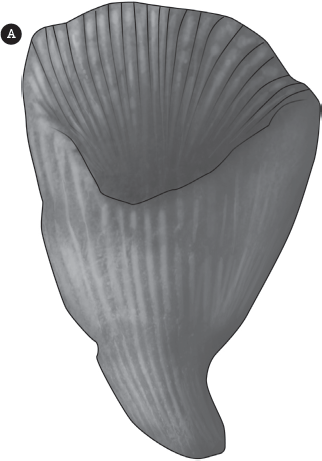
Sponges are about as simple as a multicellular animal can be. They are just a colony of independent cells that secrete a shared skeleton; otherwise they are unspecialized and can perform all of their functions individually. The next more advanced level of organization is to have cells specialized for certain functions, such as an inner or outer layer of tissue arranged around a central body cavity. In addition, organ systems like the nervous system can be well developed, so there are discrete nerve cells. This is the level of organization you find in the phylum Cnidaria (called “Coelenterata” in older books). The roots of both of these names describe the group well. Knidos is the Greek word for “stinger” or “nettles,” and all cnidarians have specialized stinger cells in their tentacles that are used to paralyze prey. The obsolete older term “Coelenterata” means “hollow gut” in Greek, which describes the fact that this phylum is the first and most primitive group of animals on Earth to have an internal body cavity (fig. 12.1A–D).
Figure 12.1 ▲
The phylum Cnidaria includes a number of distinct body plans. Some, such as the Scyphozoa, spend most of their lives in the medusa form; (A) they are known as sea jellies (formerly called “jellyfish,” although they are not related to fish). (B) The Hydrozoa are smaller colonial animals with many simple hydra polyps. (C) The Anthozoa include the corals, which have small polyps on a hard colony of calcite secreted by the individual animals, and (D) sea anemones. (Courtesy of Wikimedia Common)
You are probably familiar with several kinds of living cnidarians, especially sea jellies (formerly called “jellyfish,” but they are not true fish) and sea anemones. All cnidarians are built around the same body plan. They have a body arranged around an internal cavity, with an opening that serves as both mouth and anus at one end, and tentacles with stinger cells arranged around the mouth/anus. The internal wall of the body cavity is lined with endodermal cells, which are specialized for digesting prey that they swallow, and the external surface of the body is covered by ectodermal cells, which protect it from the outside world. They may also have masses of tissues between the endoderm and ectoderm called mesoderm or mesoglea, which provide bulk and thickness to their body walls. In a sea jelly, most of their mass is mesoglea, which makes them “jelly-like.” They also have a nervous system and muscular system that react to touch, which allows them to sting prey and pull it into their mouth using their tentacles. But they do not have eyes or most other sensory systems, nor do they have a respiratory system or excretory system. They take in all of their oxygen and get rid of all of their waste gases through the surface of their body.
From this basic body arrangement, cnidarians have two fundamental versions (fig. 12.2A): the jelly-like form, or medusa (named after the monster in the Greek myth of Perseus and Andromeda, who had snakes for hair and turned anyone to stone who saw her), and the attached anemone-like arrangement, or the polyp. In the medusa stage, like sea jellies, the tentacles hang down from the floating body, with the mouth below and the rest of the body above. In the polyp stage, the tentacles are on the top with the mouth, and the body is attached at the bottom.
Figure 12.2 ▲
The cnidarians spend their entire lives alternating between two basic arrangements of a hollow body cavity with a mouth opening surrounded by tentacles. (A) If they are attached to a surface with their tentacles catching food in the water currents, they are polyps. If they are free-floating with the mouth and tentacles hanging down, they are medusae. (B) The typical cnidarian life cycle. All cnidarians alternate generations between the polyp and medusa stage. (Illustration by Mary Persis Williams)
Although they look very different, both of these stages are actually part of the life cycle of the same organism (fig. 12.2B). Cnidarians reproduced by alternating generations between sexual and asexual reproduction. The polyps are the asexual stage, and they can bud and clone themselves asexually to form huge colonies. Eventually, however, some polyps produce tiny larvae that grow into free-swimming medusae, which have sexual organs. These swarm in the ocean and release sperm and eggs in a coordinated fashion, which fertilize to form a free-swimming larva that eventually settles and grows into a polyp again. Some cnidarians (like Hydra in a freshwater pond, or corals) spend almost all of their lives as immobile polyps, having only a brief medusa stage to spread to new habitats and colonize new seafloor. Others, like sea jellies, spend most of their lives in the medusa stage, and only exist as polyps for a brief time.
