
Before starting the formal learning of handwriting, the child spends several years experimenting and training his or her graphomotor skills: doodling, drawing and pseudo-writing. These activities are fundamental because they help the child develop motor control of the hand. Those general manual skills can then serve as a support to learn how to write (Adi-Japha and Freeman, 2001; Lurcat, 1983). For young children, fine motor skills are relatively difficult to acquire (Ajuriaguerra, 1979; Lurcat, 1983) and depend on the maturation of the motor system. From a physiological point of view, this maturation, including proper control of distal muscles, occurs at age eight or nine. In fact, van Galen, Portier, Smits-Engelsman, and Schomaker (1993) showed that performance of children handwriting was more related to their motor development than to their chronological age. With motor maturation, precision can be gained, and writing size and pen pressure decrease.
In beginning writers, the graphic trace needs to be visually guided, adjusted and corrected on-line (Adi-Japha and Freeman, 2001; Tseng and Chow, 2000), but after a few years, visual and kinesthetic aspects of letters are fully memorised (Adi-Japha and Freeman, 2001). In adults, handwriting is relatively independent from the sensory feedback (try to write feedback with your eyes closed and you will see that your trace looks completely normal). This so-called proactive mode of motor control results in decrease of writing time and number of breaks in the movement: the gesture becomes more fluent.
Finally, before becoming an ‘expert’ writer, the child has to automatise the learned movements and store them in the form of ‘motor programs’. The concept of a motor program is central in the models dedicated to motor function (see Summers and Anson, 2009, for discussion of this notion). To determine the nature of motor programs in writing, researchers have looked for movement characteristics that stay invariant among different contexts (Teulings and Schomaker, 1993; Teulings, Thomassen and Van Galen, 1986). The rationale was that even though each writer has his or her own style of writing (which can sometimes cause difficulties for recognition or identification), some features of the movement vary with the context and external constraints (Orliaguet, Kandel and Boe, 1997; Thomassen and Schomaker, 1986; Wing, Nimmo-Smith, and Eldridge, 1983; van der Plaats, Rudolf, and van Galen, 1991), while other features stay invariant. Thus, features that stay stable in various conditions would be the ones stored at a high level of representation. These high-level representations correspond to the motor programs (Teulings and Schomaker, 1993). The motor program would inform on the sequence, direction and size of strokes (Ellis, 1982; Van Galen, 1991). This spatial coding would not be limited to independent strokes but include the spatial organisation of a succession of strokes, most likely at the level of a letter (Teulings, Thomassen, and Van Galen, 1983). Stelmach (1978), Van Galen (1991) and Teulings et al. (1983) proposed that only the first element of a sequence is programmed before initiating the movement to write a whole word, with the following units being programmed while the movement is executed. According to developmental studies, children stabilise and expand the size of the motor program slowly and reach a stable performance at approximately age 12 (Meulenbroek and Van Galen, 1988). Writing development is therefore a long and demanding process. Considering how early the learning starts, and how complex the simultaneous acquisition of the motor and linguistic codes is, it becomes necessary to hypothesise that learning how to write has consequences on the organisation of the brain.
Writing is not restricted to the hand. We are all able to write with other effectors such as the mouth, foot, elbow … while keeping individual spatial characteristics of our writing. This so-called motor equivalence was described by the Russian physiologist Bernstein (cited in Bongaardt, 2001). This shows that the intention of writing is coded at an abstract level, regardless of the muscle groups involved. The concept of previously defined motor program therefore corresponds to an abstract internal representation of the shape of the units we have to produce that is not tied to a particular muscle group but can be transferred to any effector.
The questions we address in the following sections is where and how this information is stored in our brain and how it can influence our perception of written shapes.
Following a brain injury, it can happen that previously skilled adult writers become unable to produce writing gestures in the absence of any other movement or language disorder. This condition is called “apraxic agraphia” (see Roeltgen, 2003), and results in writing that is slow and completely disorganised in space. Descriptions of agraphic patients and the localisation of their brain lesions were the first available data for understanding the organisation of the brain network sustaining handwriting.
