In 1965, a mat of seaweed broke away from Iceland’s brutal southern shore and began to make for the wide, open Atlantic Ocean. But a couple of days later, this drifting vegetation hit land—new land—a small volcanic island that had started to emerge from the waves a couple of years earlier. Surtsey was still rife with volcanic activity; yet the seaweed found itself lodged in the volcanic debris on the island’s north shore. In the carpet of organic matter, there was a seed of sea rocket, which settled into the virgin soil, germinated and began to grow. The appearance of this flowering plant, so soon after the formation of this island and whilst eruptions were still ongoing, is remarkable.
The botanists keen to witness the colonisation of a virgin volcano were certainly excited. Soon after the first eruption, they recorded the presence of bacteria, fungi and more algae, many presumably arriving on the wind as spores from Iceland itself some 40 km to the north. By the time the sea rocket rocked up, there was an ashy soil-like substrate to welcome it. Within a few years, other plants like sea sandwort and lyme grass had become established and started to flower. It makes sense that all of these early settlers should have been durable, able to survive the hazards of transport and the extraordinarily hostile landscape that awaited them.
So it was for the Galápagos too. Some of the first arrivals were likely windborne spores of bacteria, fungi, algae and lichens, carried across hundreds of kilometres of open water. These early settlers are particularly hardy, able to lodge on bare rock that most other species could not dream of calling home. They all contribute to the weathering of volcanic lava flows, bringing on the formation of soil-like substrates that give plants a realistic chance of survival.
As with the geology of these islands, our knowledge of Galápagos plants really starts with Darwin. ‘Amongst other things, I collected every plant, which I could see in flower, & as it was the flowering season I hope my collection may be of some interest to you,’ he wrote from Sydney, Australia, to his old mentor John Stevens Henslow in Cambridge. ‘I shall be very curious to know whether the Flora belongs to America, or is peculiar.’ With only limited botanical expertise, he needed help.
Henslow made a token effort, describing a couple of Darwin’s Beagle cacti, one from Patagonia and the other from the Galápagos. He probably felt that this would satisfy his young friend. Yet Darwin had a far more ambitious goal: explaining where his Galápagos plants had come from. It was a project that, taken to its logical conclusion, would ultimately reveal the origin of species.
Darwin had collected sufficient specimens in the Galápagos to hit upon two key ideas. First, he realised that much of the Galápagos fauna had a distinctly South American flavour to it. Second, the species on one island seemed to differ slightly from those on the next. ‘It never occurred to me, that the productions of islands only a few miles apart, and placed under the same physical conditions, would be dissimilar,’ admitted Darwin in the first edition of his Journal, published in 1839.
His impressive collection of Galápagos plants proved to be a crucial test of this observation. So in 1838, he wrote again to Henslow to give him a prod. ‘I do not want you to take any trouble in giving me names &c &c—all I want is to know whether in casting your eye over my plants, how many cases . . . there are of near species, of the same genus;—one species coming from one island, & the other from a second island.’
When Henslow again failed to produce the goods, Darwin got him to send the languishing Beagle specimens to up-and-coming botanist Joseph Dalton Hooker, who had recently returned from his own voyage of discovery to the Antarctic. Within days of receiving the Galápagos plants towards the end of 1843, Hooker wrote to Darwin in great excitement. There were indeed remarkable differences from one island to the next. In his initial analysis, Hooker judged that Darwin had handed him 217 different plant species, more than half of which were peculiar to the Galápagos. Of these so-called endemic species, the vast majority seemed to be confined to just one island. This fact, wrote Hooker, ‘quite overturns all our preconceived notions of species radiating from a centre.’
Darwin, in turn, was ecstatic. ‘I cannot tell you how delighted & astonished I am at the results of your examination; how wonderfully they support my assertion on the differences in the animals of the different islands.’ So powerful was the case of the Galápagos plants that Darwin gave them a prominent place in the beefed-up second edition of his Journal, which appeared in 1845. ‘Reviewing the facts here given, one is astonished at the amount of creative force, if such an expression may be used, displayed on these small, barren, and rocky islands.’
Seeding the Galápagos
This was all very well, but it still didn’t tie up how the species had got there in the first place. The journey from South America to the Galápagos might be easy for us humans, catered for as we now are by more than forty flights a week. But for a plant it’s a migration that presents a series of formidable challenges. Hooker mapped out several possible routes that bits of vegetation or seeds might have taken: by sea, by air, carried by birds or brought by humans.
