure. In Varela’s image, our organism is a meshwork of “selfless selves,” and we are and live this meshwork (Varela 1991; Varela and Cohen 1989).
Let me forestall an objection that might arise at this point. The nervous system is clearly embedded in the body of the organism, and the organism in its environment (Chiel and Beer 1997). This fact seemingly contradicts the statement that the nervous system is an autonomous system and that the organism is an autonomous agent. The thought here would be that the operation of the nervous system loops through the body (via sensory and motor surfaces), and therefore it is not possible that the nervous system has operational closure (that the product of every process within the system stays within that system). Similarly, because the bodily activity of the organism loops through the environment (motor activity affects the sensory stimulation one receives back from the environment), the organism cannot have an operationally closed dynamics.
A number of points, both methodological and epistemological, need to be made in reply. The first point is that, strictly speaking, system, autonomy, and heteronomy are heuristic notions—they are cognitive aids or guides in the scientific investigation and characterization of observable phenomena and patterns of behavior. As heuristic notions, they (implicitly) refer back to and implicate the interpretive and explanatory stance of an observer (or observer community). What counts as the system in any given case, and hence whether it is autonomous or heteronomous, is context-dependent and interest-relative. For any system it is always possible to adopt a heteronomy or external-control perspective, and this can be useful for many purposes. Nevertheless, this stance does not illuminate—and indeed can obscure—certain observable patterns of behavior, namely, patterns arising from the system’s internal dynamics rather than external parameters. An organism dynamically produces and maintains its own organization as an invariant through change, and thereby also brings forth its own domain of interaction. (Although living organisms are paradigmatic in this regard, nothing apparently rules out the possibility of artificial autonomy.) A heteronomy perspective does not provide an adequate framework to investigate and understand this phenomenon; an autonomy perspective is needed.
The second point is that in any given case or for any candidate system we need to distinguish between, on the one hand, the operation of the system as such, which is a function of both its organization (the set of relations that defines it as a system) and physical structure, and, on the other hand, its performance in relation to whatever wider context in which it is observed. For example, if we wish to characterize the organization and operation of the nervous system as a finite neuronal network, then we need to characterize the nervous system as organizationally and operationally closed, such that any change of activity in a neuron (or neural assembly) always leads to a change of activity in other neurons (either directly through synaptic action or indirectly through intervening physical and chemical elements). Sensory and effector neurons are no exception because any change in the one leads to changes in the other, such that the network always closes back upon itself, regardless of intervening elements (Maturana and Varela 1980, p. 127). Nevertheless, the domain of states available to the nervous system (as an operationally closed network) is clearly a function of its history of interactions with the rest of the body (and the environment). Hence, besides characterizing the nervous system’s operation as a closed network, we need to characterize its performance in its structural coupling with the rest of the body (and the environment). Similarly, to characterize the organism as a finite cellular or multicellular entity, we need to characterize it as an organizationally and operationally closed system. At the same time, we need to characterize the organism’s performance or behavior in its structural coupling with the environment.
We can also shift perspectives and characterize the nervous system as a heteronomous system—that is, as a component system with various functions defined in relation to the organism (such as registering sensory changes or guiding movement). Notice, however—and this is the third point—that in so shifting perspectives we are ipso facto no longer talking about the same system. The system with which we are now concerned is no longer the nervous system as a finite neuronal network, but rather the larger system of the organism (in which the nervous system is seen as a component). Similarly, we can also shift perspectives and characterize the organism as a heteronomous system subject to the control of the environment (for instance, other organisms). Once again, in thus shifting perspectives we are ipso facto no longer dealing with the same system. The system now is the larger system of organism-plus-environment, not the organism as a finite cellular or multicellular entity.8
These considerations show us that there is no inconsistency between characterizing the nervous system and organism as autonomous and emphasizing their somatic and environmental embeddedness. We do, however, have to keep our logical and conceptual accounts clear, so that we know which explanatory heuristic is in play at any given time. In any case, for the enactive approach it is the autonomy perspective on natural cognitive agents that remains the reference point for understanding mind and life, not a predefined input-output task strucure.
Adopting an autonomy perspective also brings with it a certain way of thinking about semantic information or meaning. For enactive theorists, information is context-dependent and agent-relative; it belongs to the coupling of a system and its environment. What counts as information is determined by the history, structure, and needs of the system acting in its environment.