The first scientist who assumed the existence a brain center for writing was the Austrian neurologist Sigmund Exner in 1881. He postulated that this center contained the ‘motor images’ needed to produce letters. After the autopsy of several agraphic patients, he noticed brain lesions commonly located near the “foot of the second frontal convolution of the left hemisphere” (Exner, 1881), in an area that functionally corresponds to the premotor cortex, above the Broca’s area (known to be involved in the oral production of language; see Dronkers, Plaisant, Iba-Zizen, and Cabanis, 2007 for details).
This center is traditionally referred to as “Exner’s area”, and was found to be damaged in other agraphic patients (with or without language deficits) (Anderson, Damasio and Damasio, 1990; Dubois, Hécaen, and Marcie, 1969). Most lesions leading to agraphia are otherwise located in the left posterior parietal lobe in right-handed individuals (see Planton, Jucla, Roux, and Démonet, 2013, for review). An interaction between those two regions therefore seems a prerequisite for the generation of handwriting movements. Their involvement has indeed been confirmed with brain imaging tools allowing the in vivo assessment of brain activity in the course of various tasks.
Functional brain-imaging results of the last two decades were recently summarised into two meta-analyses (Planton et al., 2013 and Purcell, Turkeltaub, Eden, and Rapp, 2011; see legend of Figure 1.1 for details on functional magnetic resonance imaging [fMRI]). Both confirm the implication of left-hemisphere frontal and parietal regions, previously identified in agraphic patients, in the production of handwriting gestures. Several studies support the idea that both the superior parietal lobule and the dorsal premotor cortex respond with a certain level of specificity to the production of handwriting movements; that is, their response is stronger when participants produce handwriting relative to any other motor or linguistic task. Note, however, that the exact position of Exner’s area along the precentral sulcus is still discussed, with observed activations often extending to, or being restricted to, more dorsal portions of the precentral sulcus, whereas Exner’s area defined from clinical cases is more ventral.
Other brain regions also appear implicated in the motor control of handwriting. The right cerebellum, known for its important role in motor coordination and motor learning, is the most consistently found (Planton et al., 2013). In addition, lexical processes such as word selection, orthographic and phonological processing are thought to depend on distinct parts of the brain, mostly in the left inferior frontal and temporal cortices. See Figure 1.1 for an overview of these regions (the cerebellum is not represented in three-dimensional views of the cortex).
The question of the functional specificity of those premotor and parietal regions, that is the possibility that neurons in those locations get tuned to writing movements relative to other movements of our repertoire, has been addressed in several studies. Brain mapping by direct cortical electrical stimulation can be realised on patients undergoing pre-surgical exploration before cyst or tumor excision. Patients are awake and can perform various tasks while their brain is stimulated locally, and the stimulation transiently disturbs the neural processing occurring in the targeted region. Roux et al. (2009) have explored the frontal cortex while patients were writing to dictation as well as reading sentences and naming objects presented visually. When the stimulation was applied specifically to the premotor cortex, in the left superior frontal sulcus, transient symptoms of motor agraphia such as ill-formed graphemes, slowing of the movement and even writing arrests were observed while neither the other two linguistic tasks, nor fine motor coordination of the hand, were affected. This study is important because it shows that performance of handwriting movements is causally related to the functioning of this left superior frontal region. As shown by functional imaging studies, the location of this region is more dorsal than the region defined by clinicians on the basis of agraphia. Roux et al. (2009) have proposed to term this region “graphemic motor frontal area” and have hypothesised that it acts as an interface between orthographic processing and motor control in writing. This region could therefore be crucial for the retrieval of motor programs.
Writing impairments following a brain lesion can, in rare cases, be expressed in the form of surprising dissociations. Anderson et al. (1990), Delazer, Lochy, Jenner, Domahs, and Benke (2002) and Starrfellt (2007) have reported patients for whom the writing of letters and words was severely disturbed, while the writing of digits and figures was performed in a normal effortless way. Delazer et al. (2002) have quantified the writing velocity of their patient and matched controls and have shown that writing velocity was strongly decreased for letters but was close to normal for digits. Interestingly, in the control group, the writing of digits was slightly slower than the writing of letters.
