7
DESDE MEU PRIMEIRO DIA no laboratório, eu sabia que não deixaria Cambridge por um longo período. Partir seria estupidez, porque eu descobrira de imediato como era divertido conversar com Francis Crick. Encontrar alguém no laboratório de Max que soubesse que o DNA era mais importante do que as proteínas era uma verdadeira sorte. Além do mais, era um grande alívio não ter de passar todo o tempo aprendendo análise radiográfica de proteínas. Nossas conversas na hora do almoço logo se concentraram em como os genes se aglomeravam. Poucos dias após minha chegada, sabíamos o que fazer: imitar Linus Pauling e superá-lo em seu próprio jogo.
O sucesso de Pauling com a cadeia polipeptídica sugeriu naturalmente a Francis que o mesmo artifício poderia funcionar com o DNA. Mas, ainda que ninguém por perto pensasse que o DNA estava no coração de tudo, as potenciais dificuldades pessoais com o laboratório do King’s o refrearam de se envolver com o tema. Além disso, apesar de a hemoglobina não ser o centro do universo, os dois anos anteriores de Francis no Cavendish certamente não foram monótonos. Problemas com proteínas continuaram a aparecer em quantidade mais do que suficiente e exigiam alguém com uma inclinação para a teoria. Mas agora, comigo no laboratório querendo sempre falar sobre genes, Francis deixou de manter seus pensamentos sobre o DNA num recôndito escondido do cérebro. Ainda assim, ele não tinha intenção de abandonar o interesse por outros problemas do laboratório. Ninguém se importaria se, ao passar apenas algumas horas por semana pensando no DNA, ele me ajudasse a solucionar um problema de enorme importância.
Como consequência, John Kendrew logo percebeu que era improvável que eu o ajudasse a determinar a estrutura da mioglobina. Como era incapaz de cultivar grandes cristais de mioglobina equina, ele esperava que eu tivesse o dedo mais verde que o seu. Entretanto, nenhum esforço era necessário para ver que minhas experiências de laboratório eram menos habilidosas do que as de um químico suíço. Cerca de quinze dias após a minha chegada a Cambridge, fomos até o matadouro local para pegar um coração de cavalo para uma nova preparação de mioglobina. Se tivéssemos sorte, os danos às moléculas de mioglobina que impedem a cristalização seriam evitados com o congelamento imediato do coração do velho cavalo de corrida. Mas minhas tentativas subsequentes com a cristalização não foram mais bem-sucedidas do que as de John. De certa forma, eu estava quase aliviado. Se elas tivessem funcionado, John poderia ter me colocado para tirar fotografias de raios X.
Assim, nenhum obstáculo me impedia de passar muitas horas por dia conversando com Francis. Pensar o tempo todo era demais até mesmo para ele e, muitas vezes, quando se via frustrado com suas equações, costumava recorrer aos meus conhecimentos sobre fagos. Em outros momentos, Francis procurava preencher meu cérebro com fatos cristalográficos, disponíveis normalmente apenas por meio da leitura dolorosa de periódicos profissionais. Os raciocínios precisos necessários para entender como Linus Pauling havia descoberto a α-hélice foram particularmente importantes.
Logo me mostraram que a conquista de Pauling era um produto do senso comum, não o resultado de um raciocínio matemático complicado. Equações se misturavam ocasionalmente ao seu argumento, mas, na maioria dos casos, palavras teriam sido suficientes. A chave para o sucesso de Linus era sua confiança nas leis simples da química estrutural. A α-hélice não havia sido descoberta apenas pela contemplação de imagens de raios X; o pulo do gato, em vez disso, fora perguntar que átomos gostavam de ficar próximos uns dos outros. No lugar de papel e lápis, as principais ferramentas de trabalho eram um conjunto de modelos moleculares que se assemelhavam superficialmente a brinquedos de crianças em idade pré-escolar.
Desse modo, não víamos nenhuma razão para que não pudéssemos elucidar o DNA da mesma forma. Tudo o que tínhamos de fazer era construir um conjunto de modelos moleculares e começar a brincar – com sorte, a estrutura seria uma hélice. Qualquer outro tipo de configuração seria muito mais complexa; e preocupar-se com complicações antes de excluir a possibilidade de que a resposta fosse simples seria tolice. Pauling nunca chegou a lugar algum perseguindo confusões.