According to the received view in cognitive science, in order to explain cognitive abilities we need to appeal to information-bearing states inside the system. Such states, by virtue of the semantic information they carry about the world, qualify as representations. Cognitivists conceive of these representations as symbols in a computational “language of thought,” and connectionists as constrained patterns of network activity corresponding to phase space “attractors” (regions of phase space toward which all nearby trajectories converge). In either case there is a strong tendency to adopt an objectivist conception of representation: representations are internal structures that encode context-independent information about the world, and cognition is the processing of such information.
This objectivist notion of information presupposes a heteronomy perspective in which an observer or designer stands outside the system and states what is to count as information (and hence what is to count as error or success in representation). Information looks different from an autonomy perspective. Here the system, on the basis of its operationally closed dynamics and mode of structural coupling with the environment, helps determine what information is or can be.
A neurobiological example can help to illustrate these ideas.9 Certain kinds of cortical neurons are often described as feature detectors because they respond preferentially (fire above their base rate) to various types of stimuli, such as edges, lines, and moving spots. Such neurons are identified by recording their individual activity with a microelectrode and determining the sensory stimulus to which the neuron is most sensitive. Such neurons are said to “represent” features of objects and to make that information available for futher processing by various systems in the brain. This view lies behind the standard formulation of the so-called binding problem. This problem concerns how distinct features (shape, color, motion), as represented by cell populations in spatially distributed and functionally segregated neural pathways, can be bound together to form a complete and accurate representation of the object (so that the right shapes go with the right colors and motions). This way of thinking about the brain treats it as a heteronomous system: object features outside the organism provide informational inputs to the brain, and the brain’s information processing task is to arrive at an accurate representation of the objective world and produce an adaptive motor output.
From an autonomy perspective, it is crucial to distinguish between information about stimuli as they are defined by an observer and information in the sense of what meanings the stimuli have for the animal. Only the latter play a significant role in the brain’s operation. The notion of an object “feature” is defined by an observer who stands outside the system, has independent access to the environment, and establishes correlations between environmental features and neuronal responses. The animal’s brain has no access to features in this sense (and a fortiori has no access to any mapping from features to neuronal responses). As Freeman explains, “In the view from neurodynamics, neurons that respond to edges, lines, and moving spots are manifesting the local topological properties of neuronal maps, which extract local time and space derivatives in automatic preprocessing for spatial and temporal contrast enhancement. No objects or features are manifested at the level of the single neuron, assuredly not those used by an observer” (Freeman 1995, p. 54). From an autonomy perspective, individual neurons do not detect objectively defined features. Rather, assemblies of neurons make sense of stimulation by constructing meaning, and this meaning arises as a function of how the brain’s endogenous and nonlinear activity compensates for sensory perturbations. From this perspective, the feature-binding problem is not the brain’s problem, but the brain theorist’s problem; it is an artifact of a certain way of looking at the brain. Freeman’s description of the alternative view, based on looking at the brain as an autonomous system operating according to nonlinear causality, is well worth quoting here:
In this view the experimenter trains a subject to co-operate through the use of positive and negative reinforcement, thereby inducing a state of expectancy and search for a stimulus, as it is conceived by the subject. When the expected stimulus arrives, the activated receptors transmit pulses to the sensory cortex, where they elicit the construction by nonlinear dynamics of a macroscopic, spatially coherent oscillatory pattern that covers an entire area of sensory cortex . . . It is observed by means of the electroencephalogram (EEG) from electrode arrays on all the sensory cortices . . . It is not seen in recordings from single neuron action potentials, because the fraction of the variance in the single neuronal pulse train that is covariant with the neural mass is far too small, on the order of 0.1 percent.