The same strong current that carried the bishop of Panama to the Galápagos in 1535 will often bring miscellaneous vegetation from the continent. This much was noted more than two hundred years ago by James Colnett: ‘On several parts of the shore, there was drift-wood, of a larger size, than any of the trees, that grow on the island: also bamboos and wild sugar canes, with a few small cocoa nuts at full growth, though not larger than a pigeon’s egg,’ he wrote in 1797. Beagle captain Robert FitzRoy also noted the way the driftwood was always to be found high and dry on the south-western shores of the islands.
Still, 1,000 km by sea? It’s a heck of a way. Although Hooker observed that many of the Galápagos plants boasted large, tough seeds, something he figured would ‘probably aid them in resisting for some time the effects of salt water’, he remained sceptical that seeds could really withstand such a journey.
Darwin decided to find out and in 1855 began to play around with seeds and seawater at his home in the English countryside south of London. He kicked off with cress, radish, cabbage, lettuce, carrot, celery and onion, leaving seeds in saltwater for a week before planting them out. They all germinated, though some more convincingly than others. ‘It is quite surprising that the Radishes shd have grown, for the salt-water was putrid to an extent, which I cd not have thought credible had I not smelt it myself,’ he wrote. He went further, buying seeds of all manner of different species and gradually extending the length of time they were immersed in his seawater concoction. By the time he published On the Origin of Species in 1859, Darwin had exposed the seeds of eighty-seven different plant species to these hostile conditions. Incredibly, nearly all of them germinated after weeks stewing in brine; some still seemed in perfectly good working order after several months.
So seeds floating on the waves could easily account for the arrival of plants in the Galápagos. But in the course of his seed-salting experiments, Darwin hit upon another serious obstacle to this mode of travel. Most of the seeds he dunked sank immediately, and only a few bobbed back up to the surface. It made sense then that species with floatable seeds would stand a much better chance of taking a long-distance oceanic cruise from South America to the Galápagos.
Darwin hauled A. K. Johnston’s Physical Atlas from his bookshelf, from which he worked out the average speed of several Atlantic currents. Based on all his observations, he concluded that ‘the seeds of 14/100 plants belonging to one country might be floated across 924 miles of sea to another country’. The current that connects continental South America to the Galápagos can run considerably faster than Darwin’s Atlantic average, often around 100 km per day. So a back-of-the-envelope calculation suggests that a buoyant seed could reach the archipelago in just over a week.
More plants—even those with sinking seeds—might arrive by hitching a ride on a clod of earth attached to the roots of some big tree or carried in the carcass of a dead animal. To illustrate this possibility, Darwin fed a pigeon on seeds that would normally be ‘killed by even a few days’ immersion in sea-water’. He then sacrificed it and floated its body on salty water for a month. To his great surprise (and, one imagines, satisfaction), the delicate seeds, once dissected from the pigeon’s crop and planted out, ‘nearly all germinated’.
The Coastal Zone
Even if a floating seed does reach an island, it will face an immediate landscape that, for many, will be just too salty. It takes some nifty adaptations to make it in the coastal zone.
Plants can do this in a number of ways, some of them better suited to becoming established along the Galápagos shoreline. For a red mangrove, for instance, its roots are crucial. These drop down into the seawater, acting like props to keep the trunk out of the water, and are so specialised that they can draw water from the sea without bringing too much salt on board. The leaves of black mangroves also contain salt glands capable of shifting salt from the inside to the outside of the plant. Mangroves and many other coastal species also have dark, waxy leaves, which reflect a lot of sunlight, preventing overheating and reducing unnecessary evaporation of precious water from the tissues.
Surviving is all well and good, but if you want to colonise, then you really have to reproduce. If, as was likely the case for most new arrivals in the Galápagos, you find yourself just one of a handful (or possibly the only one) of your species, the more self-sufficient your mode of reproduction the better. If you have a fussy reproductive set-up, like a flowering plant that relies on a special insect to carry pollen to a fellow member of your species, you could be in trouble. This helps explain why the vast majority of flowering plants in the Galápagos are capable of self-pollination and, with no need to attract insects, not particularly showy (white or yellow petals are the norm). Choose a Galápagos flower at random. Take a peek at its reproductive parts. It’s probably a hermaphrodite, sporting both male and female apparatus. This is the case for the red mangrove (named for its reddish bark rather than its yellow flower). Unusually, it is viviparous too, which means that the seeds begin to sprout before they are released from the parental tree. When these so-called propagules finally drop, they can survive in salty water for more than a year, obviously useful if they find themselves washed out to sea.
