5
Creativity and Talent: Etiology of Familial Clustering
The question of whether an individual has inherited a particular skill or talent or whether skills and talent are acquired through practice and training has long been a topic of debate. On this question, Sir Francis Galton (1822–1911), a cousin of Darwin, stated: “I have no patience with the hypothesis occasionally expressed, and often implied, especially in tales written to teach children to be good, that babies are born pretty much alike, and that the sole agencies in creating differences between boy and boy, and man and man, are steady application and moral effort.” Galton undertook several efforts to address the issue in an empirical manner. In his books Hereditary Genius: An Inquiry into Its Laws and Consequences (1869) and English Men of Science: Their Nature and Nurture (1874), he investigated whether talent and giftedness possibly descend through ancestry. Through an inquiry into the relationships of eminent Englishmen, Galton observed high interrelatedness. Those included in his study more frequently had eminent fathers than grandfathers, and eminent sons than grandsons. Consequently, in the first- or second-degree relationship the degree of resemblance was higher than in third-degree relationships.
Familial clustering has also been described in various case studies of legendary families. For example, the Bach family showed a remarkable concentration of musical talent, and the Amati family introduced the violin into Italy. However, familial does not equal genetic (Martin, Boomsma, & Machin, 1997), and early exposure to favorable environments may explain the clustering that has been observed in pedigrees such as that of the Bach family. To investigate whether social advantages rather than biology underlie familial clustering, Galton introduced an early variant of the “adoption design,” in which he used the adopted sons of Popes and their nephews as a control group.
Assessing the influence of genetics and environment is essential for our understanding of the etiology of individual differences in behavior and complex traits, including talents. In addition to the adoption design, Galton pioneered several techniques that have become critical methods in the field of behavior genetics. These methods include the use of twins and families to estimate the impact of heredity on individual differences. In behavioral genetics, the central quest is to find out to what extent individual differences in a measured trait (phenotype) arise from differences in genotype and differences in environment (i.e., the sum of all nongenetic influences). The variance in a phenotype is decomposed into components due to the effects of polymorphic genes at all locations across the genome, and those due to the effects of environment. Studies where this decomposition of variance is performed often result in an estimated measure called “heritability.” Heritability is the proportion of phenotypic variance that can be accounted for by differences between individuals. Genetic influences are sometimes portioned into additive and nonadditive effects of alleles (an allele is a particular variant of a gene), while environmental effects are portioned into the effects of common environment shared by individuals growing up in the same family and the effects of nonshared environment (Plomin, DeFries, McClearn, & McGuffin, 2008).
The perception of talents and abilities as innate versus as products of environmental factors can have a large societal impact. A consequence of the belief that innate talent is a precondition for high achievement may be that young people who are not identified as having the talent could be more likely to be denied the help and encouragement they would need to attain high levels of competence (Howe, Davidson, & Sloboda, 1998). Moreover, insight into the etiology of traits, whether genetic, environmental, or both, potentially influences the definition and operationalization of that trait. In Galton’s work, talent was operationalized in a rather unique manner. It was defined according to a person’s enduring reputation, called eminence. By this, Galton (1869, p. 33) referred to “the opinion of contemporaries, revised by posterity . . . the reputation of a leader of opinion, of an originator, of a man to whom the world deliberately acknowledges itself largely indebted.” In other works, talent has often been used interchangeably with the term genius. Successful composers, artists, poets, writers, and scientists began to be called geniuses and achieved fame for the realization of their special abilities. However, large variety exists within the term genius. Those who are highly intelligent (e.g., Einstein), highly eminent (e.g., Nobel Prize winners), or highly creative (e.g., Napoleon Bonaparte) are all referred to as geniuses (Simonton, 1999). In their review, Howe et al. (1998) assign five properties to talent: (1) it originates in genetically transmitted structures and hence is at least partly innate; (2) its full effects may not be evident at an early stage, but there will be some advance indications, allowing trained people to identify the presence of talent before exceptional levels of mature performance have been demonstrated; (3) these early indications of talent provide a basis for predicting who is likely to excel; (4) only a minority are talented, for if all children were, then there would be no way to predict or explain differential success; and finally, (5) talents are relatively domain specific (Howe et al., 1998, p. 399).
In this chapter, we aim to extend our prior work on talents (Vinkhuyzen, van der Sluis, Posthuma, and Boomsma, 2009) in which we studied the contribution of genetic factors to a series of talents in a general population sample of adolescent and young adult Dutch twins. The talents included music, arts, chess, writing, memory, knowledge, languages, and mathematics. Participants were asked to rate themselves on each of these domains. Other studies have also focused on the genetic architecture of some of these talents (Howe et al., 1998; Lubinsky et al., 2006; McGue et al., 1993; Ruthsatz et al., 2008). We briefly review the main findings for each of these talents below.
