Animal Hearing and Sight
“Bumblebee bat, how do you see at night?—I make a squeaky sound that bounces back from whatever it hits. I see by hearing.”
—Darrin Lunde, entomologist and author
Because hearing and balance involve the inner workings of the ear and are closely related in how they detect and transmit information to the brain, it is logical to discuss them together. But talk about a Rube Goldberg apparatus! The inner ear, where the structures that handle both of these senses are found, is astonishingly intricate and convoluted, with many moving parts. If this structure from many different vertebrates were shown to an engineer, there would probably be no way she or he would figure out what it was for, which is a reminder that evolution does not produce perfect or even logical structures in organisms. But one feature would be certain: the engineer would recognize that they all have the same basic parts and so are modifications of one basic structural apparatus.
If the engineer were then shown insect hearing organs, she or he would be even more confused, because the hearing apparatus of these animals more than likely evolved many times independently, yielding structures that don’t seem to have commonality at all even though they all have the same function. The ancestor of all insects was deaf, and the story of insect ears is about converting an existing structure in the insect body to a hearing organ.
But why would an insect want to hear in the first place? After all, hundreds of thousands of species of insects living today don’t have hearing apparatuses and so don’t really hear. For instance, of the 350,000 known beetle species, only a small proportion have hearing. It is evident through evolutionary analysis that hearing arose independently in many groups of insects at about sixty-five million years ago. Something big must have triggered this shift in sensing the world in insects. In one word, it was bats.
Bats hunt by a process called echolocation. In fact, some cave-dwelling birds and some marine mammals also use echolocation to “hear” their surroundings. In echolocation, an animal makes a sound and then literally listens for the echo of the sound. By measuring the time the sound takes to bounce back to the ears, the echolocating animal can get a good idea of whether there is something else out there, and if so where it is. Because the animal has two ears, this makes for an efficient sound reception system—the animal can actually hear differences in the two ears and use them to further refine where objects are, including flying insects. So, what an insect with hearing does is intercept the sound from the echolocating bat and use that information to attempt to avoid the predator. Bats and flying insects have engaged in an ever-escalating arms race over the past sixty-five million years with respect to echolocation and ears. Oh, and of course, as with most things in nature, hearing in some insects has been involved in enhancing copulation.
There are two major groups of bats: microbats (Microchiroptera) and megabats (Megachiroptera). The Megachiroptera include the fruit bats and other relatives and, except for one species, do not echolocate to find food. The one species of Megachiroptera that does echolocate uses a short-click vocalization that is very different from the vocalization used in echolocation by microbats.
A microbat echolocates by emitting high-frequency sound waves from its larynx that range between 14,000 and 100,000 hertz and are then broadcast from its mouth or its nose. The more vibrations per second, the higher the frequency. Humans perceive sounds in the range of 20 to 20,000 hertz, so we can hear only some bat echolocation screams. Microbat vocalizations are also somewhat species specific, because in adapting to a particular habitat a bat will also adapt the echolocation frequency for the environment. Even though some bat species overlap in the frequency of their echolocation calls, researchers have developed echolocation call libraries for bats much like the bird-call libraries used by ornithologists.
But how does an organism develop ears? One way is through the slow but steady accumulation of change by which Charles Darwin thought all life evolved. This process of imperceptible change was articulated in On the Origin of Species as a universal principle for how life evolved.
Darwin was smitten by the work of the geologist Charles Lyell, and Lyell’s Principles of Geology was one of the books Darwin took with him on the voyage of HMS Beagle. Lyell made clear that changes in the Earth’s geology, such as the emergence of mountain ranges and erosion, were best described as gradual, and Darwin felt likewise, that change in living organisms over evolutionary time was also gradual. This concept of gradualism was taken up by early evolutionary biologists and dominated the way evolutionists thought about change in the organic world until the 1970s, when Niles Eldredge and Stephen Jay Gould suggested that evolution might occur in a punctuated fashion. The second mechanism conjures up the “hopeful monster.” This is a term first used by the nineteenth-century zoologist Richard Goldschmidt and resurrected by Gould in the 1970s to explain the appearance of novelties in the divergence process. We have already discussed one potential hopeful monster in Chapter 3—the Cyclops—which, although it is monstrous, might not be so hopeful as a survivor in nature. One problem with hopeful monsters is that, although large-scale changes in traits do occur, they are usually accompanied by lethality in the individuals with the mutation for the change. This side effect, which is very common in experimental genetics and developmental biology, occurs because genes usually interact in more than a single pathway of development. This phenomenon is called pleiotropy and is another evolutionary pathway that organismal life can take to generate novel structures and behaviors. So, although a mutation in a gene might produce a hopeful monster in one pathway, it will be lethal in the other pathway or pathways that it affects. How hearing and balance evolved in insects is a beautiful story of evolution at work in the realm of hopeful monsters and pleiotropy.