The emergent pattern is not a representation of a stimulus . . . It is a state transition that is induced by a stimulus, followed by a construction of a pattern that is shaped by the synaptic modification among cortical neurons from prior learning. It is also dependent on the brain stem nuclei that bathe the forebrain in neuromodulatory chemicals. It is a dynamic action pattern that creates and carries the meaning of the stimulus for the subject. It reflects the individual history, present context, and expectancy, corresponding to the unity and wholeness of intentionality. Owing to dependence on history, the patterns created in each cortex are unique to each subject. (Freeman 1999b, pp. 149–150)
The distinction between autonomous meaning-construction and heteronomous information processing needs to be placed in the broader context of the embodied dynamicist way of thinking about information. To explain this way of thinking, it will be helpful to go back to ideas introduced by Howard Pattee (1977). Pattee made an important distinction between two modes of description of a complex system—the linguistic mode, which describes the system in terms of discrete, rate-independent, symbolic elements, and the dynamical mode, which describes the system in terms of continuous, rate-dependent processes, and thus explicitly includes the flow of time. Pattee raised the following question: “How do we know we are not intepreting certain structures as descriptions, only because we recognize them as consistent with rules of one of our own languages?” (1977, p. 262). In other words, how do we know our linguistic descriptions are not simply observer-relative, but rather correspond to symbolic structures that belong to the system itself and play a role in its operation? And he answered: “we must further restrict our model of a complex system to remove the case of the external observer reading a message that is not really in the system itself. This restriction is achieved by requiring that a complex system must read and write its own messages” (1977, p. 262).
Pattee’s example is a living cell. When we describe DNA triplets as “coding for” amino acids, we employ the linguistic mode of description. Which amino acid a given DNA triplet specifies is supposed to be rate-independent—it does not matter how fast the triplet is “read” in the course of protein synthesis. It is also supposed to be arbitrary, in the sense that “[i]t is hard to see why a code in which GGC means glycine and AAG means lycine is either better or worse than one in which the meanings are reservsed” (Maynard Smith 1986, p. 19). According to Pattee, the linguistic mode of description in this case is not observer-relative because the cell is a self-describing system that “reads and writes its own messages.” The writing of its own messages corresponds to DNA replication (the production of a complement of the original DNA molecule through a template); the reading of its own messages corresponds to protein synthesis (DNA “transcription” to RNA and RNA “translation” to protein).
Pattee then makes a number of crucial points. First, for the code to be read there must ultimately be a transduction or conversion within the cell from the linguistic mode to the dynamical mode. This conversion occurs when the rate-independent linear array of amino acids folds to become a three-dimensional enzyme. Within the life cycle of the cell, there is thus a transformation from the enzyme as something designated in the genome to the enzyme as an operational component of metabolism. Second, this transformation (the protein folding) is not itself linguistically described in the cell, but rather happens according to physical law (under the higher-order constraint of the DNA-specificed amino acid sequence). Third, if the transformation were linguistically described, the speed and precision with which it is accomplished would be considerably compromised. Pattee’s conclusion is that “we would not expect a complete formal description or simulation of a complex system to adapt or function as rapidly or reliably as the partially self-describing, tacit dynamic system it simulates” (1977, p. 264).
Pattee emphasizes the complementarity of the linguistic and dynamical modes of description, but also suggests that symbolic information emerges from and acts as a constraint on dynamics. This idea is important for embodied dynamicism and the enactive approach. Let us return to the example of the cell. In general, nucleotide triplets are capable of predictably specifying an amino acid if and only if they are properly embedded in the cell’s metabolism, that is, in a multitude of enyzmatic regulations in a complex biochemical network. This network has a chicken-and-egg character at several levels. First, proteins can arise only from a DNA/RNA “reading” process, but this process cannot happen without proteins. Second, the DNA “writing” and “reading” processes must be properly situated within the intracellular environment, but this environment is a result of those very processes. Finally, the entire cell is an autopoietic system—that is, an autonomous system defined by an operationally closed network of molecular processes that simultaneously both produces and realizes the cell concretely in the physical space of its biochemical components.