CT scans indicated that lesions leading to this highly specific deficit were located either in the left premotor cortex close to Exner’s area (Anderson et al.,1990), or in the left superior parietal and parieto-occipital regions (Delazer et al., 2002). In the latter case, lesions could not be precisely located (Starrfelt, 2007). This indicates that the retrieval of motor programs in memory could functionally depend on the nature of the symbols to be written. The motor system could code motor information as a function of the type of elements to produce, and the stability of motor programs corresponding to letters could be more strongly dependent on the integrity of the parieto-frontal regions described above. To address the question of neural specificity for letters versus digits writing in healthy participants, Longcamp et al. (2014) ran a brain-imaging study where the graphic trace produced on a digitising tablet and the variations in brain activity were recorded simultaneously. Participants had to write letters or digits to dictation, and the produced graphic traces were displayed visually in real time. The authors observed that a set of three brain regions, including the left premotor cortex at a location very similar to the previously identified graphemic motor frontal area, was more strongly activated when participants were writing letters than digits. The on-line recording of writing kinematics confirmed that differences in brain activity could not be explained by low-level differences in the execution of the movements but emerged from the mobilisation of more abstract representations. A possible explanation of this stronger functional specificity for linguistic symbols is that letters occur more frequently and in a more grouped way in written texts so that their representation in the brain regions controlling the hand would tend to cluster in a more restricted cortical space (see Polk and Farah, 1998, for a similar account of functional specificity in letters recognition). During development, kinematic features of the graphic movements get a dissociation between writing letters and drawing geometrical shapes at around age six or seven. This could be the origin of the preferential response of this premotor region to letters.
In addition, most of the time, the handwriting movement co-occurs with visual feedback: the graphic trace is perceived on-line. Coincident visual and motor experience with letters is also thought to lead to strong links in the organisation of visual and motor networks allowing the processing of written language.
Interestingly, one of the agraphic patients described earlier, who produced severely distorted letters following a lesion of the left premotor cortex, also displayed important difficulties in reading and visual recognition of single letters (Anderson et al., 1990). Reading of digits was preserved. Based on this observation, Anderson and his collaborators hypothesised a “possible role of premotor cortices in the coactivation of precise sequences of motor and sensory activity patterns involved in reading and writing” (Anderson et al., 1990: 80). This suggests that writing knowledge, built through repeated production of letters and letter strings, has a role in visual recognition of written language.
One argument supporting the idea of a participation of writing knowledge to reading can be found in the description of pure alexic patients. Pure alexia is a condition occurring most often after a left occipito-temporal lesion involving associative visual cortex, where patients suddenly display important difficulties in reading. The recognition of single letters is severely impaired. In some cases, patients spontaneously adopt strategies where they form the letters with their fingers in the air. This facilitates the retrieval of letter names and reading (Bartolomeo, Bachoud-Le´vi, Chokron, and Degos, 2002a, 2002b; Seki, Yajima, and Sugishita, 1995). Activation of the motor representation of letters may therefore help restoring access to the identity of letters when their visual representation is disturbed in pathological conditions. But it is also used in non-pathological conditions when for instance the shape of the characters is highly complex, as in Japanese. It is well known that Japanese readers spontaneously use in-air writing to retrieve the identity and meaning of infrequent complex characters when they read (Sasaki, 1987). In ideographic writing systems like Japanese or Chinese, learning of characters relies on repeated copying of each character. This has been shown to facilitate memorisation (Naka and Naoi, 1995).