The ancestors of hearing insects, and all insects for that matter, probably had pretty good balance. Part of balance in insects is accomplished with what are called chordotonal organs, small patches of neural cells that communicate to the brain how stretchy the tissue between body or appendage segments is. By judging how much two adjacent segments are stretched relative to each other, the insect can determine where its body is in space. This is a basic proprioceptive mechanism that insects need for efficient locomotion. The chordotonal organs are made up of mechanoreceptor cells that also react to vibrations and hence were preadapted or exapted (see Chapter 4) to be able to detect sound of different frequencies. Anywhere a junction of segments of structures exists, a chordotonal organ is usually placed as a proprioception organ to assist in balance. What this means is that the potential for the development of an ear exists in bizarre body parts with this kind of proprioception, such as on the abdomen and legs. And in fact, it appears that evolution has taken advantage of converting as many of these primordial chordotonal organs as possible into ears.
Another possible evolutionary mechanism for ears to evolve is, as I mentioned above, through exaptation. Some insects have exapted the chordotonal organs of their antennae to make ears, and some have also used these organs in their mouthparts. Different groups of insects have accomplished this exaptation independently about ten times. Some insects have the antennal version of the ear. These are mostly diptera, like flies and mosquitoes, where the proprioception organ, called Johnston’s organ, has been converted to a hearing organ to interpret wingbeat sounds in members of their species. Wingbeats are an important part of courtship in these dipterans. Other insects have used the chordotonal organ hinges in their wings and legs and converted these to ears. Finally, the area between adjacent segments has chordotonal organs, too, and these have been converted into ears several independent times. In some insects these organs produce ears on the abdomen of the insect called tympanal ears. Most of the time a single pair of chordotonal organs is transformed (one on each side of the insect), but in the bladder grasshopper Bullacris membracioides, six chordotonal organs on each side of the insect have been converted into ears, making for twelve abdominally located tympanal ears.
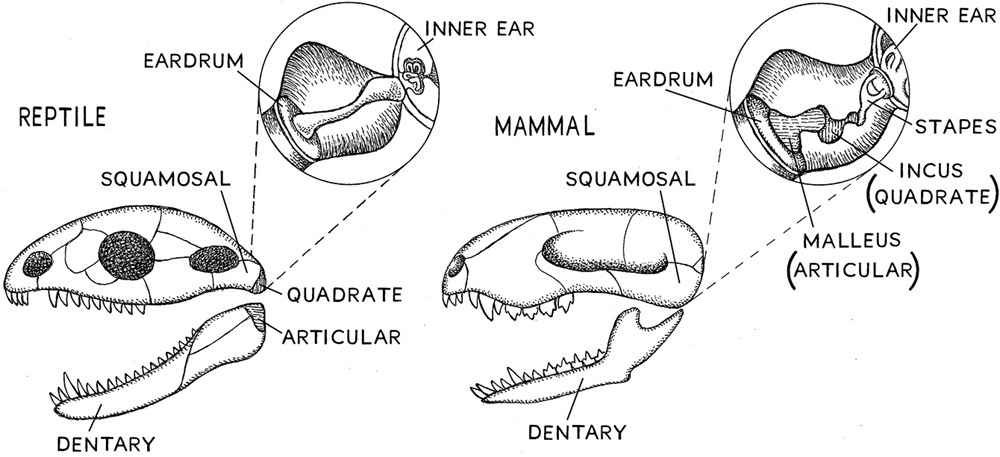
It is more than likely that the vertebrate ears with which we are so familiar arose only once (thus evolution works one way in one group of organisms and another way in other groups of organisms). Vertebrates have been around for more than five hundred million years and insects for about four hundred million years. If the evolutionary process was playing on a level surface in both groups, then we might expect ears to arise independently about the same number of times in the two lineages. But ears arise ten times more frequently in insects than in vertebrates. Understanding why vertebrate ears have stayed so static requires following five bones near the jaw and side of the head through the evolution of vertebrates.