Individual differences in musical abilities have been attributed to genetic factors by Fuller and Thompson (1978). Heritability estimates range from 10 percent for nonschool musical performance to 71 percent for vocal performance (Coon & Carey, 1989). Pitch recognition is heritable for 70 to 80 percent (Drayna, Manichaikul, de Lange, Snieder, & Spector, 2001). Absolute pitch has also been found to aggregate in families (Baharloo et al., 2000).
Regarding chess, the nature-nurture debate was intensified by the Hungarian psychologist and chess teacher László Polgár. In his book Bring Up Genius! he emphasized the importance of specialized training (Polgár, 1989). He argued that by fully tapping into the potential of children, one could become capable of simply reading the chessboard and making decisions based on intuition. This belief was confirmed by a study of De Bruin and colleagues (2008) where most of the variation in performance was accounted for by deliberate practice. However, the question whether extensive practice reflects a genetic disposition to enjoy and benefit from playing and practicing chess remains unanswered.
Concerning the ability to learn and acquire a new language as well as impairments in language development, several studies have addressed the role of genetic factors and high heritability estimates (Dale et al., 1998; Newbury et al., 2005; Spinath et al., 2004; Stromswold, 2001; Viding et al., 2003). Recently, a twin study in young adolescents examined the heritability of second-language acquisition (Dale et al., 2010) and suggested a substantial heritability of 67 percent. The remaining variance was explained by shared environmental influences (13 percent) and unique environmental influences (20 percent). The authors mentioned that the influence of genetics in the ability to speak multiple foreign languages is higher than estimates in first-language acquisition, and that large overlap exists between the genes for first- and second-language acquisition (Dale et al., 2010).
Studies into the decomposition of the variance in what is called “general knowledge” derives, for example, from the Information subtest of the WAIS-III (1997). Heritability for the information subtest is estimated at 75 percent (Rijsdijk et al., 2002). Regarding memory function, consensus exists that genetic factors explain around 50 percent of individual differences (Bouchard, 1998; Finkel et al., 1995).
Mathematics has been studied from both biological and genetic perspectives. The biological basis for mathematical talent was explored, for example, in a study on sex differences in mathematics (Benbow & Lubinski, 1993). Hormonal influences, medical and bodily conditions, and specific brain activations are associated with mathematical achievement. Twin studies have reported heritability estimates ranging from 19 up to 90 percent (Alarcon et al., 2000; Thompson et al., 1991; Wijsman et al., 2004).
Overall, the impression from earlier studies is that both genetics and the environment contribute to variation in talents. Extending the classical twin design to a multigeneration design has several advantages, including a direct assessment of and test for the genetic and cultural transmission from one generation to the next. In addition, data from spouses (the parents of twins) provide information on nonrandom mating.
In this chapter, we include information on self-reported talents from twins and from their parents. The inclusion of parents in a parent-offspring model offers the possibility to test for several hypotheses regarding the genetic architecture of complex traits (e.g., Eaves, 1972; Martin, Boomsma, & Machin, 1997; Maes et al., 2009; Fulker, 1989). One of those hypotheses considers the relationship between parental phenotypes and those of their children and addresses the question of cultural versus genetic inheritance. In addition, having data on spouses offers the possibility to study assortative or nonrandom mating. This type of mating refers to the phenomenon where the phenotypes of the parents are correlated (Eaves, Last, Martin, & Jinks, 1977), for example, men with a particular talent may more often tend to marry women who also are talented. The consequences of assortative mating depend on the mechanism that leads to assortment, including social homogamy, social interaction, and phenotypic assortment (Heath & Eaves, 1985; Reynolds et al., 2006; van Leeuwen et al., 2008). Social homogamy refers to the phenomenon where individuals coming from similar social backgrounds are more likely to meet and marry each other. Social interaction occurs when spouses mutually influence each other because they spend time together. Phenotypic assortment refers to the phenomenon where individuals select each other based on their phenotype. If assortment is phenotypic, this can have important consequences for the genetic architecture of traits because it increases the additive genetic variance in the offspring generation and the covariance between additive genetic factors in first-degree relatives (Eaves et al., 1989; van Grootheest et al., 2008).
The combination of parental data and data from monozygotic (MZ) and dizygotic (DZ) twins makes for a strong design to establish and screen for genetic influences in behavioral dimensions, such as talents. The different degree of genetic relatedness of MZ and DZ twins is used to identify the relative contribution of genes and environment. MZ twins derive from one fertilized egg (one zygote) and are genetically (nearly) identical, whereas DZ twins derive from two different fertilized eggs (two zygotes) and share on average half of their segregating genes, just like nontwin brothers and sisters. If the resemblance for a particular trait is larger in MZ than in DZ twins, this strongly suggests that their larger genetic resemblance is important. From the observed phenotypic differences between twins, the heritability can be estimated, which, if greater than zero, implies that genetic factors influence a trait (Boomsma et al., 2002).