The representation of the movements learned through tracing can bind to the visual information in memory to access the identity of a letter. This binding most likely depends on the training conditions. Our group showed for instance that when 5-year-old children learn letters through handwriting, their subsequent recognition performance is better than when they learn letters through a keyboard (Longcamp, Zerbato-Poudou, and Velay, 2005). The difference between the training modalities is that with a keyboard, letters are not traced anymore but only pointed to in space. Motor representation becomes less differentiated and less complex, providing little information on the shape of the letter. Interestingly, recent data show that in China, increased use of computers and smartphones to write in children has adverse effects on reading scores (Tan, Xu, Chang, and Siok, 2012). In a series of studies, Karin James and her collaborators have shown that at the brain level, the posterior visual brain regions of children get more strongly activated when children reading letters who have been trained through handwriting (James and Engelhardt, 2012). The important factor for this effect to arise is the actual motor execution of writing because when children are trained only by viewing the experimenter trace the letters, the subsequent enhancement of visual activations is smaller. Finally, even more recently, the same group has also shown that the visual feedback produced in the case of active self-generated writing movements has a certain importance in the building of visual representations: writing by hand, indeed, generates a variable visual trace on the paper, and the repeated viewing of this variable trace, combined with the motor commands used to generate it, could help the visual system getting more flexible and more efficient when facing different instances of a given letter (Li and James, 2016).
These data support the idea that motor representations built through repeated performance of handwriting movements have a functional impact on reading, at the level of single letters. Extra evidence comes from neuro-imaging studies.
Longcamp, Anton, Roth and Velay (2003) have addressed this issue by having volunteers presented with either single letters or with unknown shapes resembling characters. They observed that the mere visual observation of letters, but not of unknown characters, gave rise to activations of a premotor region of the left hemisphere in right-handers, in addition to the typically expected visual cortical activations. This premotor region, as described earlier, is considered crucial for the maintenance and retrieval of writing movements in memory. Longcamp et al. (2003) actually observed that this same premotor region was indeed strongly activated when participants were in the course of producing writing. It did not respond when participants observed the unknown characters, because no motor representation could be evoked. Comparable results have been found by James and Gauthier (2006). Interestingly, a symmetrical region, in the right premotor cortex, is preferentially activated during letter recognition in left-handed participants, because in left-handers, the left writing hand is controlled by the right hemisphere (Longcamp et al., 2005). The activation of this premotor region in reading was shown to be driven by the level of literacy: its function is therefore modified by the cultural acquisition of written language (Dehaene et al., 2010).
This automatic motor activation, which has been termed ‘motor resonance’, is present whatever the font observed. For instance in the Longcamp et al. (2003, 2005) studies, stimuli were standard upper-case printed letters that had little correspondence with the shapes participants would produce spontaneously. However, although it seems that resonance is triggered by the letter identity, experimental evidence indicates that it gets stronger when the font itself conveys dynamical information, as it is the case for handwritten letters. Longcamp et al. (2011) found for instance that the dorsal premotor cortex, together with the primary motor cortex and other cortical regions involved in motor control, were more strongly activated when participants observed handwritten rather than printed letters.
In addition, visual presentation of handwritten letters triggered responses in visual regions usually involved in visual motion processing, such as area V5/MT whose neurons are sensitive to stimuli moving in the visual field. The dynamical information present static handwritten traces seems to be re-enacted in the brain, provided that the perceived shapes belong to the repertoire of the individual (Nakatsuka, Thabit, Koganemaru, Nojima, Fukuyama, and Mima, 2012). Nakamura, Kuo, Pegado, Cohen, Tzeng and Dehaene (2012) have shown that motor cortical activations are maximal if cursive words are presented dynamically, with strokes unfolding in the direction that would be naturally produced by the writer. When the dynamical information does not comply with the rules of production, motor resonance drops. Up to now, no study has tested which of the dynamical visual features present in the graphic trace is actually critical to obtain those neural effects. It seems, however, that the eccentricity of the curves present in the handwriting, and their slant, are important elements. When the eccentricity of elliptic shapes is close to the eccentricity that the observer would spontaneously produce (as biomechanically, the motor system produces some eccentricities more easily than other), the perception of the ellipses is facilitated (Wamain, Tallet, Zanone and Longcamp, 2011). One possible function of the motor reactivation in the course of perceiving handwritten traces could be the recognition of the writer (Barton et al., 2010; Wamain Tallet, Zanone, and Longcamp, 2012). Interestingly, it has been noticed that patients who become unable to recognise faces (a condition termed prosopagnosia), following a brain lesion in the right visual system, also lose this ability to recognise the writer of a handwritten text (Barton et al., 2010). Embodiment would therefore mediate social recognition of “who” wrote. Going one step further, some authors have also proposed that aesthetic experience during the observation of artwork is affected by embodied simulation of the actions of the painter, that gave rise to the visible trace, and that these mechanisms are universal (Freedberg and Gallese, 2007).