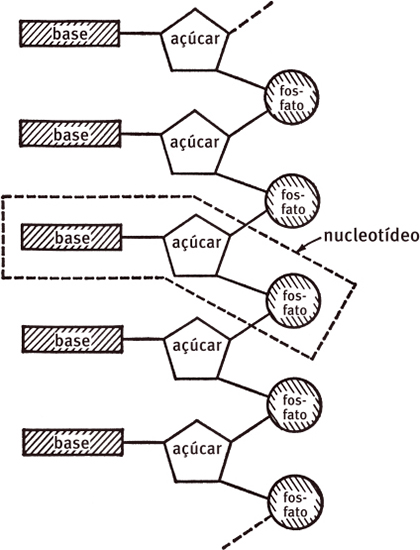
Desde nossas primeiras conversas, presumimos que a molécula de DNA continha um número muito grande de nucleotídeos ligados linearmente de modo regular. Mais uma vez, nosso raciocínio se baseava, em parte, na simplicidade. Apesar de os químicos orgânicos do laboratório vizinho, de Alexander Todd, acreditarem que era essa a organização básica, eles ainda estavam longe de estabelecer quimicamente que todas as ligações entre os nucleotídeos eram idênticas. Se não fosse o caso, no entanto, não víamos como as moléculas de DNA se compactavam para formar os agregados cristalinos estudados por Maurice Wilkins e Rosalind Franklin. Dessa forma, a não ser que encontrássemos um bloqueio completo ao nosso progresso futuro, o melhor caminho era considerar o esqueleto açúcar-fosfato como algo extremamente regular e procurar uma configuração helicoidal tridimensional na qual todos os grupos do esqueleto tivessem ambientes químicos idênticos.
Vimos de imediato que a solução para o DNA poderia ser mais complicada do que a da α-hélice. Na α-hélice, uma única cadeia de polipeptídeo (um grupo de aminoácidos) se torce em um arranjo helicoidal unido por ligações de hidrogênio entre grupos da mesma cadeia. Maurice havia dito a Francis, entretanto, que o diâmetro da molécula de DNA seria mais grosso caso existisse apenas uma cadeia de polinucleotídeo (um grupo de nucleotídeos). Isso o levou a pensar que a molécula de DNA era uma hélice composta, formada por diversas cadeias de polinucleotídeos torcidas umas sobre as outras. Se isso fosse verdade, antes de começar a criar um modelo sério, seria necessário decidir se as cadeias seriam mantidas unidas por ligações de hidrogênio ou por conexões de sal envolvendo os grupos fosfato de carga negativa.
Uma complicação adicional surgiu do fato de quatro tipos de nucleotídeos serem encontrados no DNA. Nesse sentido, o DNA não era uma molécula regular, mas altamente irregular. Os quatro nucleotídeos não eram, entretanto, completamente diferentes, já que cada um deles continha os mesmos açúcares e fosfatos. Sua singularidade estava nas bases nitrogenadas, que eram ou uma purina (adenina e guanina) ou uma pirimidina (citosina e timina). Mas, como as ligações entre os nucleotídeos envolviam apenas os grupos açúcar e fosfato, nossa suposição de que o mesmo tipo de elo químico ligava todos os nucleotídeos não foi afetada. Ao construir um modelo, nós presumiríamos que o esqueleto açúcar-fosfato era muito regular e a ordem das bases necessárias, muito irregular. Se as sequências de base fossem sempre as mesmas, todas as moléculas de DNA seriam idênticas e não existiria a variação que distingue um gene de outro.
Uma pequena parte do DNA, como vislumbrada pelo grupo de pesquisa de Alexander Todd em 1951. Eles pensavam que todos os elos entre nucleotídeos fossem ligações fosfodiéster, que juntavam o átomo de carbono nº 5 do açúcar ao átomo de carbono nº 3 do açúcar do nucleotídeo adjacente. Como químicos orgânicos, eles estavam preocupados em como os átomos se ligavam uns aos outros, deixando para os cristalógrafos o problema do arranjo tridimensional dos átomos.