When the shared environment is likely to have some influence on a trait, the parents-twins model provides the opportunity to estimate the influence of the phenotypes of the mother and father on the shared environment of the offspring. This implies that it is possible to partition the shared environmental effects found in talent among offspring into the effects of the parental phenotype on offspring behavior (vertical cultural transmission), environmental effects that are shared among offspring but not shared with parents (horizontal cultural transmission), and the effects of assortative mating. These effects can be separated by simultaneously modeling the correlations between parents, between parents and their offspring, and between offspring. When the spouse correlation is significant, part of the shared environmental effects may be explained by assortative mating. If the parent-offspring correlations are larger than what would be expected under genetic transmission alone, this could imply that the shared environmental effects found in offspring are the results of vertical cultural transmission. A lower parent-offspring correlation compared with DZ twin and sibling correlations can indicate that part of the environmental effects is shared between twins and siblings only, that part of the genetic factors acts in a dominant manner, or that different genetic factors affect talent in the two generations.
Netherlands Twin Registry (NTR) Study
The data for this study were collected in 1991 through a survey sent out by the Netherlands Twin Registry (NTR) established in 1987 (Boomsma et al., 2006). Adolescent and young adult twins and their parents were recruited through city council registrations and contacted by mail and invited to complete a survey (for details, see Koopmans, van Doornen, & Boomsma, 1994; Willemsen, Posthuma, & Boomsma, 2005). Data from 3,331 twins and 2,995 parents from 1,693 families were available. The twin sample consisted of 557 monozygotic males (MZM), 473 dizygotic males (DZM), 759 monozygotic females (MZF), 590 dizygotic females (DZF), 952 dizygotic opposite sex (DOS), and 1,407 fathers and 1,588 mothers. The mean age of the twins was 17.73 years (SD = 2.37, range = 12–50 years). The mean age for the parents was 46.6 years (SD = 5.4, range = 33–71 years).
The talents were selected from the Talent Inventory developed by McGue et al. (1993), and included self-report data on music, arts, chess, writing, memory, knowledge, language, and mathematics. Participants were asked to rank their own competence, in comparison to the general population, on an ordinal four-point scale representing different categories. The first category represents people who classify themselves as less competent than most people. The second category represents the average (as competent as most people), the third category the above average (more competent than most people), and the fourth category represents people who classify themselves at the top end, that is, as being exceptionally skilled.
Talent for Music refers to singing or playing one or more instruments. A talent for Arts reflects artistic and creative activities (e.g., painting, acting). Chess refers to the ability to play games like chess, backgammon, and mah-jong. Talent for Writing is defined as creative writing (e.g., letters, manuscripts, books). Memory reflects general mnemonic skills (e.g., events, numbers, and facts). Knowledge refers to general and specific knowledge of facts. Language is defined as the ability to speak one or more foreign languages. Finally, talent for Mathematics refers to mathematical and numerical ability.
The analysis of the categorical data was based on a “liability” model (Falconer & Mackay, 1996; Neale & Cardon, 1992). In this model, a categorical variable is assumed to reflect an imprecise measurement of an underlying normal distribution of liability. A threshold divides the population into two or more groups (categories) of people. For the study of talent—which was measured on a four-point scale—categories 3 and 4 were merged, and scores on the liability distribution could fall into three categories that were defined by two thresholds. Since the liability is a theoretical construct, its scale is arbitrary. The liability was assumed to be standard normally distributed with zero mean and unit variance. Correlations among family members on the liability scale and thresholds were estimated. Thresholds were allowed to vary as a function of sex (0 for male and 1 for female) and age (standardized). Age coefficients were similar for parents and for threshold 1 or 2. All analyses were conducted using structural equation modeling in Mx (Neale, Boker, Xie, & Maes, 2003).
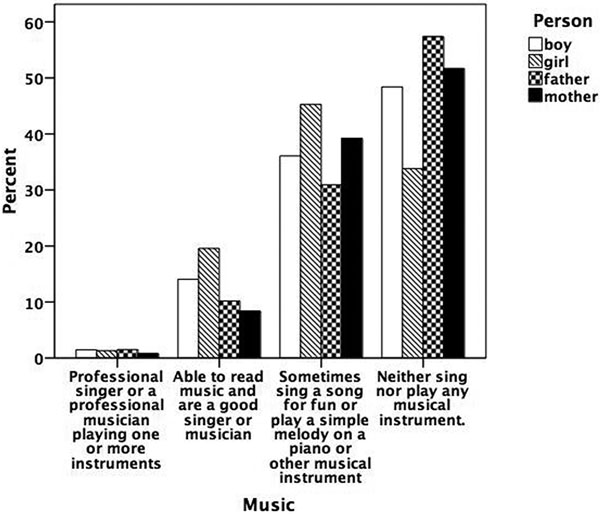
Figures 5.1 to 5.8 show the prevalence of each talent in the adolescent boys and girls and in their fathers and mothers. In general, the proportion of individuals with exceptional talent was small. The percentage of individuals falling into the exceptionally talented category is typically less than 10 percent, except for talent in Languages. The proportion of individuals with above average but not exceptional aptitude is somewhat larger (ranging between 3 and 40 percent). When the two above average categories are combined, the percentage of individuals with above average talent ranges from 4.6 percent for Mathematics in mothers, to 42.5 percent for Memory in the offspring generation. From this figure it can also be seen that there are some clear generation and sex differences in prevalence of talent. Adolescent and young adult twins rate themselves more often as above average talented for Music, Arts, Memory, and Languages than their parents. We also see a clear tendency for men to report themselves more often as above average talented than women for Memory, Knowledge, Chess, and Mathematics. Adolescent girls rate themselves as most talented for Music, Arts, Writing, and Languages, compared to adolescent boys and their parents.
