26
MEU ESQUEMA SE PARTIU em pedaços no meio do dia seguinte. Havia contra mim o fato químico desagradável de que eu escolhera as formas tautoméricas erradas da guanina e da timina. Antes que essa verdade perturbadora fosse revelada, eu havia tomado um café da manhã rápido no Whim e passado no Clare para responder a uma carta de Max Delbrück, na qual ele contava que meu manuscrito sobre genética bacteriana parecera frágil aos geneticistas da Caltech. No entanto, ele aceitaria meu pedido para que o enviasse ao Proceedings of the National Academy. Dessa forma, ainda seria jovem ao cometer a tolice de publicar uma ideia estúpida. E poderia me regenerar antes que minha carreira tomasse para sempre um rumo temerário.
No início, essa mensagem teve o efeito perturbador desejado. Mas, àquela hora, com meu humor animado pela possibilidade de ter a estrutura autoduplicadora, reiterei a fé de que eu sabia o que se passava quando as bactérias se reproduziam sexualmente. Além disso, não poderia me abster de acrescentar uma frase, dizendo que havia acabado de idealizar uma linda estrutura do DNA que era completamente diferente da estrutura de Pauling. Por alguns segundos, considerei dar detalhes do que me ocupava, mas, como estava na correria, desisti, joguei a carta rapidamente na caixa de correio e voei para o laboratório.
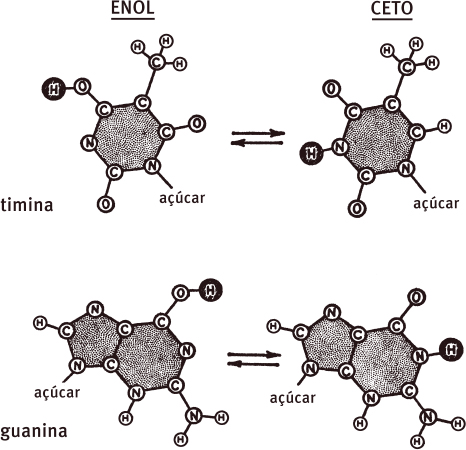
Não fazia nem uma hora que eu tinha postado a carta quando percebi que minha ideia não fazia sentido. Logo depois de chegar ao escritório e começar a explicar meu esquema, o cristalógrafo americano Jerry Donohue declarou que ela não funcionaria. As formas tautoméricas que eu copiara do livro de Davidson estavam, na opinião dele, designadas incorretamente. Minha réplica imediata de que muitos outros textos também mostravam guanina e timina na forma enólica não convenceu Jerry. Ele revelou alegremente que, durante anos, químicos orgânicos vinham favorecendo arbitrariamente formas tautoméricas específicas em detrimento de alternativas baseadas em parâmetros muito frágeis. De fato, os livros de química orgânica estavam cheios de imagens de formas tautoméricas altamente improváveis. A imagem da guanina que apresentara estava quase certamente distorcida. Toda a sua intuição química lhe dizia que ela ocorria na forma cetônica. Ele estava igualmente certo de que a timina também fora designada equivocadamente na forma enólica. Mais uma vez, ele favorecia bastante a alternativa cetônica.
Jerry, entretanto, não deu uma razão infalível para preferir as formas cetônicas. Ele reconheceu que apenas uma estrutura cristalina estava por trás do problema. Era a dicetopiperazina, cuja configuração tridimensional havia sido solucionada com cuidado no laboratório de Pauling anos antes. Nesse caso, não havia dúvida de que a forma cetônica, e não a enólica, estava presente. Além do mais, ele tinha certeza de que os argumentos da mecânica quântica que mostraram o porquê de a dicetopiperazina ter forma cetônica também poderiam valer para a guanina e a timina. Dessa forma, eu estava fortemente impelido a não perder mais tempo com meu esquema tolo.
Apesar de minha reação imediata ser esperar que Jerry estivesse blefando, não rejeitei suas críticas. Depois do próprio Linus, Jerry sabia mais sobre ligações de hidrogênio do que qualquer outra pessoa no mundo. Ele trabalhara na Caltech com estruturas cristalinas de moléculas orgânicas pequenas durante muitos anos, portanto eu não podia ter a ilusão de que ele não compreendia o nosso problema. Durante os seis meses em que ocupou uma mesa em nosso escritório, nunca o ouvi abrir a boca para falar de assuntos que não conhecia.
As formas tautoméricas contrastantes da guanina e da timina que podem ocorrer no DNA. Os átomos de hidrogênio que podem sofrer mudanças de posição (uma troca tautomérica) estão sombreados.
Bastante preocupado, voltei para minha mesa, esperando que surgisse algum esquema para resgatar a ideia do semelhante-com-semelhante. Mas era óbvio que as novas tarefas eram o golpe fatal. A mudança dos átomos de hidrogênio para seus locais cetônicos tornou ainda mais importante a diferença de tamanho entre as purinas e as pirimidinas do que seria se as formas enólicas existissem. Somente por um anseio muito especial eu poderia imaginar o esqueleto de polinucleotídeos se dobrando o bastante para acomodar sequências de bases irregulares. Até mesmo essa possibilidade desvaneceu quando Francis chegou. Ele percebeu de imediato que uma estrutura semelhante-com-semelhante só poderia causar uma repetição cristalográfica de 34 Å se cada cadeia tivesse uma rotação completa a cada 68 Å. Mas isso significaria que o ângulo de rotação entre bases sucessivas seria de apenas 18 graus, um padrão que Francis acreditava ter sido completamente descartado em suas manipulações recentes dos modelos. Francis também não gostava do fato de que a estrutura não apresentava explicação para as regras de Chargaff (adenina equivale a timina, guanina equivale a citosina). Eu, entretanto, mantive minha reação indiferente aos dados de Chargaff. Então, fiquei satisfeito com a chegada da hora do almoço, quando a falação alegre de Francis levou temporariamente meus pensamentos para o porquê de os alunos de graduação não conseguirem satisfazer as garotas au pair.