Now, when we employ the linguistic mode of description and state that DNA/RNA “codes” for proteins, we restrict our focus to one particular sequence of this overall circular causality. We abstract away from the many intervening and necessary causal steps in the actual dynamic process of protein synthesis, and we bracket out the essential participation of many other molecular elements (such as RNA polymerase enzymes, and positive and negative regulatory proteins). We “thus reduce our description to a skeleton that associates a certain part of a nucleic acid with a certain protein segment. Next we observe that this kind of simplified description of an actual dynamic process is a useful one in following the sequences of reproductive steps from one generation to the other, to the extent that the dynamic process stays stable (i.e., the kinds of dynamics responsible for bonding, folding, and so on) . . . A symbolic explanation, such as the description of some cellular components as genes, betrays the emergence of certain coherent patterns of behavior to which we choose to pay attention” (Varela 1979, p. 75). It is the emergence of such coherent dynamic patterns that underwrites the symbolic informational level of description: “An object or event is a symbol only if it is a token for an abbreviated nomic chain that occurs within the bounds of the system’s organizational closure. In other words, whenever the system’s closure determines certain regularities in the face of internal or external interactions and perturbations, such regularities can be abbreviated as a symbol, usually the initial or terminal element in the nomic chain” (Varela 1979, pp. 79–80). Thus, when we talk about DNA “coding” for proteins we are not referring to a special type of symbolic causal relation or a special type of intrinsically informational molecule that rises above the dynamic fray. Rather, we are abbreviating a lengthy but remarkably stable dynamic pattern of biochemical events. It is precisely the stability and predictability of the entire pattern that allows us to telescope it in a linguistic mode of description by treating nucleotide triplets as in effect “standing for” amino acids.
This mode of description is unobjectionable (and has heuristic value) as long as it is remembered that the genetic code is no more than a rule of causal specificity based on the fact that cells use nucleic acids as templates for the primary structure (amino acid sequence) of proteins (Godfrey-Smith 2000b; Thompson 1997). Yet it is unacceptable to say that DNA contains the information for phenotypic design, because this statement attributes an intrinsic semantic-informational status to one particular type of component and thereby divests this component of its necessary embedding in the dynamics of the autopoietic network. It is this network in its entirety that specifies the phenotypic characteristics of a cell, not one of its components, and it is this network as a whole that serves as the precondition and causal basis of DNA replication (“writing”) and protein synthesis (“reading”) (see Moss 1992). Information is not intrinsic to the static linear array of the DNA sequence, but is rather dynamically constituted in and by the cell as an autopoietically organized, three-dimensional entity—by the cell as a body. In summary, the linguistic mode is emergent from the dynamical mode, and information exists only as dynamically embodied.
With these points having been made, we return to the difference between autonomous meaning-construction and heteronomous information processing. Information is formed within a context rather than imposed from without. Gregory Bateson used to say, “information is a difference that makes a difference” (Bateson 1972, p. 315). We could elaborate this insight by saying that information, dynamically conceived, is the making of a difference that makes a difference for somebody somewhere (see Oyama 2000b). Information here is understood in the sense of informare, to form within (Varela 1979, p. 266). An autonomous system becomes informed by virtue of the meaning formation in which it participates, and this meaning formation depends on the way its endogenous dynamics specifies things that make a difference to it (Kelso and Kay 1987; Turvey and Shaw 1999).
For another example we can return to the finger coordination study of Haken, Kelso, and Bunz (1985). There the switching from in-phase to anti-phase happens without any command from a motor program; rather, it occurs spontaneously as an emergent property of the system’s self-organizing dynamics. The collective variable or order parameter of relative phase is informational in the sense that it specifies coherent patterns or relations that inform the system and that can be physically or physiologically realized in multiple ways. As Kelso explains:
Instead of treating dynamics as ordinary physics using standard biophysical quantities such as mass, length, momentum, and energy, our coordination or pattern dynamics is informational from the very start. The order parameter, ϕ [relative phase], captures the coherent relations among different kinds of things. Unlike ordinary physics, the pattern dynamics is context dependent: the dynamics are valid for a given biological function or task, but largely independent of how this function is physiologically implemented. Thus, if we accept that the same order parameter, Φ, captures coherent spatiotemporal relations among different kinds of things, and the same equations of motion describe how different coordination patterns form, coexist, and change, it seems justified to conclude that order parameters in biological systems are functionally specific, context-sensitive informational variables; and that the coordination dynamics are more general than the particular structures that instantiate them.