Figure 5.1
Prevalence of talent for music.
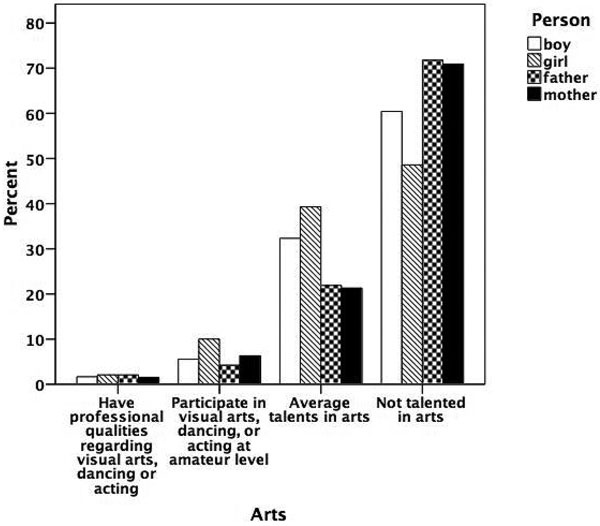
Figure 5.2
Prevalence of talent for arts.
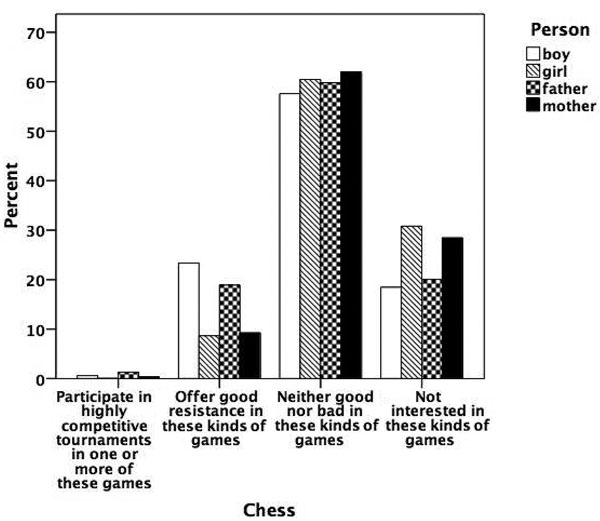
Figure 5.3
Prevalence of talent for chess.
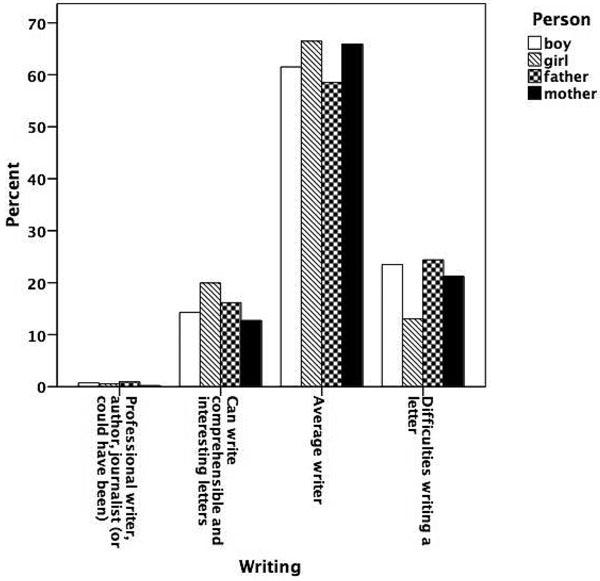
Figure 5.4
Prevalence of talent for writing.