The Arid Zone
Alongside the seed-floating route to the Galápagos, Hooker imagined other modes of transport too. Owing to ‘the excessive minuteness’ of the spores of lower plants like mosses and ferns, he was in little doubt ‘that their diffusion by the winds is a never-ceasing though invisible operation’, just as on the Icelandic island of Surtsey. Birds, he suggested, could be important too.
Darwin agreed. ‘Living birds can hardly fail to be highly effective agents in the transportation of seeds,’ he wrote in the Origin. Obvious as it might be, he still sought evidence, going out into his garden in search of bird droppings. ‘In the course of two months,’ he wrote, ‘I picked up in my garden 12 kinds of seeds, out of the excrement of small birds, and these seemed perfect, and some of them, which I tried, germinated.’ In addition, the occasional seed might get stuck to a foot or a beak. Although some Galápagos plants clearly came on ocean currents and others on the wind, it’s thought that most Galápagos plants probably hitched a ride with birds.
If a settling seed is fortunate enough to land beyond the salty coastal region, it could still be in for a shock. The lowest reaches of each Galápagos volcano are dominated by the arid zone, where water—or the lack of it—is the perennial problem. It can be unbelievably hot too. On Santiago, Darwin stuck a thermometer into some brown soil, and the mercury rocketed up to almost 60°C. It could have been yet still hotter, but Darwin’s thermometer only went so far.
There are several ways to cope in this situation. As water escapes through a plant’s leaves, those species with fewer, smaller leaves and fewer pores that open up to the atmosphere are going to be able to survive longer in an arid environment. This is why Darwin found the plants beyond San Cristóbal’s shoreline to be ‘such wretched-looking little weeds’, the landscape ‘covered by stunted, sun-burnt brushwood’. From a distance, this appeared ‘as leafless as our trees during winter’.
Herman Melville found the plant life more wretched still. ‘On most of the isles where vegetation is found at all, it is more ungrateful than the blankness of Atacama,’ he wrote in the first of his literary sketches on ‘the Encantadas’. Whereas Darwin eagerly collected specimens to be pressed, Melville only saw ‘tangled thickets of wiry bushes, without fruit and without a name, springing up among deep fissures of calcined rock and treacherously masking them’.
Many of these arid-dwelling species have interesting roots. Take the prickly pear cacti of the genus Opuntia, for example. These have two kinds: one superficial set of finer roots whose job it is to suck up every last drop of water in the aftermath of a downpour and a deeper ‘tap’ root, which gives it a stronger hold on the rocks and searches out deeper sources of water. The thorny Acacia bushes have also gone in for this approach, with a root network that grows during drought to cover a greater area of soil. Some of this spiky vegetation will be festooned with curtains of lichen, fine filaments that are able to trap moisture from the atmosphere. This then falls in steady drips to the earth around the tree.
Another approach is to specialise in water storage. This is something that cacti are famous for. The lava cactus is notable for its ability to establish on fresh lava flows, and specimens can be seen in Sullivan Bay on Santiago, in Punta Moreno on Isabela and around the rim of Genovesa. It’s a stout cactus that comes in clusters. Its leaves have been so modified that we call them spines, a perfect way to cut down on water loss. This species is incredibly slow growing and doesn’t often blossom. When it does, the window for pollination is just a matter of hours; then the petals shrivel, and the flower drops.
Compared with the lava cactus, the prickly pear opuntias are a much more common sight. It’s well known that giant tortoises are long lived, and many visitors to the Galápagos often imagine that one of the tortoises Darwin saw back in 1835 might still be alive today, that they have seen one and the same animal. Although this seems vanishingly unlikely, it could certainly be the case for some of the biggest Galápagos opuntias. It’s reckoned that these take around fifty years to reach maturity and start to produce flowers and fruits. They can then probably do so for a couple of hundred years.
In the Galápagos, there are six different species (with a couple further subdivided into distinct varieties). All of them have flattened oval branches (or pads as we tend to call them) that lobe off a central trunk and store the water. As with the smaller lava cactus, the leaves are no more than spines that stick out from the pad, considerably reducing loss of water.
Apart from these general similarities, there are plenty of differences between species. Darwin collected just one, a prickly pear opuntia specimen from Santiago, the one Henslow described back in 1837. But he would have been fascinated to have had a chance to look at the other forms that exist throughout the archipelago. The most obvious difference between species is their size. The Santiago species, at around 2m high with a trunk some 30 cm wide, turns out to be somewhere in the middle of the opuntia range. The northern islands of Darwin, Wolf and Genovesa are home to one of the smallest varieties, a low-lying form in which the trunk is largely obscured by a riot of pads. On Santa Fé, by contrast, there is a colossal species that can reach up to 12m high with a trunk that’s over 1m wide.