Notice, coordination dynamics is not trapped (like ordinary physics) by its (purely formal) syntax. Order parameters are semantic, relational quantities that are intrinsically meaningful to system functioning. What could be more meaningful to an organism than information that specifies the coordinative relations among its parts or between itself and the environment? This view turns the mind-matter, information-dynamics interaction on its head. Instead of treating dynamics as ordinary physics and information as a symbolic code acting in the way that a program relates to a computer, dynamics is cast in terms that are semantically meaningful. (Kelso 1995, p. 145)
Let me connect these points to the autonomy perspective. As we have seen, from the autonomy perspective a natural cognitive agent—an organism, animal, or person—does not process information in a context-independent sense. Rather, it brings forth or enacts meaning in structural coupling with its environment. The meanings of an autonomous system’s states are formed within (informare) the context of the system’s dynamics and structural coupling. Therefore, if we wish to continue using the term representation, then we need to be aware of what sense this term can have for the enactive approach. Representational “vehicles” (the structures or processes that embody meaning) are temporally extended patterns of activity that can crisscross the brain-body-world boundaries, and the meanings or contents they embody are brought forth or enacted in the context of the system’s structural coupling with its environment.
Another way to make this point would be to say that autonomous systems do not operate on the basis of internal representations in the subjectivist/objectivist sense. Instead of internally representing an external world in some Cartesian sense, they enact an environment inseparable from their own structure and actions (Varela, Thompson, and Rosch 1991, p. 140). In phenomenological language, they constitute (disclose) a world that bears the stamp of their own structure. As Merleau-Ponty puts it, quoting Goldstein, in the passage cited earlier: “the environment emerges from the world through the being or actualization of the organism.” In the case of animal life, the environment emerges as a sensorimotor world through the actualization of the organism as a sensorimotor being. The organism is a sensorimotor being thanks to its nervous system. The nervous system connects anatomically distant sensory and motor processes, subsuming them in operationally closed sensorimotor networks. Through their coherent, large-scale patterns of activity these networks establish a sensorimotor identity for the animal—a sensorimotor self. In the same stroke, they specify what counts as “other,” namely, the animal’s sensorimotor world.
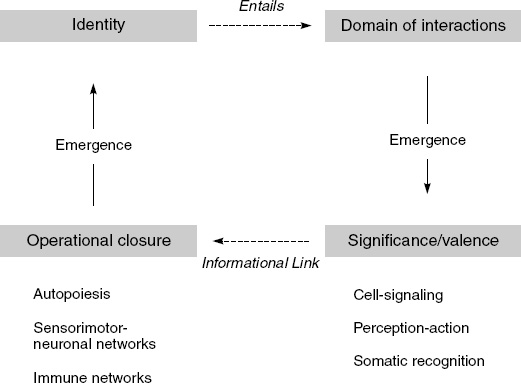
This idea of a sensorimotor world—a body-oriented world of perception and action—is none other than von Uexküll’s original notion of an Umwelt. An Umwelt is an animal’s environment in the sense of its lived, phenomenal world, the world as it presents itself to that animal thanks to its sensorimotor repertoire: “all that a subject perceives becomes his perceptual world and all that he does, his effector-world. Perceptual and effector worlds together form a closed unit, the Umwelt” (von Uexküll 1957, p. 6). The logic of this co-emergence is depicted in Figure 3.3.
In this figure, information is the intentional relation of the system to its milieu, established on the basis of the system’s autonomy (organizational-operational closure). One of the main scientific tasks for embodied dynamicism and the enactive approach is to explain how the pattern dynamics of brain, body, and behavior are informational in this sense (see Kelso 1995, p. 288).
Another key idea of the enactive approach that needs elaboration here is the idea of emergence or emergent processes. In complex systems theory, an emergent process is one that results from collective self-organization. An emergent process belongs to an ensemble or network of elements, arises spontaneously or self-organizes from the locally defined and globally constrained or controlled interactions of those elements, and does not belong to any single element. The enactive approach builds on this notion of emergence but reformulates it as “dynamic co-emergence,” in which part and whole co-emerge and mutually specify each other.
Let me first introduce emergence in the complex systems sense. A standard example of this kind of emergence is the formation of “Bénard cells,” a dynamic geometrical pattern of fluid flow, in liquids or gases subject to an imposed temperature gradient (see Kelso 1995, pp. 7–8; Solé and Goodwin 2000, pp. 13–17). The emergence of Bénard cells can be seen in the behavior of cooking oil in a frying pan. Applying heat to the pan increases the temperature difference between the cooler layer of oil at the top and the hotter layer of oil at the bottom. When the temperature difference between top and bottom is small, there is no large-scale or global motion of the oil, but eventually when the difference becomes large enough instability occurs and the liquid starts to roll in an orderly fashion known as convection rolls. In other words, the system undergoes a state transition, described mathematically as a bifurcation, as the new self-organizing behavior and spatial structures of convection rolls emerge. As the temperature gradient is increased still further, the convection rolls undergo another transition or bifurcation and give rise to an array of hexagonal up-and-down flow patterns called Bénard cells.