Não estava ansioso para voltar ao trabalho depois do almoço, porque temia que, ao tentar enquadrar as formas cetônicas em algum novo esquema, desse com um muro de pedra e tivesse de encarar o fato de que nenhum esquema regular de ligações de hidrogênio era compatível com as evidências de raios X. Enquanto eu ficasse lá fora contemplando os açafrões, era possível manter a esperança de que uma bela organização de bases me ocorreria. Por sorte, quando subimos as escadas, percebi que tinha uma desculpa para adiar o passo crucial da construção de modelos por pelo menos algumas horas. Os modelos metálicos de purinas e pirimidinas, necessários para checar sistematicamente todas as possibilidades de ligações de hidrogênio concebíveis, não haviam ficado prontos a tempo. Ao menos mais dois dias seriam necessários antes que estivessem em nossas mãos. Isso era muito tempo para ficar no limbo, até mesmo para mim, portanto passei o resto da tarde cortando representações precisas das bases em cartolina. Mas, quando ficaram prontas, percebi que a resposta teria de ser adiada até o dia seguinte. Depois do jantar, eu me juntaria a um grupo da hospedaria de Pop no teatro.
Quando cheguei ao escritório ainda vazio na manhã seguinte, tirei logo os papéis da minha mesa para ter uma superfície grande e plana onde formar pares de bases unidos por ligações de hidrogênio. Apesar de começar com certo preconceito contra a ideia do semelhante-com-semelhante, vi muito bem que isso não levava a lugar algum. Quando Jerry entrou, olhei em sua direção, vi que não era Francis e comecei a trocar as posições das bases para criar outras possibilidades de pareamento. De repente, percebi que um par de adenina-timina unido por duas ligações de hidrogênio tinha forma idêntica a um par de guanina-citosina unido por pelo menos duas ligações de hidrogênio. Todos os elos de hidrogênio pareciam se formar naturalmente; não era preciso trapacear para tornar os dois tipos de pares de bases idênticos na forma. Rapidamente, chamei Jerry para perguntar se agora ele tinha alguma objeção aos meus novos pares de bases.
Quando ele disse que não, meu moral explodiu, pois suspeitei que agora tínhamos a resposta para o enigma da equivalência exata entre o número de resíduos de purinas e o de pirimidinas. Duas sequências de bases irregulares poderiam ser comprimidas regularmente no centro de uma hélice se uma purina sempre se ligasse por meio de hidrogênio a uma pirimidina. Além do mais, a necessidade de haver uma ligação de hidrogênio significava que a adenina sempre faria par com a timina, enquanto a guanina só poderia parear com a citosina. As regras de Chargaff se destacaram repentinamente como consequência de uma estrutura duplo-helicoidal para o DNA. Ainda mais animador era que esse tipo de dupla hélice sugeria um esquema de replicação muito mais satisfatório do que o meu pareamento semelhante-com-semelhante considerado por breves instantes. Parear sempre adenina com timina e guanina com citosina significava que as sequências de bases das duas cadeias entrelaçadas eram complementares uma à outra. Dada a sequência de bases de uma cadeia, a do seu par era automaticamente determinada. Conceitualmente, era muito fácil visualizar como uma cadeia única poderia ser o molde para a síntese de uma cadeia com a sequência complementar.
Ao chegar, Francis mal havia atravessado a porta quando deixei escapar que a resposta para tudo estava em nossas mãos. Embora ele tivesse mantido o ceticismo durante alguns momentos por questão de princípio, os pares com formatos similares A-T e G-C tiveram o impacto esperado. A junção apressada das bases de diferentes maneiras não revelou outro modo que satisfizesse as regras de Chargaff. Minutos depois, ele percebeu que duas ligações glicosídicas (unindo base e açúcar) de cada par de bases estavam relacionadas de modo sistemático por um eixo diádico, perpendicular ao eixo helicoidal. Assim, os dois pares podiam ser intercambiados e as ligações glicosídicas continuariam viradas na mesma direção. A consequência importante disso era que uma determinada cadeia poderia conter tanto as purinas quanto as pirimidinas. Ao mesmo tempo, sugeria com força que os esqueletos das duas cadeias poderiam seguir em direções opostas.

Os pares de bases adenina-timina e guanina-citosina, usados para construir a dupla hélice (as ligações de hidrogênio estão pontilhadas). A formação de uma terceira ligação de hidrogênio entre guanina e citosina foi considerada, mas rejeitada porque um estudo cristalográfico da guanina indicou que seria muito fraca. Agora se sabe que essa hipótese estava errada. Três ligações fortes de hidrogênio podem ser traçadas entre a guanina e a citosina.
A questão passou a ser descobrir se os pares de bases A-T e G-C se encaixariam facilmente na configuração do esqueleto imaginada nas duas semanas anteriores. À primeira vista, parecia ser uma boa aposta, porque eu deixara livre uma grande área vazia para as bases no centro. Entretanto, nós dois sabíamos que não voltaríamos para casa até que um modelo completo com todos os contatos estereoquímicos satisfatórios fosse construído. Havia também o fato óbvio de que as implicações de sua existência eram muito importantes para arriscar um alarme falso. Por isso, senti uma leve náusea quando, na hora do almoço, Francis entrou voando no Eagle para contar a todos que pudessem ouvir que tínhamos descoberto o segredo da vida.