It’s often said that the different heights have come about as a result of the selective force imposed by the local herbivores. So on islands that have never had tortoises (like Darwin, Wolf and Genovesa, for instance), the local prickly pear opuntia is spread out near to the ground. On islands with tortoises, by contrast, the opuntias seem to have found a way to grow their succulent pads out of the reptiles’ reach. Though possible, this explanation is probably a bit too simplistic. Although Santa Fé once had tortoises, would they really have driven the local opuntia to such a giant extreme?
The spines of the different Opuntia species also present something of a puzzle, one that would have intrigued Darwin. These differ markedly in number, length and strength. On Santiago, for instance, the fleshy pads are covered with clusters of between five and thirty-five spines that can measure up to 7.5 cm long. Henslow described these as ‘strongly resembling hog’s bristles.’ On Genovesa, by contrast, the spines come in clusters of between seven and twenty-eight and are shorter and softer, more like sun-bleached hair. It’s been suggested that these wavy spines have appeared on islands where there is no herbivorous threat; the plants just don’t need the same defences they used to. Alternatively, soft spines may make it easier for birds like the cactus finch to come for pollen and nectar. On an island like Genovesa with few insect pollinators, such bird-mediated pollination could be particularly important.
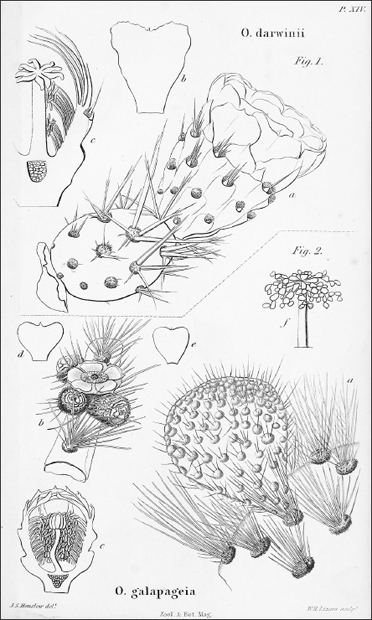
FIGURE 4.1. The prickly pear cactus Opuntia galapageia (Figure 2). Charles Darwin collected this species from the lowlands of Santiago. It is clearly different from another Opuntia he collected from mainland South America (Figure 1). Reproduced from John Stevens Henslow, Magazine of Zoology and Botany (1837): 466–468.
If times get really tough, it’s always possible to shut down completely. The prickly pear opuntias can do this by dropping pads, a strategy employed by another common arid-zone species, the palo santo tree. In the cool season from June to December, the palo santo is not in leaf, and its silvery lichen-clad bark shines out from the dark volcanic backdrop. When the hot season hits and rain begins to fall, its leaves begin to sprout, and the scattered stands become a much greener affair. The same goes for the deciduous guayabillo, pega-pega and matazarno that can be found further up, as the arid zone transitions into the moister highlands.
The Highlands
Given the challenges imposed by the coastal and arid zones, it seems extraordinary that plants should have made it any further. But with the aid of wind and birds (and indeed humans), plants can get to most places, and those islands that rise more than a few hundred metres above sea level boast some of the choicest plant habitats to be had.
During the cool season, between June and November, the garúa hovers at between 500m and 1,000m above sea level. This creates a lush moist zone, overrun with effulgent flora of an altogether different kind. Driving from Puerto Ayora up the road that cuts its way across Santa Cruz towards Baltra, this transition is quite clear. In and around the town, it’s hot, it’s dry and the vegetation is sparse. At the small town of Bellavista at around 200m above sea level, everything will have changed. There is no sign of lava any more. It is there, of course, but buried beneath a choking quilt of purest green. This is why Bellavista (and the smaller farming community of Santa Rosa) exists at all: the climate here is perfect for growing crops and grazing cattle. Travelling higher still, one finds a misty, luxuriant landscape carpeted with grass and sedge (see Appendix C, Figure 5).