Most cnidarians, including freshwater hydras, sea anemones, and sea jellies, are soft-bodied and have no hard parts, so they are very rarely fossilized. However, one group of cnidarians—corals—leaves an excellent fossil record. Coral reefs are built of hundreds of polyps that looked like tiny sea anemones, but the base of their ectoderm secretes a large amount of calcite that combines with their neighboring polyps to form a huge coral “skeleton” or coral reef. When we look at a piece of modern coral or a coral fossil, it’s hard to imagine this rock was once a living organism, but at one time it was covered with a layer of hundreds of polyps, all busily trapping tiny prey with their tentacles while building their enormous skeletons (see fig. 12.1C).
Some corals can live in cold water, or in deep water without much light, but they tend to secrete very small skeletons. The vast majority of reef-building corals have symbiotic algae that live in their tissues. The algae are plants, which provide the coral with oxygen and use up the carbon dioxide they produce. Algae also helps the coral secrete the enormous volume of calcite needed to make their limestone reefs. All plants require light for photosynthesis, so large reef-building corals can only live in very shallow tropical oceans where there is good light penetration and where sand or mud from nearby land does not make the water dark and murky.
Thus coral reefs grow in only a few parts of the world today, mostly in tropical or subtropical shallow oceans far from the influence of nearby rivers and their muds. These areas include the Bahamas, the east coast of Florida (but not most of the Gulf of Mexico side, where the Mississippi mud makes the water dark), and other parts of the Caribbean and Yucatán Peninsula of Mexico; the Persian Gulf; and the South Pacific islands and Great Barrier Reef of Australia; and just a few other places.
Paleontologists believe that ancient corals had similar restrictions, yet we find fossil corals in places like Iowa. How could this be? At various times in geological history, especially during the early Paleozoic, the entire planet was much warmer, sea level was higher, and shallow warm tropical seas drowned the continents. As a result of these factors, fossil corals can be found in many places that are now far from the ocean. Let’s take a closer look at three main groups of corals that are commonly fossilized: tabulate corals, horn corals, and modern corals.
Tabulate corals (Order Tabulata). Tabulate corals are built of dense clusters of tiny tubes (corallites) all packed together like a box of drinking straws. Each corallite had lots of tiny dividing walls inside it, known as tabulae (“little table” in Latin), so this feature gives the group its name. A tiny coral polyp lived in the top of each tube.
Tabulate corals were extremely important as reef builders in the early Paleozoic, especially in the Silurian and Devonian. The “honeycomb coral” (Favosites) is a typical Silurian-Devonian coral found in many localities in the Midwest, and it gets its appearance from its “honeycomb” appearance when it is sliced across the top (fig. 12.3A). Another common Silurian reef builder is the “chain coral” (Halysites), whose corallites form little loops or chains with gaps between them in top view (fig. 12.3B). Tabulate corals were nearly wiped out during the great Late Devonian extinction event, but they straggled on through the rest of the Paleozoic, only to vanish in the great Permian extinction.
Figure 12.3 ▲
The tabulate corals were built of dense clusters of tube-shaped corallites. The top hole of the tube would have been occupied by a tiny polyp. (A) The tabulate coral Favosites, with the densely packed cluster of corallites giving it a porous appearance on the surface. (B) The chain coral Halysites, which had it tubular corallites arranged in chains. (Courtesy of Wikimedia Commons)
Horn corals (Order Rugosa). The other common group of Paleozoic corals was the horn corals, or rugosids. Many are shaped like the curved horns of a cow, hence their common name. In life, a typical horn coral like Streptelasma (fig. 12.4A) or Zaphrentis (fig. 12.4B) would have sat on the seafloor with the narrow pointed end embedded in the sediment, and a large anemone-like animal would have lived in the bowl-shaped cavity on the top of the cone, or calyx. Most rugosids had a wrinkled outer surface on the “horn,” hence their name (“rugose” means “wrinkled” in Latin). The most extreme example of this is the lumpy, irregularly wrinkled Devonian coral known as Heliophyllum (fig. 12.4C). Its name means “sun leaf,” possibly because, if you look down on the calyx, the septa radiate outward like the rays of the sun.