Let’s follow the arrangement of these bones in three kinds of organisms (bony fish, reptiles, and mammals) and in so doing learn how traits like the jaw and the ears can change over time. To make the exercise fruitful, we need to consider which structures are ancestral and which have evolved—so we need what evolutionary biologists call an outgroup. Two pretty good outgroups for looking at bony fish, reptiles, and mammals are the primitive-looking hagfish and lampreys. The five bones to follow are the dentary, articular, squamosal, quadrata, and stapes bones. Because the bones have changed in the evolution of fish, reptiles, and mammals, the inner ear of these three organisms contains different structures. The inner ear also contains the balance organs, which are pretty well developed in fish, so we will also bring hagfish and lamprey balance organs into the discussion.
Hagfish and lampreys do not have jaws, but they do have inner ear vesicles. They feed primarily by grasping prey with their tongue or by attaching to their food item, so they get by without jaws fairly well. The anatomy of their heads with respect to the five bones is pretty simple. Since they don’t have a jaw, they either don’t have the five bones we are following, or it is too difficult to see them, and hence we simply cannot make a statement about their existence. The inner ears of these two fish have fairly simple structures, which suggests that the common ancestor of jawless and jawed vertebrates had an inner ear. But because the bones of the heads of these two primitive-looking organisms don’t look like any of the bones in jawed vertebrates, all we can say is that the inner ear is an ancestral characteristic of both jawless and jawed vertebrates. It is in this inner ear area where both sounds and balance are managed.
The balance organs in the inner ear exist in the outgroups but in a modified form. The inner ear of a hagfish has only one semicircular canal, a tubelike structure responsible for balance. Near the semicircular canal in the inner ear of a hagfish resides a patch of neural cells that have hairs on them that act as balance organs. The lamprey has two semicircular canals and also patches of neural cells with hairs on them that act as the mechanism for mechanoreceptors in balance. More advanced vertebrates (bony fish, reptiles, and mammals) have three of these canals arranged like the three axes (X, Y, and Z) of a three-dimensional graph. That the semicircular canals are involved in balance is well known, and as discussed previously with respect to primates, a sense of balance is critical for survival. In some mammals, balance was not critical, and the evolution of the inner ear in these organisms presents biologists with yet another important proof of the occurrence of evolution and its importance in the natural world (box 5.1).
Fish attain balance by yet another structure called the lateral line. This balance system, found in most fish, is used to detect motion and vibrations in the external environment. The lateral line is made up of cells on the skin that are called hair cells. Motion or vibrations displace the hairs on these cells of the lateral line, and such displacement is converted into a neural signal to the brain through a mechanoreceptor mechanism. The lateral line is exactly what its name suggests—a line of hair cells extending from the gill covers to the tail. Although most fish use the lateral line hair cells to detect vibrations around them, the evolutionary process has modified the lateral line hair cells in many species to detect electrical impulses, too. So, in addition to detecting motion or vibrations around the body, these lateral line cells detect electrical impulses from other animals too. Of course, for these modified electroreception cells to be useful they need an electrical field to detect. So, there are two ways such fields are produced. First, some fish use these receptors to detect electric fields that they themselves produce. In a process called electrolocation, these fish use the hair cell organs to detect objects, somewhat the same way that echolocation works, only with electrical fields. Second, all organisms passively produce the other source of electric fields, because the nervous system of any organism is essentially producing electrical impulses all the time. Fish with electroreceptors use this passive source of electrical fields to locate prey and predators.
Electroreception is not unique in the animal world, because it appears to have evolved several times. Perhaps the most notable case of this convergence can be found in the duck-billed platypus. This strange mammal, a member of a small group of mammals called monotremes, has an electroreception system that did not evolve from the lateral line but instead developed from cells in skin glands that have been converted through the evolutionary process into cells with free nerve endings. Monotremes in general have electroreception, but the platypus is the most impressive, with forty thousand of these modified skin cells arranged along the bill as stripes. Platypuses are pretty good hunters, and more than likely these electroreceptors assist in their acute hunting ability. Other vertebrates with electroreceptor capacity include some dolphins, and the anatomical origin of these electroreceptors is independent of the lateral line and also the monotreme adaptation.