When Darwin experienced this transition, it struck him too. Landing at Black Beach on Floreana (where the small town of Puerto Velasco Ibarra now stands), he and FitzRoy were met by the vice governor of the islands, Nicholas Lawson, who then walked them around 8 km inland to the small penal colony he was supervising on behalf of the Ecuadorian government. ‘The wood gradually becomes greener during the ascent,’ Darwin wrote in his diary. ‘Passing round the side of the highest hill; the body is cooled by the fine Southerly trade wind & the eye refreshed by a plain green as England in the Spring time.’ After the barren landscapes of Peru and Chile, he was thrilled ‘to find black mud & on the trees to see mosses, ferns & Lichens & Parasitical plants adhaering.’
He was in Scalesia forest, a habitat named after the giant daisy tree Scalesia pedunculata. This is a species that would normally dominate the vegetation at this altitude, though it—and the wider habitat—has been severely affected by clearance for agriculture and pressures from introduced species. On Santa Cruz, the visitor site known as Los Gemelos boasts a relatively intact patch of highland forest, the upstanding S. pedunculata standing guard above the turmoil of vegetation tumbling into the depths of these collapsed twin craters. The trees grow incredibly quickly, dwarfing a two-story house within a matter of years as they shoot up in search of the sun. Their leaves cluster at the skyward tips of the branches to form a kind of canopy, and just beneath is a crowded orgy of mosses, liverworts, orchids, and passion flower, all using the trunk and branches as a kind of ladder. There are other trees here, including Galápagos pisonia, the Galápagos guava and cat’s claw, though they struggle against the dominant position that S. pedunculata has assumed.
At around five years old and about 8m tall, the giant daisy tree will begin to flower. Tiny white petals emerge on top of the canopy. The seeds will fall to the forest floor, but in this shady environment they will not germinate until their parent plants have come to the end of their fast, short lives at about age fifteen. Then, as the canopy starts to die back and sunlight streams to the floor, the seedlings and saplings emerge, all racing together to replace their parents as if in some kind of shared adventure. Between each unfolding canopy, the trees maintain daylight, a gap of around 30 cm. If I may indulge in a touch of anthropomorphism, it is as if each specimen is acutely aware of its neighbour’s sensitivities, the forest a paragon of mutual respect.
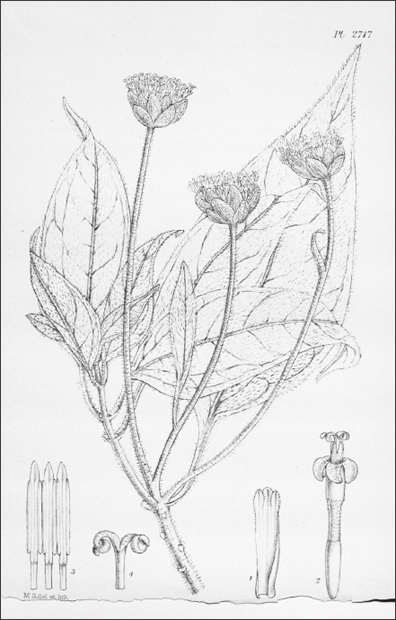
FIGURE 4.2. The giant daisy tree Scalesia pedunculata. This is just one of at least fifteen closely related species found only in the Galápagos, but it is certainly the most spectacular, a slender tree that grows rapidly to a height of some 20m. In the past, this species would have dominated much of the highlands—‘a very handsome species’, according to botanist Joseph Hooker. Reproduced from Joseph Hooker, Icones Plantarum 28 (1905): pl. 2717.
Scalesia pedunculata is not just a pretty tree; it is also part of a fascinating evolutionary story. For it’s not the only Scalesia in town. In fact, the genus comprises at least fifteen different species (some of which are further divided into subspecies), all of them endemic to the Galápagos and found from the arid zone to the highlands. In spite of considerable differences in outward appearance, its thought that all these species are probably descended from a single daisy-like ancestor that managed to establish in the arid zone. From this unpropitious origin, subsequent generations spread higher and higher, adapting to the increasing humidity along the way and dispersing to other islands. Small, low-lying islands tend to support one shrubby Scalesia species. Larger, elevated islands have more. On Santa Cruz, for instance, there are at least six different species.
Beyond the Scalesia forest, on established islands like San Cristóbal and Santa Cruz that rise above 400m, there are more rarefied habitats still. There are no Scalesia here. They give way to lower-lying, bushy thickets of Miconia (the nesting site for the critically endangered Galápagos petrel), then, even higher, to the pampa zone, a boggy moor-like expanse of grass, sedge, moss and fern.
All this vegetation, from the salty coast, through the desert-like arid zone and the humid highlands, to the chilly, thinned-out pampa, provides the backdrop for the arrival of land-dwelling creatures from tiny insects to hulking reptiles to humans. It is to them that we now turn.