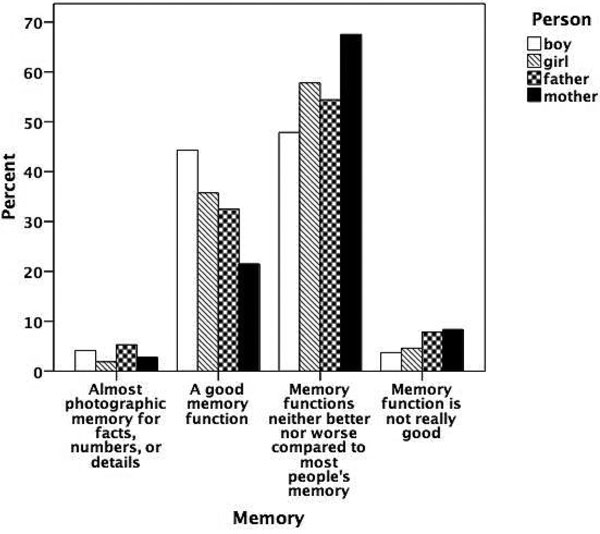
Figure 5.5
Prevalence of talent for memory.
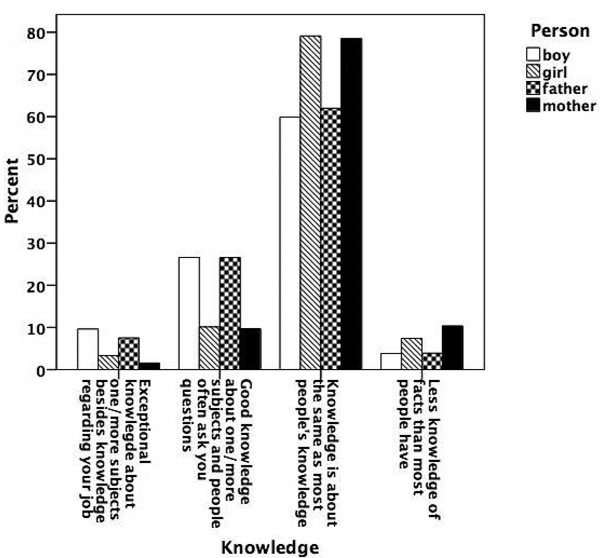
Figure 5.6
Prevalence of talent for knowledge.
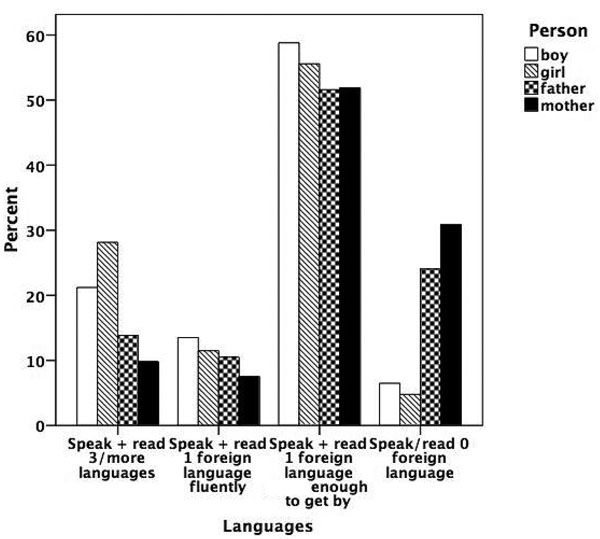
Figure 5.7
Prevalence of talent for languages.
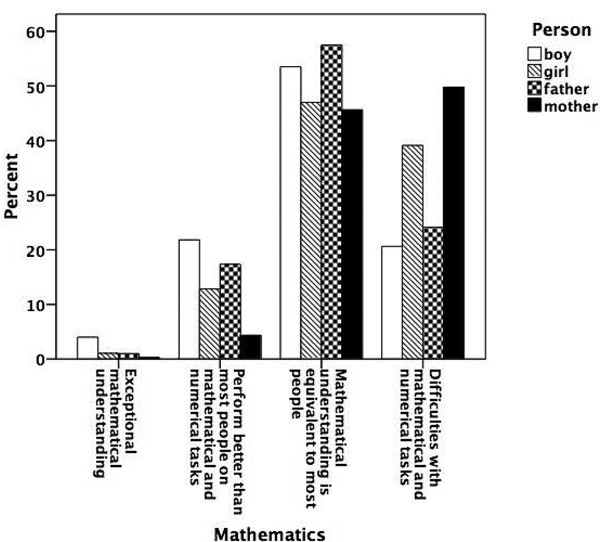
Figure 5.8
Prevalence of talent for mathematics.
The polychoric correlations among family members are displayed in table 5.1. For all variables, MZ twin correlations exceeded the DZ twin correlations, suggesting genetic influences. MZ twin correlations ranged between 0.49 and 0.78. DZ twin correlations ranged from 0.09 up to 0.48. Only for Music was the DZ correlation larger than half the MZ correlation, which could be an indication of shared environmental influences. For some variables, including Chess, Writing, Memory, and Mathematics, the DZ correlation is smaller than half the MZ correlation, suggesting the influence of nonadditive genetic factors. When inspecting the parent-offspring correlations, it becomes clear that they are all significantly larger than zero and often very similar to the DZ twin correlations, suggesting that differences in genetic effects across generations are unlikely. Importantly, the fact that both DZ correlations and parent-offspring correlations are not larger than half the MZ correlations strongly suggests that cultural transmission is an unlikely explanation for the familial clustering of talent behavior. Spouse correlations are mostly significantly different from zero, except for Memory. This indicates that for all aptitudes except Memory there is significant nonrandom mating. Nonrandom mating is largest for Languages (0.48).