This example illustrates several basic points about collective self-organization and dynamic emergence. The temperature gradient is the control parameter for the transitions or bifurcations. It leads the system through a variety of possible states but does not dictate, prescribe, or code for the emerging flow patterns. Nor is there any homunculus or program inside the system determining those patterns. In Kelso’s words: “Such spontaneous pattern formation is exactly what we mean by self-organization: the system organizes itself, but there is no ‘self,’ no agent inside the system doing the organizing” (Kelso 1995, p. 8). The order parameter of the system is the amplitude of the convection rolls. It is created by the interactions of the fluid molecules, but at the same time it governs or constrains their behavior by drastically reducing the immense number of degrees of freedom of motion that the individual molecules would otherwise have.
Emergence through collective self-organization thus has two aspects. One is local-to-global determination, as a result of which novel macrolevel structures and processes emerge. The other is global-to-local determination whereby global structures and processes constrain local interactions. These global-to-local influences do not take the same form as the local-to-global ones: they typically manifest themselves through changes in control parameters (the temperature gradient in the example above) and boundary conditions rather than through changes to the individual elements (the fluid molecules). Coherent and ordered global behaviors, which are described by collective variables or order parameters, constrain or govern the behavior of the individual components, entraining them so that they no longer have the same behavioral alternatives open to them as they would if they were not interdependently woven into the coherent and ordered global pattern. At the same time, the behavior of the components generates and sustains the global order. This two-sided or double determination is known as circular causality (Haken 1983).
Emergence and circular causality are crucially important in the context of neurodynamics. Neuroscience indicates that cognition, emotion, and action require the transient integration of numerous, widely distributed, and constantly interacting brain regions and areas. An outstanding question for neuroscience today is to determine the mechanisms of this large-scale integration. From a dynamic-systems perspective, large-scale integration corresponds to the formation of transient dynamic links between widespread neural populations (Varela et al. 2001). On the one hand, large-scale dynamic patterns emerge from distributed local neuronal activities. On the other hand, large-scale patterns constrain these local activities. According to a number of theorists, dynamic instability or metastability is crucial to large-scale integration because it permits a flexible repertoire of global states without the system becoming trapped in any one particular state.10
Emergence and circular causality can also be illustrated by neurodynamical studies of epilepsy (Thompson and Varela 2001). Focal epileptic seizures originate in specific parts of the cortex; they can remain confined to those areas or spread to other parts of the brain. Their clinical manifestations depend on the cortical area in which they originate, how widely they spread, and how long they last. Local epileptic activity can modify the subject’s mental competencies and give rise to various kinds of mental experiences, such as visual or auditory illusions and hallucinations, and memory phenomena involving the vivid actual recall of a past event or déjà-vu illusions. These mental phenomena can also be induced by direct electrical stimulation of the temporal lobe in epileptic patients, as classically described by Wilder Penfield (1938). Thus local neuronal activity at the level of an epileptogenic zone can produce large-scale effects, eventually influencing the global level of a moment of consciousness. This is a case of local-to-global emergence in the brain.
The converse—global-to-local influence—though less documented and more controversial, seems also to be the case. The basic idea is that cognitive activity, which reflects large-scale integration in the brain, can affect local activity. For example, the subject can voluntarily affect local epileptic activity, as indicated by numerous patient reports and a few clinically reported cases (see Le Van Quyen and Petitmengin 2002). As long ago as 1954, Penfield and Jasper described the blocking of a parietal seizure by the initiation of a complex mathematical calculation (Penfield and Jasper 1954), and recently more extensive observations have confirmed such cognitive influences (Schmid-Schonbein 1998). We can assume that such intervention is possible because the epileptogenic zones are embedded in a complex network of other brain regions that actively participate in the large-scale integration underlying cognitive acts. It also seems reasonable to assume that these global patterns of integration can influence local events, including eventually the local epileptogenic zones, whose activity can thus be taken as an indication of the global influence.