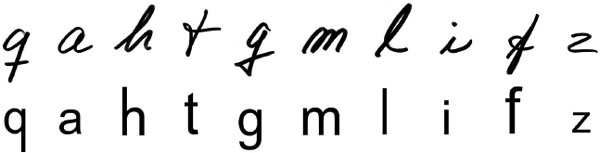
Figure 1.2 Examples of the handwritten and typed stimuli used in Longcamp et al.’s (2011) study.
A follow-up issue was to understand if motor resonance could be manipulated, as a function of the experience with the characters. To this end, Longcamp et al. (2008) designed a training protocol very similar to the one carried out in preschool children (Longcamp et al., 2005), where adult participants learned unknown Tamil characters, either by hand, or through an adapted computer keyboard. In this adult sample, behavioral results were very similar to results of children: characters trained by hand were better recognised a few days after the end of training sessions. Participants displayed fewer confusions between the characters and their mirror images, when they were trained through handwriting. Handwriting therefore seems to provide the visual system with crucial information regarding the spatial configuration of characters. In addition, brain imaging recordings indicated stronger activations in a network of brain regions involved in programming and execution of manual actions, including the left premotor cortex, for characters trained through handwriting. Typewriting training, perhaps because movements remain nonspecific, did not elicit preferential brain activations relative to handwriting training.
The way we learn how to write may therefore shape our cortical representations of written language (Velay and Longcamp, 2012; James and Engelhardt, 2012; James and Atwood, 2009). Writing habits are undergoing massive changes with the now massive use of digitising tools. It is therefore possible that this strong motor perceptual coupling that normally develops in the course of literacy acquisition will no longer exist in the near future. Consequences of those changes are currently investigated from various perspectives (Kiefer and Velay, 2016). Beyond single characters and letters, reading whole words involves linguistic (semantic, phonological, orthographic) processes, and one can wonder whether motor resonance also plays a role not only in visuospatial analysis, but also in linguistic processing of written traces, in the context of actual reading. In the last study described in this chapter, we investigated whether learning a second language through writing would facilitate memorisation and subsequent reading of this language. We chose Chinese as a test language, given the specificity of the ideographic writing system, where each character has strong linguistic features.
The aim of this study was to understand how the neural activity organises when French students learn to read Chinese. For readers of alphabetic language, learning to read Chinese is very challenging. Each Chinese character has a very complex layout, associated with a specific meaning and their pronunciation cannot be derived directly from the visual form. In this context, the stability of the orthographic representation of Chinese characters is essential. Our hypothesis was that learning Chinese characters by writing should stabilise the orthographic representation and strengthen the integration of semantic information.
We taught French students who were undertaking Chinese classes at the university 48 new Chinese characters. The students followed five consecutive days of training during which they had to learn half of these characters by writing and the other half by pronunciation. Two days after the end of their training, the subjects were scanned in fMRI. We measured their brain activity when they read newly learned characters. We tested to which extent the neural substrates of reading and writing Chinese words overlapped and whether the type of training (handwriting or pronunciation) had a direct impact this neural organisation. We observed that when subjects read Chinese characters, an extended brain network was activated. Included in this network were six regions that matched those found when the participants were actually writing. Among those regions, we found the left premotor and superior parietal regions described as writing specific in the previous sections and the right cerebellum, whose involvement in writing is also consistently reported. This constitutes a clear argument supporting the idea that the neural substrates of writing are mobilised during linguistic processing of written characters when meaning has to be assessed, in the course of reading. In addition, the level of activation of those regions, that is the level of motor resonance during actual reading, was stronger when the characters had been trained by hand than by pronunciation. Finally, we noted that the response of the left fusiform gyrus, a region typically found in the context of orthographic processing (Cohen et al., 2000; Cohen and Dehaene, 2004, region 4 in Figure 1.1), was also modulated by the type of training and by the linguistic properties of characters.