Table 5.1
Twin, parent-offspring, and spouse correlations for various talents.
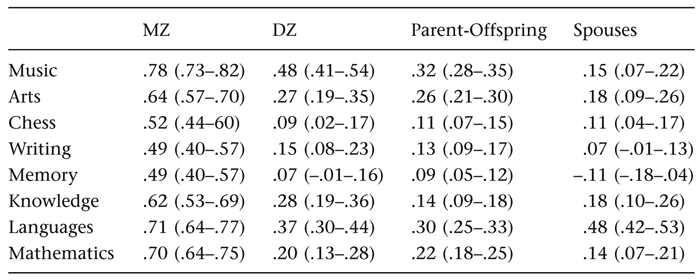
Notes: Correlation estimates with 95 percent confidence intervals.
Conclusions
To summarize, the familial clustering for a variety of talents, already reported as early as in the end of the nineteenth century by pioneer Sir Francis Galton, is confirmed in our study. The unique features of the parents-twins design made it possible to demonstrate that this familial clustering is unlikely to be explained by cultural transmission from one generation to the next, but is rather due to genetic transmission. This holds for all the talents that were included in this study: Music, Arts, Chess, Writing, Memory, Knowledge, Languages, and Mathematics. In addition, nonrandom mating was observed for all talents except Memory. Some evidence for nonadditive genetic influences was found for Chess, Writing, Memory, and Mathematics, as indicated by the low DZ correlations, compared to the MZ correlations. Parent-offspring correlations for these talents were similarly low. This suggests that the familial correlations that were observed are best explained by genetic inheritance, that genetic effects include nonadditivity, and that environmental influences do not contribute to familial resemblance over and above the resemblance that is accounted for by genetic effects.
Our findings indicate that children who grow up with talented parents also are more often exposed to favorable environmental factors. There is no evidence for a “main effect” of cultural transmission, but this double advantage will often offer children with innate talent the opportunity to practice the skills that are required for the expression of talent. Moreover, those with talent will benefit from practice (for example, for music and chess) and to experience the rewards that practice can bring them. Unlike children without any potential for a particular talent, the reward they experience may act as a stimulus to continue practicing, whereby individual differences in practicing themselves become influenced by genotype.
Our findings also have implications for gene-finding studies. Since talented behavior is so clearly heritable, what are the genes that constitute this heritability? Recently, genome-wide association (GWA) studies have turned out to be quite successful for some complex polygenic physical and medical traits, such as height, body mass index, and type II diabetes (Lango Allen et al., 2010; Saxena et al., 2007; Speliotes et al., 2010; Yang et al., 2010). GWA studies have more difficulties in pinpointing the genetic variants that influence more complex cognitive, affective, and behavioral phenotypes, although some progress has been made (e.g., De Moor et al., 2010; Sullivan et al., 2009; Vink et al., 2009). For example, GWA studies for intelligence have come up with a limited number of genetic variants of small effect that may influence intelligence, but a recent study shows that the genome-wide polymorphisms are able to explain a large portion of the heritability of intelligence (Davies et al., 2011). To our knowledge, no such studies have been conducted for the talents discussed in this chapter. Yet, the findings concerning general intelligence could provide important information on the underlying molecular genetic basis of some of the talents presented here, such as Memory, Knowledge, Languages, and Mathematics.
References
Alarcon, M., Knopik, V. S., & DeFries, J. C. (2000). Covariation of mathematical achievement and general cognitive ability in twins. Journal of School Psychology, 38, 63–77.
Baharloo, S., Service, S. K., Risch, N., Gitschier, J., & Freimer, N. B. (2000). Familial aggregation of absolute pitch. American Journal of Human Genetics, 67, 755–758.
Benbow, C. P., & Lubinski, D. (1993). Psychological profiles of the mathematically talented: Some gender differences and evidence supporting their biological basis. In K. Ackerill (Ed.), The origins and development of high ability (pp. 44–66). New York: John Wiley.
Boomsma, D. I., Busjahn, A., & Peltronen, I. (2002). Classical twin studies and beyond. Nature Reviews: Genetics, 3, 872–882.
Boomsma, D. I., de Geus, E. J. C., Vink, J. M., Stubbe, J. M., Distel, M. A., Hottenga, J. J., et al. (2006). Netherlands Twin Register: From twins to twin families. Twin Research and Human Genetics, 9, 849–857.