BOX 5.1 | VESTIGIAL BALANCE IN SLOTHS
By studying mammals that rely little on balance, researchers have examined the plasticity or expendability of the semicircular canals. One group of mammals that do not use balance to any great degree are the three-toed sloths. These mammals move extremely slowly, and as the authors of a paper examining their semicircular canals state, “Significant travelling distances, speed and agility are not part of their locomotor repertoire.” Sloths do descend their tree top perches once in a while to defecate, but overall they do not need an acute sense of balance. In defense of Darwin, evolutionary biologist Jerry Coyne has pointed out that Darwin himself predicted that traits not under natural selection might show greater degrees of variation and even decay, leading to his ideas about vestigial organs. When the inner ears of several three-toed sloths were examined, Guillaume Billet and his colleagues discovered that in every conceivable way of measuring the structure of the semicircular canals, these sloths varied more than other mammals. This higher degree of variation is indicative of very relaxed natural selection on the structures. Coyne has pointed out that the beauty of the study is not in showing that vestigial organs occur, because evolutionary biologists can cite many great examples of vestigial structures. What is significant about the work is actually catching the semicircular canals on their way to becoming vestigial.
The hagfish, as noted, does not have a jaw, so the next innovation involves organisms with jaws. The anatomical structure we call the jaw is an ancestral characteristic of bony and cartilaginous fish, reptiles, birds, and mammals (fig. 5.1). The five bones we are following are also sometimes hard to find in jawed fish. In primitive sturgeons, the bones of the head are fused into a single sheet, so discerning the individual bones is extremely difficult. But when advanced bony fish are examined, the individual bones of the jaw become recognizable. In bony fish, we easily recognize and localize three of these bones—the dentate, articular, and squamosal bones–but there is no trace of the quadrate and stapes bones. Fish do not have an outer ear, nor do they have a middle ear, so it is safe to say that the three bones that are recognizable in fish jaws reside well outside of the ear. These two bones appear in reptiles, though, where the quadrate is found outside of the inner ear, and the stapes is found inside of the inner ear connected to a membrane called the eardrum. Note that the articulation of the jaw in bony fish is produced by the contact of the quadrate and articular bones, and this is a universal characteristic of jaws in fish, reptiles, birds, turtles, and amphibians, but not in mammals. What has happened in this last group results in the incredibly complex inner ear that mammals have. The quadrate and articular bones of the jaw hinge in mammals are converted into inner ear bones called the malleus (the same as the articular) and the incus (the same as the quadrate). This new arrangement of the five bones we have been following, especially the last three (articular, quadrate, and stapes or malleus, incus, and stapes), makes up the three-boned inner ear in humans and other mammals. The mammalian jaw is made up of the articulation of the squamosal and dentary bones. This is quite a transition, but ultimately it is a frugal use of the jaw bones. There has been and continues to be strong selection on maintaining these bone structures and articulations in mammals, and hence there is little variation in mammals’ overall inner ear bone arrangement. But the range of sound wavelengths that can be detected by mammals can vary as a result of altering other structures of the inner ear, or even some of the structure of the middle and outer ear. One need only look at the size of the outer ear in mammals that require acute sound detection (larger ears are usually correlated with greater acuity of sound detection) to see how the outer ear can be altered as a response to selection for collecting and transmitting very specific sounds to the brain.
Figure 5.1. Comparison of bones of the jaw of a mammal and a reptile.
A modification of the inner ear of mammals that can produce variation in the range of hearing of mammals is the development of a coiled membranous extension at the bottom of the inner ear called the cochlea. Mammals are the only vertebrate with well-developed cochlear structures, and there is considerable variation in the makeup and length of this membranous structure. The cochlea varies in several aspects—total volume of the cochlea, number of spirals, total length of cochlea, diameter of the cochlear tube, tightness of the spiraling of the cochlear tube, and curviness of the cochlea—all of which have been correlated to the relative frequency of wavelengths that are detectable by the mammalian ear. For instance, mammals with large cochlear diameters are more sensitive to high-frequency sounds. So, one way mammals can adjust to the wavelengths that they can detect is through evolving different cochlear dimensions. The degree of variation of mammalian cochlear structures is impressive, reflecting the wide range of hearing capacity in mammals.