The constant evolution of brain imaging techniques allows a better characterisation of the neural underpinnings of writing, and of motor resonance during reading. Because of strong coupling between dynamical visual information and gesture production in the course of learning how to read and write, the visual and motor networks are in constant interplay when we process written language. With the massive changes occurring in the current society of computers and mobile devices, handwriting is getting replaced by other modes of written language production, and this visual–motor coupling is already changing. The challenge in the next few years will be to understand how the organisation of writing in the brain, and its relationship with other perceptual and linguistic skills, adapts to material constraints of the digital revolution
Adi-Japha, E., and Freeman, N. H. (2001). Development of differentiation between writing and drawing systems. Developmental Psychology, 37(1), 101114.
Ajuriaguerra, J. de. (1979). L’écriture de l’enfant: L’ évolution de l’écriture et ses difficultés. Neuchatel: Delachaux et Niestlé.
Anderson, S. W., Damasio, A. R., and Damasio, H. (1990). Troubled letters but not numbers. Domain specific cognitive impairments following focal damage in frontal cortex. Brain: A Journal of Neurology, 113, 749766.
Bartolomeo, P. (2008). The neural correlates of visual mental imagery: An ongoing debate. Cortex, 44(2), 107108.
Bartolomeo, P., Bachoud-Le´vi, A.-C., Chokron, S., and Degos, J.-D. (2002a). The relationship between visual perception and visual mental imagery: A reappraisal of the neuropsychological evidence. Cortex, 38(3), 357378.
Bartolomeo, P., Bachoud-Le´vi, A.-C., Chokron, S., and Degos, J.-D. (2002b). Visually- and motor-based knowledge of letters: Evidence from a pure alexic patient. Neuropsychologia, 40, 1363–1371.
Barton, J. J., Sekunova, A., Sheldon, C., Johnston, S., Iaria, G., and Scheel, M. (2010). Reading words, seeing style: The neuropsychology of word, font and handwriting perception. Neuropsychologia, 48(13), 3868–3877.
Bongaardt, R. (2001). How Bernstein conquered movement: Revisiting the work of Nikolai Aleksandovitsch Bernstein, In M. Latash and V. M. Zatsiorsky (Eds.) Classics in Movement Science. Champaign, Ill: Human Kinetics.
Cohen, L., and Dehaene, S. (2004). Specialization within the ventral stream: The case for the visual word form area. NeuroImage, 22(1), 466476.
Cohen, L., Dehaene, S., Naccache, L., Lehéricy, S., Dehaene-Lambertz, G., Hénaff, M. A., and Michel, F. (2000). The visual word form area: Spatial and temporal characterization of an initial stage of reading in normal subjects and posterior split-brain patients. Brain: A Journal of Neurology, 123(Pt 2), 291307.
Dehaene, S., Pegado, F., Braga, L. W., Ventura, P., Filho, G. N., Jobert, A., … Cohen, L. (2010). How learning to read changes the cortical networks for vision and language. Science, 330(6009), 13591364.
Delazer, M., Lochy, A., Jenner, C., Domahs, F., and Benke, T. (2002). When writing 0 (zero) is easier than writing O (o): A neuropsychological case study of agraphia. Neuropsychologia, 40(12), 21672177.
Dronkers, N. F., Plaisant, O., Iba-Zizen, M. T., and Cabanis, E. A. (2007). Paul Broca’s historic cases: High resolution MR imaging of the brains of Leborgne and Lelong. Brain, 130(5), 14321441.
Dubois, J., Hecaen, P., and Marcie, P. (1969). L’agraphie “pure”. Neuropsychologia, 3, 271286.
Ellis, A. W. (1982). Normality and Pathology in Cognitive Functions. London: Academic Press.
Exner, S. (1881). Untersuchungen Über die Localisation der Functionen in der Grosshirnrinde des Menschen. Vienna: Wilhelm Braumüller.
Freedberg, D., and Gallese, V. (2007). Motion, emotion and empathy in esthetic experience. Trends in Cognitive Science, 11(5), 197–203.
James, K. H., and Atwood, T. P. (2009). The role of sensorimotor learning in the perception of letter-like forms: Tracking the causes of neural specialization for letters. Cognitive Neuropsychology, 26(1), 91–110.