Figure 12.4 ▲
The rugosids, or “horn corals,” lived on the sea bottom with their pointed end embedded in the sediment and an anemone-like animal in the top bowl or calyx. (A) The common Ordovician horn coral Streptelasma. (B) The Silurian-Devonian coral Zaphrentis. (C) The wrinkled Devonian coral Heliophyllum. (D) The colonial rugosid Hexagonaria. Its corallites are tightly packed together like the cells in a honeycomb, giving it the nickname “honeycomb corals.” (E) Entire colonies of Hexagonaria are often eroded into pebbles, known as “Petoskey stones.” ([A] Illustration by Mary Persis Williams; [B–E] courtesy of Wikimedia Commons)
Horn corals like Heliophyllum have proven to be important for other reasons. If you slice one lengthwise and polish it, under the microscope you can see their daily growth bands as well as the annual growth bands caused by the seasons. In the 1960s, paleontologist John Wells counted the number of daily growth bands recorded in a year and found that there were 400 days in a year in the Devonian (about 400 million years ago), not the 365.25 days in a year we know today. In other words, the earth used to spin around its axis much faster, and it has been slowing down ever since. Other studies of different kinds of fossils with daily and annual growth bands confirmed this pattern. This phenomenon was predicted many years ago by astronomers who realized that the earth was being slowed down by the gravitational drag of the moon’s tidal pull on the earth. Many million years from now, the earth will finally slow down to a stop, no longer spinning on its axis. This has already happened to the moon, which has been slowed down by the earth’s tidal pull of gravity so that one side always faces the earth. The other side of the moon has only been seen by Apollo astronauts who flew around it and photographed it for the first time.
Although most horn corals were solitary, some rugosids clustered together into dense colonies. The most famous of these is Hexagonaria, an important reef-building coral of the Devonian (figs. 12.4D–E). When you slice across Hexagonaria, you can see that the corallites were packed together tightly to form a hexagonal honeycomb pattern, hence their name. Polished pebbles of Hexagonaria are common on the shores of the Great Lakes and have come to be known as “Petoskey stones.” These are the official State Rock of Michigan. Other important late Paleozoic colonial rugosids were Lithostrotion and Syringopora.
Like tabulates, rugose corals were hit hard by the Late Devonian mass extinction, which wiped out the immense reefs made of tabulate and corals and stromatoporoid sponges. The rugosids straggled on through the rest of the Paleozoic, but they were finally wiped out by the great Permian extinction.
Modern corals (Order Scleractinia, or hexacorals). Both tabulate and rugose corals vanished in the great Permian extinction, and there were no corals or reefs of any kind in the Early Triassic. By the Middle Triassic, some group of soft-bodied anemones evolved the ability to build big calcite skeletons again, and the modern group of scleractinian corals evolved (see fig. 12.1C). They are also called hexacorals because the internal dividing walls (septa) are arranged in multiples of six. (In contrast, the pattern for rugose corals is multiples of four, hence their old name, “Tetracorallia.”)
Scleractinian corals have adopted a wide variety of body shapes, from the branching corals to massive dense corals like the “brain coral,” and many other shapes. These corals have been evolving rapidly ever since the Triassic, but they have gone through numerous crises as well when other reef builders pushed them out of their preferred habitats. For example, during the Cretaceous coral reefs were almost displaced by huge reefs of colonial reef-building oysters known as rugosids.
Although it’s hard to imagine anything eating stony corals, they have numerous predators. Parrotfish have hard beaks that are excellent for crushing hard coral, and sudden population explosions of the crown-of-thorns sea star destroyed some tracts of the Great Barrier Reef. In addition, specialized crabs, polychaete worms, snails, and echinoids are adapted to feeding on coral polyps, and many organisms (clionid sponges, bivalves, and certain worms) are specialized to bore and burrow into coral skeletons.
Today, one of the great worries of biologists and environmental scientists is that global warming has been causing the world’s oceans to get warmer and more acidic, driving out their symbiotic algae and thus killing the world’s coral reefs in a process known as bleaching. In just the past 30 years, marine biologists have noted a dramatic worldwide dying off of coral reefs that were healthy and thriving in the 1980s and 1990s. Many reef biologists predict that coral reefs will be wiped out all over the planet within a few decades, causing a mass extinction in the huge number of animals that depend on coral reefs for their lives.