Experimental work by Michel Le Van Quyen, Francisco Varela, and their colleagues provides evidence for such global-to-local influence in the case of a patient with an unusually focal and stable occipitotemporal epileptic discharge.11 This patient showed no evidence of cognitive impairment and was willing to participate in simple cognitive tasks of visual and auditory discrimination. For the visual task, he was asked to press a button when the target stimulus appeared, but not when the two other nontarget stimuli were shown. The temporal intervals between successive discharges of the epileptic spike pattern were analyzed. Dynamical inspection (in the form of a phase space known as a first-return map) showed that the distribution of the intervals followed a particular kind of unstable dynamic pattern. The spikes displayed a distinct periodic activity for a short time before they diverged away along another unstable direction, a kind of dynamic pattern known as an unstable periodic orbit. Furthermore, this activity covaried with the specific mental state of the patient during the perceptual task and appeared to be modulated by the gamma frequency (30–70 Hertz) activity associated with his cognitive states. (Gamma frequency activity is widely reported to be associated with a variety of cognitive processes, including attention, perception, and memory.) These findings suggest that the patient’s act of perception contributed in a highly specific way to “pulling” the epileptic activities toward particular unstable periodic orbits. Such global-to-local influence mobilized by cognitive activity might open up possibilities for cognitive strategies of control of epileptic seizures (Le Van Quyen and Petitmengin 2002).
Let me conclude this chapter by linking these ideas about circular causality and emergence back to autonomy. An autonomy perspective brings with it a certain way of thinking about emergence. What emerges in the case of an autonomous system such as a cell is a self-producing entity that also brings forth its own domain of interactions (see Figure 3.3). This sort of emergence takes a major step beyond dynamic pattern formation in physical dissipative systems:
[A]lthough the phenomenon of self-organization always involves the generation and maintenance of a global (or high-level) pattern or correlation that constrains the (low-level) dynamics of the components of the system, in standard dissipative structures this occurs only provided that the system is put under the appropriate boundary conditions. If those (externally controlled) conditions are changed (in particular, if the input of matter or energy is outside a certain range), the self-organizing dynamic vanishes. Therefore, there is an important difference between the typical examples of “spontaneous” dissipative structures and real autonomous systems: in the former case, the flow of energy and/or matter that keeps the system away from equilibrium is not controlled by the organization of the system (the key boundary conditions are externally established, either by the scientist in the lab or by some natural phenomenon that is not causally dependent on the self-organizing one), whereas in the latter case, the constraints that actually guide energy/matter flows from the environment through the constitutive processes of the system are endogenously created and maintained. (Ruiz-Mirazo and Moreno 2004, p. 238)
An autonomous system, such as a cell or multicellular organism, is not merely self-maintaining, like a candle flame; it is also self-producing and thus produces its own self-maintaining processes, including an active topological boundary that demarcates inside from outside and actively regulates interaction with the environment. In the single-cell, autopoietic form of autonomy, a membrane-bounded, metabolic network produces the metabolites that constitute both the network itself and the membrane that permits the network’s bounded dynamics. Other autonomous systems have different sorts of self-constructing processes and network topologies. Whether the system is a cell, immune network, nervous system, insect colony, or animal society, what emerges is a unity with its own self-producing identity and domain of interactions or milieu, be it cellular (autopoiesis), somatic (immune networks), sensorimotor and neurocognitive (the nervous system), or social (animal societies).
Dynamic co-emergence best describes the sort of emergence we see in autonomy. In an autonomous system, the whole not only arises from the (organizational closure of) the parts, but the parts also arise from the whole.12 The whole is constituted by the relations of the parts, and the parts are constituted by the relations they bear to one another in the whole. Hence, the parts do not exist in advance, prior to the whole, as independent entities that retain their identity in the whole. Rather, part and whole co-emerge and mutually specify each other.
Biological life, seen from the perspective of autopoiesis, provides a paradigm case of dynamic co-emergence. A minimal autopoietic whole emerges from the dynamic interdependence of a membrane boundary and an internal chemical reaction network. The membrane and reaction network (as well as the molecules that compose them) do not preexist as independent entities. Rather, they co-emerge through their integrative, metabolic relation to each other. They produce and constitute the whole, while the whole produces them and subordinates them to it. We will come back to this paradigm case of dynamic co-emergence in Part II.