James, K. H., and Engelhardt, L. (2012). The effects of handwriting experience on functional brain development in pre-literate children. Trends in Neuroscience and Education, 1(1), 3242.
James, K. H., and Gauthier, I. (2006). Letter processing automatically recruits a sensory-motor brain network. Neuropsychologica 44: 2937–2949.
Kiefer, M., and Velay, J. L. (2016). Writing in the digital age. Trends in Neuroscience and Education, 15(3): 77–156.
Li, J. X., and James, K. H. (2016). Handwriting generates variable visual output to facilitate symbol learning. Journal of Experimental Psychology: General, 145(3), 298313.
Longcamp, M., Anton, J.-L., Roth, M., and Velay, J.-L. (2003). Visual presentation of single letters activates a premotor area involved in writing. NeuroImage, 19(4), 14921500.
Longcamp, M., Anton, J.-L., Roth, M., and Velay, J.-L. (2005). Premotor activations in response to visually presented single letters depend on the hand used to write: A study on left-handers. Neuropsychologia, 43(12), 18011809.
Longcamp, M., Boucard, C., Gilhodes, J.-C., Anton, J.-L., Roth, M., Nazarian, B., and Velay, J.-L. (2008). Learning through hand- or typewriting influences visual recognition of new graphic shapes: Behavioral and functional imaging evidence. Journal of Cognitive Neuroscience, 20(5), 802815.
Longcamp, M., Hlushchuk, Y., and Hari, T. (2011). What differs in the visual recognition of handwritten vs printed letters? An fMRI study. Human Brain Mapping 32: 1250–1259.
Longcamp, M., Lagarrigue, A., Nazarian, B., Roth, M., Anton, J.-L., Alario, F.-X., and Velay, J.-L. (2014). Functional specificity in the motor system: Evidence from coupled fMRI and kinematic recordings during letter and digit writing. Human Brain Mapping, 35(12), 60776087.
Longcamp, M., Zerbato-Poudou, M.-T., and Velay, J.-L. (2005). The influence of writing practice on letter recognition in preschool children: A comparison between handwriting and typing. Acta Psychologica, 119(1), 6779.
Lurcat, L. (1983). Le graphisme et l’écriture chez l’enfant. Revue française de pédagogie, 65, 718.
Meulenbroek, R. G. J., and Van Galen, G. P. (1988). The acquisition of skilled handwriting: Discontinuous trends in kinematic variables. In A. M. Colley and I. R. Beech (Eds.), Cognition and Action in Skilled Behaviour (pp. 273–281). Amsterdam: North Holland.
Naka, M., and Naoi, H. (1995). The effect of repeated writing on memory. Memory & Cognition, 23(2), 201212.
Nakamura, K., Kuo, W.-J., Pegado, F., Cohen, L., Tzeng, O. J. L., and Dehaene, S. (2012). Universal brain systems for recognizing word shapes and handwriting gestures during reading. Proceedings of the National Academy of Sciences, 109(50), 2076220767.
Nakatsuka, M., Thabit, M. N., Koganemaru, S., Nojima, I., Fukuyama, H., and Mima, T. (2012). Writing’s shadow: Corticospinal activation during letter observation. Journal of Cognitive Neuroscience, 24(5), 11381148.
Orliaguet, J.-P., Kandel, S., and Boë, J.-L. (1997). Visual perception of motor anticipation in cursive handwriting: Influence of spatial and movement information on the prediction of forthcoming letters. Perception, 26, 905912.
Planton, S., Jucla, M., Roux, F.-E., and Démonet, J.-F. (2013). The “handwriting brain”: A meta-analysis of neuroimaging studies of motor versus orthographic processes. Cortex: A Journal Devoted to the Study of the Nervous System and Behavior, 49(10), 27722787.
Polk, T. A., and Farah, M. J. (1998). The neural development and organization of letter recognition: Evidence from functional neuroimaging, computational modeling, and behavioral studies. Proceedings of the National Academy of Sciences, 95, 847–852.
Purcell, J. J., Turkeltaub, P. E., Eden, G. F., and Rapp, B. (2011). Examining the central and peripheral processes of written word production through meta-analysis. Frontiers in Psychology, 2, 239.