Bouchard, T. I., Jr. (1998). Genetic and environmental influences on adult intelligence and special mental abilities. Human Biology, 70, 257–279.
Coon, H., & Carey, G. (1989). Genetic and environmental determinants of musical ability in twins. Behavior Genetics, 19, 183–193.
Dale, P. S., Harlaar, N., Haworth, C. M. A., & Plomin, R. (2010). Two by two: A twin study of second-language acquisition. Psychological Science, 21, 635–640.
Dale, P. S., Simonoff, E., Bishop, D. V. M., Eley, T. C., Oliver, B., Price, T. S., et al. (1998). Genetic influence on language delay in 2-year old children. Nature Neuroscience, 1, 324–328.
Davies, G., Tenesa, A., Payton, A., Yang, J., Harris, S. E., Liewald, D., et al. (2011). Genome-wide association studies establish that human intelligence is highly heritable and polygenic. Molecular Psychiatry, 16, 996–1005.
De Bruin, A. B., Smits, N., Rikers, R. M., & Schmidt, H. G. (2008). Deliberate practice predicts performance over time in adolescent chess players and drop-puts: A linear mixed models analysis. British Journal of Psychology, 99, 473–497.
De Moor, M. H. M., Costa, P. T., Terracciano, A., Krueger, R. F., de Geus, E. J. C., Tanaka, T., et al. (2010). Meta-analysis of genome-wide association studies for personality. Molecular Psychiatry; Epub ahead of print. doi:10.1038/mp.2010.
Drayna, D., Manichaikul, A., de Lange, M., Snieder, H., & Spector, T. (2001). Genetic correlates of musical pitch recognition in humans. Science, 291, 1969–1972.
Eaves, L. J. (1972). Computer simulation of sample size and experimental design in human psychogenetics. Psychological Bulletin, 77, 144–152.
Eaves, L. J., Last, K., Martin, N. G., & Jinks, J. L. (1977). A progressive approach to non additivity and genotype-environmental covariance in the analysis of human differences. British Journal of Mathematical and Statistical Psychology, 30, 1–42.
Eaves, L. J., Fulker, D. W., & Heath, A. C. (1989). The effects of social homogamy and cultural inheritance on the covariances of twins and their parents: A LISREL model. Behavior Genetics, 19, 113–122.
Falconer, D. S., & Mackay, T. F. C. (1996). An introduction to quantitative genetics. Essex: Longmans, Green, Harlow.
Finkel, D., Pedersen, N., & McGue, M. (1995). Genetic influences on memory performance in adulthood: Comparison of Minnesota and Swedish twin data. Psychology and Aging, 10, 437–446.
Fulker, D. W. (1989). Genetic and cultural transmission in human behavior. In B. S. Weir, E. J. Eissen, M. M. Goodman, & G. Namkoong (Eds.), Proceedings of the Second International Conference on Quantitative Genetics (pp. 318–340). Sunderland, MA: Sinauer.
Fuller, J. L., & Thompson, W. R. (1978). Foundations of behavior genetics. Saint Louis: Mosby.
Galton, F. (1874). English men of science: Their nature and nurture. London: Macmillan.
Galton, F. (1869). Hereditary genius. London: Macmillan.
Heath, A. C., & Eaves, L. J. (1985). Resolving the effects of phenotype and social back-ground on mate selection. Behavior Genetics, 15, 15–30.
Howe, M. J. A., Davidson, J. W., & Sloboda, J. A. (1998). Innate talents, reality or myth. Behavioral and Brain Sciences, 21, 399–407.
Koopmans, J. R., van Doornen, L. J. P., & Boomsma, D. I. (1994). Smoking and sports participation. In U. Goldbourt, U. DeFaire, & K. Berg (Eds.), Genetic factors in coronary heart disease (pp. 217–235). Dordrecht: Kluwer Academic.
Lango Allen, H., Estrada, K., Lettre, G., Berndt, S. I., Weedon, M. N., Rivadeneira, R., et al. (2010). Hundreds of variants clustered in genomic loci and biological pathways affect human height. Nature, 467, 832–838.
Lubinsky, D., Benbow, C. P., Webb, R. M., & Bleske-Rechek, A. (2006). Tracking exceptional human capital over two decades. Psychological Science, 17, 194–199.
Maes, H. H., Neale, M. C., Medland, S. E., Keller, M. C., Martin, N. G., Heath, A. C., et al. (2009). Flexible Mx specification of various extend twin kinship designs. Twin Research and Human Genetics, 12, 26–34.
Martin, N. G., Boomsma, D. I., & Machin, G. A. (1997). A twin-pronged attack on complex traits. Nature Genetics, 17, 387–392.
McGue, M., Hirsch, B., & Lykken, D. T. (1993). Age and the self-perception of ability: A twin study analysis. Psychology and Aging, 8, 72–80.
Neale, M. C., Boker, S. H., Xie, G., & Maes, H.H. (2003). Mx: Statistical modeling (6th Ed.). Richmond: Virginia Commonwealth University.