Apesar de Pauling ter chegado à α-hélice praticamente sem as provas de raios X, ele sabia de sua existência e, em certo grau, levou isso em consideração. Devido aos dados radiográficos, uma grande variedade de configurações tridimensionais possíveis para a cadeia de polipeptídeo foi rapidamente descartada. Os dados exatos de raios X nos ajudariam a avançar bem mais rápido com a molécula mais sutilmente construída do DNA. Uma simples observação da imagem radiográfica do DNA preveniria um bom número de inícios equivocados. Felizmente, já existia uma fotografia razoável na literatura publicada. Ela tinha sido feita cinco anos antes pelo cristalógrafo inglês W.T. Astbury e poderia ser usada para dar início à nossa jornada. Possuir fotografias cristalinas muito superiores às de Maurice poderia economizar de seis meses a um ano de trabalho. O doloroso fato de que elas pertenciam a Maurice não poderia ser evitado.
Não havia nada a fazer além de conversar com ele. Para nossa surpresa, Francis não teve dificuldade em persuadir Maurice a vir a Cambridge para um fim de semana. E não foi preciso forçá-lo a concluir que a estrutura era uma hélice. Não só era a sugestão óbvia, como Maurice já falara em termos de hélices em um encontro de verão em Cambridge. Cerca de seis semanas antes de eu chegar lá pela primeira vez, ele havia mostrado fotografias de difração de raios X do DNA, que revelavam uma ausência marcante de reflexos no meridiano. Essa era uma característica que seu colega, o teórico Alex Stokes, lhe dissera ser compatível com uma hélice. Posto isso, Maurice suspeitou que três cadeias de polinucleotídeos fossem usadas para construir a hélice.
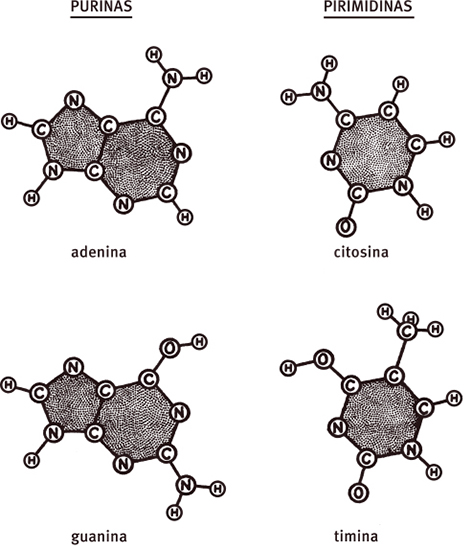
As estruturas químicas das quatro bases do DNA, como costumavam ser representadas por volta de 1951. Pelo fato de os elétrons nos anéis de cinco e seis membros não serem localizados, cada base tem um formato plano com uma espessura de 3,4 Å.
Ele, entretanto, não compartilhava da nossa crença de que o jogo de criação de modelos de Pauling determinaria rapidamente a estrutura, pelo menos não até que fossem obtidos mais resultados de raios X. A maioria de nossas conversas, em vez disso, estava focada em Rosy Franklin. Ela estava gerando mais problema do que nunca. Agora insistia que nem mesmo Maurice deveria tirar mais fotos de raios X do DNA. Ao tentar chegar a um acordo com Rosy, Maurice fez um péssimo negócio. Ele havia cedido a ela todos os bons DNAs cristalinos usados em seu trabalho original e concordado em restringir seus estudos a outro DNA, que ele descobriu depois que não cristalizava.
Eles chegaram ao ponto em que Rosy nem ao menos contava a Maurice seus últimos resultados. Ele soube como estavam as coisas somente três semanas depois, no meio de novembro. Rosy faria, então, uma apresentação sobre o trabalho realizado nos seis meses anteriores. Naturalmente, fiquei encantado quando Maurice disse que eu seria bem-vindo à apresentação dela. Pela primeira vez tinha um incentivo real para aprender algo de cristalografia: não queria que Rosy me desse lições.