Roeltgen, D. P. (2003). Agraphia. In M. Kenneth, M. D. Heilman, and M. D. Edward Valenstein (Eds.) (4th ed.). Oxford: Oxford University Press, pp. 126–146.
Roux, F-E., Dufor, O., Giussani, C., Wamain, Y., Draper, L., Longcamp, M., and Démonet, J.-F. (2009). The graphemic/motor frontal area Exner’s area revisited. Annals of Neurology, 66(4), 537545.
Sasaki, M. (1987). Why do Japanese write characters in space? International Journal of Behavioral Development, 10(2), 135149.
Seki, K., Yajima, M., and Sugishita, M. (1995). The efficacy of kinesthetic reading treatment for pure alexia. Neuropsychologia, 33(5), 595597.
Starrfelt, R. (2007). Selective alexia and agraphia sparing numbers—a case study. Brain and Language, 102(1), 5263.
Stelmach, G. (1978). Information Processing in Motor Control and Learning. New York: Academic Press.
Summers, J. J., and Anson, J. G. (2009). Current status of the motor program: Revisited. Human Movement Science, 28(5), 566577.
Tan, L. H., Xu, M., Chang, C. Q., and Siok, W. T. (2012). China’s language input system in the digital age affects children’s readings development. Proceedings of the National Academy of Sciences of the United States of America 110(3): 1119–1123.
Teulings, H-L., and Schomaker, L. R. (1993). Invariant properties between stroke features in handwriting. Acta Psychologica, 82(1–3), 69–88.
Teulings, H-L., Thomassen, A. J. W. M., and van Galen, G. P. (1983). Preparation of partly precued handwriting movements: The size of movement units in handwriting. Acta Psychologica, 54(1–3), 165–177.
Teulings, H-L., Thomassen, A. J. W. M., and Van Galen, G. P. van. (1986). Invariants in handwriting: The information contained in a motor program. In Dans H. S. R. Kao, G. P. van Galen, and R. Hoosain (Éds.), Graphonomics: Contemporary Research in Handwriting (pp. 305–315). Amsterdam: Elsevier.
Thomassen, A. J. W. M., and Schomaker, L. R. B. (1986). Between-letter context effects in handwriting trajectories. In Graphonomics: Contemporary Research in Handwriting (Vol. 37, pp. 253–272). Amsterdam: North-Holland.
Tseng, M. H., and Chow, S. M. K. (2000). Perceptual-motor function of school-age children with slow handwriting speed. American Journal of Occupational Therapy, 54, 8388.
van der Plaats, R. E., and van Galen, G. P. (1991). Allographic variability in adult handwriting. Human Movement Science, 10(2–3), 291–300.
van Galen, G. P. (1991). Handwriting: Issues for a psychomotor theory. Human Movement Science, 10(2–3), 165–191.
van Galen, G. P., Portier, S. J., Smits-Engelsman, B. C. M., and Schomaker, L. R. B. (1993). Neuromotor noise and poor handwriting in children. Acta Psychologica, 82(1–3), 161178.
Velay, J.-L., and Longcamp, M. (2012). Handwriting versus typewriting: Behavioural and cerebral consequences in letter recognition. In M. Torrance, D. Alamargot, M. Castelló, F. Ganier, O. Kruse, and A. Mangen (Eds.), Learning to Write Effectively: Current Trends in European Research (pp. 371–373). Leiden: Brill.
Wamain, Y., Tallet, J., Zanone, P. G., and Longcamp, M. (2011). “Biological geometry perception”: Visual discrimination of eccentricity is related to individual motor preferences. PLOS One, 6(1), e15995.
Wamain, Y., Tallet, J., Zanone, P. G., and Longcamp, M. (2012). Brain responses to handwritten and printed letters differentially depend on the activation state of the primary motor cortex. Neuroimage, 63(3), 1766–1773.
Wing, A. M., Nimmo-Smith, M. I., and Eldridge, M. A. (1983). The consistency of cursive letter formation as a function of position in the word. Acta Psychologica, 54(1–3), 197–204.