Neale, M. C., & Cardon, L. R. (1992). Methodology for genetic studies of twins and families. Dordrecht: Kluwer Academic.
Newbury, D. F., Bishop, D. V. M., & Monaco, A. P. (2005). Genetic influences on language impairment and phonological short-term memory. Trends in Cognitive Sciences, 9, 528–534.
Plomin, R., DeFries, J. C., McClearn, G. E., & McGuffin, P. (2008). Behavioral genetics (5th Ed.). New York: Worth Publishers.
Polgár, L. (1989). Nevelj zsenit! (Bring up genius!). Budapest: Kossuth Kiadó ZRT.
Reynolds, C. A., Barlow, T., & Pedersen, N. L. (2006). Alcohol, tobacco, and caffeine use: Spouse similarity processes. Behavior Genetics, 36, 201–215.
Rijsdijk, F. V., Vernon, P. A., & Boomsma, D. I. (2002). Application of hierarchical genetic models to Raven and WAIS subtests: A Dutch twin study. Behavior Genetics, 32, 199–210.
Ruthsatz, J., Detterman, D., Griscom, W., & Collins, J. M. (2008). Becoming an expert in the musical domain: It takes more than just practice. Intelligence, 36, 330–338.
Saxena, R., Voight, B. F., Lyssenko, V., Burtt, N. P., de Bakker, P. I. W., Chen, H., et al. (2007). Genome-wide association analysis identifies loci for type 2 diabetes and triglyceride levels. Science, 316, 1331–1336.
Simonton, D. K. (1999). Origins of genius: Darwinian perspectives on creativity. Oxford: Oxford University Press.
Speliotes, E. K., Willer, C. J., Berndt, S. I., Monda, K. L., Thorleifsson, G., Jackson, A. U., et al. (2010). Association analyses of 249,796 individuals reveal 18 new loci associated with body mass index. Nature Genetics, 42, 937–948.
Spinath, F. M., Price, T. S., Dale, P. S., & Plomin, R. (2004). The genetic and environmental origins of language disability and ability: A study of language at 2, 3, and 4 years of age in a large community sample of twins. Child Development, 75, 445–454.
Stromswold, K. (2001). The heritability of language: A review and meta-analysis of twin, adoption, and linkage studies. Language, 77, 647–723.
Sullivan, P. F., de Geus, E. J. C., Willemsen, G., James, M. R., Smit, J. H., Zandbelt, T., et al. (2009). Genomewide association for major depressive disorder: A possible role for the presynaptic protein piccolo. Molecular Psychiatry, 14, 359–375.
Thompson, L. A., Detterman, D. K., & Plomin, R. (1991). Associations between cognitive-abilities and scholastic achievement-genetic overlap but environmental differences. Psychological Science, 2, 158–165.
van Grootheest, D. S., van den Berg, S. M., Cath, D. C., Willemsen, G., & Boomsma, D. I. (2008). Marital resemblance for obsessive-compulsive, anxious, and depressive symptoms in a population-based sample. Psychological Medicine, 38, 1731–1740.
van Leeuwen, M., van den Berg, S. M., & Boomsma, D. I. (2008). A twin-family study of general IQ. Learning and Individual Differences, 18, 76–88.
Viding, E., Price, T. S., Spinath, F. M., Bishop, D. V. M., & Dale, P. S. (2003). Genetic and environmental mediation of the relationship between language and nonverbal impairment in 4-year-old twins. Journal of Speech, Language, and Hearing Research: JSLHR, 46, 1271–1282.
Vink, J. M., Smit, A. B., de Geus, E. J., Sullivan, P., Willemsen, G., Hottenga, J. J., et al. (2009). Genome-wide association study of smoking initiation and current smoking. American Journal of Human Genetics, 84, 367–379.
Vinkhuyzen, A. A. E., van der Sluis, S., Posthuma, D., & Boomsma, D. I. (2009). The heritability of aptitude and exceptional talent across different domains in adolescents and young adults. Behavior Genetics, 9, 380–392.
WAIS-III. (1997). Manual. Dutch version. Lisse: Swets and Zeitlinger.
Wijsman, E. M., Robinson, N. M., Ainsworth, K. H., Rosenthal, E. A., Holzman, T., & Raskind, W. H. (2004). Familial aggregation patterns in mathematical ability. Behavior Genetics, 24, 51–62.
Willemsen, G., Posthuma, D., & Boomsma, D. I. (2005). Environmental factors determine where the Dutch live: Results from the Netherlands Twin Register. Twin Research and Human Genetics, 8, 312–317.
Yang, J., Benyamin, B., McEvoy, B. P., Gordon, S., Henders, A. K., Nyholt, D. R., et al. (2010). Common SNPs explain a large proportion of the heritability for human height. Nature Genetics, 42, 565–569.